Descargar la presentación
La descarga está en progreso. Por favor, espere

1
SEMINARIO 8 Aguilar, Rosario Bastos, Luis Marcone, María Inés Martí, María Consuelo Rudi, Juan Manuel Soria, Mariel Vaca, Alicia

2
UN NUEVO TIPO DE MUTACIÓN CAUSA UN DEFECTO EN EL SPLICING DEL GEN ATM Franco Pagani, Emanuelle Buratti, Cristiana Stuani, Regina Bendix, Thilo Dörk & Francisco E. Baralle Nature Genetics – Volumen 30 – Abril 2002

3
ATAXIA-TELANGIECTASIA

4
ATAXIA-TELANGIECTASIA

5
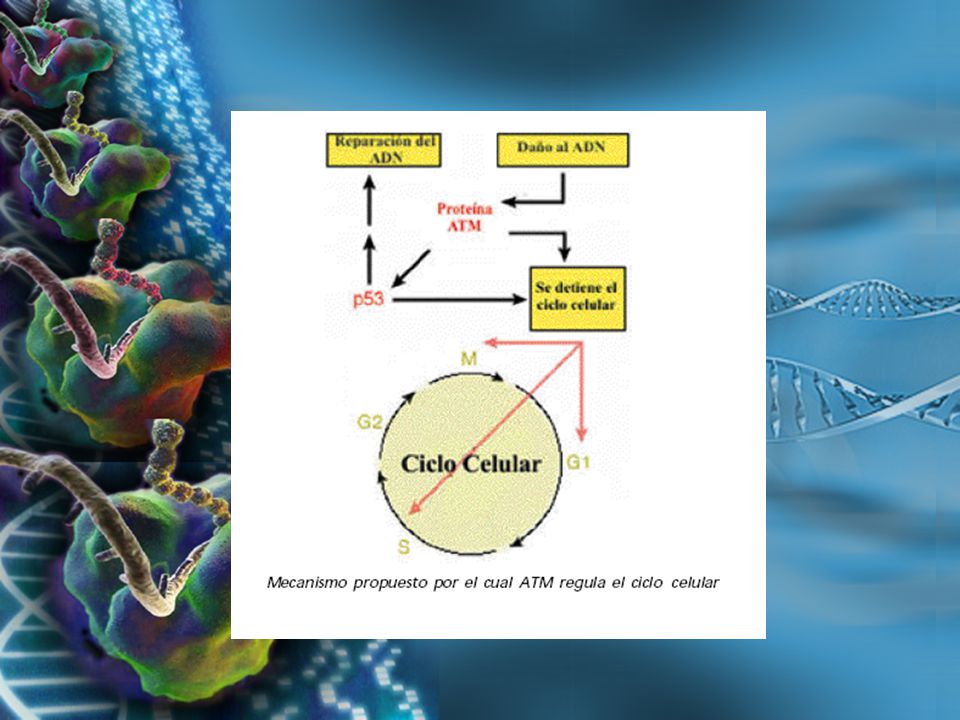
ATAXIA-TELANGIECTASIA Herencia autosómica recesiva Herencia autosómica recesiva 1/40000 – 1/100000 nacidos vivos 1/40000 – 1/100000 nacidos vivos Locus 11q 22 – 23 (1980) Locus 11q 22 – 23 (1980) Gen ATM Gen ATM Proteína Quinasa Serina Treonina Proteína Quinasa Serina Treonina Traducción de señales Traducción de señales Arresto del Ciclo Celular: G1, Fase S, G2 Arresto del Ciclo Celular: G1, Fase S, G2
 Locus 11q 22 – 23 (1980) Gen ATM Gen ATM Proteína Quinasa Serina Treonina Proteína Quinasa Serina Treonina Traducción de señales Traducción de señales Arresto del Ciclo Celular: G1, Fase S, G2 Arresto del Ciclo Celular: G1, Fase S, G2")
6
ATM

