Descargar la presentación
La descarga está en progreso. Por favor, espere

1
elementos necesarios en eucariotas en procariotas
TRADUCCIÓN ARN de transferencia Aminoacil tRNA sintetasa Ribosomas El proceso de traducción elementos necesarios en eucariotas La información contenida en los genes es perpetuada con la replicación del material genético, el objetivo último de la información contenida en los genes es ser expresada, ejerciendo así la función que les es única. La información genética se perpetúa con el proceso de replicación del ADN. La expresión de dicha información sigue un camino en dos etapas, la primera de ellas es la transcripción, en la que se obtiene una cadena de ARN. La segunda etapa es la traducción, que convierte la secuencia de nucleótidos del ARN en la secuencia de aminoácidos que conforma la proteína. en procariotas

2
aa tRNA sintetasa cargado
ribosoma mRNA Los elementos que participan en la traducción son: los ARN de transferencia, las aminoacil tRNA sintetasas que permiten cargar a los ARN de transferencia con el aminoácido que les es específico, el ARN mensajero y el ribosoma. A continuación se describirán cada uno de estos elementos, a excepción de ARN mensajero que ya fue descrito en las sesiones anteriores. Role of aminoacyl-tRNA formation in the elongation phase of protein synthesis. An uncharged tRNA is first aminoacylated with the appropriate amino acid to generate an aminoacyltRNA, which then interacts with a translation elongation factor. This allows delivery of the aminoacyl-tRNA to the ribosomal A site, where its anticodon can interact with the corresponding codon in mRNA. The example shown illustrates how this leads to the translation of the codon GGU as tryptophan.

3
tRNAs Adaptan física e informacionalmente
al mRNA con la proteína codificada La interpretación del mensaje contenido en el ARNm se realiza por medio de los ARN de transferencia, mismos que son los únicos capaces de relacionar tripletes de nucleótidos con un aminoácido. La estructura secundaria de cientos de tRNA ajusta con la forma de un trébol mantenida por apareamiento de bases entre regiones complementarias.

4
Prokaryotic tRNA genes
Los ARN de transferencia están codificados por genes, que en procariotas son mas o menos uniformes en tamaño (75 a 95 bp). Las secuencias de dichos genes varían considerablemente entre los genes pero aún así se pueden distinguir cuatro o cinco repetidos invertidos que definen los elementos característicos regularmente espaciados de la molécula ARN de transferencia : los tallos y rizos, que se moldean en la forma conocida de trébol. Todos los genes conocidos de E. coli carecen de intrones y presentan una secuencia CCA en 3’. En coli, los 79 genes para los 46 ARNt (o 41 anticodones) están esparcidos por el genoma en 41 unidades de transcripción. De estos 79 genes solo 20 existen como genes simples, los demás están organizados en operones policistrónicos, conteniendo ya sea genes para el mismo tRNA, para diferentes tRNA o bien genes para proteínas. Hay que destacar que en coli, 14 genes de tRNA se encuentran en 7 operones de ARN ribosomales, localizados en las regiones espaciadoras entre los genes que codifican para los ARNr de 16S y 23S.
. Las secuencias de dichos genes varían considerablemente entre los genes pero aún así se pueden distinguir cuatro o cinco repetidos invertidos que definen los elementos característicos regularmente espaciados de la molécula ARN de transferencia : los tallos y rizos, que se moldean en la forma conocida de trébol. Todos los genes conocidos de E. coli carecen de intrones y presentan una secuencia CCA en 3’. En coli, los 79 genes para los 46 ARNt (o 41 anticodones) están esparcidos por el genoma en 41 unidades de transcripción. De estos 79 genes solo 20 existen como genes simples, los demás están organizados en operones policistrónicos, conteniendo ya sea genes para el mismo tRNA, para diferentes tRNA o bien genes para proteínas. Hay que destacar que en coli, 14 genes de tRNA se encuentran en 7 operones de ARN ribosomales, localizados en las regiones espaciadoras entre los genes que codifican para los ARNr de 16S y 23S.")
6
Eukaryotic tRNA genes Transcripción de genes eucarióticos de tRNA implica el concurso, entre otros, de factores de transcripción asociados a secuencias localizadas en un par de cajas, A y B. Los complejos que se originan son reconocidos por la RNA pol III.

