Descargar la presentación
La descarga está en progreso. Por favor, espere

1
UNIDAD TEMÁTICA: INTRODUCCIÓN A LAS PATOLOGÍAS DEL SISTEMA INMUNE
UNIVERSIDAD NACIONAL DE ASUNCIÓN FACULTAD DE CIENCIAS QUÍMICAS CÁTEDRA DE INMUNOLOGÍA CLÍNICA UNIDAD TEMÁTICA: INTRODUCCIÓN A LAS PATOLOGÍAS DEL SISTEMA INMUNE

2
OBJETIVOS Analizar los diferentes mecanismos que actúan en la regulación del sistema inmune Discernir diferencias entre los mecanismos de tolerancia central y periférica

3
CONTENIDOS 1.2 Regulación de la respuesta inmune
Elementos que participan en la regulación del sistema inmune: antígenos, anticuerpos, factores genéticos, sistema endócrino y sistema nervioso 1.2.2 Rol de la subpoblación de células T en la regulación: Th17 y T reguladoras. Tolerancia a lo propio 1.3.1 Mecanismos de tolerancia del sistema inmune 1.3.2 Tolerancia central 1.3.3 Tolerancia Periférica 1.3.4 Tolerancia artificial

4
REGULACIÓN DEL SI 1. El antígeno 2. Los anticuerpos 3. Determinadas células del sistema inmune 4. Las citoquinas 5. Los factores genéticos 6. El sistema regulador neuroendocrino 7. Linfocitos T reguladores 8. Homoestasis de la respuesta inmune antimicrobiana 9. Tolerancia oral

5
ANTÍGENO NATURALEZA DOSIS VIA DE ENTRADA
Microorganismos intracelulares y sustancias como la sílice estimulan una respuesta celular. Microorganismos extracelulares y antígenos solubles una respuesta humoral. Los antígenos capsulares bacterianos de polisacárido. IgM DOSIS Altas dosis de antígeno pueden inducir tolerancia de células TyB VIA DE ENTRADA Los antígenos administrados por vía subcutánea o intradérmica evocan una respuesta inmunitaria activa. Los administrados por vía intravenosa, oral o en aerosol pueden causar tolerancia o desviación inmunitaria.

6
Antígeno intracelular/antígeno extracelular Th1/Th2.

7
Anticuerpos Administración pasiva de IgM junto con el antígeno mejora la respuesta inmune (anticuerpos mono y policlonales). Inmunocomplejos IgM son captados por receptores Fc O C3 de las APC y procesados con mayor eficacia. Los inmunocomplejos que tienen IgM estimulan la respuesta antiidiotípica frente a la IgM que amplifica la respuesta inmune. Mientras que anticuerpos IgG suprimen la respuesta.
. Inmunocomplejos IgM son captados por receptores Fc O C3 de las APC y procesados con mayor eficacia. Los inmunocomplejos que tienen IgM estimulan la respuesta antiidiotípica frente a la IgM que amplifica la respuesta inmune. Mientras que anticuerpos IgG suprimen la respuesta.")
8
Regulación por anticuerpos:
Retrorregulación negativa basada en la supresión mediada por los anticuerpos (fundamentalmente IgG), que bloquean las interacciones entre el epitopo y el BCR especifico de los linfocitos B, por entrecruzamiento de receptores Fc unidos a inmunoglobulinas o inmunocomplejos en diferentes células: Inhibición de las células B por entrecruzamiento de receptores
, que bloquean las interacciones entre el epitopo y el BCR especifico de los linfocitos B, por entrecruzamiento de receptores Fc unidos a inmunoglobulinas o inmunocomplejos en diferentes células: Inhibición de las células B por entrecruzamiento de receptores.")
9
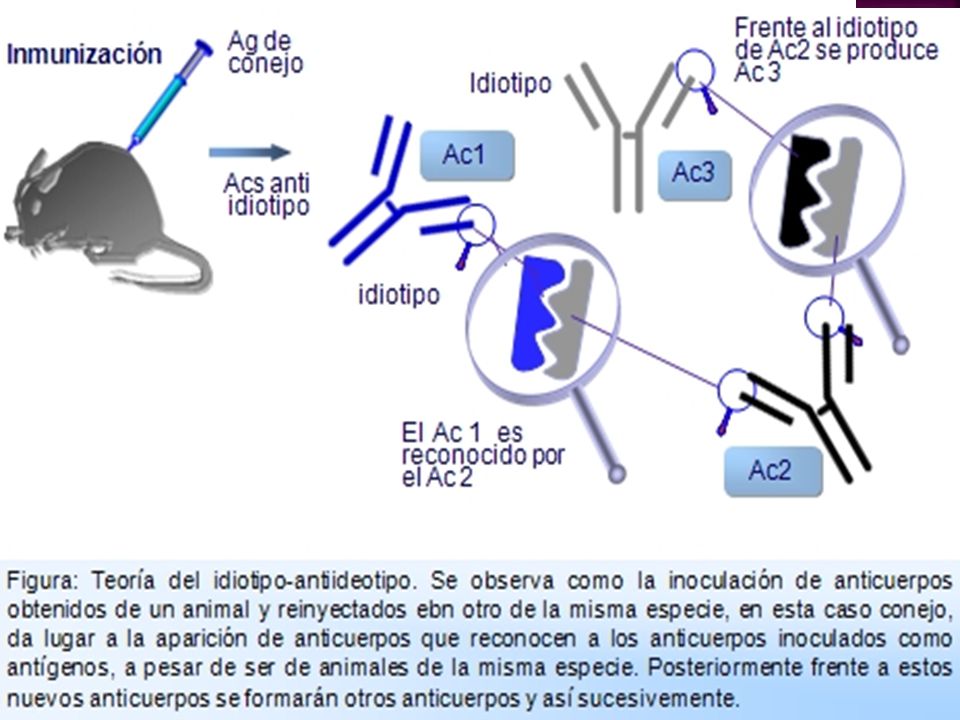
Redes idiotípicas: Durante el periodo neonatal las regiones idiotípicas de
las inmunoglobulinas son un repertorio de inmunógenos para el sistema inmune. Los anticuerpos que se formarían frente a estos idiotipos se denominan anticuerpos anti-idiotipo, y podrían modificar el curso de las reacciones inmunitarias. Jerne propuso la existencia de una red o entramado idiotipo-antiidiotipo que, en estado de reposo del S.I. tiene un equilibrio cuantitativo y cualitativo

10
Estos autoantígenos están en muy pequeña cantidad para ser destruidos por el sistema inmune, pero cuando un determinado clon es estimulado por su antígeno, el sistema inmune ya no podría ignorarlo y originaría unas respuestas contra ellos en forma de segundos anticuerpos (antiidiotipo), que encajarían con los primeros y tendrían además un efecto modulador que puede ser positivo o negativo. A este proceso se le denominó "teoría de la red de anticuerpos" (Figura: Teoría del idiotipo-antiidiotipo).
..")
12
RESPUESTAS ANTI-IDIOTÍPICAS
Las respuestas anti-idiotípicas pueden asociarse a tolerancia. La supresión de un idiotipo por los anticuerpos antiidiotípicos puede alterar de forma significativa la respuesta…la tolerancia será parcial porque solo afecta a las células B que portan ese idiotipo.

