Descargar la presentación
La descarga está en progreso. Por favor, espere

1
FISIOLOGÍA GENERAL TEMA 2. Diferenciación celular. Organización funcional del cuerpo humano. TEMA 3. Medio interno. Homeostasis. Mecanismos y sistemas de control. TEMA 4. Compartimientos del organismo. Líquidos corporales. TEMA 5. Funciones de las membranas celulares. Paso de sustancias. Mensajeros químicos. Receptores. TEMA 6. Excitabilidad. Potenciales de membrana. TEMA 7. Potencial de acción y teoría iónica del impulso nervioso. TEMA 8. Conducción del impulso nervioso y fisiología general de las fibras nerviosas. TEMA 9. Transmisión sináptica. TEMA 10. Sinapsis colinérgicas y catecolaminérgicas. Otros tipos de sinapsis. TEMA 11. Efectores. Excitación y contracción del músculo esquelético. TEMA 12. Excitación y contracción del músculo liso. Músculo cardíaco. TEMA 13. Organización funcional del sistema nervioso. Reflejos TEMA 14. Sistema nervioso autónomo.

2
Tema 8. Conducción del impulso nervioso y fisiología general de las fibras nerviosas.
Introducción. Morfología de la neurona. Tipos de células del sistema nervioso. Conducción del impulso nervioso. Tipos de fibras nerviosas.

3
1. Introducción → Ramón y Cajal: teoría neuronal de organización del SN. → La unidad anatómico funcional del SN es la neurona. → Las ramificaciones neuronales se entrelazan entre sí. → Los puntos de contacto entre neuronas son las sinapsis. La compleja estructura del sistema nervioso hizo que la identificación microscópica y demostración de tales elementos celulares fuera compleja y objeto de numerosas polémicas. De hecho, inicialmente la opinión científica se inclinaba por la conocida teoría reticularista, la cual suponía que las prolongaciones de las neuronas se fundían formando una red en la que no existía la identidad celular individual.Tanto el enunciado como las implicaciones conceptuales de la teoría celular no se aplicaron al estudio del tejido nervioso hasta más de medio siglo después de su enunciación. La teoría reticularista fue desechada gracias a los estudios histológicos de Cajal, quien aportó argumentos esenciales para el establecimiento de la teoría neuronal y considerar así a las neuronas como células individuales. Pero no fue hasta mediados del siglo XX, con el desarrollo de la microscopía electrónica, cuando se pudieron observar la membrana plasmática y los contactos sinápticos entre las neuronas, confirmando plenamente los postulados sobre la unidad anatómica de las neuronas de Cajal.

5
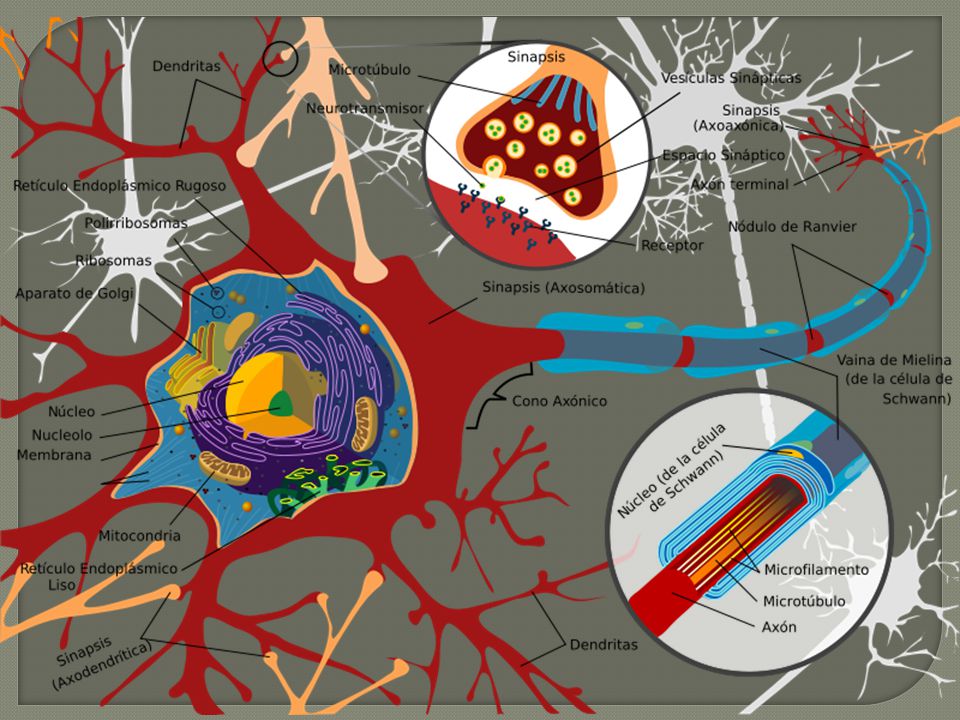
2. Morfología de la neurona
Las neuronas presentan grandes diferencias en su morfología. Dendritas (árbol dendrítico): - Prolongaciones cortas MP ricas en receptores Actúan como una antena que detecta cambios en el entorno neuronal - Sinapsis con los axones de otras neuronas Soma: cuerpo celular central. El núcleo posee una elevada actividad transcripcional. Axón: prolongación larga que parte del cono axónico, desde el que se aleja el impulso nervioso. - Isodiamétrico (0,5-20 mm) - Longitud variable (hasta 1m). - Termina en ramificaciones (telodendrón) que contiene los terminales o botones sinápticos que contactan con otras neuronas - El citoesqueleto permite el tránsito bidireccional de orgánulos (mitocondrias) y vesículas de neurotransmisores Árbol dendrítico Soma Axón Núcleo Cono axónico A diferencia con el resto de tejidos, las neuronas muestran una tremenda disparidad en su morfología. Gracias a la diversidad morfológica y funcional entre las neuronas es posible la compleja actividad del sistema nervioso y de los epifenómenos derivados como la memoria y el aprendizaje. A pesar de esta enorme heterogeneidad