7
7 pb 5 pb 3-4 pb Cada uno de los brazos que aparecen en el esquema se caracterizan ya sea por su estructura o su función. En el sentido de las manecillas del reloj, el primer brazo es el brazo aceptor consistente en un tallo con bases apareadas que termina con una base no apareada y su OH aminoacilado. Los otros tres brazos consisten de tallos apareados y rizos no apareados. El segundo brazo es conocido como TyC ya que contiene el codón para un aminoácido poco común (pseudouridina). El tercer brazo es el brazo anticodón ya que cuenta con el triplete anticodón al centro del rizo. El cuarto brazo se conoce como brazo D por la presencia de dihidrouridina (una base modificada del tRNA). El número de bases en los tRNA puede ir desde 74 hasta 95, las diferencias obedecen básicamente a variaciones en dos de los brazos, el D y otro localizado entre el TyC y el anticodón. Dicho brazo se llama brazo variable, su composición determina el tipo de tRNA del que se trata entre dos clases. La clase 1 presenta este brazo extra pequeño, de 3 a 5 bases, representa el 75% de los tRNA. La clase 2 cuenta con el brazo extra de un tamaño largo (13 a 21 bases y 5 pares de bases en el tallo).
. El tercer brazo es el brazo anticodón ya que cuenta con el triplete anticodón al centro del rizo. El cuarto brazo se conoce como brazo D por la presencia de dihidrouridina (una base modificada del tRNA). El número de bases en los tRNA puede ir desde 74 hasta 95, las diferencias obedecen básicamente a variaciones en dos de los brazos, el D y otro localizado entre el TyC y el anticodón. Dicho brazo se llama brazo variable, su composición determina el tipo de tRNA del que se trata entre dos clases. La clase 1 presenta este brazo extra pequeño, de 3 a 5 bases, representa el 75% de los tRNA. La clase 2 cuenta con el brazo extra de un tamaño largo (13 a 21 bases y 5 pares de bases en el tallo).")
8
Elementos de identidad en los tRNAs

9
Elementos de identidad en los tRNAs que permiten la identificacion especifica con la rRNA sintetasa.

11
Estructura terciaria de un tRNA

13
AA + ATP AARS AA-AMP PPi AARS AA-AMP + tRNA AA-tRNA AMP
Aminoacyl-tRNAs are synthesized by the 3’-esterification of tRNAs with the appropriate amino acids. For the majority of aminoacyl-tRNAs this is accomplished by direct aminoacylation of a particular tRNA with the corresponding amino acid in a two-step reaction: where AA is an amino acid and AARS is the corresponding aminoacyl-tRNA synthetase.

14
La aminoacilación de tRNA es catalizada por una familia de enzimas llamadas aminoacil-tRNA sintetasas Cada aminoacil tRNA sintetasa es responsable del establecimiento del código genético a través de la aminoacilación específica de las contrapartes del tRNA, asociando un aminoácido con sus tripletes anticodones correspondientes.

15
enlace covalente del aminoácido al extremo 3´CCA del tRNA

16
Activación y anclaje covalente del aminoácido al tRNA

18
Todas las aminoacil tRNA sintetasas catalizan la misma reacción, en la primera etapa de la misma, ATP y aminoácido se unen al sitio activo de la sintetasa, el carboxilato del aminoácido realiza un ataque nucleofílico sobre el fosfato del ATP, esto permite la formación de un complejo enzima-aminoacil-adenilato y la liberación de un grupo pirofosfato inorgánico.

19
En la segunda etapa de la reacción ocurre un ataque nucleofílico entre el OH en 2’ o 3’ de la adenosina terminal del tRNA sobre el alfa carbonilo del aminoacil-adenilato, obteniéndose la esterificación en 3’ del tRNA con el aminoácido y la liberación de AMP, seguido de la liberación del producto.

20
E. coli GlnRS·tRNAgln · ATP Complejo del tipo I
Yeast AspRS·tRNAasp·ATP Complejo del tipo II A pesar de lo conservado que es esta reacción de 3’ esterificación del tRNA con el aminoácido requerido, las sintetasas se agrupan en dos clases diferentes. Dichas clases presentan características de secuencia exclusivas que reflejan topologías exclusivas del sitio activo. Las sintetasas de la clase I tienen la particularidad de presentar en su sitio activo un dominio de unión a dinucleótido tipo Rossmann. Las sintetasas de tipo II carecen de dicho plegamiento, en su lugar presentan un plegamiento tipo beta antiparalelo. Como resultado de los plegamientos del sitio activo en ambos tipos de sintetasa, la unión a ATP en las enzimas de la clase I se realiza en una conformación extendida mientras que en las enzimas de la clase II la unión es “torcida”. La unión al tRNA también es diferente entre las dos clases de enzima. Derivado de estudios estructurales se sabe que las enzimas de clase I se unen al brazo aceptor del tRNA desde el surco menor de modo que el brazo variable se encuentra del lado del solvente. En las enzimas de clase II, la aproximación de la enzima al tRNA se realiza por el lado del surco mayor del brazo aceptor de modo que el brazo variable está del lado de la sintetasa.