13
FACTORES GENÉTICOS La capacidad de respuesta inmune depende de la genética del individuo. Mediante experimentos de genética molecular se ha determinado la existencia de genes que condicionan o modulan la respuesta inmune (en el ratón, genes Ir). • Influencia del CMH: El polimorfismo de secuencias CMH que cada individuo hereda (Tema 6). • Genes no ligados al CMH: Controlan la capacidad de respuesta global del organismo a antígenos complejos, como la capacidad del macrófago para procesar Ag o la tasa de proliferación de linfocitos B.
. • Influencia del CMH: El polimorfismo de secuencias CMH que cada. individuo hereda (Tema 6). • Genes no ligados al CMH: Controlan la capacidad de respuesta global. del organismo a antígenos complejos, como la capacidad del macrófago para procesar Ag o la tasa de proliferación de linfocitos B.")
14
NIVELES DE REGULACION DE LA EXPRESIÓN GENICA
EXPOSICIÓN DE UN DETERMINADO LOCUS A LA MAQUINARIA TRANSCRIPCIONAL. LA TRANSCRIPCIÓN SE INCICIA POR FACTORES DE TRANSCRIPCION NIVELES DE COMPACTACIÓN DE LA CROMATINA LA ACETILACIÓN DE HISTONAS MODIFICACIONES POSTRANSCRIPCIONALES TRANSPORTE DE TRANSCRIPTOS DESDE EL NUCLEO CELULAR LA ESTABILIDAD DEL RNA MENSAJERO

15
EN EL SI, LOS FACTORES DE TRANSCRIPCIÓN CONTROLAN PROCESOS BIOLOGICOS ESENCIALES
MADURACIÓN DE LOS LINFOCITOS ACTIVACION LINFOCITARIA DIFERENCIACIÓN DE CELULAS TCD4 ACTIVIDAD CITOTOXICA DE LINFOCITOS T Y NK DIFERENCIACIÓN DE CELULAS B FENOMENOS DE HOMEOSTASIS Y TOLERANCIA SELECCIÓN NEGATIVA ANERGIA CLONAL GENERACIÓN DE CEL T REG APOPTOSIS DE CÉLULAS T EFECTORAS

17
SISTEMA NERVIOSO- SISTEMA ENDÓCRINO
Actualmente existen evidencias que sugieren la existencia de una compleja red de comunicación tridireccional entre el sistema inmune, el sistema nervioso y el sistema endocrino. El estrés o una infección (inflamación) pueden activar y modificar el equilibrio de las citoquinas pro-inflamatorias, y activar tanto al eje hipotálamo-hipófisisadrenal (HPA) como al sistema simpático-adrenal, produciendo la liberación de hormonas como ACTH, glucocorticoides, y catecolaminas (adrenalina y noradrenalina) que inducen cambios cualitativos y cuantitativos en el sistema inmunitario.
 pueden activar y modificar el equilibrio de las citoquinas pro-inflamatorias, y activar tanto al eje hipotálamo-hipófisisadrenal (HPA) como al sistema simpático-adrenal, produciendo la liberación de hormonas como ACTH, glucocorticoides, y catecolaminas (adrenalina y noradrenalina) que inducen cambios cualitativos y cuantitativos en el sistema inmunitario.")
18
Actualmente existen evidencias que sugieren la existencia de una compleja red
de comunicación tridireccional entre el sistema inmune, el sistema nervioso y el sistema endocrino (Fig. 2). El estrés o una infección (inflamación) pueden activar y modificar el equilibrio de las citoquinas pro-inflamatorias, y activar tanto al eje hipotálamo-hipófisisadrenal (HPA) como al sistema simpático-adrenal, produciendo la liberación de hormonas como ACTH, glucocorticoides, y catecolaminas (adrenalina y noradrenalina) que inducen cambios cualitativos y cuantitativos en el sistema inmunitario. Hoy es conocido como el sistema endocrino y el sistema nervioso pueden contribuir a la regulación del sistema inmune. La confirmación de este fenómeno se realizó en los años setenta cuando por primera vez se demostró que los niveles de glucocorticoides se elevaban durante la respuesta inmunitaria produciendo un efecto supresor sobre la misma. Esta conexión ha sido después ampliamente demostrada en ambas direcciones de tal manera que, en su conjunto, se mantiene la homeostasis del organismo de una forma integrada con participación de los sistemas inmune, endocrino y nervioso. (Figura: Interacción diferentes sistemas).
. El estrés o una infección (inflamación) pueden activar y modificar el equilibrio. de las citoquinas pro-inflamatorias, y activar tanto al eje hipotálamo-hipófisisadrenal. (HPA) como al sistema simpático-adrenal, produciendo la liberación de. hormonas como ACTH, glucocorticoides, y catecolaminas (adrenalina y. noradrenalina) que inducen cambios cualitativos y cuantitativos en el sistema. inmunitario. Hoy es conocido como el sistema endocrino y el sistema nervioso pueden contribuir a la regulación del sistema inmune. La confirmación de este fenómeno se realizó en los años setenta cuando por primera vez se demostró que los niveles de glucocorticoides se elevaban durante la respuesta inmunitaria produciendo un efecto supresor sobre la misma. Esta conexión ha sido después ampliamente demostrada en ambas direcciones de tal manera que, en su conjunto, se mantiene la homeostasis del organismo de una forma integrada con participación de los sistemas inmune, endocrino y nervioso. (Figura: Interacción diferentes sistemas).")
19
Hoy es conocido como el sistema endocrino y el sistema nervioso pueden contribuir a la regulación del sistema inmune. La confirmación de este fenómeno se realizó en los años setenta cuando por primera vez se demostró que los niveles de glucocorticoides se elevaban durante la respuesta inmunitaria produciendo un efecto supresor sobre la misma. Esta conexión ha sido después ampliamente demostrada en ambas direcciones de tal manera que, en su conjunto, se mantiene la homeostasis del organismo de una forma integrada con participación de los sistemas inmune, endocrino y nervioso.

