Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Ricardo Ariel Floridia
Inmunología de las parasitosis Ricardo Ariel Floridia

2
Definición: El parásito
Ser vivo que de manera temporal o permanente vive a expensas de otro organismo de distinta especie (huésped), obteniendo de éste nutrición y hábitat, al que puede producir daño de variada intensidad y con el que tiene una dependencia obligada y unilateral. Clasificación: Protozoos Helmintos Artrópodos
, obteniendo de éste nutrición y hábitat, al que puede producir daño de variada intensidad y con el que tiene una dependencia obligada y unilateral. Clasificación: Protozoos. Helmintos. Artrópodos.")
3
¿Cuál es el éxito del parásito?
El parasitismo es un fenómeno biológico interdependiente y dinámico entre dos organismos. El éxito del parásito se mide, no por los trastornos que le causa a su huésped, sino por su capacidad para adaptarse e integrarse al mismo. Desde el punto de vista inmunológico, el éxito del parásito se da si se puede integrar al huésped de manera que este no lo considere exógeno.

5
Infecciones parasitarias
Desencadenan una serie mecanismos de defensa inmunitaria característicos. Algunos mediados por anticuerpos y otros por células. La eficacia de cada uno depende del parasito implicado y de la fase de la infección. Es importante la vía de ingreso de los parásitos, que difiere en muchos de ellos, donde la respuesta inmune va a ser diferente también, como así también el sitio final donde se alojará.

7
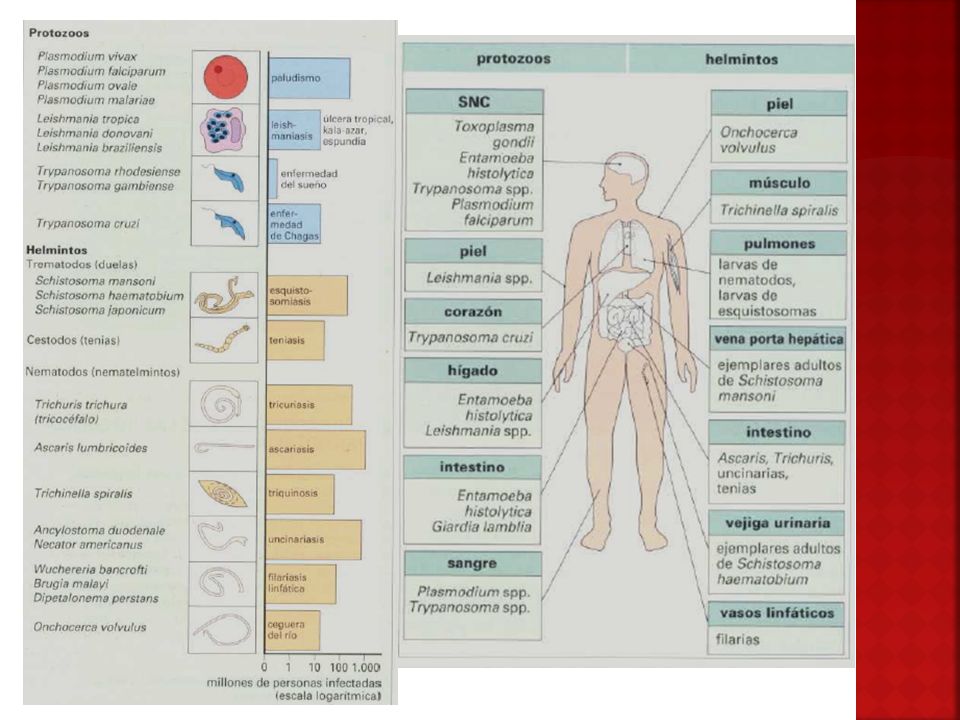
Localizaciones de los parásitos
Enteroparasitosis Histoparasitosis Hemoparasitosis Ectoparasitosis

8
Antígenos parasitarios
Pueden ser: Proteínas Lípidos Polisacárido Ácidos nucleicos Las proteínas se encuentran conjugadas con Hidratos de carbono y los lípidos constituyen complejos con proteínas y polisacáridos. También presentan Haptenos como fosforilcolina o AG de Forssman