21
Clase I sintetasa

22
Clase II sintetasa

24
Estructura y función del ribosoma
El ensamblaje de aminoácidos en proteínas gracias al ribosoma, que es una ribonucleoproteína compacta que consiste de 2 subunidades. Cada subunidad está conformada por varias proteínas asociadas con una molécula de ARN. Los ribosomas son descritos en función de las unidades de sedimentación medidas en Svedbergs. Ribosomas bacterianos sedimentan generalmente a 70S, los eucarióticos son generalmente mas grandes, del orden de 80S. Las subunidades ribosomales se disocian al reducir la concentración de Mg++.

25
sitios funcionales en el ribosoma
5’ 3’

26
Estructura de la subunidad grande
1. rRNA 16S 2. rRNA 5S 3. proteínas (LSU) 4. centro peptidil-transferasa y mecanismo catalítico
 4. centro peptidil-transferasa y. mecanismo catalítico.")
27
Estructura cristalografica del ribosoma 70S de Thermus thermofilus
Estructura cristalografica del ribosoma 70S de Thermus thermofilus. En A se ve la estructura del ribosoma desde atrás de 30S. B muestra la vista desde el lado, mostrando la interfase entre 30S y 50S, a la izquierda y derecha respectivamente, se distingue en la interfase el tRNA posicionado en A. C muestra al ribosoma desde atrás de 50S, en la parte inferior se muestra la salida del canal de salida del polipeptido. D muestra 50S a la izquierda y 30S a la derecha, en rojo se ve el tRNA que ocupa el sitio P.

28
rRNAs 23S y 5S en la estructura terciaria del ribosoma

29
estructura terciaria del rRNA 5s

30
Bases conservadas en la estructura terciaria de la subunidad grande
bases muy conservadas (rojo) ligado de substrato y factores actvidad catalítica estabilización de estructura tridimensional (gran número de adeninas) extensiones variables (verde)
 ligado de substrato y factores. actvidad catalítica. estabilización de estructura tridimensional (gran número de adeninas) extensiones variables. (verde)")
31
Estructura de proteínas ribosomales
13/30 proteínas presentan extensiones largas: estabilizan el plegamiento terciario rellenan los huecos funcionan como un pegamento molecular 23/29 interaccionan con más de un dominio rRNA 23s - 1157/2923 nucleótidos hacen contactos de van der Waals con proteínas

32
Localización de proteínas en la subunidad grande del ribosoma
Estructura cristalográfica del ribosoma de Haloarcula marismortui a 2.4Å

33
Túnel de salida de la proteína y extensiones cercanas de proteínas ribosomales

34
Túnel de salida de la proteína en la subunidad grande del ribosoma

35
Comparación de las estructuras secundarias predichas para los rRNAs 16S de (a) Halobacterium volcanii, (b) levadura y (c) mitocondria bovina
 Halobacterium volcanii, (b) levadura y (c) mitocondria bovina")
36
Traducción 0.1 mm Fibroína
Protein synthesis is accomplished through the process of translation of an mRNA sequence into a polypeptide chain. This process can be divided into three distinct stages: initiation, elongation and termination. During the initiation phase, the two subunits of the ribosome are brought together to the translation start site on the mRNA where the polypeptide chain is to begin. Extension of the polypeptide chain occurs when a specific aminoacyl-tRNA, as determined by the template mRNA, binds an elongating ribosome. The protein chain is released from the ribosome when any one of three stop codons in the relevant reading frame on the mRNA is reached. Individual reactions at each one of these stages are catalyzed by a number of initiation, elongation and release factors, respectively. En el proceso de la traducción, la identidad de un aminoácido insertado en una posición particular durante la síntesis de la proteína, está determinada por el apareamiento de un codón en el ARNm con un aminoacil-tRNA particular. La fidelidad del proceso de síntesis depnde de la precisión de dos procesos en particular a) El reconocimiento del codón-anticodón b) La síntesis del aminoacil-tRNA 0.1 mm Fibroína
 El reconocimiento del codón-anticodón. b) La síntesis del aminoacil-tRNA. 0.1 mm. Fibroína.")
37
Role of aminoacyl-tRNA formation in the elongation phase of protein synthesis. An
uncharged tRNA is first aminoacylated with the appropriate amino acid to generate an aminoacyltRNA, which then interacts with a translation elongation factor. This allows delivery of the aminoacyl-tRNA to the ribosomal A site, where its anticodon can interact with the corresponding codon in mRNA. The example shown illustrates how this leads to the translation of the codon GGU as tryptophan.

