Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Aspecto molecular en la Enfermedad del Parkinson.
Dr. Alex Espinoza Giacomozzi. Neurología. Hospital DIPRECA. Dr Chana.

4
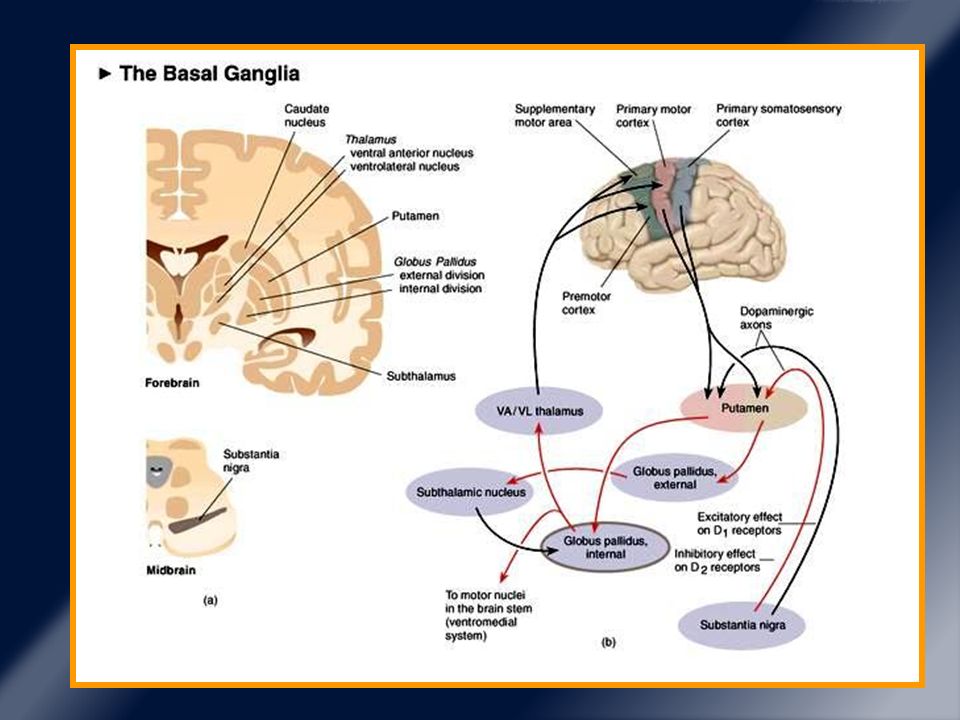
Neuronas espinudas medias del putamen.

6
Estimulación no fisiológica del receptor de dopamina.
El sistema dopaminérgico nigroestriatal opera tónicamente. La actividad en estas neuronas es de una frecuencia modal 4 Hz, excepto cuando son interrumpidas brevemente por disparos fásicos bursts (en primates) Schultz W, et al, Rev Neurol 1994, 150: 634 – 9.
 Schultz W, et al, Rev Neurol 1994, 150: 634 – 9.")
7
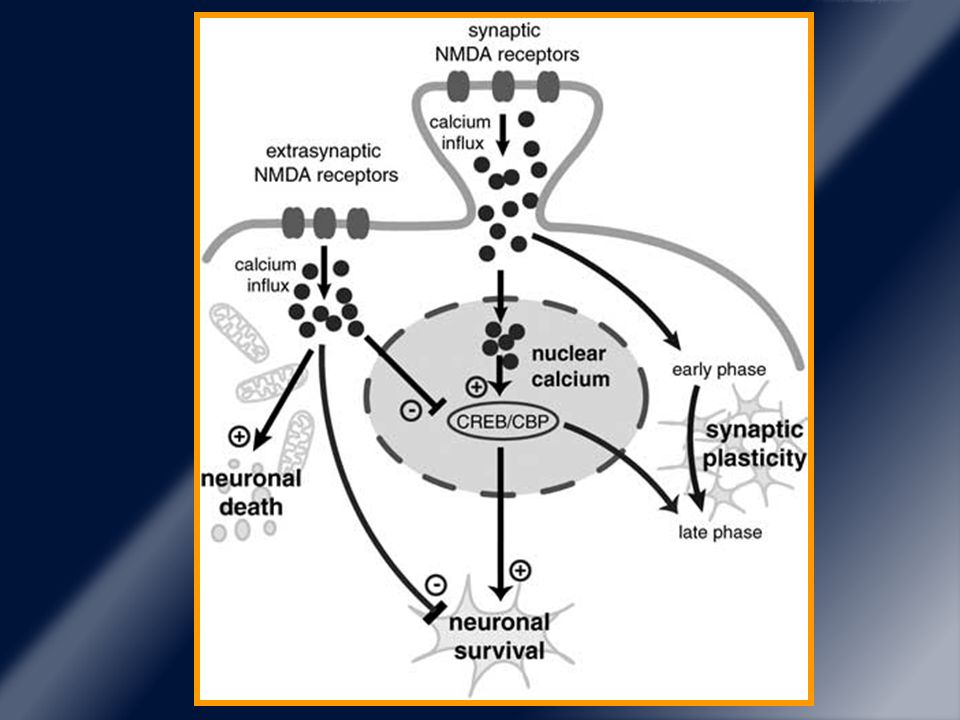
La mayoría de los receptores dopaminérgicos son extrasinápticos.
La cantidad de transmisor liberado refleja la frecuencia de descarga el receptor DA intrasináptico es expuesto a concentraciones estables de dopamina. La mayoría de los receptores dopaminérgicos son extrasinápticos. Yung et al, Neuroscience 1995, 65: 709 – 30. Caillé et al, Brain Res 1996, 730: 17 – 31. Y son activados por difusión de la DA desde la hendidura sináptica al espacio extracelular. Zoli et al, Brain Res 1998, 26:136 – 47.