9
Ubicación de los antígenos
AG Somáticos (estructurales o endoantígenos) Originan gran variedad de anticuerpos de una menor importancia, porque para su reconocimiento el parásito debe ser destruido. AG Metabólicos (productos de secreción, excreción o exoantígenos) Muchos de ellos son enzimas. Originan anticuerpos mas protectores y mas específicos. Permiten llegar a diferenciar cepas.
 Originan gran variedad de anticuerpos de una menor importancia, porque para su reconocimiento el parásito debe ser destruido. AG Metabólicos (productos de secreción, excreción o exoantígenos) Muchos de ellos son enzimas. Originan anticuerpos mas protectores y mas específicos. Permiten llegar a diferenciar cepas.")
10
Propiedades del Antígeno
Inmunogénicos: es la capacidad de inducir una respuesta inmunológica, ya sea mediada por anticuerpos o por células. Antigenicidad: es la capacidad de combinarse específicamente con el producto final de la respuesta inmunológica.

11
Inmunogenicidad

12
Hapteno Inmunógeno

13
Respuestas inmunes Anticuerpos: son necesarios para eliminar parásitos extracelulares en la sangre, intestino y otros líquidos tisulares. Célular: para eliminar parásitos intracelulares. El tipo de respuesta depende del parásito involucrado, y del ciclo, nunca son respuesta puras. Ejemplo en el paludismo

14
El sistema inmune Inmunidad innata: Inmunidad adquirida:
Barreras Físicas: Piel, mucus, cilios, tos, estornudo, peristaltismo intestinal. Barreras químicas: pH ácido, Sales biliares, ácidos grasos, lisozima, lactoferrina, transferrina. Barreras biológicas: microbiota normal Componentes del sistema inmune: Células: Fagocitos, Macrófagos, PMN, NK, complemento Inmunidad adquirida: Linfocitos: T y B Anticuerpos

16
Receptores tipo toll Son una familia de proteínas que conforman receptores del sistema inmunitario innato. Reconocen patrones moleculares expresados por un amplio espectro de agentes infecciosos, y estimulan una variedad de respuestas inflamatorias. Forman parte de los “pattern recognition receptors” (PRRs) Reconocen a los PAMPs (Patrones moleculares asociados a patógenos) Esta señalización representa el vínculo entre la respuesta inmune innata y la adaptativa.
 Reconocen a los PAMPs (Patrones moleculares asociados a patógenos) Esta señalización representa el vínculo entre la respuesta inmune innata y la adaptativa.")
17
Receptores tipo Toll

18
Pamps (Patrones moleculares asociados a patógenos)
PAMPs: “pathogen associated molecular patterns”. Estructuras conservadas en distintos grupos de microorganismos, generalmente esenciales para su sobrevida Reconocen sus ligandos en forma de dímeros desde la membrana celular o desde endosomas Los diferentes TLRs estan apropiadamente ubicados en estos distintos compartimentos celulares porque reconocen distintos PAMPs Membrana plasmática TLR1, 2, 4 , 5, 6, 11 Componentes de membrana de microorganismos, lípidos, lipoproteínas, proteínas Endosomes, endolisosomas TLR3, TLR7, 8 y 9 Acidos Nucleicos

19
Características de los PAMPs
son patrimonio de los patógenos pero no de sus huéspedes son esenciales para la sobrevida o patogenicidad del microorganismo son estructuras invariantes compartidas por clases enteras de patógenos (conservadas evolutivamente) Ejemplos de PAMPs LPS Peptidoglicano Acido lipoteicoico manosa de oligosacáridos microbianos DNA conteniendo motivos CpG no metilados RNA doble cadena
 Ejemplos de PAMPs. LPS. Peptidoglicano. Acido lipoteicoico. manosa de oligosacáridos microbianos. DNA conteniendo motivos CpG no metilados. RNA doble cadena.")
20
Relación entre tlr y respuesta adaptativa