20
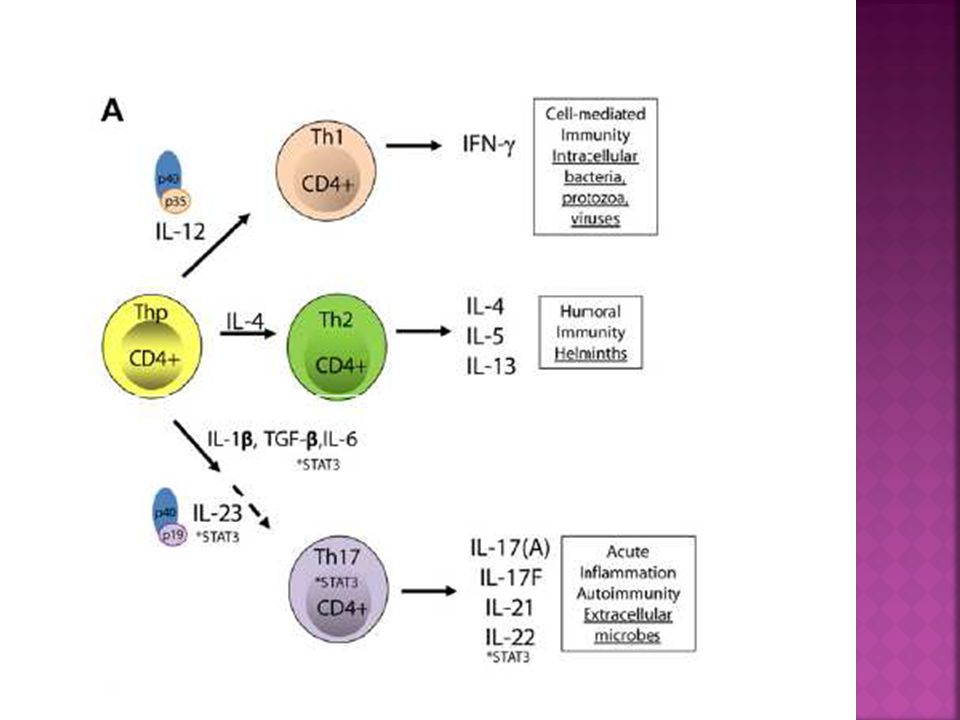
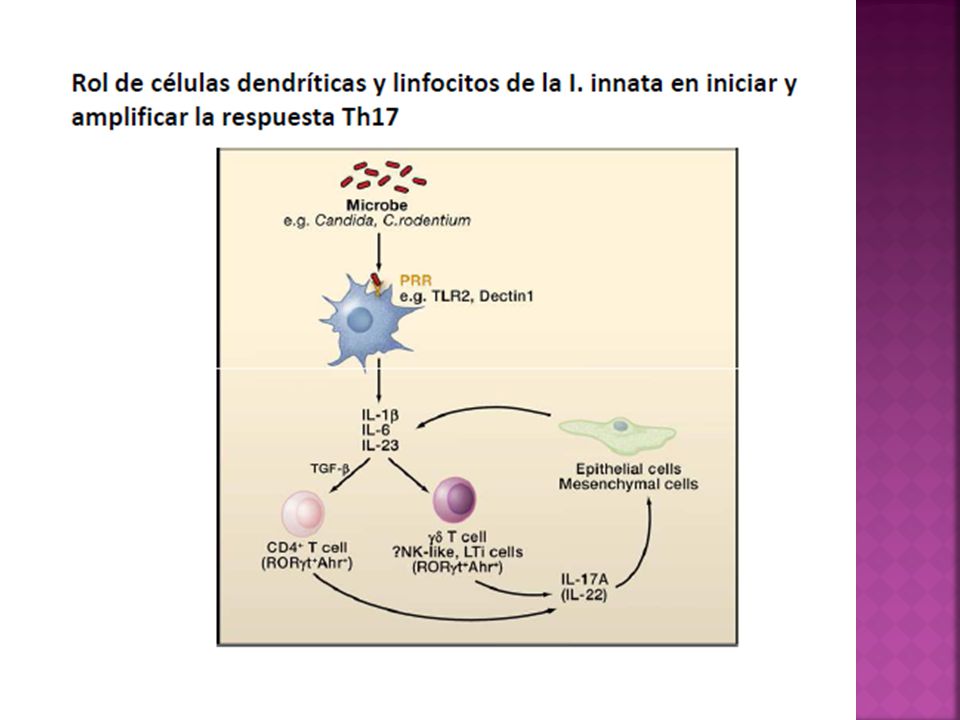
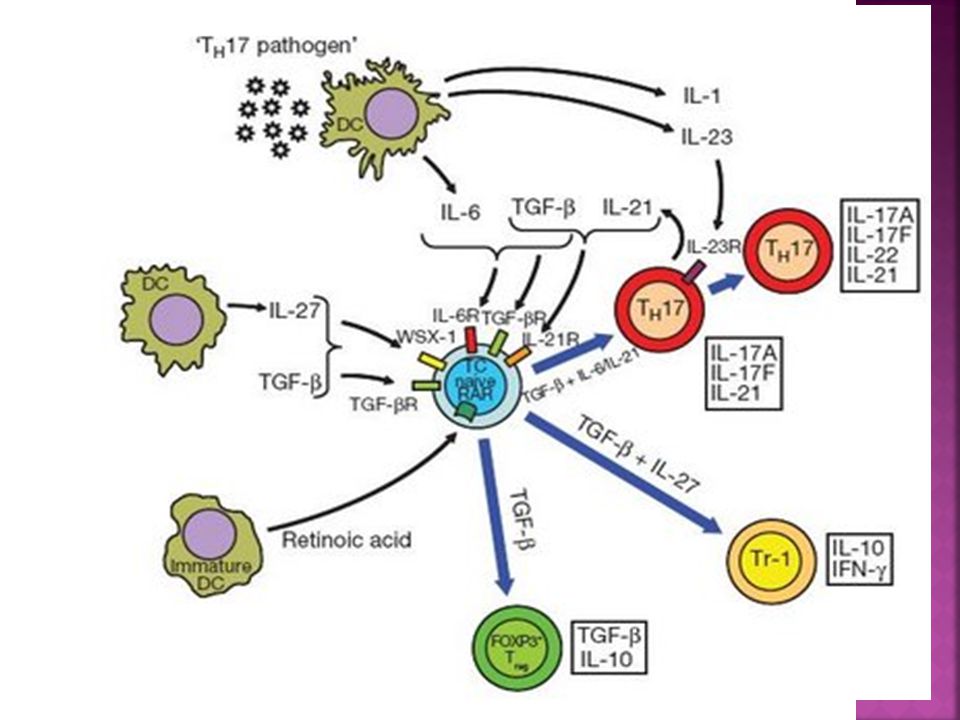
Th17 La combinación de TGF-β e IL-6, y los factores de transcripción RORγt, RORα y Stat3 son esenciales para comprometer el subtipo Th17. IL-23 juega un papel clave en la estabilización del fenotipo y de la actividad patogénica de células productoras de IL-17. La citocina IL-21 producida por células Th17 participa en un mecanismo de retroalimentación para favorecer el desarrollo de células productoras de IL-17, mientras que las citocinas IL-27, IL-4, IFN-γ, IL-25 e IL-2 limitan el fenotipo Th17.