8
Normalmente las concentraciones de DA permanecen dentro de un rango bajo y estable.
Skirboll et al, Exp Neurol 1990, 110: 187 – 93. Sarre et al, Arch Pharmacol 1994, 350: 15 – 21. Esta situación cambia en EP cuando los receptores de DA entran en contacto con dosis y patrones de estimulación no fisiológicas . Juncos et al, Ann Neurol 1989, 25: 473 – 8. Mouradian et al, Ann neurol 1989, 25: 523 – 6. Lo primero que ocurre es la degeneración de neuronas dopaminérgicas y caída de los niveles de DA (apareciendo la clínica).
.")
9
Luego se agrega la terapia con DA.
Al quedar pocas neuronas dopaminérgicas, las dosis estándares someten a los receptores postsinápticos a períodos intermitentes de alta estimulación. La alta intensidad está demostrada por la excesiva DA y su metabolito en LCR con terapia convencional La intermitencia es la consecuencia de la pérdida neuronal, la cual suprime los mecanismos de almacenaje vesicular de DA.

10
La alta intensidad de estimulación no es más fisiológica que la estimulación subumbral atendiendo a la denervación dopaminérgica. Por lo que la transmisión dopaminérgica se mantiene no fisiológica se trate o no el paciente. Cualquier tipo de estimulación anormal en los receptores DA puede ser capaz de alterar la función de las neuronas espinudas contribuyendo al severo aumento de wearing – off fluctuations y diskinesias de peak de dosis.

11
Neuronas espinudas medias estriatales.
Tanto la estructura como la función de estas neuronas se altera con la exposición crónica de sus receptores de DA a estimulación no fisiológica. En modelos animales los efectos funcionales incluyen cambios en la DA receptor – linked signaling afecta el estado de fosforilación eficacia sináptica de receptores glutamatérgicos ionotrópicos

12
La administración de 6 – OHDA en ratas produce EP, y la terapia con DA crónica dada intermitentemente produce en pocas semanas alteraciones motoras tipo on – off fluctuaciones. En aquellas que recibieron en forma continua no desarrollaron estas manifestaciones motoras. La denervación dopaminérgica y dosis intermitentes de DA, produce de la sensibilidad de AMPA y NMDA.

13
Resultado, la estimulación glutamatérgica en estas neuronas hace que aumente la metabolización de GABA sintetasa. Aumenta encefalina, dinorfina y neurotensina, en estas condiciones, pero no en las estimuladas continuamente. Observaciones neurofisiológicas y neurofarmacológicas indican que las modificaciones de transmisor reflejan un en la tasas de descargas de las neuronas espinudas medias.