21
Activación del sistema inmune adaptativo

22
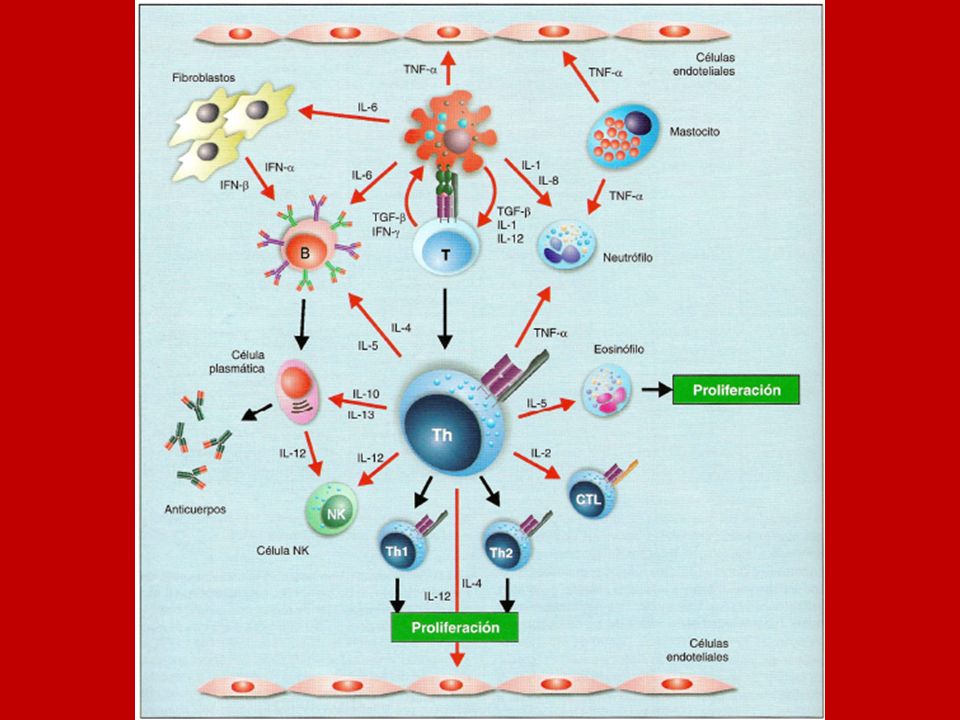
Mecanismos efectores LT Th1 Citoquinas LT Th2 Citoquinas
L TCD8 LT Citotóxica efectora Citoquinas LT Th1 LT Th2 Macrófago activado L TCD4 Anticuerpos neutralizantes Y Célula Plasmática Anticuerpos aglutinantes L B Y Citoquinas Anticuerpos fijadores de Complemento

23
A nivel de receptores

24
La polarización de la respuesta T es influenciada por la interacción PAMPs -PRR
INNATA ADAPTATIVA Discriminación PAMP-PPR Instrucción CITOQUINAS MOLEC. COESTIMUL Polarización TH1 TH 2 Treg

25
Primera línea de defensa
Formada por macrófagos, neutrófilos, eosinófilos, y plaquetas. Evidencias: Eliminación de estas célula en ratones produce aumento de la susceptibilidad frente a la infección contra cercarias de S. mansoni Macrófagos hepáticos y fagocitos del bazo eliminan tripanosomas y parásitos del paludismo, en forma mas eficiente si los mismos se encuentran opsonizados con el complemento. Los macrófagos tiene doble función: como efectoras fagocíticas y como presentadoras de antígenos. Secretan moléculas que regulan respuestas inflamatorias (IL 1, IL12, TNFa, CFS)activan he inducen proliferación. Moléculas antiinflamatorias e inmunosupresoras como IL 10, prostaglandinas, TGFb
activan he inducen proliferación. Moléculas antiinflamatorias e inmunosupresoras como IL 10, prostaglandinas, TGFb.")
26
Los macrófagos Importante frente a parásitos de menor tamaño.
Fagocitosis Secreción de factores tóxicos. Activación por citoquinas: pueden destruir tanto pequeños parásitos extracelulares, fases eritrocitarias del paludismo, o larvas de esquistosomas. Pueden actuar como asesinas mediante un mecanismo de citotoxicidad mediada por células y dependiente de anticuerpos. Secretan citoquinas como IFNa, IL1 que pueden inducir resistencia de los hepatocitos a los parásitos del paludismo.