existe, a partir de una serie de características comunes, el concepto generalizado de neurona como una célula constituida por: las dendritas (árbol dendrítico), el soma (cuerpo celular) y el axón (árbol axónico). Evidentemente existen excepciones que no se ajustan completamente a esta constitución general. Por ejemplo existen neuronas sin dendritas o reducidas a una mínima expresión como en las neuronas ganglionares, otras carecen de axón como las células amacrinas de retina y bulbo olfatorio incluso pueden carecer de dendritas y axón, como las células cromafines adrenales. La transmisión nerviosa ocurre en dirección desde las dendritas hasta el axón. Las dendritas se extienden desde el soma neuronal como ramas a partir del tronco de un árbol, de ahí que al conjunto de dendritas de una neurona individual se le denomine árbol dendrítico. Así pues las dendritas actúan como una gran antena receptora de los acontecimientos que ocurren en el entorno neural, de hecho tienen la función especializada de la recepción de contactos sinápticos. El árbol dendrítico es la zona de recepción de información procedente de los axones de otras neuronas, aunque no son las únicas zonas de recepción, de hecho también existen espinas somáticas y axónicas (a nivel del cono axónico). En el tema siguiente se realizará un estudio pormenorizado de sobre la estructura y función de las dendritas en contacto sináptico. Soma o cuerpo neuronal Usualmente ocupa la porción central de la célula. Contiene un gran núcleo en estrecha colaboración con los procesos citoplasmáticos y con una elevada actividad transcripcional. Esto queda patente en la cromatina distendida, un nucleolo hipertrófico y la elevada concentración de poros nucleares que presenta. La elevada actividad en la traducción y síntesis proteica se manifiesta en los abundantes polirribosomas y áreas de retículo endoplasmático rugoso, alrededor del núcleo, conocido como grumos de Nissl. Axón Tiene su origen en el cono axónico, pegado al soma neuronal, región importante en el desencadenamiento del potencial de acción. A partir de él, el impulso nervioso se aleja del cuerpo celular. Normalmente es único, largo y delgado. Su diámetro, aunque puede variar de menos de una micra a muchas micras, es isodiamétrico (mantiene su diámetro en su extensión). Su longitud también es variable, llegando a alcanzar más de un metro en algunas neuronas motoras. Puede presentar alguna ramificación, las cuales se disponen en ángulo recto y en su extremo terminal presentan una profusa ramificación denominada telodendrón o arborización terminal. En el telodendrón se localizan una dilataciones denominadas botones terminales o botones sinápticos, lugar donde se produce el contacto sináptico con otras neuronas. Al seccionar un axón se observa que en su axoplasma (citoplasma axónico) apenas existe maquinaria de síntesis proteica (casi no existen proteínas), sin embargo abundan los neurotúbulos (microtúbulos) y neurofilamentos, los cuales se encuentran distribuidos en sentido longitudinal del axón. Estos componentes estructurales del citoesqueleto son muy útiles en el transporte axonal bidireccional