14
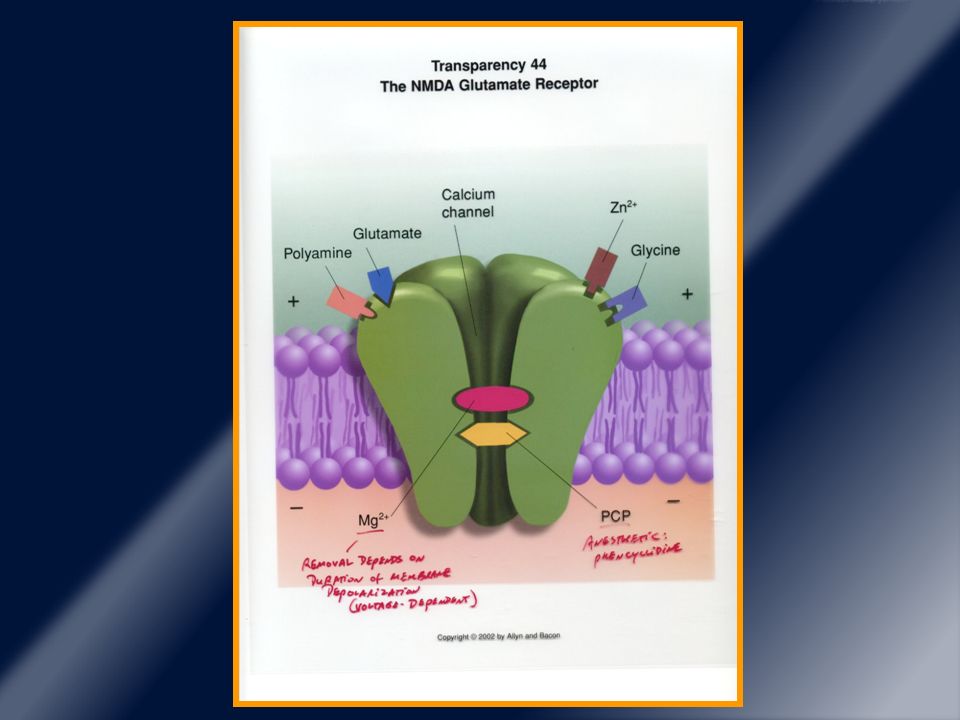
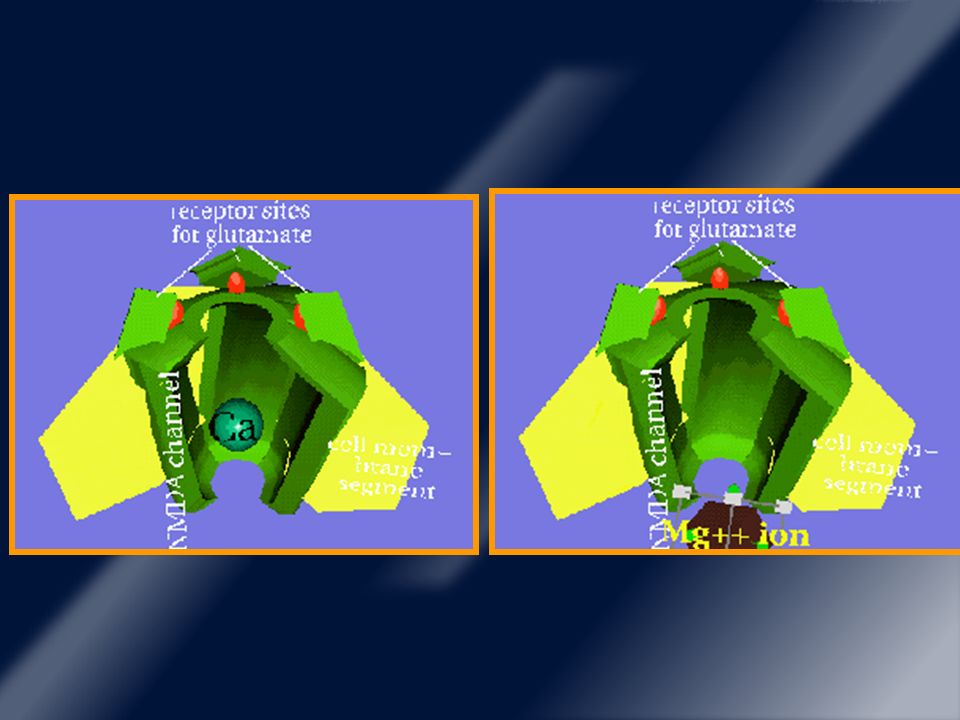
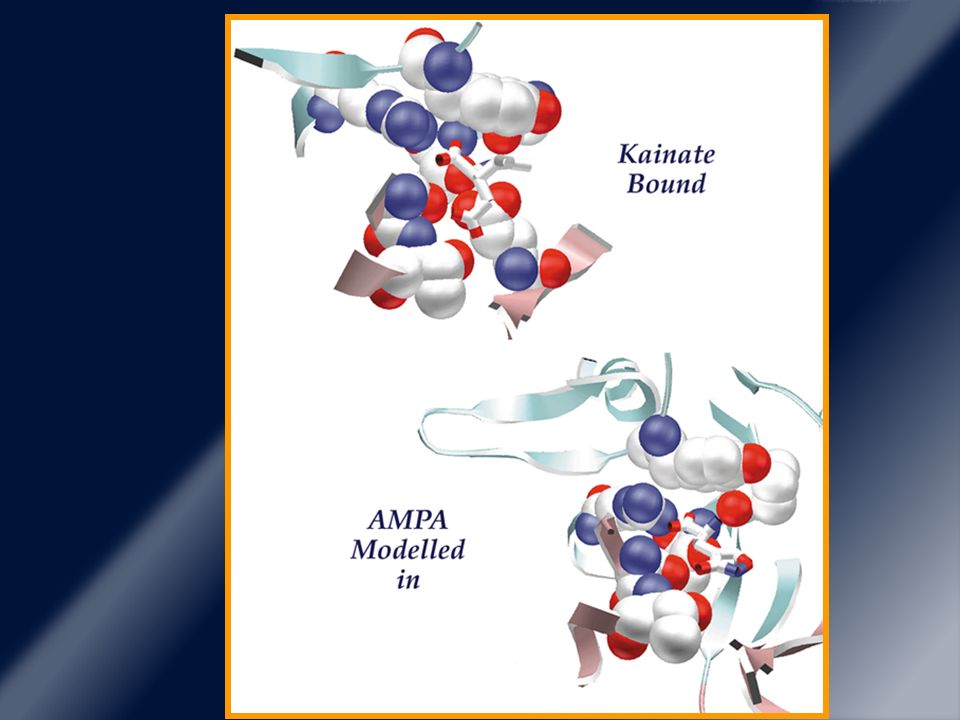
Receptores glutamatérgicos ionotrópicos.
Este receptor NMDA funciona ligado a un canal iónico (Ca2+). AMPA (canales de Na+ y otros iones monovalentes). NMDA es un tetrámero, con subunidad NR1 y NR2 y las isoformas NR2B y menos frecuente NR2A están en estriado.
. AMPA (canales de Na+ y otros iones monovalentes). NMDA es un tetrámero, con subunidad NR1 y NR2 y las isoformas NR2B y menos frecuente NR2A están en estriado.")
15
AMPA es un tetrámero con subunidad GluR 1 – 4
AMPA es un tetrámero con subunidad GluR 1 – 4. El subtipo 4 es más frecuente en neuronas espinudas medias. La regulación de estos ocurre por fosforilación proteica (sitios carboxi intracell). Fosforilación de tirosina apertura. Fosforilación de serina/treonina cierre. Alteración en la fosforilación NMDA y AMPA + denervación DA + estimulación intermitente activación de cascada intracell que expresión de receptores DA y glutamina.
. Fosforilación de tirosina apertura. Fosforilación de serina/treonina cierre. Alteración en la fosforilación NMDA y AMPA + denervación DA + estimulación intermitente activación de cascada intracell que expresión de receptores DA y glutamina.")
16
Cambios en la fosforilación de receptor.
Tirosina. Como resultado de la denervación DA + drogas DA intermitentes fosforilación de residuos de tirosina en receptores glutamatérgicos. NMDA NR2A y NR2B, no NR1, son directamente fosforilados por tirosin kinasa. Existiría relación entre fosforilación tirosina de receptores NMDA y regulación dopaminérgica en la regulación motora.