27
Los macrófagos Poco tiempo después de la infección se produce una estimulación de las funciones efectoras de los macrófagos. Macrofagos y fagocitos inducen compuestos reactivos del oxigeno frente a la fagocitosis de parásitos (T. cruzi, T. gondii, leishmania, paludismo, filarias, esquistosomulas). Oxido nítrico: potente toxina no dependiente del oxígeno, estimulada por IFNg, TNFa con gran respuesta frente a las dos citoquinas en conjunto.
. Oxido nítrico: potente toxina no dependiente del oxígeno, estimulada por IFNg, TNFa con gran respuesta frente a las dos citoquinas en conjunto.")
28
Los macrófagos Para la activación específica es necesario las citoquinas secretadas de células T: (IFNg, GM-CSF, IL3, IL4), también se activan por medio de vías no dependientes de células T: Células NK secretan IFNg cuando son estimuladas por IL12 Macrófagos secretan TNFa en respuesta a productos parasitarios. TNFa estimula a macrofagos, eosinófilos, y plaquetas para destruír parásitos, efecto potenciado por IFNg. El TNFa puede tener efectos negativos según la cantidad secretada y la circulacion del compuesto
, también se activan por medio de vías no dependientes de células T: Células NK secretan IFNg cuando son estimuladas por IL12. Macrófagos secretan TNFa en respuesta a productos parasitarios. TNFa estimula a macrofagos, eosinófilos, y plaquetas para destruír parásitos, efecto potenciado por IFNg. El TNFa puede tener efectos negativos según la cantidad secretada y la circulacion del compuesto.")
29
Los neutrófilos Tienen naturaleza fagocítica igual a los macrófagos.
Destruyen a parásitos por los siguiente mecanismos: Dependientes del oxígeno (Estallido respiratorio mas intenso que los macrófagos Oxido nítrico Reacciones citotoxicas mediada por anticuerpos (Receptores FC) Son activados por citoquinas (IFNg, TNFa, GM-CSF. Se acumulan en regiones infectadas de parásitos
 Son activados por citoquinas (IFNg, TNFa, GM-CSF. Se acumulan en regiones infectadas de parásitos.")
30
Los eosinófilos Están asociados a infecciones por helmintos.
Actúan mediante mecanismos dependientes e independientes del oxígeno Son menos eficientes como fagocitos que los neutrófilos. Se desgranulan fácilmente cuando se unen a la IgE o IgG que opsoniza al parásito. Son estimulados por las citoquinas: TNFa, GM-CSF, pero necesitan de su unión al antígeno. Las lesiones son causadas por la proteína básica principal (MBP)
")
31
Los eosinófilos Actúan coordinadamente con los mastocitos.
Los antígenos parasitarios producen la desgranulación de los mastocitos dependiente de IgE liberando mediadores. Los mastocitos atraen eosinofilos a la zona de infección. Sus gránulos liberan: Proteína básica principal Proteína Cationica Neurotoxina Peroxidasa

32
Mastocitos Es una célula diseñada para organizar la inflamación de emergencia. Están muy próximos a los vasos sanguíneos Pueden regular localmente la permeabilidad vascular. EI patógeno los activa liberando grandes cantidades de mediadores inflamatorios preformados. Gránulos Aminas vasoactivas (histamina) Proteasas Neutras Factores Quimiotácticos Citoquinas (TNFa) Metabolitos del ac. Araquidónico
 Proteasas Neutras. Factores Quimiotácticos. Citoquinas (TNFa) Metabolitos del ac. Araquidónico.")
33
Acción de los mastocitos