21
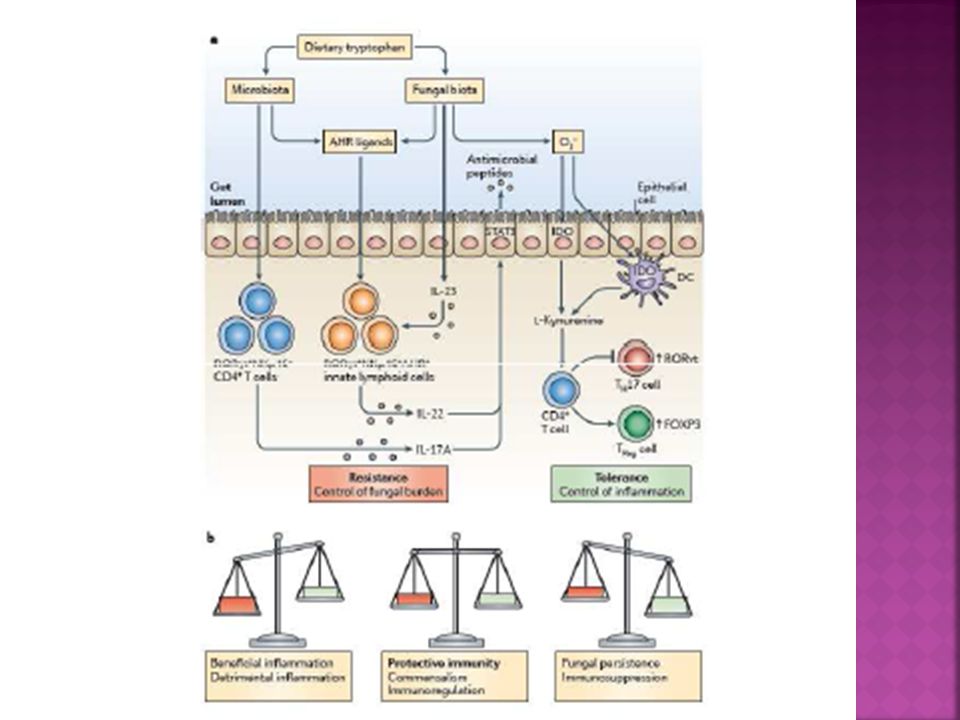
RESPUESTA TH17 Las células TCD4+ efectoras productoras de IL-17 representan un perfil de diferenciación. Las células TH17 cumplen un papel destacado no solo en las infecciones micóticas, sino también en la inmunidad frente a bacterias extracelulares y fenómenos de autoinmunidad. Constituye una línea de defensa propia de la inmunidad adaptativa en las superficies mucosas en el aparato digestivo, respiratorio y piel y nos protegen de las infecciones invasivas.

23
Las citocinas IL-1, IL-6, IL-21, IL23 y TGF-B desempeñan un papel importante en la diferenciación de un perfil TH17. IL-1 e IL-6 cumple una función crítica en la diferenciación. IL 21 e IL-23 contribuyen en la expansión.

24
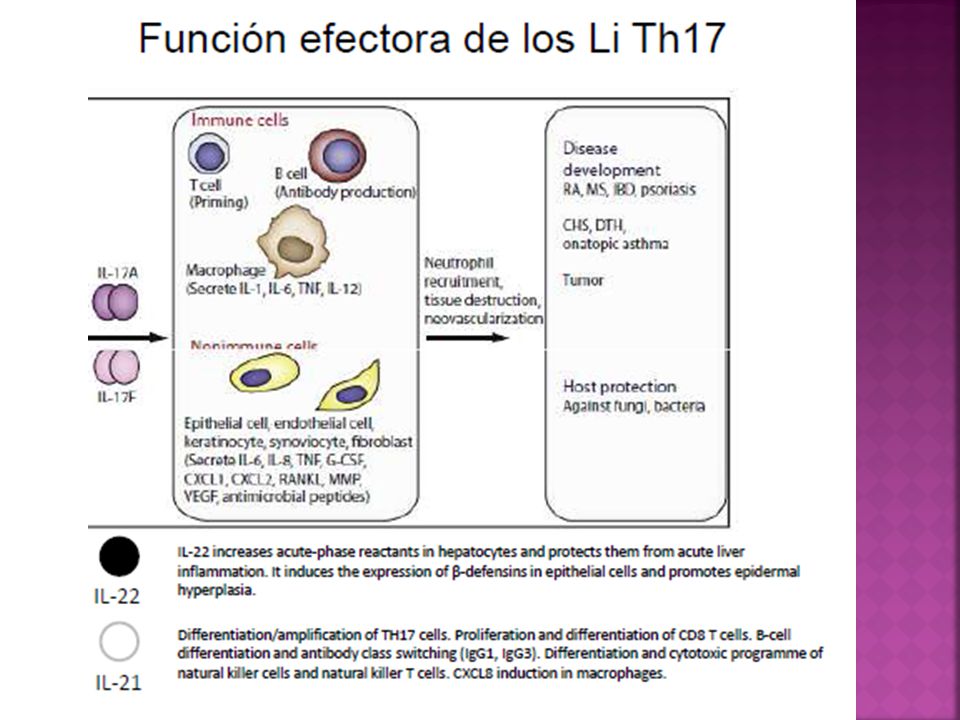
Mecanismos efectores Las células TH17 producen citocinas IL-17A, IL17F, IL- 21 e IL-22. Las dos primeras expresan una notoria capacidad proinflamatoria. Actual sobre un abanico de tipos celulares activando el factor NF-Kb estimulando la producción de citocinas inflamatorias ( TNF-α, IL-1, IL-6, GM-CSF y G-CSF), quimiocinas inflamatorias, mucinas, péptidos antimicrobianos y metaloproteasas que promueven la infiltración de tejido afectado por células reclutadas desde la circulación, particularmente granulocitos neutrófilos.
, quimiocinas inflamatorias, mucinas, péptidos antimicrobianos y metaloproteasas que promueven la infiltración de tejido afectado por células reclutadas desde la circulación, particularmente granulocitos neutrófilos.")
25
La IL-22 media también efectos proinflamatorios similares pero su receptor se encuentra en queratinocitos, cel. epiteliales, endoteliales y fibroblastos pero no en células inmunitarias.

27
Las IL-17 A e IL-17F no solo son producidas por células TH17, sino que también son producidas en altas cantidades durante la fase temprana de los procesos infecciosos por diferentes tipos celulares de la inmunidad innata como NKT y linfocitos Tγδ que puede cumplir un papel importante en el control de infecciones bacterianas y micóticas nacientes, en las mucosas intestinal y respiratorias

30
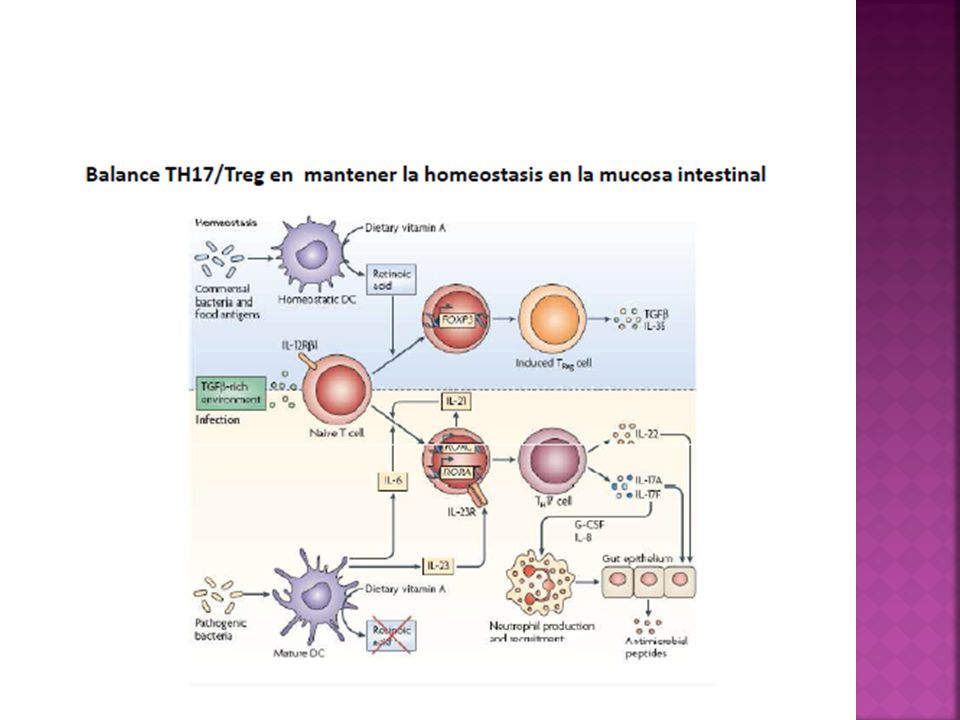
Células T reguladoras Las células T y B autorreactivas presentes en la periferia deben ser silenciadas a fin de evitar el desarrollo de enfermedades autoinmunes. Las células T reguladoras cumplen en este sentido un papel fundamental, por otra parte, modulan también los mecanismos propios de la inmunidad antiinfecciosa.