17
Aplicación de 6 – OHDA + inhibidor de tirosin kinasa produjo menores efectos motores.
En rata modelos tratadas con levodopa intermitentemente + inhibidor de tirosin kinasa, normalizó las respuestas motoras y se atenuó la fosforilación de NR2A y 2B. Hiperfosforilación de residuos de tirosina en NMDA, debido a activación de kinasa o inhibición de fosfatasa contribuye a la disfunción motora por denervación DA y TTO intermitente

18
Tirosin kinasa.

19
Serina. Fosforilación de residuos serina/treonina es afectado tanto por denervación DA, como por TTO crónico de dopaminérgicos. Esta fosforilación ocurriría en NR2A, pero no en NR2B. TTO crónico agonista D fosforilación serina en NR2A y NR2B. TTO crónico agonista D fosforilación NR2B y no afecta 2A.

20
Efecto de inhibidor de Kinasa.
3 de los pples 2dos mensajeros (kinasa PKA, PKC y CaMKII) están implicados en la alteración de la fosforilación de serina en NMDA y AMPA en la estimulación no fisiológica de receptores DA. Inhibición de PKA y CaMKII en neuronas espinudas medias de ratas modelos, limitó la eficacia antiparkinsoniana, pero redujo las alteraciones motoras por la terapia con levodopa. Inhibición de fosfatasa 2A produce efectos inversos.
 están implicados en la alteración de la fosforilación de serina en NMDA y AMPA en la estimulación no fisiológica de receptores DA. Inhibición de PKA y CaMKII en neuronas espinudas medias de ratas modelos, limitó la eficacia antiparkinsoniana, pero redujo las alteraciones motoras por la terapia con levodopa. Inhibición de fosfatasa 2A produce efectos inversos.")
21
Lo anterior apoya la idea de que la denervación DA y la estimulación intermitente de receptores DA en neuronas espinudas medias activan cascadas internas que afectan el estado de fosforilación de receptores glutamatérgicos.

22
El resultante cambio en los input corticales glutamatérgicos a las neuronas espinudas medias modifica el output de éstas lo que favorece la clínica parkinsoniana y las alteraciones motoras por el TTO a largo plazo.

23
Sensibilización del receptor glutamatérgico.
Estimulación no fisiológica en los receptores dopaminérgicos estriatales altera la eficacia de los receptores glutamatérgicos como consecuencia de los cambios en su fosforilación. La sensibilidad de los AMPA y NMDA se altera con la denervación DA y la manipulación farcológica.

24
NMDA RECEPTOR. Este receptor se expresa ppalmente en las dendritas de las neuronas espinudas medias estriatales. Antagonismo de NMDA receptor efecto antiparkinsoniano y alteraciones inducidas por levodopa. Este antagonismo tiene actividad preferencial sobre D2 (vía indirecta), por ej, amantadina (antagonista no conpetitivo NMDA) tiene limitada acción antiparkinsoniana (excepto en etapas tempranas), pero gran de diskinesias por levodopa.
, por ej, amantadina (antagonista no conpetitivo NMDA) tiene limitada acción antiparkinsoniana (excepto en etapas tempranas), pero gran de diskinesias por levodopa.")
25
Sensibilización de NMDA:
La inhibición selectiva en NR2B es la que las diskinesias por levodopa. Antagonismo selectivo NR2A potencia respuesta antiparkinsoniana, pero diskinesias inducidas por levodopa. Sensibilización de NMDA: fosforilación de tirosina de subunidad NR2B junto con fosforilación en serina de NR2A en D2 receptores es crucial favorece aparición de parkinsonismo y alteraciones motoras (diskinesias y fluctuación). La efectividad de antagonistas NMDA no selectivos, se explica por que NR2A se encuentra en varias zonas cerebrales.
. La efectividad de antagonistas NMDA no selectivos, se explica por que NR2A se encuentra en varias zonas cerebrales.")
31
AMPA receptores. Estudios en ratas modelos sugieren que antagonistas de este receptor parkinsonismo y LID. Algunos antagonistas (NBQX) potencian acción antiparkinsoniana D1 y no actúan sobre D2. Acción viceversa tiene el antagonista AMPA LY Por ende, la capacidad de estos antagonistas de denervación parkinsoniana influencia sobre output D1 nigroestriatal y su capacidad de mitigar LID relacionada con efecto sobre D2.
 potencian acción antiparkinsoniana D1 y no actúan sobre D2. Acción viceversa tiene el antagonista AMPA LY Por ende, la capacidad de estos antagonistas de denervación parkinsoniana influencia sobre output D1 nigroestriatal y su capacidad de mitigar LID relacionada con efecto sobre D2.")
32
Receptor AMPA

34
Integración sináptica.
Las dendritas de las neuronas espinudas no sólo reciben proyecciones dopaminérgicas de la sust nigra y glutamatérgica de corteza. También expresan receptores para 5 HT 2A, adenosina A2a, adrenérgicos alfa 2, canibinoides CB – 1. A través de cascadas de fosforilación – desfosforilación, estos receptores modulan los efectos de la estimulación DA no fisiológica sobre receptor glutamatérgico.