34
Células NK Actividad citolítica innata(citotoxicidad mediada por receptores NK y secreción de IFN-g) Inmunidad contra infecciones por bacterias y parásitos intracelulares (activación de macrófagos por secreción de IFN-g) CCDA: citotoxicidad celular dependiente de Ac (receptores para fracción constante de Ac), liberan granzimas y perforinas. Regulación de la respuesta inmune a través de un diálogo recíproco con células dendríticas y secreción de citoquinas. A diferencia de LT CD8+ no tienen TCR
 CCDA: citotoxicidad celular dependiente de Ac (receptores para fracción constante de Ac), liberan granzimas y perforinas. Regulación de la respuesta inmune a través de un diálogo recíproco con células dendríticas y secreción de citoquinas. A diferencia de LT CD8+ no tienen TCR.")
35
Células NK Tienen receptores innatos capaces de detectar la ausencia de moléculas de histocompatibilidad. Un receptor innato activador (por ejemplo, NKRP1) reconoce azucares comunes en las células y otro receptor innato, pero inhibidor (KIR), sondea la membrana. Si hay MHC, no hay lisis porque el KIR la prohíbe. Si no hay MHC, el KIR no puede salvar a la diana. Además, como pasaba con los fagocitos, la opsonizaci6n de la célula infectada permite su reconocimiento por los Iinfocitos NK a través de sus receptores FcR.
 reconoce azucares comunes en las células y otro receptor innato, pero inhibidor (KIR), sondea la membrana. Si hay MHC, no hay lisis porque el KIR la prohíbe. Si no hay MHC, el KIR no puede salvar a la diana. Además, como pasaba con los fagocitos, la opsonizaci6n de la célula infectada permite su reconocimiento por los Iinfocitos NK a través de sus receptores FcR.")
36
Las plaquetas Atacan a muchos parásitos (T. cruzi, T. gondii)
Su actividad citotoxica se potencia con citoquinas (IFNg, TNFa) Tienen receptores para IgE
 Tienen receptores para IgE.")
37
La célula T Son centrales en el control de la infección:
Ayudan a eliminar la infección secretando citoquinas pro- inflamatorias (IFNg, IL3, GM-CSF, IL5) Modulan la infección: (IL10, IL4) Según el parásito y la fase del ciclo del mismo se van a activar las células T CD4+ o CD8+ que ejercen funciones protectoras diferentes. TCD4+ secretan diferentes citoquinas entre ellas INFg TCD8+ que además de secretar IFNg produce citoxicidad celular. Estimula a células NK secretando IL12
 Modulan la infección: (IL10, IL4) Según el parásito y la fase del ciclo del mismo se van a activar las células T CD4+ o CD8+ que ejercen funciones protectoras diferentes. TCD4+ secretan diferentes citoquinas entre ellas INFg. TCD8+ que además de secretar IFNg produce citoxicidad celular. Estimula a células NK secretando IL12.")
38
La células T En el comienzo de una infección las células T CD4 pueden ser TH1, TH2, TH17, y luego este perfil se va modificando según la evolución de la enfermedad. TH1 y TH2 son antagónicas TH1 y TH17: son perfiles inflamatorios celulares. TH2: perfil anti-inflamatorio, mediado por anticuerpos

39
Células TH1 Potencia respuestas defensivas frente a protozoos intracelulares. El INFg activa macrófagos para que destruya los protozoos que viven en el interior (Leishmania, tripanosoma, toxoplasma) y desactiva respuesta TH2. Potente respuestas frente a otros parásitos intracelulares
 y desactiva respuesta TH2. Potente respuestas frente a otros parásitos intracelulares.")
40
Células TH2 Las células Th2 están involucradas en la respuesta inmune humoral y la defensa del huésped contra los parásitos extracelulares. Se caracterizan por la producción de IL4, IL5, IL6, IL10 y IL13 Citocinas como la IL4 en general, estimulan la producción de anticuerpos dirigidos hacia los grandes parásitos extracelulares, mientras que la IL5 estimula la respuesta de eosinófilos a los grandes parásitos extracelulares.