38
Resumen del tema de la traducción

39
Bioquímica de la traducción
1. Activación del aminoácido (ATP, aminoacil-tRNA sintetasa) 2. Pegado covalente del AA al 3´OH del CCA terminal del tRNA 3. Unión del tRNA al mRNA por apareaminento codón-anticodón 4. Formación del enlace peptídico en el ribosoma 5. Terminación del polipéptido Procariotes: 20aa/s = 20s por 400 AA/proteína Eucariotes: 2 aa/s
 2. Pegado covalente del AA al 3´OH del CCA terminal del tRNA. 3. Unión del tRNA al mRNA por apareaminento codón-anticodón. 4. Formación del enlace peptídico en el ribosoma. 5. Terminación del polipéptido. Procariotes: 20aa/s = 20s por 400 AA/proteína. Eucariotes: 2 aa/s.")
41
INICIO DE LA TRADUCCIÓN
Comprende las reacciones que preceden la formación del enlace peptídico entre los dos primeros aminoácidos. Initiation of translation in the majority of eukaryotic cellular mRNAs depends on the 5’-cap (m7GpppN) and involves ribosomal scanning of the 5' untranslated region (5’-UTR) for an initiating AUG start codon. Therefore, this mechanism is often called cap-dependent translation initiation. Proximity to the cap, as well as the nucleotides surrounding an AUG codon, influence the efficiency of the start site recognition during the scanning process. However, if the recognition site is poor enough, scanning ribosomal subunits will ignore and skip potential starting AUGs, a phenomenon called leaky scanning. Leaky scanning allows a single mRNA to encode several proteins that differ in their amino-termini. Several eukaryotic cell and viral mRNAs initiate translation by an alternative mechanism that involves internal initiation rather than ribosomal scanning. These mRNAs contain complex nucleotide sequences, called internal ribosomal entry sites, where ribosomes bind in a cap-independent manner and start translation at the closest downstream AUG codon.
 and involves ribosomal scanning of the 5 untranslated region (5’-UTR) for an initiating AUG start codon. Therefore, this mechanism is often called cap-dependent translation initiation. Proximity to the cap, as well as the nucleotides surrounding an AUG codon, influence the efficiency of the start site recognition during the scanning process. However, if the recognition site is poor enough, scanning ribosomal subunits will ignore and skip potential starting AUGs, a phenomenon called leaky scanning. Leaky scanning allows a single mRNA to encode several proteins that differ in their amino-termini. Several eukaryotic cell and viral mRNAs initiate translation by an alternative mechanism that involves internal initiation rather than ribosomal scanning. These mRNAs contain complex nucleotide sequences, called internal ribosomal entry sites, where ribosomes bind in a cap-independent manner and start translation at the closest downstream AUG codon.")
42
INICIO DE LA TRADUCCIÓN
C A P DEPENDIENTE Translation initiation is a complex process in which the Met-tRNAi initiator, 40S, and 60S ribosomal subunits are assembled by eukaryotic initiation factors (eIFs) into an 80S ribosome at the start codon of an mRNA. The basic mechanism for this process can be described as a series of five steps:
 into an 80S ribosome at the start codon of an mRNA. The basic mechanism for this process can be described as a series of five steps:")
43
Formation of a pool of free 40S subunits
The 80S ribosome dissociates into free 40S (small) and 60S (large) ribosomal subunits. Each ribosomal subunit is constituted by several individual ribosomal proteins and rRNA.
 and 60S (large) ribosomal subunits. Each ribosomal subunit is constituted by several individual ribosomal proteins and rRNA.")
44
2) formation of the ternary complex (Met-tRNAi/eIF2/GTP), and subsequently, the 43S complex (comprising the 40S subunit, Met-tRNAi/eIF2/GTP, eIF3 and eIF1A),