9
INTRODUCCIÓN Exon 5´ Punto de Ramificaciòn Intròn Sitio de Corte y Empalme 5´ Región rica en Pirimidina = 15b Sitio de corte y empalme 3` Exón 3` 80 80100 10060 70 60 80 80 90100 80 100 70 80 45 Pre mRNA Frecuencia de Apariciòn % Las únicas bases casi invariables son las 5´GU 3`AG 20 – 50 Kb

10
INTRODUCCIÓN Exón Críptico 20 21 DNA RNA cDNA

11
Splicing anormal Exón Críptico Deleción GTAA En el intrón 20, a 1870 y 570 pb de los exones vecinos (ex 20 y 21) Situada en una secuencia nucleotídica similar a los sitios consenso de splicing SON NECESARIOS 30 – 40 NUCLEOTIDOS, EN CADA EXTREMO DEL INTRÓN, PARA QUE EL SPLICING SEA NORMAL
 Situada en una secuencia nucleotídica similar a los sitios consenso de splicing SON NECESARIOS 30 – 40 NUCLEOTIDOS, EN CADA EXTREMO DEL INTRÓN, PARA QUE EL SPLICING SEA NORMAL")
12
HIPÓTESIS La secuencia de nucleótidos (CAGGTAAGT) del intrón 20, que contiene la deleción GTAA, pese a estar alejada de 5´ ss y de 3´ ss no es neutra en el splicing del pre-mRNA
 del intrón 20, que contiene la deleción GTAA, pese a estar alejada de 5´ ss y de 3´ ss no es neutra en el splicing del pre-mRNA")
13
Splicing entre exones 20 y 21 Evaluar el splicing entre los exones 20 y 21 en el paciente con A-T y en un individuo normal OBJETIVO

14
Splicing entre exones 20 y 21 MATERIALES Y MÉTODOS ARN (Línea celular linfoide HA174) ARN (Línea celular linfoide HA174) RT-PCR: random primer (ex 20 dir, ex 21 rev) RT-PCR: random primer (ex 20 dir, ex 21 rev) Gel de agarosa Gel de agarosa Masculino heterocigota para - Mutación salto en exón 16 ( 2250G→A ) - Mutación exón críptico entre exones 20 y 21 Masculino heterocigota para - Mutación salto en exón 16 ( 2250G→A ) - Mutación exón críptico entre exones 20 y 21 PACIENTE
 ARN (Línea celular linfoide HA174) RT-PCR: random primer (ex 20 dir, ex 21 rev) RT-PCR: random primer (ex 20 dir, ex 21 rev) Gel de agarosa Gel de agarosa Masculino heterocigota para - Mutación salto en exón 16 ( 2250G→A ) - Mutación exón críptico entre exones 20 y 21 Masculino heterocigota para - Mutación salto en exón 16 ( 2250G→A ) - Mutación exón críptico entre exones 20 y 21 PACIENTE")
15
Splicing entre exones 20 y 21 RESULTADOS Calle 1 Inserción de 65 nt (exón críptico) Calle 2 Splicing normal
 Calle 2 Splicing normal")
16
Splicing entre exones 20 y 21 Exón Críptico RNA m. 16 dir 22 rev RT PCR CDNA

17
Splicing alelo-específico Evaluar el splicing alelo-específico entre los exones 20 y 21 OBJETIVO

18
Splicing alelo-específico MATERIALES Y MÉTODOS Gel de agarosa Gel de agarosa RT-PCR: random primer (ex 16 dir, ex 22 rev) El alelo 2250G→A no podrá amplificarse (salto del exón 16) RT-PCR: random primer (ex 16 dir, ex 22 rev) El alelo 2250G→A no podrá amplificarse (salto del exón 16) ARN ARN
 El alelo 2250G→A no podrá amplificarse (salto del exón 16) RT-PCR: random primer (ex 16 dir, ex 22 rev) El alelo 2250G→A no podrá amplificarse (salto del exón 16) ARN ARN")
19
Splicing alelo-específico RESULTADOS Calle 1 Exón críptico Calle 2 Splicing normal