34
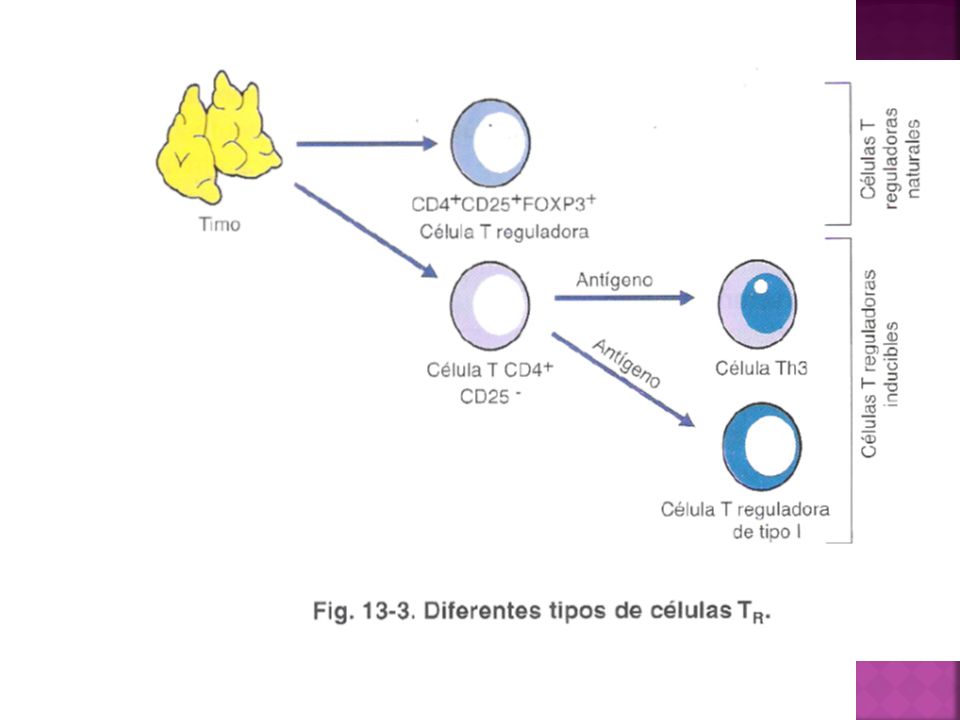
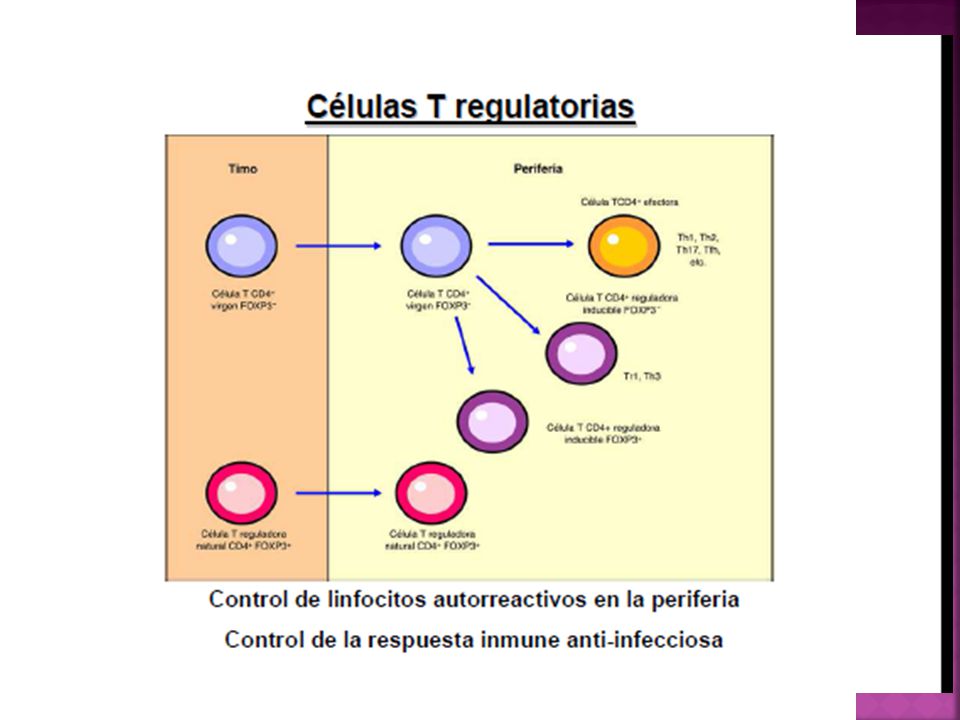
Las células T reguladoras pueden mostrar a grandes rasgos 2 orígenes diferentes.
Celulas T reguladoras naturales: emigran del timo expresando constitutivamente el factor de transcripción FOXP3, la molécula CD25 y una funcionalidad supresora. Células T virgenes convencionales pueden diferenciarse en los órganos linfáticos secundarios en T reguladoras

36
Células T reguladoras inducibles
Tr1 son células T reguladoras FOXP3- que median su actividad supresora a través de la producción de IL-10 TH3 son células T reguladoras FOXP3- que median su actividad supresora a través de la producción de TGF-B. Tolerancia Oral Tercera población que al activarse expresan FOXP3 y median un efecto supresor através de varios mecanismos

37
Células reguladoras inducibles
Adquieren su perfil supresor luego de activarse por el antígeno en los órganos linfáticos secundarios. Principalmente producen IL-10 y TGF-B. Numerosas evidencias sugieren que su inducción es dirigida por células dendríticas semimaduras presentes en los órganos linfáticos secundarios, que expresan un fenotipo caracterizado por baja expresión de CD40, baja producción de IL12 y alta producción de IL-10. CD que no han recibido las señales de peligro y arriban a los órganos linfáticos secundarios sin alcanzar su madurez.