: - Prolongaciones cortas. MP ricas en receptores. Actúan como una antena que detecta cambios en el entorno neuronal. - Sinapsis con los axones de otras neuronas. Soma: cuerpo celular central. El núcleo posee una elevada actividad transcripcional. Axón: prolongación larga que parte del cono axónico, desde el que se aleja el impulso nervioso. - Isodiamétrico (0,5-20 mm) - Longitud variable (hasta 1m). - Termina en ramificaciones (telodendrón) que contiene los terminales o botones sinápticos que contactan con otras neuronas. - El citoesqueleto permite el tránsito bidireccional de orgánulos (mitocondrias) y vesículas de neurotransmisores. Árbol dendrítico. Soma. Axón. Núcleo. Cono axónico. A diferencia con el resto de tejidos, las neuronas muestran una tremenda disparidad en su morfología. Gracias a la diversidad morfológica y funcional entre las neuronas es posible la compleja actividad del sistema nervioso y de los epifenómenos derivados como la memoria y el aprendizaje. A pesar de esta enorme heterogeneidad existe, a partir de una serie de características comunes, el concepto generalizado de neurona como una célula constituida por: las dendritas (árbol dendrítico), el soma (cuerpo celular) y el axón (árbol axónico). Evidentemente existen excepciones que no se ajustan completamente a esta constitución general. Por ejemplo existen neuronas sin dendritas o reducidas a una mínima expresión como en las neuronas ganglionares, otras carecen de axón como las células amacrinas de retina y bulbo olfatorio incluso pueden carecer de dendritas y axón, como las células cromafines adrenales. La transmisión nerviosa ocurre en dirección desde las dendritas hasta el axón. Las dendritas se extienden desde el soma neuronal como ramas a partir del tronco de un árbol, de ahí que al conjunto de dendritas de una neurona individual se le denomine árbol dendrítico. Así pues las dendritas actúan como una gran antena receptora de los acontecimientos que ocurren en el entorno neural, de hecho tienen la función especializada de la recepción de contactos sinápticos. El árbol dendrítico es la zona de recepción de información procedente de los axones de otras neuronas, aunque no son las únicas zonas de recepción, de hecho también existen espinas somáticas y axónicas (a nivel del cono axónico). En el tema siguiente se realizará un estudio pormenorizado de sobre la estructura y función de las dendritas en contacto sináptico. Soma o cuerpo neuronal. Usualmente ocupa la porción central de la célula. Contiene un gran núcleo en estrecha colaboración con los procesos citoplasmáticos y con una elevada actividad transcripcional. Esto queda patente en la cromatina distendida, un nucleolo hipertrófico y la elevada concentración de poros nucleares que presenta. La elevada actividad en la traducción y síntesis proteica se manifiesta en los abundantes polirribosomas y áreas de retículo endoplasmático rugoso, alrededor del núcleo, conocido como grumos de Nissl. Axón. Tiene su origen en el cono axónico, pegado al soma neuronal, región importante en el desencadenamiento del potencial de acción. A partir de él, el impulso nervioso se aleja del cuerpo celular. Normalmente es único, largo y delgado. Su diámetro, aunque puede variar de menos de una micra a muchas micras, es isodiamétrico (mantiene su diámetro en su extensión). Su longitud también es variable, llegando a alcanzar más de un metro en algunas neuronas motoras. Puede presentar alguna ramificación, las cuales se disponen en ángulo recto y en su extremo terminal presentan una profusa ramificación denominada telodendrón o arborización terminal. En el telodendrón se localizan una dilataciones denominadas botones terminales o botones sinápticos, lugar donde se produce el contacto sináptico con otras neuronas. Al seccionar un axón se observa que en su axoplasma (citoplasma axónico) apenas existe maquinaria de síntesis proteica (casi no existen proteínas), sin embargo abundan los neurotúbulos (microtúbulos) y neurofilamentos, los cuales se encuentran distribuidos en sentido longitudinal del axón. Estos componentes estructurales del citoesqueleto son muy útiles en el transporte axonal bidireccional.")