20
El splicing defectuoso del exón 20 no afecta el splicing regulado por elementos canónicos Observar que el splicing defectuoso del exón que posee la deleción no produce un transcripto normal OBJETIVO

21
El splicing defectuoso del exón 20 no afecta el splicing regulado por elementos canónicos MATERIALES Y MÉTODOS Los productos amplificados fueron separados en geles de agarosa Los productos amplificados fueron separados en geles de agarosa Se amplificaron por RT-PCR, usando primers específicos, los exones 19 al 22 Se amplificaron por RT-PCR, usando primers específicos, los exones 19 al 22 PUROMICINA: es un antibiótico que inhibe la síntesis proteica, pues su estructura es similar a la terminación aminoacil de un aminoacil ARNt. De esta manera libera las cadenas de pre-RNA antes de que se complete su síntesis PUROMICINA: es un antibiótico que inhibe la síntesis proteica, pues su estructura es similar a la terminación aminoacil de un aminoacil ARNt. De esta manera libera las cadenas de pre-RNA antes de que se complete su síntesis

22
RESULTADOS Calle 1 Tratamiento sin puromicina Calle 2 Tratamiento con puromicina El splicing defectuoso del exón 20 no afecta el splicing regulado por elementos canónicos

23
CONCLUSIÓN El tratamiento con puromicina incrementa la cantidad relativa del producto que contiene la inserción de 65 nt, a partir de la línea celular que se obtiene del individuo afectado. De lo contrario, se obtiene poco producto del splicing defectuoso, ya que, la inserción posee un codon stop que trunca la transcripción antes de lo normal

24
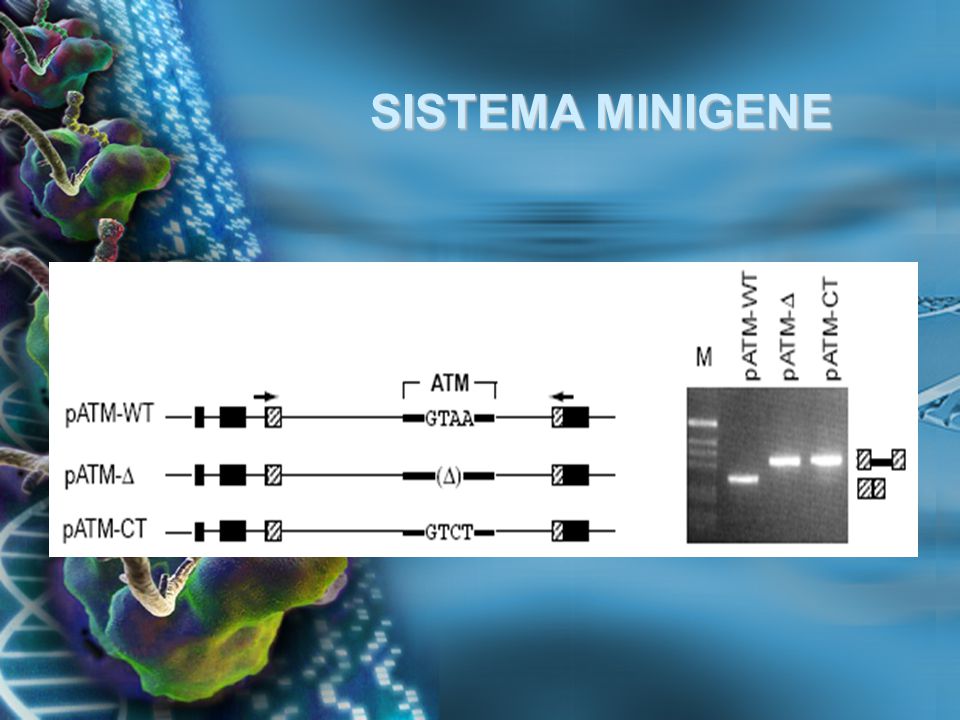
SISTEMA MINIGENE El sistema minigene es una importante herramienta para la identificación y análisis in vivo de reguladores como elementos cis- y factores trans-, que establecen un splicing eficiente y regulan el splicing alternativo