38
Células reguladoras inducibles
La administración de un antígeno por vía oral o la estimulación in vitro de células T CD4+CD25- en presencia de factor de crecimiento transformador B induce la generación de células T CD4+ productoras de TGFB (Células Th3). Inhibición de las vías de señalización empleadas por el TGFB produce la activación espontánea de células T y la inducción de cuadros inflamatorios in vivo….….th3 tendrían un rol en los mecanismos de escape tumoral frente a la respuesta inmune.. El tgf-b ejerce su actividad supresora inhibiendo la diferenciación de linfocitos T CD4 y TCD8 naive en células efectoras y suprimiendo la actividad de estas células cuando ya se han diferenciado..bloqueando la transcripción de los factores transcripcionales Gata-3 y T-bet…factores críticos en la diferenciación de células TCD4 en TH1 y TH2…
. Inhibición de las vías de señalización empleadas por el TGFB produce la activación espontánea de células T y la inducción de cuadros inflamatorios in vivo….….th3 tendrían un rol en los mecanismos de escape tumoral frente a la respuesta inmune.. El tgf-b ejerce su actividad supresora inhibiendo la diferenciación de linfocitos T CD4 y TCD8 naive en células efectoras y suprimiendo la actividad de estas células cuando ya se han diferenciado..bloqueando la transcripción de los factores transcripcionales Gata-3 y T-bet…factores críticos en la diferenciación de células TCD4 en TH1 y TH2…")
40
A diferencia de……… Se generan en órganos linfáticos secundarios en el momento de reconocer al antígeno en condiciones TOLEROGÉNICAS. AUSENCIA DE CITOCINAS PROINFLAMATORIAS PRESENCIA DE IL-10 Y TGFB BAJA EXPRESIÓN DE MOLECULAS COESTIMULATORIAS PRODUCCIÓN DE ÁCIDO RETINOICO POR DC EN LA MUCOSA INTESTINAL

42
Tolerancia es un estado de falta de respuesta frente a un antígeno determinado

43
TOLERANCIA Experimentos que demostraron que es posible inducir un estado de tolerancia cuando los linfocitos inmaduros se exponen a antígenos extraños. 1960 Premio nobel de Medicina. Peter Medawar y , Frank Burnet. Peter Brian Medawar (28 de febrero de 1915 – 2 de octubre de 1987) – Biólogo brasileño nacionalizado inglés nacido en Río de Janeiro, Brasil. Estudió en la Universidad de Oxford y obtuvo su doctorado en Fue profesor de zoología en la Universidad de Birmingham. En 1949, Frank Burnet propuso la teoría de que el sistema inmunitario puede adquirir tolerancia a lo trasplantes de tejidos, evitando el rechazo, si se injertan células del donante al receptor cuando éste es aún un embrión o un recién nacido. Podo después, Medawar descubrió en el ganado vacuno que lo gemelos no idénticos acetan trasplantes de piel uno del otro, lo que verificaba la teoría de Burnet. Realizó el experimento definitivo, injertar células extrañas en embriones de ratón y comprobar despúes, con el ratón ya adulto, que acepta trasplantes de piel del mismo individuo. El descubrimiento revolucionó la inmunología y abrió el camino hacia los trasplantes de órganos en los seres humanos. En 1960, conjuntamente con Burnet, ganó el Premio Nobel de Fisiología y Medicina.
 – Biólogo brasileño nacionalizado inglés nacido en Río de Janeiro, Brasil. Estudió en la Universidad de Oxford y obtuvo su doctorado en Fue profesor de zoología en la Universidad de Birmingham. En 1949, Frank Burnet propuso la teoría de que el sistema inmunitario puede adquirir tolerancia a lo trasplantes de tejidos, evitando el rechazo, si se injertan células del donante al receptor cuando éste es aún un embrión o un recién nacido. Podo después, Medawar descubrió en el ganado vacuno que lo gemelos no idénticos acetan trasplantes de piel uno del otro, lo que verificaba la teoría de Burnet. Realizó el experimento definitivo, injertar células extrañas en embriones de ratón y comprobar despúes, con el ratón ya adulto, que acepta trasplantes de piel del mismo individuo. El descubrimiento revolucionó la inmunología y abrió el camino hacia los trasplantes de órganos en los seres humanos. En 1960, conjuntamente con Burnet, ganó el Premio Nobel de Fisiología y Medicina.")
44
Tolerancia…raton recien nacido

45
Conclusiones Dr. Medawar. Inducción de tolerancia a aloantígenos.
Ratón de la cepa A antes de su nacimiento, le implantó células alogénicas provenientes de un ratón B. Cuando el ratón A alcanzó el estado de adulto aceptó el injerto proveniente de B. La adquisición activa de tolerancia fue específica. La administración en el ratón de cepa A de células no tolerizadas fue capaz de romper el estado de tolerancia. En resumen los experimentos permitieron introducir un estado de tolerancia cuando los linfocitos inmaduros se exponen a antígenos extraños y definir la tolerancia inmunitaria como la falta de respuesta inmune a un antígeno provocado por la exposión anterior a ese antígeno.

46
La tolerización inducida por el reconocimiento antigénico en los linfocitos inmaduros es sólo aplicable a antígenos extraños?. TOLERANCIA CENTRAL

47
Mecanismos de Tolerancia Inmune

48
Tolerancia inmune Estado de “no respuesta” a un antígeno inducido por una exposición previa a ese antígeno. Por lo tanto es específica El desarrollo de tolerancia a auto- antígenos es necesario ya que al azar se genera una amplia diversidad de receptores específicos para antígenos y algunos de éstos, son autoreactivos Las células que expresan estos receptores deben ser eliminadas, ya sea física o funcionalmente

49
La tolerancia se puede desarrollar de un modo natural, como cuando un animal en desarrollo se vuelve incapaz de responder a sus propias moléculas (autototolerancia). Cuando este sistema falla, se producen patologías por autoinmunidad.

50
Tolerancia Inmune a lo Propio
Estado fisiológico que se adquiere en el cual el sistema inmune reconoce lo propio pero no establece una respuesta inmune que conlleve a la destrucción de los autoantígenos No hay una diferencia fundamental entre moléculas propias o extrañas para inducir un estado de tolerancia siempre que el antígeno se introduzca en el momento y lugar adecuado La mayor parte de la clase nos vamos a referir al desarrollo de tolerancia a ags propios aunque algunos de los mecanismos que mantienen la tolerancia a lo propio son similares o idénticos a mecanismos que permiten el desarrollo de tolerancia a otros antígenos. Y porque esto es necesario, básicamente para prevenir el desarrollo de respuestas inmunes patológicas: prevenir rechazo, enf. alérgicas, autoinmunes etc etc

51
Consecuencias del encuentro entre los linfocitos específicos y el antígeno
Antígeno inmunogénico Se genera una respuesta Activación Proliferación y diferenciación Anergia (No genera respuesta) Antígeno tolerogénico Tolerancia Apoptosis Muerte celular
 Antígeno tolerogénico. Tolerancia. Apoptosis. Muerte celular.")
52
Formas de Tolerancia Tolerancia Central Tolerancia Periférica
Garantiza la no respuesta a autoantígenos ubicuos Organos Linfoides Primarios Tolerancia Periférica Garantiza la no respuesta a autoantígenos específicos de tejidos Organos Linfoides Secundarios y Tejidos Pero no todas las proteinas propias se expresan en el timo, los timocitos que presentan especificidad para estos Ag no expresados en el timo, se escapan y se enc con el Ag propio en periferia

53
TOLERANCIA CENTRAL. Maduración de clonas que no reconocen antígenos
Precursor linfoide Maduración de clonas que no reconocen antígenos propios Eliminación de linfocitos específicos que reconocen antígenos propios presentes en órganos generativos.