6
3. Generación del potencial de acción
Las dendritas actúan como antena receptoras de señales: sinapsis de otras neuronas. Estas señales pueden ser activadoras o inhibidoras (siguiente tema). La despolarización se transmite a través del soma hasta el cono axónico. Si la despolarización llega a un cierto umbral, se dispara un potencial de acción que se transmite por todo el axón hasta los contactos sinápticos
. La despolarización se transmite a través del soma hasta el cono axónico. Si la despolarización llega a un cierto umbral, se dispara un potencial de acción que se transmite por todo el axón hasta los contactos sinápticos.")
7
4. Conducción del potencial de acción
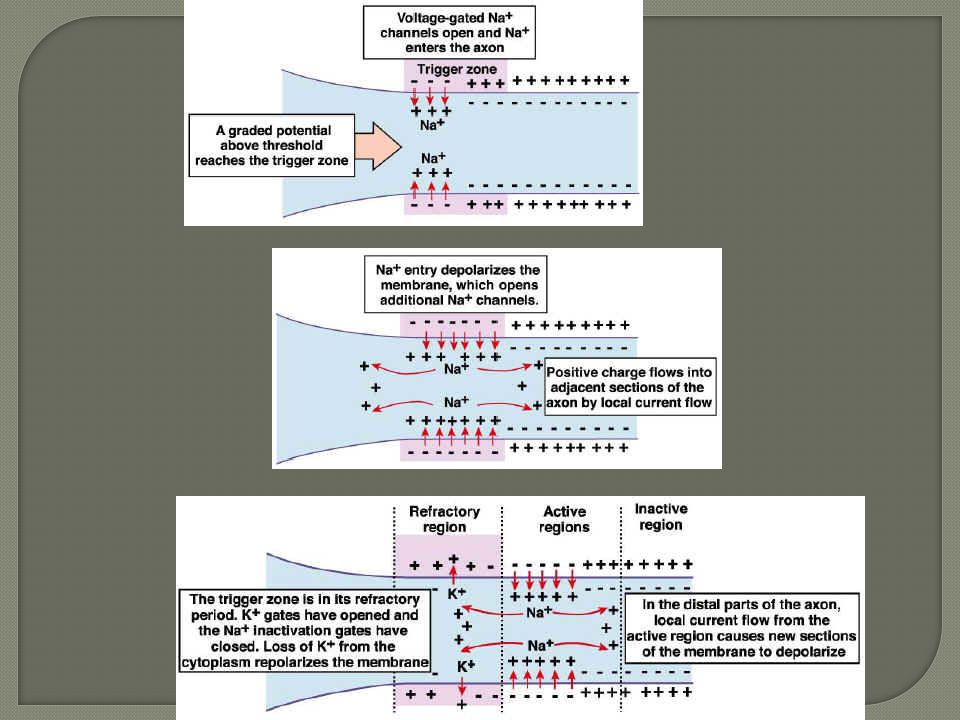
El potencial de acción es conducido a lo largo de las fibras nerviosas (axones) sin reducir su intensidad. La forma y la amplitud del potencial de acción es siempre la misma: para codificar información se utilizan variaciones de frecuencia. La frecuencia máx. Está limitada por la duración del PRA (1 ms): 1000 impulsos por segundo. La conducción se lleva a cabo por corrientes de circuitos locales: las regiones adyacentes a la zona despolarizada se acercan al umbral de disparo y desencadenan potenciales de acción (conducción electrotónica).
 sin reducir su intensidad. La forma y la amplitud del potencial de acción es siempre la misma: para codificar información se utilizan variaciones de frecuencia. La frecuencia máx. Está limitada por la duración del PRA (1 ms): 1000 impulsos por segundo. La conducción se lleva a cabo por corrientes de circuitos locales: las regiones adyacentes a la zona despolarizada se acercan al umbral de disparo y desencadenan potenciales de acción (conducción electrotónica).")
9
4. Conducción del potencial de acción
La velocidad de conducción electrotónica depende de las propiedades eléctricas del citoplasma y de la MP. A mayor diámetro de la fibra, mayor velocidad de conducción electrotónica… … a menor resistencia interna del axón, menor será la caída electrotónica con la distancia Una fibra nerviosa de 10 micras conduciría a 0,5 m/s: 4 segundos para retirar el pie!!!

10
4. Conducción del potencial de acción
Los axones están recubiertos de mielina La cubierta de mielina aisla electricamente el axón, aumentando la resistencia eléctrica de la membrana: - Menor pérdida de señal conducida - Mayor velocidad de conducción Los intercambios de iones ocurren en los nódulos de Ranvier.