25
SISTEMA MINIGENE Los usos más comunes de los minigenes son: Determinar la variante alélica que afecta la eficiencia del splicing, entre otros usos Determinar la variante alélica que afecta la eficiencia del splicing, entre otros usos Identificar elementos exónicos e intrónicos que favorecen o reprimen el splicing Identificar elementos exónicos e intrónicos que favorecen o reprimen el splicing Determinar el rol de los sitios de splicing en establecer el nivel basal de reconociemiento del exón Determinar el rol de los sitios de splicing en establecer el nivel basal de reconociemiento del exón

26
SISTEMA MINIGENE La expresión de preRNAs mediante el Sistema Minigene, por transfección transciente, provee un rápido ensayo para conocer la pérdida de función y/o el análisis de la pérdida o ganancia de elementos cis- y/o factores trans- que afectan la regulación de la transcripción

27
SISTEMA MINIGENE

30
Estudio de la secuencia de ATM afectada por la deleción ISPE Elemento de procesamiento de splicing del intrón Secuencia del ISPE de ATM Sitios de splice 3’ y 5’ respectivamente HOMOLOGAS

31
Estudio de la secuencia de ATM afectada por la deleción Complementariedad de bases entre ISPE y snRNA U1 12 bp fuertemente complementarias Unión no clásica del snRNA U1

32
Estudio de la secuencia de ATM afectada por la deleción HIPÓTESIS La unión del snRNA U1 al ISPE es crucial en el proceso de modulación del pre RNAm del intrón 20 Mutaciones puntuales en ISPE Sistema de minigen de ATM Mutagénesis sitio dirigida RT- PCR Gel de agarosa

33
Estudio de la secuencia de ATM afectada por la deleción CONFIRMACIÓN DE LA HIPÓTESIS Mutaciones puntuales en el ISPE REMOCIÓN COMPLETA DEL INTRÓN Cierta inclusión del INTRÓN INCLUSIÓN COMPLETA DEL INTRÓN 5G y 6G excepción

34
CONCLUSIONES Secuencia intrónica (ISPE) distante de los sitios clásicos de splice Secuencia intrónica (ISPE) distante de los sitios clásicos de splice Mutaciones puntuales en la secuencia intrónica (ISPE), alteran la unión clásica de snRNA U1, actuando como el ISPE con la deleción y por lo tanto generando la inclusión del intrón Mutaciones puntuales en la secuencia intrónica (ISPE), alteran la unión clásica de snRNA U1, actuando como el ISPE con la deleción y por lo tanto generando la inclusión del intrón Activación del exon críptico producida por la de una interacción no clásica del snRNA U1 con el ISPE Activación del exon críptico producida por la de una interacción no clásica del snRNA U1 con el ISPE
 distante de los sitios clásicos de splice Secuencia intrónica (ISPE) distante de los sitios clásicos de splice Mutaciones puntuales en la secuencia intrónica (ISPE), alteran la unión clásica de snRNA U1, actuando como el ISPE con la deleción y por lo tanto generando la inclusión del intrón Mutaciones puntuales en la secuencia intrónica (ISPE), alteran la unión clásica de snRNA U1, actuando como el ISPE con la deleción y por lo tanto generando la inclusión del intrón Activación del exon críptico producida por la de una interacción no clásica del snRNA U1 con el ISPE Activación del exon críptico producida por la de una interacción no clásica del snRNA U1 con el ISPE")
35
¿La deleción de 4pb o mutación CT rompe la interación entre U1 snRNP y ATM ISPE?