54
TOLERANCIA PERIFÉRICA.
Antígeno propio de tejidos periféricos Antígeno extraño ELIMINACIÓN ANERGIA

55
Mecanismos de Tolerancia
La muerte celular por apoptosis (eliminación clonal) La inactivación funcional sin muerte celular (anergia clonal) La supresión de la activación y de las funciones efectoras por los linfocitos reguladores/supresores Algunos autoantígenos son ignorados por el sistema inmune de forma que los linfocitos autorreactivos permanecen viables y funcionales, pero no reaccionan contra los autoantigenos en forma detectable (ignorancia clonal). Estrictamente no corresponde a un mecanismo de tolerancia inmune La apoptosis es el principal mecanismo de tolerancia central y los tres mecanismos funcionan para establecer tolerancia periférica.
 La inactivación funcional sin muerte celular (anergia clonal) La supresión de la activación y de las funciones efectoras. por los linfocitos reguladores/supresores. Algunos autoantígenos son ignorados por el sistema inmune. de forma que los linfocitos autorreactivos permanecen. viables y funcionales, pero no reaccionan contra los. autoantigenos en forma detectable (ignorancia clonal). Estrictamente no corresponde a un mecanismo de tolerancia inmune. La apoptosis es el principal mecanismo de tolerancia central y los tres mecanismos funcionan para establecer tolerancia periférica.")
56
Selección clonal de LT en el timo
Las células pre-T llegan de médula ósea al timo, donde ocurre el proceso de selección Es un proceso riguroso que resulta en la muerte del 90-95% de los LT La selección se da con base en la afinidad del TCR al complejo CMH-péptido Corteza Médula Selección negativa Selección positiva APOPTOSIS

57
Timo: Apoptosis por negligencia (selección +)
")
58
Tolerancia en el timo: Apoptosis durante la selección -
“Muerte activa” Depende de la concentración de antígeno y de la afinidad del TCR

59
Pasos en la selección y maduración de los linfocitos T

60
TOLERANCIA PERIFÉRICA DE CÉLULAS T
1. ANERGIA: la activación de células T requiere señales coestimulatorias además de las señales derivadas de la unión al antígeno. Ante la ausencia de señales coestimulatorias, la célula se convierte en “anérgica”. ANERGIA: Ausencia de respuesta funcional

61
LINFOCITOS T 1.- TOLERANCIA CENTRAL 2.- TOLERANCIA PERIFÉRICA Deleción clonal Anergia clonal Ignorancia inmunológica Células T reguladoras LINFOCITOS B

62
Si las células T CD4+ reconocen un antígeno peptídico
La inactivación funcional o anergia es inducida por el reconocimiento del antígeno sin la coestimulación suficiente Si recibe las dos señales TCR MHC-pep T CD28 B7 Si las células T CD4+ reconocen un antígeno peptídico presentado por una CPA que no tenga coestimulación, las células T se vuelven incapaces de responder al antígeno T ANERGIA

63
Se ha implicado señales a través de receptores
alternos en el proceso de anergia. T ANERGIA (-) CTLA-4 B7 CTLA-4 homólogo a CD28 y se une a B7, trasmite señales que inhiben la activación de la célula T
 CTLA-4. B7. CTLA-4 homólogo a CD28 y se une a B7, trasmite señales que inhiben la activación de la célula T.")
64
ANERGIA

65
2. Muerte celular inducida por la activación: apoptosis
Se proponen 2 mecanismos: a) En la cél T el reconocimiento del Ag induce producción de proteínas proapoptóticas . En la respuesta a patógenos éstas son contrarestadas por proteínas anti-apoptóticas inducidas por co-estimulación y 2º señales. Los Ag propios no estimulan la producción de proteínas anti-apoptóticas Apoptosis
 En la cél T el reconocimiento del Ag induce producción de proteínas proapoptóticas . En la respuesta a patógenos éstas son contrarestadas por proteínas anti-apoptóticas inducidas por co-estimulación y 2º señales. Los Ag propios no estimulan la producción de proteínas anti-apoptóticas. Apoptosis.")
66
Activación de cascada de enzimas citosólicas
b) En los LT CD4+ la activación repetida lleva a la co-expresión del receptor Fas y su ligando FasL. FasL T Fas Los Ag propios pueden eliminar cél T específicas porque están presentes a lo largo de toda la vida y son capaces de estimular repetidamente. Activación de cascada de enzimas citosólicas Apoptosis
 En los LT CD4+ la activación repetida lleva a la co-expresión del receptor Fas y su ligando FasL. FasL. T. Fas. Los Ag propios pueden eliminar cél T específicas porque están presentes a lo largo de toda la vida y son capaces de estimular repetidamente. Activación de cascada de enzimas citosólicas. Apoptosis.")
67
Tolerancia periférica T

68
3. Mecanismos reguladores
3.1. La reactividad de las células T podría ser controlada indirectamente por una subpoblación diferente de células T con funciones reguladoras. 3.2. Citoquinas reguladoras

69
Generación de cel. T reguladoras naturales
Downloaded from: StudentConsult (on 2 May :00 PM) © 2005 Elsevier
 © 2005 Elsevier.")
70
Ignorancia Inmunológica
Secuestro de antígenos Hay sitios anatómicos donde existe mayor dificultad de que las células del sistema inmune tengan acceso; SNC, ojo, testículos

71
Tolerancia de LB Las células B en su mayoría son dependientes de las células T Central: en la médula ósea Eliminación Anergia Reducción expresión receptor Ag Edición del receptor Periférica: en el centro germinal Ausencia de ayuda T Apoptosis Se reuiere para establecer toelrancia a CHs y lipidos

72
1. Eliminación de células B
Tolerancia central 1. Eliminación de células B Se da en la médula ósea: Las células B en diferenciación que expresan receptores de inmunoglobulinas, con alta afinidad de unión para antígenos propios son eliminadas por apoptosis 7 7

73
2. Anergia de Células B Se produce cuando las células B, con el potencial para responder a proteínas propias solubles, inhiben sus receptores de superficie IgM La union se da con menor afinidad que cuando se induce apoptosis 8 8

74
3. Edición del receptor de la célula B
Se da en la fase de LB inmaduros (IgM+) Ocurre una recombinación adicional de la cadena liviana
 Ocurre una recombinación adicional de la cadena liviana.")
75
4. Disminución en la expresión del receptor de LB:

76
Tolerancia periférica
1. Apoptosis

77
Apoptosis via Fas-FasL

78
2. Anergia Downloaded from: StudentConsult (on 2 May 2005 07:00 PM)
© 2005 Elsevier

79
Tolerancia Artificial
Puede inducirse artificialmente por varios esquemas que eventualmente pueden utilizarse clínicamente, para evitar el rechazo de transplantes y para el tratamiento de enfermedades autoinmunes, alergias 10 10

80
La tolerancia inducida experimentalmente es un estado de ausencia de respuesta a un antígeno que normalmente sería inmunogénico. Para ello el antígeno ha de ser administrado bajo ciertas condiciones.

81
Los antígenos extraños pueden ser administrados por vías que preferencialmente inhiben respuestas inmunes por la inducción de tolerancia en linfocitos específicos. La inducción de la tolerancia inmunológica puede ser explotada como una alternativa terapéutica para prevenir respuestas inmunes dañinas.