 formation of the ternary complex (Met-tRNAi/eIF2/GTP), and subsequently, the 43S complex (comprising the 40S subunit, Met-tRNAi/eIF2/GTP, eIF3 and eIF1A),")
45
3) activation of the mRNA upon binding of the cap-binding complex eIF4F, and factors eIF4A, eIF4B and eIF4H, with subsequent binding to the 43S complex, 4) ribosomal scanning and start codon recognition. The initiation codon of a eukaryotic mRNA is normally the first AUG triplet downstream of the 5’-terminal cap and is usually separated from it by nt. After Cap-mediated attachment to mRNA, a 43S complex is thought to scan downstream from the 5’-end until it encounters the initiation codon.
 ribosomal scanning and start codon recognition. The initiation codon of a eukaryotic mRNA is normally the first AUG triplet downstream of the 5’-terminal cap and is usually separated from it by nt. After Cap-mediated attachment to mRNA, a 43S complex is thought to scan downstream from the 5’-end until it encounters the initiation codon.")
46
5) GTP hydrolysis and joining of the 60S ribosomal subunit.
 GTP hydrolysis and joining of the 60S ribosomal subunit.")
47
ELONGACIÓN DE LA TRADUCCIÓN
Comprende las reacciones desde la síntesis del primer enlace peptídico hasta la adición del último aa. Elongation of a polypeptide chain depends on the stepwise movement of the ribosome along the mRNA chain. A ribosome contains one binding site for the mRNA molecule and two binding sites for tRNAs. The peptidyl-tRNA-binding site (P-site) holds the tRNA molecule that is linked to the growing end of the polypeptide chain. The aminocyl-tRNA-binding site (A-site) holds the incoming tRNA molecule charged with an amino acid. The elongation process can be divided in three discrete steps. First, an aminoacyl-tRNA molecule binds to the A-site on the ribosome by forming base pairs with three mRNA nucleotides (codon). Second, the carboxyl end of the polypeptide chain is uncoupled from the tRNA molecule in the P-site and forms a new peptide bond with the amino acid that is in the A-site. This central reaction is catalyzed by a peptidyl transferase enzyme. Third, the ribosome moves a distance of three nucleotides along the mRNA chain, ejects the unlinked tRNA and �resets� the ribosome so that the next aminoacyl-tRNA molecule can bind. This three-step cycle is then repeated over and over until a stop codon is reached, causing translation to be terminated.
 holds the tRNA molecule that is linked to the growing end of the polypeptide chain. The aminocyl-tRNA-binding site (A-site) holds the incoming tRNA molecule charged with an amino acid. The elongation process can be divided in three discrete steps. First, an aminoacyl-tRNA molecule binds to the A-site on the ribosome by forming base pairs with three mRNA nucleotides (codon). Second, the carboxyl end of the polypeptide chain is uncoupled from the tRNA molecule in the P-site and forms a new peptide bond with the amino acid that is in the A-site. This central reaction is catalyzed by a peptidyl transferase enzyme. Third, the ribosome moves a distance of three nucleotides along the mRNA chain, ejects the unlinked tRNA and �resets� the ribosome so that the next aminoacyl-tRNA molecule can bind. This three-step cycle is then repeated over and over until a stop codon is reached, causing translation to be terminated.")
48
La elongación ocurre cuando el siguiente aminoacil tRNA (después del de la Met) llega al sitio A. El anticodón reconoce el codón correspondiente (valina en este ejemplo). Ahora Met y Val se encuentran adyacentes en los sitios P y A respectivamente. Por medio de una peptidil transferasa se forma un enlace peptídico entre Met y Val.
. Ahora Met y Val se encuentran adyacentes en los sitios P y A respectivamente. Por medio de una peptidil transferasa se forma un enlace peptídico entre Met y Val..")
49
Mecanismo de formación del enlace peptídico