36
EMSA (Electroforetic Movility Shift Assay) Puede determinar si una proteína o mezclas de proteínas, son capaces de unirse a un determinado DNA o RNA Puede determinar si una proteína o mezclas de proteínas, son capaces de unirse a un determinado DNA o RNA Gel Shift Assay, Band Shift Assay o Gel Retardation Assay Gel Shift Assay, Band Shift Assay o Gel Retardation Assay Interacción proteína-DNA o proteína-RNA Interacción proteína-DNA o proteína-RNA
 Puede determinar si una proteína o mezclas de proteínas, son capaces de unirse a un determinado DNA o RNA Puede determinar si una proteína o mezclas de proteínas, son capaces de unirse a un determinado DNA o RNA Gel Shift Assay, Band Shift Assay o Gel Retardation Assay Gel Shift Assay, Band Shift Assay o Gel Retardation Assay Interacción proteína-DNA o proteína-RNA Interacción proteína-DNA o proteína-RNA")
37
Separación electroforética de una mezcla proteína-DNA en geles de poliacrilamida o agarosa, por un período corto de tiempo Separación electroforética de una mezcla proteína-DNA en geles de poliacrilamida o agarosa, por un período corto de tiempo La velocidad de la corrida está dada por el tamaño y carga, y en menor medida por la forma La velocidad de la corrida está dada por el tamaño y carga, y en menor medida por la forma Visualización: DNA o RNA marcado con fluorescentes o radioactivos Visualización: DNA o RNA marcado con fluorescentes o radioactivos EMSA

38
EMSA Oligonucleótidos de RNA sc de ATM- ISPE normal, e ISPE con deleción (ATM-Δ) o con sustitución CT (ATM-CT) con 5’ terminal marcado Oligonucleótidos de RNA sc de ATM- ISPE normal, e ISPE con deleción (ATM-Δ) o con sustitución CT (ATM-CT) con 5’ terminal marcado Extracto nuclear de HeLA Extracto nuclear de HeLA CONCLUSIÓN: Sólo el ATM ISPE normal induce la formación del complejo Proteína-RNA específico
 o con sustitución CT (ATM-CT) con 5’ terminal marcado Oligonucleótidos de RNA sc de ATM- ISPE normal, e ISPE con deleción (ATM-Δ) o con sustitución CT (ATM-CT) con 5’ terminal marcado Extracto nuclear de HeLA Extracto nuclear de HeLA CONCLUSIÓN: Sólo el ATM ISPE normal induce la formación del complejo Proteína-RNA específico")
39
EMSA Análisis de competición Oligonucleótidos ATM-ISPE, ATM-Δ o ATM-CT, no marcados, en exceso Oligonucleótidos ATM-ISPE, ATM-Δ o ATM-CT, no marcados, en exceso ATM ISPE marcado ATM ISPE marcado Incubación: 25 minutos a T ambiente Incubación: 25 minutos a T ambiente CONCLUSIÓN: Las variantes ATM-Δ o ATM-CT no pueden desplazar al ATM ISPE del complejo proteína-RNA

40
¿Está presente U1 snRNP en el complejo Proteína-RNA específico?

41
Ensayo de clivaje con RNasa H Incubación del extracto nuclear de HeLa, con Oligonucleótido complementario U1 AS (antisense) y RNasa H Incubación del extracto nuclear de HeLa, con Oligonucleótido complementario U1 AS (antisense) y RNasa H T = 30ºC durante 15 y 30 minutos T = 30ºC durante 15 y 30 minutos Control negativo: Oligonucleótido CAS no relacionado Control negativo: Oligonucleótido CAS no relacionado
 y RNasa H Incubación del extracto nuclear de HeLa, con Oligonucleótido complementario U1 AS (antisense) y RNasa H T = 30ºC durante 15 y 30 minutos T = 30ºC durante 15 y 30 minutos Control negativo: Oligonucleótido CAS no relacionado Control negativo: Oligonucleótido CAS no relacionado")
42
Análisis de super-movilidad Se agrega anticuerpo policlonal de conejo anti U1 A Se agrega anticuerpo policlonal de conejo anti U1 A Control: anticuerpo policlonal de conejo específico Control: anticuerpo policlonal de conejo específico CONCLUSIÓN: snRNP U1 participa en el complejo Proteína-RNA formado por ATM ISPE