82
Propiedades de la inducción de la tolerancia
La tolerancia es un estado adquirido ("aprendido"), no innato. Se induce más fácilmente en linfocitos inmaduros. Se induce cuando no hay señal coestimulatoria. Requiere que el antígeno persista para que dicho estado permanezca.
, no innato. Se induce más fácilmente en linfocitos inmaduros. Se induce cuando no hay señal coestimulatoria. Requiere que el antígeno persista para que dicho estado permanezca.")
83
NEONATO Injerto de CEPA B CEPA A Injerto de CEPA B CEPA A
6 semanas 7-14 DÍAS CEPA A Inyección de leucocitos de CEPA B Injerto de CEPA B 6 semanas 7-14 DÍAS CEPA A Inyección de leucocitos de CEPA B Injerto de CEPA C 7-14 DÍAS 6 semanas

84
Inducción de tolerancia
Naturaleza del antígeno: soluble (tolerogénico); particulado (inmunogénico) Vía de entrada: oral, sanguíneo (tolerogénico); intradérmico, muscular (inmunogénico) APCs DC tolerogénicas, B cells, APC no profesionales (tolerogénicas) DC Citocinas IL-10, TGF-b suelen ser tolerogénicas
; particulado (inmunogénico) Vía de entrada: oral, sanguíneo (tolerogénico); intradérmico, muscular (inmunogénico) APCs. DC tolerogénicas, B cells, APC no profesionales (tolerogénicas) DC. Citocinas. IL-10, TGF-b suelen ser tolerogénicas.")
85
CÉLULAS DENDRÍTICAS INMADURAS
CÉLULAS DENDRÍTICAS INMADURAS. Mantenimiento de la tolerancia a autoantígenos Se describió una población de células TR CD8+CD28- capaces de inducir en las CD la expresión de ILT3 e ILT4 (immunoglobulin-like transcripst) que inhiben tanto la expresión de moléculas co-estimulatorias como la señalización a través de CD40.
 que inhiben tanto la expresión de moléculas co-estimulatorias como la señalización a través de CD40.")
86
Tolerancia periférica de linfocitos B
Linfocitos B capaces de reconocer autoantígenos que hayan escapado a los mecanismos de delección propios de la tolerancia central podrían madurar sin convertirse en peligrosos ya que no contarían con linfocitos T autorreactivos que puedan colaborar con ellos para su expansión clonal y diferenciación.

87
Homeostasis de la respuesta inmune antimicrobiana
Las células T efectoras son células de vida media corta. La eliminación del antígeno priva a las células T efectoras de señales co-estimulatorias y citocinas (ambas producidas durante el reconocimiento antigénico) y conduce a una rápida pérdida en la expresión de proteínas antiapoptóticas.
 y conduce a una rápida pérdida en la expresión de proteínas antiapoptóticas.")
88
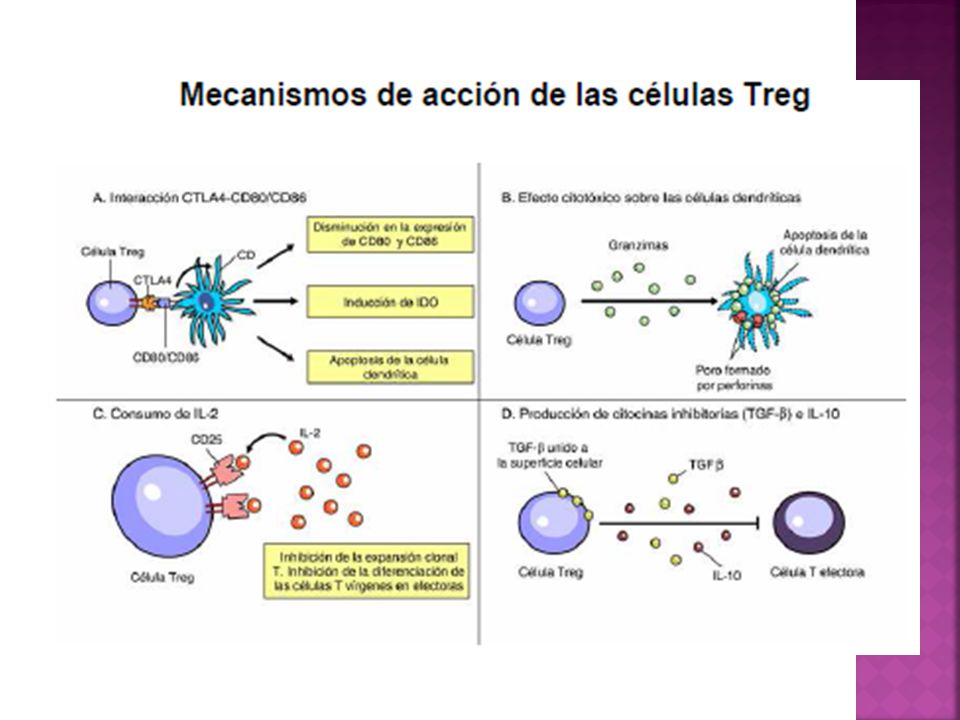
Inhibición de la expansión clonal T mediada por CTLA-4
El receptor CTLA-4 se expresa en los linfocitos T luego de su activiación y envía señales que inhiben la transcripción de la IL-2 y la progresión de las células T en el ciclo celular. CTLA-4 posee alta afinidad por las moléculas CD80/CD86 sustancialmente mayor que CD28

89
Muerte celular inducida por el sistema Fas/FasL
Mientras que las células T expresan en forma constitutiva la molécula Fas, su activación conduce a la expresión de FasL. La interacción FasL-Fas desencadena la apoptosis de los linfocitos T efectores.

90
Las células T reguladoras cumplen una importante función no sólo en el control de la autorreactividad periférica, sino también en el de la respuesta inmune antimicrobina y antitumoral.

91
Las células que han escapado a la selección negativa en el timo siguen sujetas a control en la periferia secuestro. El autoantígeno se oculta físicamente o funcionalmente (carece de APC) del sistema inmunitario. Sitio privilegiado. El fas o las citocinas (TGFB, IL10) impiden que las células T respondan al autoantígeno. Homeostasis. El tamaño del reservorio de los linfocitos T está controlado por la apoptosis (muerte celular inducida por activación o muerte celular programada), la competencia por factores de crecimiento. Células t reguladoras naturales e inducidas.
 del sistema inmunitario. Sitio privilegiado. El fas o las citocinas (TGFB, IL10) impiden que las células T respondan al autoantígeno. Homeostasis. El tamaño del reservorio de los linfocitos T está controlado por la apoptosis (muerte celular inducida por activación o muerte celular programada), la competencia por factores de crecimiento. Células t reguladoras naturales e inducidas.")
92
Los antígenos solubles inducen tolerancia con facilidad
Puede inducirse tolerancia en neonatos y adultos administrnado antígenos solubles.. Las células T se hacen tolerantes desde el bazo y el timo después de dosis muy bajas del antígeno y en unas horas.. La tolerancia de las células B del bazo requiere mucho más tiempo y dosis más altas de antígeno

93
Una proliferación clonal amplia puede dar lugar al agotamiento y la tolerancia
La exposición antigénica repetida puede estimular a todas las células que responden al antígeno para que se diferencien en células finales de vida corta, sin dejar células que puedan responder a una exposición posterior al antígeno

94
BIBLIOGRAFIA Fainboim Geffner. Introducción a la Inmunología Humana 6 edición. Editorial Panamericana Abbas AK, Lichtman AH. Basic Immunology. Functions and disorders of the immune system, 3rd edition. Saunders (Elsevier); 2009
;")
Presentaciones similares