11
5. Conducción saltatoria del potencial de acción
El recubrimiento de mielina es llevado a cabo por los oligodendrocitos en el SNC y las células de Schwann en el SNP a intervalos regulares (entre 1 y 3mm).
.")
12
5. Conducción saltatoria del potencial de acción
El potencial de acción “salta” de un nódulo de Ranvier a otro. Entre ellos la corriente sufre conducción electrotónica. En la MP del nódulo es donde hay canales de Na+ y K+ y bomba Na+/K+ La conducción saltatoria permite mayor velocidad de conducción 100 veces mayor, y con menor movimiento de iones y menor gasto energético Debajo de la vaina de mielina la corriente fluye longitudinalmente por el axolema (no hay conducción activa)hasta el siguiente nodo donde la apertura de los canales Na+ permitirá la propagación del potencial de acción. De forma que el potencial recobra su amplitud y sigue viajando pasivamente hasta el próximo nodo. Estos saltos de potencial de acción de un nodo al siguiente se denominan conducción saltatoria. Este mecanismo es más rápido que el hallado en las fibras amielínicas (120 m/s en comparación con 0,5 m/s). Ha desempeñado un papel importante en la evolución de organismos mayores y más complejos cuyos sistemas nerviosos necesitan transmitir rápidamente potenciales de acción a largas distancias. Sin conducción saltatoria, la velocidad de conducción requeriría incrementos drásticos en el diámetro del axón, a tal punto que podrían resultar en la formación de sistemas nerviosos excesivamente grandes para los cuerpos que deben alojarlos.
hasta el siguiente nodo donde la apertura de los canales Na+ permitirá la propagación del potencial de acción. De forma que el potencial recobra su amplitud y sigue viajando pasivamente hasta el próximo nodo. Estos saltos de potencial de acción de un nodo al siguiente se denominan conducción saltatoria. Este mecanismo es más rápido que el hallado en las fibras amielínicas (120 m/s en comparación con 0,5 m/s). Ha desempeñado un papel importante en la evolución de organismos mayores y más complejos cuyos sistemas nerviosos necesitan transmitir rápidamente potenciales de acción a largas distancias. Sin conducción saltatoria, la velocidad de conducción requeriría incrementos drásticos en el diámetro del axón, a tal punto que podrían resultar en la formación de sistemas nerviosos excesivamente grandes para los cuerpos que deben alojarlos.")
13
FACTORES QUE CONDICIONAN LA VELOCIDAD DE CONDUCCIÓN
5. Conducción saltatoria del potencial de acción Temperatura Edad FACTORES QUE CONDICIONAN LA VELOCIDAD DE CONDUCCIÓN Características propias de la fibra: Presencia o ausencia de mielina Diámetro Tres factores influyen directamente en la velocidad de transmisión del impulso nervioso: 1.- Diámetro de la neurona. 2.- Espesor de la capa mielínica. 3.- Temperatura. A mayor diámetro, espesor o temperatura mayor es la velocidad de conducción. En general, la conducción nerviosa es rápida pero bastante más lenta que la electricidad. Propagación En los axones amielínicos, los potenciales de acción se propagan como una interacción pasiva entre la despolarización que se desplaza por la membrana y los canales de sodio regulados por voltaje. Los potenciales de acción de membrana pueden representarse uniendo varios circuitos RC, cada uno representando un trozo de membrana. Cuando una parte de la membrana celular se despolariza lo suficiente como para que se abran los canales de sodio dependientes de voltaje, los iones de sodio entran en la célula por difusión facilitada. Una vez dentro, los iones positivos de sodio impulsan los iones próximos a lo largo del axón por repulsión electrostática, y atraen los iones negativos desde la membrana adyacente. Como resultado, una corriente positiva se desplaza a lo largo del axón, sin que ningún ion se esté desplazando muy rápido. Una vez que la membrana adyacente está sufiencientemente despolarizada, sus canales de sodio dependientes de voltaje se abren, realimentando el ciclo. El proceso se repite a lo largo del axón, generándose un nuevo potencial de acción en cada segmento de la membrana. [editar] Velocidad de propagación Los potenciales de acción se propagan más rápido en axones de mayor diámetro, si los demás parámetros se mantienen. La principal razón para que ocurra es que la resistencia axial de la luz del axón es menor cuanto mayor sea el diámetro, debido a la mayor relación entre superficie total y superficie de membrana en un corte transversal. Como