41
Células t cd8+ Tras la infección con virus, bacterias o protozoos, el antígeno específico de linfocitos T CD8 + genera un proceso de diferenciación y proliferación para dar origen a células efectoras. Liberan IFNg

42
Células dendríticas Tienen largas prolongaciones citoplasmáticas repletas de receptores antigénicos. Se ubican en: Órganos linfáticos Epitelio de la piel y la mucosa del aparato digestivo y respiratorio. Pueden ser Mieloides Plasmocitoides

43
Células dendríticas Son células presentadoras de antígenos profesionales. Son nexo entre las respuesta inmune innata y la adptativa. Pueden endocitar y procesar los antígenos para ser presentado en la superficie, unido al complejo mayor de histocompatibilidad. Posee moléculas coestimulante para activar mejor al linfocito T. Activan linfocitos T vírgenes quienes se diferencian en células efectoras Activan la inmunidad celular: macrófagos y linfocitos T efectores. Activan la respuesta humoral por estimulación de linfocitos B y la producción de anticuerpos.

44
Acción de los anticuerpos
Dañan directamente los protozoos, o por acción del complemento. Neutralizan directamente a un parásito Estimulan la fagocitosis Opsonización y citoxicidad mediada por anticuerpos. El incremento de IgE se relaciona con algunas parasitosis. La IgG4 bloque efectos de la IgE Son IgG (85%), IgA, IgM, IgE
, IgA, IgM, IgE.")
45
El carácter crónico de una infección
Fin del parásito: perdurar en el tiempo. Trata que el huésped no muera hasta que se asegure la transferencia a otro individuo. ¿Qué modifica la respuesta inmune hacia la cronicidad? La presencia de: Antígenos circulantes Persistencia de la estimulación antigénica Formación de inmunocomplejos Fenómenos inmunopatológicos Inmunosupresión

46
Reacción frente al paludismo
Intervienen tanto células TH1 como TH2. Las células TH1 actúan durante la fase hepática, ya que interviene el INFg que secretan estas células, eliminando los esporozoitos. Las células TH2 que colaboran en la producción de anticuerpos permiten la eliminación de los parásitos palúdicos durante la fase eritrocitaria, efectuada en el bazo por las células efectoras activadas y citotoxicidad mediada por anticuerpos.

47
Respuesta inmune frente a la infección por leishmania
IL12 producida por macrófagos y células B promueve el crecimiento de las células TH1 activadas y de NK que también produce IFNg (citoquinas anti- leishmania). Así se suprime el desarrollo de las células TH2. Este último perfil hace progresar la enfermedad.
. Así se suprime el desarrollo de las células TH2. Este último perfil hace progresar la enfermedad.")
48
Mecanismos de destrucción T. cruzi
Respuesta protectora Mecanismos de destrucción T. cruzi NADPH NADP+ O2 O2- TNF ONOO- IFN ON IRF1 iNOS L-arginina

49
Infecciones por Helmintos
Es característico la elevación de las IgE y de los eosinófilos, lo que es dependiente de las células TH2. Las células TH1 están relacionadas con la secreción de INFg y la eliminación de las larvas. La generación de huevos por parte del helminto cambia la respuesta a TH2 (baja el INFg y sube la IL5). Si la respuesta no puede eliminar la infección, el parásito se aísla con células inflamatorias atraídas por citoquinas en la región (Granulomas).
. Si la respuesta no puede eliminar la infección, el parásito se aísla con células inflamatorias atraídas por citoquinas en la región (Granulomas).")
50
Eliminación de helmintos intestinales
Las células TH2 son esenciales en la eliminación de helmintos intestinales Se induce la generación de IL4, IL5, promoviendo la producción de IgE y la hipertrofia de mastocitos intestinales. Se generan además citoquinas IL3, GM-CSF y TNFa y proteasas. Hay aumento de la permeabilidad intestinal, de la descamación del epitelio y de la secreción de complemento y anticuerpos (hipergamaglobulinemia) hacia la luz intestinal. Se incrementa el peristaltismo.
 hacia la luz intestinal. Se incrementa el peristaltismo.")
51
Eliminación de helmintos intestinales