35
Receptores de adenosina A2a.
Este receptor está ampliamente en las neuronas espinudas, ppalmente cerca de D2 receptores. Actúa a través de la acción de PKA y PKC, afectando la regulación de receptor glutamatérgico ionotrópico. Las sust que bloquean este receptor tiene efecto antiparkinsoniano y especialmente anti LID. Baja la dosis de levodopa, LID. Prolonga eficacia de la X de levodopa ( fluctuaciones). Esto mediado por modificación de la señal de las kinasas, afectando la regulación del receptor glutamatérgico.
. Esto mediado por modificación de la señal de las kinasas, afectando la regulación del receptor glutamatérgico.")
36
Receptor de adenosina A2a.

37
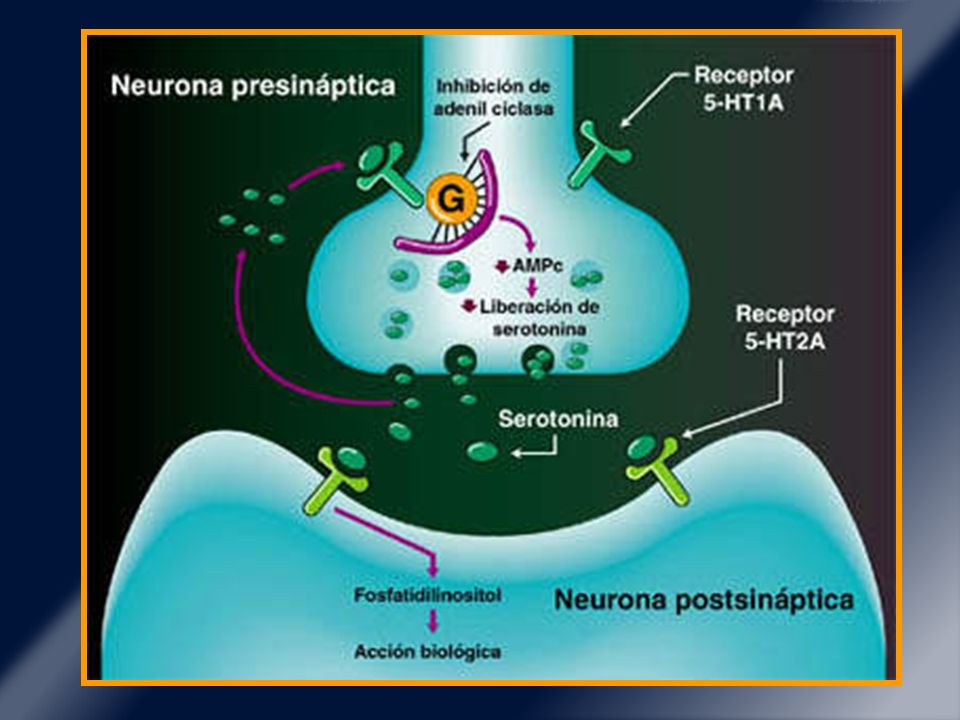
Receptor de serotonina 5HT 2A.
También ubicado en las dendritas de las neuronas espinudas medias. Su estimulación activa cascadas con AMPc y kinasas activadas por calcio (PKA y PKC). Quetiapina (actividad anti 5 HT2A/C y D2, D3) normaliza respuesta acortada a levodopa en tto crónicos y LID. fosforilación en receptor AMPA(efecto al estimular el receptor 5 HT 2A).
. Quetiapina (actividad anti 5 HT2A/C y D2, D3) normaliza respuesta acortada a levodopa en tto crónicos y LID. fosforilación en receptor AMPA(efecto al estimular el receptor 5 HT 2A).")
39
Conclusiones finales. Estudios en modelos animales y pctes con EP, sugieren que la estimulación no fisiológica de receptores de dopamina, permite cambios plásticos en neuronas espinudas medias estriatales. La aparición de EP y sus complicaciones motoras, está asociado con la activación aberrante de estas neuronas y fosforilación de sus receptores glutamatérgicos ionotrópicos NMDA y AMPA.

40
El resultado de la sensibilización de los receptores AMPA y NMDA, los inputs excitatorios corticales hacia estas neuronas, alterando el output estriatal motor.

41
La disregulación de las neuronas espinudas medias por denervación DA y estimulación no fisiológica contribuye a las alteraciones motoras.

42
Estrategias farmacológicas.
1.- Protein kinasa, cuya anómala activación por estimulación no fisiológica, modifica el estado foasforilado de receptor glutamatérgico. 2.- Receptores glutamatérgicos ionotrópicos cuya sensibilización altera el output estriatal. 3.- Cell de superficie capaces de modular cascadas inducidas por receptores DA y glutamatérgicos.

Presentaciones similares