43
CONCLUSIONES Sólo el ATM ISPE normal induce la formación del complejo Proteína-RNA específico Sólo el ATM ISPE normal induce la formación del complejo Proteína-RNA específico Las Variantes ATM-Δ o ATM-CT no pueden desplazar al ATM ISPE del complejo proteína-RNA Las Variantes ATM-Δ o ATM-CT no pueden desplazar al ATM ISPE del complejo proteína-RNA snRNP U1 participa en el complejo proteína-RNA formado por ATM ISPE snRNP U1 participa en el complejo proteína-RNA formado por ATM ISPE

44
Interacción funcional entre U1 snRNA e ISPE Determinar si la complementariedad entre mutantes de U1 snRNA, denominadas ∆-U1 y CT-U1, y las variantes complementarias ATM-∆ y ATM-CT induce el procesamiento normal del intrón in vivo OBJETIVO

45
Interacción funcional entre U1 snRNA e ISPE

46
MATERIALES Y MÉTODOS Amplificación de DNA genómico y preparación de plásmidos pATM-WT, pATM-∆ y pATM-CT con minigenes Amplificación de DNA genómico y preparación de plásmidos pATM-WT, pATM-∆ y pATM-CT con minigenes Creación de las variantes ∆-U1 y CT-U1 empleando el clon pG3U1 (clon parental de U1 snRNA) Creación de las variantes ∆-U1 y CT-U1 empleando el clon pG3U1 (clon parental de U1 snRNA) Co-transfección de células Hep3B con las variantes de los minigenes de ATM y con las mutantes del U1 snRNA (cantidades diferentes) Co-transfección de células Hep3B con las variantes de los minigenes de ATM y con las mutantes del U1 snRNA (cantidades diferentes) Preparación del RNA total mediante extracción con ácido guanidino-fenol y transcripción por RT-PCR Preparación del RNA total mediante extracción con ácido guanidino-fenol y transcripción por RT-PCR Visualización mediante geles de agarosa Visualización mediante geles de agarosa
 Creación de las variantes ∆-U1 y CT-U1 empleando el clon pG3U1 (clon parental de U1 snRNA) Co-transfección de células Hep3B con las variantes de los minigenes de ATM y con las mutantes del U1 snRNA (cantidades diferentes) Co-transfección de células Hep3B con las variantes de los minigenes de ATM y con las mutantes del U1 snRNA (cantidades diferentes) Preparación del RNA total mediante extracción con ácido guanidino-fenol y transcripción por RT-PCR Preparación del RNA total mediante extracción con ácido guanidino-fenol y transcripción por RT-PCR Visualización mediante geles de agarosa Visualización mediante geles de agarosa")
47
RESULTADOS +, 0,5 µg; +, 1,5 µg La interacción entre pATM-∆ y pATM-CT con sus respectivas variantes complementarias del U1 snRNA produce la omisión del exón críptico Interacción funcional entre U1 snRNA e ISPE

48
La restauración de la complementariedad entre U1 snRNA e ISPE genera un splicing normal, es decir, permite la remoción eficiente de los intrones CONCLUSIÓN

49
Conversión de exones en intrones OBJETIVO Analizar el efecto del ISPE en un contexto diferente HIPÓTESIS Si la interacción de U1 snRNP con el ISPE provoca la correcta remoción de intrones, entonces su localización en un exón convertiría secuencias exónicas en secuencias intrónicas

50
Conversión de exones en intrones ESTRATEGIA EXPERIMENTAL 1 Introducción de la secuencia core del ISPE en diferentes posiciones del exón 9 del gen CFTR, utilizando una versión modificada del minigen globina-fibronectina EDB