la superficie de la membrana es el obstáculo principal para la propagación del potencial en axones amielínicos, el incremento de esta tasa es una forma especialmente efectiva de incrementar la velocidad de la transmisión. Un ejemplo extremo de un animal que utiliza el aumento de diámetro de axón como regulador de la velocidad de propagación del potencial de membrana es el calamar gigante. El axón del calamar gigante controla la contracción muscular asociada con la respuesta de evasión de depredadores del animal. Este axón puede sobrepasar 1 mm de diámetro, y posiblemente sea una adaptación para permitir una activación muy rápida del mecanismo de escape. La velocidad de los impulsos nerviosos en estas fibras es una de las más rápidas de la naturaleza. Mecanismo detallado El principal obstáculo para la velocidad de transmisión en axones amielínicos es la capacitancia de la membrana. La capacidad de un condensador puede disminuirse bajando el área de un corte transversal de sus placas, o incrementando la distancia entre las placas. El sistema nervioso utiliza la mielinización para reducir la capacitancia de la membrana. La mielina es una vaina protectora creada alrededor de los axones por las células de Schwann y los oligodendrocitos, células de la neuroglía que aplastan sus citoplasmas formando láminas de membrana y plasma. Estas láminas se arrollan en el axón, alejando las placas conductoras (el plasma intra y extracelular) entre sí, disminuyendo la capacitancia de la membrana. El aislamiento resultante redunda en un conducción rápida (prácticamente instantánea) de los iones a través de las secciones mielinizadas del axón, pero impide la generación de potenciales de acción en estos segmentos. Los potenciales de acción sólo se vuelven a producir en los nodos de Ranvier desmielinizados, que se sitúan entre los segmentos mielinizados. En estos anillos hay un gran número de canales de sodio dependientes de voltaje (hasta cuatro órdenes de magnitud superior a la densidad de axones amielínicos), que permiten que los potenciales de acción se regeneren de forma eficaz en ellos. Debido a la mielinización, los segmentos aislados del axón actúan como un cable pasivo: conducen los potenciales de acción rápidamente porque la capacitancia de la membrana es muy baja, y minimizan la degradación de los potenciales de acción porque la resistencia de la membrana es alta. Cuando esta señal que se propaga de forma pasiva alcanza un nodo de Ranvier, inicia un potencial de acción que viaja de nuevo de forma pasiva hasta que alcanza el siguiente nodo, repitiendo el ciclo.
 entre sí, disminuyendo la capacitancia de la membrana. El aislamiento resultante redunda en un conducción rápida (prácticamente instantánea) de los iones a través de las secciones mielinizadas del axón, pero impide la generación de potenciales de acción en estos segmentos. Los potenciales de acción sólo se vuelven a producir en los nodos de Ranvier desmielinizados, que se sitúan entre los segmentos mielinizados. En estos anillos hay un gran número de canales de sodio dependientes de voltaje (hasta cuatro órdenes de magnitud superior a la densidad de axones amielínicos), que permiten que los potenciales de acción se regeneren de forma eficaz en ellos. Debido a la mielinización, los segmentos aislados del axón actúan como un cable pasivo: conducen los potenciales de acción rápidamente porque la capacitancia de la membrana es muy baja, y minimizan la degradación de los potenciales de acción porque la resistencia de la membrana es alta. Cuando esta señal que se propaga de forma pasiva alcanza un nodo de Ranvier, inicia un potencial de acción que viaja de nuevo de forma pasiva hasta que alcanza el siguiente nodo, repitiendo el ciclo.")
14
CLASIFICACIÓN DE LAS FIBRAS NERVIOSAS
La clasificación de las fibras nerviosas se puede realizar en función de diferentes criterios. Una de las clasificaciones más generalizadas es la presentada en esta diapositiva. En primer lugar se muestra una clasificación general en la que se dividen en mielínicas y amielínicas. Siendo las fibras tipo A las que poseen envoltura mielínica y las tipo C las que no la poseen. A su vez las tipo A se subdividen en función de su diámetro, y por tanto en función de su velocidad de conducción en alpha, beta, gamma, delta. A su vez los nervios sensoriales tipo A se pueden clasificar como I, II, III y los tipo C como IV. Guyton (McGraw Hill, 1992)
")
Presentaciones similares