52
Respuestas efectoras frente a esquistosómulas

53
Mecanismo de evasión Permite que la infección parasitaria tenga éxito:
Utilizan receptores del complemento para penetrar macrófagos (leishmania). Ejemplares adultos de S. mansoni son estimulados por el TNFa para la postura de huevos. T. brucei utiliza INFg como factor de crecimiento Resistencia a efectos destructivos del complemento Activación policlonal no específica en T. cruzi.
. Ejemplares adultos de S. mansoni son estimulados por el TNFa para la postura de huevos. T. brucei utiliza INFg como factor de crecimiento. Resistencia a efectos destructivos del complemento. Activación policlonal no específica en T. cruzi.")
54
Resistencia a la lisis por complemento
Cubierta superficial de lipo-fosfoglicano (L.major): activa el complemento pero a continuación desprende el complejo y evita la lisis. Glucoproteína de superficie con actividad parecida a la del factor acelerador de la degradación (DAF) (T. cruzi)
: activa el complemento pero a continuación desprende el complejo y evita la lisis. Glucoproteína de superficie con actividad parecida a la del factor acelerador de la degradación (DAF) (T. cruzi)")
55
Los parásitos intracelulares evitan la destrucción
Ingresar por vías no fagocíticas (T. gondii) evita que se desencadene el estallido respiratorio Ingresar por unión a receptores del complemento (Leishmania). Presencia de superoxido dismutasa (leishmania) Cubiertas de lipofosfoglucanos protege frente a ataques enzimáticos. Presencia de glicoproteína (Gp63) que inhibe las enzimas de los lisosomas de los macrófagos. Reprimir expresión de las moléculas CPH de clase II en macrófagos infectados.
 evita que se desencadene el estallido respiratorio. Ingresar por unión a receptores del complemento (Leishmania). Presencia de superoxido dismutasa (leishmania) Cubiertas de lipofosfoglucanos protege frente a ataques enzimáticos. Presencia de glicoproteína (Gp63) que inhibe las enzimas de los lisosomas de los macrófagos. Reprimir expresión de las moléculas CPH de clase II en macrófagos infectados.")
56
Mecanismos desarrollados por el parásito
Variación antigénica: ejemplo tripanosomas africanos cambian una proteína de superficie (VSG: glucoproteina variable de superficie). Cada generación de parásitos es antigénicamente diferente a los anteriores. Recubrir con moléculas del huésped su superficie: Ejemplo esquistosómas pueden adquirir moléculas de superficie que contienen los determinantes A, B, H de los grupos sanguíneos, o también CPH.
. Cada generación de parásitos es antigénicamente diferente a los anteriores. Recubrir con moléculas del huésped su superficie: Ejemplo esquistosómas pueden adquirir moléculas de superficie que contienen los determinantes A, B, H de los grupos sanguíneos, o también CPH.")
57
Estrategias de defensa de parásitos extracelulares
Formación de quistes protectores, como Entamoeba histolytica, o T. spiralis. Se recubren con colágeno como O. volvulus Los nematodos poseen una gruesa cutícula exterior para protegerse de los tóxicos. Secreción de inhibidor de elastasas por parte de las tenias anulando la capacidad de atraer neutrófilos. Secreción de glutatión peroxidasa de superficie por parte de filarias Secreción de proteasas que escinden inmunoglobulinas separando el fragmento Fc.

58
Interferencia en la respuesta inmunitaria
Inmunosupresión celular y humoral. Ejemplo larvas de Trichinella spiralis que secreta un factor soluble linfocitotoxico. Secreción incrementada de polisacáridos que desbordan la capacidad de acción de los macrófagos, interfieren con las células presentadoras de antígenos en el procesamiento de los mismos. En el paludismo la hemozoína acumulada en macrófagos interfiere con la función. Filarias y tenias pueden secretar prostaglandinas capaces de suprimir la respuesta inflamatoria. Distracción inmunitaria: P. falciparum. Libera antígenos solubles que provocan activación policlonal.