51
Conversión de exones en intrones ESTRATEGIA EXPERIMENTAL 1

52
Conversión de exones en intrones ANÁLISIS DE LOS PRODUCTOS RT-PCR mRNA RT cDNA Amplificación

53
Conversión de exones en intrones OBSERVACIONES En células transfectadas con pCF, el análisis por RT-PCR muestra un predominio de inclusión del exón 9 En células transfectadas con pCF, el análisis por RT-PCR muestra un predominio de inclusión del exón 9 En células transfectadas con pCF-ISPE 5´ y en las transfectadas con pCF-ISPE 3´ se observa un predominio de exclusión del exón 9 En células transfectadas con pCF-ISPE 5´ y en las transfectadas con pCF-ISPE 3´ se observa un predominio de exclusión del exón 9 En células transfectadas con pCF-ATM y en las transfectadas con pCF-ATM CT, se observa un aumento importante de exclusión del exón 9 con respecto al pCF En células transfectadas con pCF-ATM y en las transfectadas con pCF-ATM CT, se observa un aumento importante de exclusión del exón 9 con respecto al pCF

54
Conversión de exones en intrones ESTRATEGIA EXPERIMENTAL 1 Co-transfección de las variantes ISPEs con su respectivo U1 snRNP complementario utilizando la línea celular Hep 3B

55
Conversión de exones en intrones ANÁLISIS DE LOS PRODUCTOS RT-PCR

56
Conversión de exones en intrones OBSERVACIONES La co-transfección de las dos variantes ISPE con sus respectivas U1 snRNP complementarias muestran una inhibición específica de la inclusión del exón 9 La co-transfección de las dos variantes ISPE con sus respectivas U1 snRNP complementarias muestran una inhibición específica de la inclusión del exón 9

57
La presencia del ISPE en un exón lo convierte en un intrón La presencia del ISPE en un exón lo convierte en un intrón La interacción entre ISPE y U1 sn RNP también convierte secuencia exónicas en intrónicas en un contexto heterólogo La interacción entre ISPE y U1 sn RNP también convierte secuencia exónicas en intrónicas en un contexto heterólogo CONCLUSIONES

58
CONCLUSIONES FINALES Se identificó un nuevo mecanismo que provoca A-T y que involucra un elemento procesador del splicing intrónico (ISPE) en ATM Se identificó un nuevo mecanismo que provoca A-T y que involucra un elemento procesador del splicing intrónico (ISPE) en ATM La deleción GTAA dentro del ISPE interrumpe la interacción no canónica U1 snRNP y activa un exón aberrante, manteniendo los sitios de splicing consenso 5´ y 3´ potenciales adyacentes La deleción GTAA dentro del ISPE interrumpe la interacción no canónica U1 snRNP y activa un exón aberrante, manteniendo los sitios de splicing consenso 5´ y 3´ potenciales adyacentes La deleción ocurre 12 pb río abajo y 53 pb río arriba respecto de los sitios de splicing del exón críptico (esas secuencias ya están presentes en el intrón wild-type) La deleción ocurre 12 pb río abajo y 53 pb río arriba respecto de los sitios de splicing del exón críptico (esas secuencias ya están presentes en el intrón wild-type)
 en ATM Se identificó un nuevo mecanismo que provoca A-T y que involucra un elemento procesador del splicing intrónico (ISPE) en ATM La deleción GTAA dentro del ISPE interrumpe la interacción no canónica U1 snRNP y activa un exón aberrante, manteniendo los sitios de splicing consenso 5´ y 3´ potenciales adyacentes La deleción GTAA dentro del ISPE interrumpe la interacción no canónica U1 snRNP y activa un exón aberrante, manteniendo los sitios de splicing consenso 5´ y 3´ potenciales adyacentes La deleción ocurre 12 pb río abajo y 53 pb río arriba respecto de los sitios de splicing del exón críptico (esas secuencias ya están presentes en el intrón wild-type) La deleción ocurre 12 pb río abajo y 53 pb río arriba respecto de los sitios de splicing del exón críptico (esas secuencias ya están presentes en el intrón wild-type)")
59
PERSPECTIVAS Se sabe que U1 snRNP también está implicada en otros eventos del procesamiento de pre-mRNA: Regulación de la formación del sitio 3´ terminal Regulación de la formación del sitio 3´ terminal Refuerzo de los bordes intrónicos por unión a tripletes G Refuerzo de los bordes intrónicos por unión a tripletes G Inhibición del splicing por unión a moduladores negativos del splicing de intrones en ROUS sarcoma virus Inhibición del splicing por unión a moduladores negativos del splicing de intrones en ROUS sarcoma virus Splicing tejido específico de los transcriptos del elemento P de Drosophila Splicing tejido específico de los transcriptos del elemento P de Drosophila