59
Antígenos libres como mecanismo evasor

60
Sistema para evitar la digestión de parásitos intracelulares

61
Algunos mecanismos evasores

62
Consecuencias inmunopatológicas de las parasitosis
Hepatomegalia y esplenomegalia en respuesta al incremento de la acción de los macrófagos (Leishmania, tripanosomas, paludismo) Granulomas de células T (Esquistosomiasis) Elefantiasis (filarias) Inmunocomplejos (Paludismo) Liberación de mediadores de mastocitos por incremento en concentración de IgE. Shock anafilactico Activación policlonal: autoanticuerpos a linfocitos, eritrocitos, ADN (Tripanosomiasis, paludismo) Exceso de citoquinas causan anemia, diarrea, alteraciones pulmonares Inmunosupresión inespecífica
 Granulomas de células T (Esquistosomiasis) Elefantiasis (filarias) Inmunocomplejos (Paludismo) Liberación de mediadores de mastocitos por incremento en concentración de IgE. Shock anafilactico. Activación policlonal: autoanticuerpos a linfocitos, eritrocitos, ADN (Tripanosomiasis, paludismo) Exceso de citoquinas causan anemia, diarrea, alteraciones pulmonares. Inmunosupresión inespecífica.")
63
Aparición de Anemia en el paludismo
Se destruyen mas hematíes que los que se encuentran parasitados. Lisis por liberación de esquizonte. Inmunocomplejo unidos a hematíes Autoanticuerpos contra hematíes TNFa inhibe eritropoyesis

64
Fenómenos inmunopatológicos
Inmunosupresión: T.cruzi: El parásito inhibe la proliferación de las células T dependiente de CPAs (presentadoras) Cruzipaina induce activación alternativa de macrófagos (multiplicación de los parásitos). Autoinmunidad: En T. cruzi. Activación policlonal, o por mimetismo molecular (cruzipaina, B13, - (similar a miosina cardiaca), proteínas ribosomasles P del parásito, - (Receptores muscarínicos). Granulomas: Los huevos de S. mansoni colocados en las vénulas intestinales. Falla renal: Por inmunocomplejos circulantes Alteran función: Antígenos de Fasciola hepática alteran células dendríticas, atenúan desarrollo de respuestas Th17
 Cruzipaina induce activación alternativa de macrófagos (multiplicación de los parásitos). Autoinmunidad: En T. cruzi. Activación policlonal, o por mimetismo molecular (cruzipaina, B13, - (similar a miosina cardiaca), proteínas ribosomasles P del parásito, - (Receptores muscarínicos). Granulomas: Los huevos de S. mansoni colocados en las vénulas intestinales. Falla renal: Por inmunocomplejos circulantes. Alteran función: Antígenos de Fasciola hepática alteran células dendríticas, atenúan desarrollo de respuestas Th17.")
65
inmunodiagnóstico Enzimas: ELISA, western blot
Hemaglutinación indirecta Inmunofluorescencia indirecta Fijación del complemento Isotopos radiactivos: radioinmunoensayo Uso de citometría de flujo

66
Vacunas En veterinaria se utilizaron algunas vacunas a parásitos vivo atenuados. No existe ninguna vacuna para uso humano El principal problema es la complejidad antigénica y los mecanismos de evadir las respuestas por parte de los parásitos. También constituye un problema las distintas fases en la que podemos encontrar un parásito. No se ha podido preparar un vacuna inmunogénica Hay ensayos clínicos fase I y II para paludismo, leishmaniosis, e hidatidosis. El Instituto Oswaldo Cruz desarrollo una vacuna en humanos para esquistosomiasis que comenzó a probarse en 2012 con muy buenos resultados

67
Bibliografía Inmunología. Roitt, Brostoff, Male. Quinta edición.
Inmunología. Biología y patología del sistema inmune. Regueiro González, Larrea, González Rodriguez, Martinez Naves. Parasitosis Humanas. David Botero, Marcos Restrepo. Tercera edición. Manual Práctico de Parasitología médica. Dra. Nélida G. Saredi Parasitosis regionales. Sixto Raúl Costamagna. Elena C. Visciarelli.

Presentaciones similares





, engullendo la bacteria del carbunco (naranja).>")





 ESPECÍFICAS (Respuesta inmunitaria) – La unión antígeno anticuerpo es específica.>")