60
PERSPECTIVAS El complejo que forma U1 snRNP con la secuencia ISPE debe interferir con los posibles complejos en sitios crípticos de splicing adyacentes El complejo que forma U1 snRNP con la secuencia ISPE debe interferir con los posibles complejos en sitios crípticos de splicing adyacentes En transcriptos de elementos P de Drosophila, el reclutamiento de U1 snRNP a un sitio que nunca es utilizado para splicing resulta en una concomitante reducción del complejo U1 snRNP/ISPE en el sitio apropiado de slicing En transcriptos de elementos P de Drosophila, el reclutamiento de U1 snRNP a un sitio que nunca es utilizado para splicing resulta en una concomitante reducción del complejo U1 snRNP/ISPE en el sitio apropiado de slicing

61
PERSPECTIVAS Se requiere una distancia crítica entre los sitios 3´ y 5´ de splicing para definir un exón Se requiere una distancia crítica entre los sitios 3´ y 5´ de splicing para definir un exón Elementos que contienen sitios de splicing 3´ y 5´ adyacentes (similares al ISPE) pueden representar una señal de re-splicing que regula y asegura el correcto procesamiento de intrones largos (Drosophila Ubx) ó comportarse como un sitio (aceptor y donor) de splicing alternativo Elementos que contienen sitios de splicing 3´ y 5´ adyacentes (similares al ISPE) pueden representar una señal de re-splicing que regula y asegura el correcto procesamiento de intrones largos (Drosophila Ubx) ó comportarse como un sitio (aceptor y donor) de splicing alternativo En este contexto, los sitios de pseudo-splicing frecuentemente encontrados en intrones deben ser considerados no solo como “falsos exones” sino también como elementos importantes para el eficiente procesamiento intrónico En este contexto, los sitios de pseudo-splicing frecuentemente encontrados en intrones deben ser considerados no solo como “falsos exones” sino también como elementos importantes para el eficiente procesamiento intrónico
 pueden representar una señal de re-splicing que regula y asegura el correcto procesamiento de intrones largos (Drosophila Ubx) ó comportarse como un sitio (aceptor y donor) de splicing alternativo Elementos que contienen sitios de splicing 3´ y 5´ adyacentes (similares al ISPE) pueden representar una señal de re-splicing que regula y asegura el correcto procesamiento de intrones largos (Drosophila Ubx) ó comportarse como un sitio (aceptor y donor) de splicing alternativo En este contexto, los sitios de pseudo-splicing frecuentemente encontrados en intrones deben ser considerados no solo como falsos exones sino también como elementos importantes para el eficiente procesamiento intrónico En este contexto, los sitios de pseudo-splicing frecuentemente encontrados en intrones deben ser considerados no solo como falsos exones sino también como elementos importantes para el eficiente procesamiento intrónico")
62
UN NUEVO TIPO DE MUTACIÓN CAUSA UN DEFECTO EN EL SPLICING DEL GEN ATM MUCHAS GRACIAS!

Presentaciones similares