50
El ribosoma es translocado un codón hacia la derecha permitiendo la eyección del tRNA descargado. El proceso se repite adicionándose un aminoácido cada vez qu el ribosoma se mueve.

51
Formación del enlace peptídico

52
TERMINACIÓN DE LA TRADUCCIÓN
Incluye los pasos necesarios para la liberación del polipéptido completo, al mismo tiempo que se disocia el ribosoma del ARNm. The arrival of any of the three stop codons (UAA, UAG and UGA) into the ribosomal A-site triggers the binding of a release factor (RF) to the ribosome and subsequent polypeptide chain release. In eukaryotes, the RF is composed of two proteins, eRF1 and eRF3. eRF1 is responsible for the hydrolysis of the peptidyl-tRNA, while eRF3 provides a GTP-dependent function. The ribosome releases the mRNA and dissociates into its two complex subunits, which can reassemble on another molecule to begin a new round of protein synthesis.
 into the ribosomal A-site triggers the binding of a release factor (RF) to the ribosome and subsequent polypeptide chain release. In eukaryotes, the RF is composed of two proteins, eRF1 and eRF3. eRF1 is responsible for the hydrolysis of the peptidyl-tRNA, while eRF3 provides a GTP-dependent function. The ribosome releases the mRNA and dissociates into its two complex subunits, which can reassemble on another molecule to begin a new round of protein synthesis.")
53
Ningún anticodón en tRNA corresponde con el codón de terminación, en su lugar un factor de liberación es acoplado al codón STOP. En este momento se acciona la peptidil transferasa provocando la unión del último aminoácido, la liberación del polipéptido y la eyección del tRNA descargado.

54
Mecanismo de terminación del polipéptido

55
El factor de liberación es se desacopla y se desensamblan las subunidades ribosomales.

58
Mecanismo de inicio de la traducción en E. coli
En procariotas, el inicio de la traducción se da en el sitio de unión a ribosoma (RBS), secuencia que se encuentra antes de la secuencia codificante. La reacción ocurre en dos pasos: Reconocimiento del ARNm ocurre cuando una pequeña subunidad (30S) se une y forma el complejo de inicio en el RBS. Los factores de inicio se encuentran presentes solamente en subunidades 30S disociadas (libres). b) Entonces una subunidad grande (50S) se une al complejo de inicio y genera un ribosoma completo (70S). Cuando se asocia la subunidad 50S con la unidad 30S se liberan los factores de inicio. La liberación de los IFs distingue las dos etapas del inicio de la traducción en procariotas. Los factores de inicio son: IF1, IF2 e IF3. IF1 parece estabilizar el complejo de inicio uniéndose a la subunidad 30S. IF2 se une al tRNA especial de inicio y lo conduce al complejo de inicio. IF3 se necesita para que la subunidad 30S se una específicamente al complejo de inicio. Estabiliza 30S libres y les permite unirse a mRNA. Una vez que IF3 se une a 30S, no se puede unir 50S, necesita disociarse IF3 para que esta conjunción ocurra (30S + 50S). También es importante que IF3 esté unido a 30S porque de otro modo no se puede formar el complejo de inicio. Pero la especificidad de la unión depende únicamente del ribosoma, la unión de IF3 es necesaria para la reacción pero no para la selección del sitio. La secuencia del ARNm a la que se une 30S con IF3 se conoce como Shine-Dalgarno, que es parte del RBS, este último contiene un codón de inicio especial, normalmente es el triplete AUG, que representa metionina. En bacetrias, el extremo amino de la metionina inicial ha sido formilado
, secuencia que se encuentra antes de la secuencia codificante. La reacción ocurre en dos pasos: Reconocimiento del ARNm ocurre cuando una pequeña subunidad (30S) se une y forma el complejo de inicio en el RBS. Los factores de inicio se encuentran presentes solamente en subunidades 30S disociadas (libres). b) Entonces una subunidad grande (50S) se une al complejo de inicio y genera un ribosoma completo (70S). Cuando se asocia la subunidad 50S con la unidad 30S se liberan los factores de inicio. La liberación de los IFs distingue las dos etapas del inicio de la traducción en procariotas. Los factores de inicio son: IF1, IF2 e IF3. IF1 parece estabilizar el complejo de inicio uniéndose a la subunidad 30S. IF2 se une al tRNA especial de inicio y lo conduce al complejo de inicio. IF3 se necesita para que la subunidad 30S se una específicamente al complejo de inicio. Estabiliza 30S libres y les permite unirse a mRNA. Una vez que IF3 se une a 30S, no se puede unir 50S, necesita disociarse IF3 para que esta conjunción ocurra (30S + 50S). También es importante que IF3 esté unido a 30S porque de otro modo no se puede formar el complejo de inicio. Pero la especificidad de la unión depende únicamente del ribosoma, la unión de IF3 es necesaria para la reacción pero no para la selección del sitio. La secuencia del ARNm a la que se une 30S con IF3 se conoce como Shine-Dalgarno, que es parte del RBS, este último contiene un codón de inicio especial, normalmente es el triplete AUG, que representa metionina. En bacetrias, el extremo amino de la metionina inicial ha sido formilado.")
59
Elongación de la traducción
Elongación de la traducción. Una vez que el ribosoma está unido al codón de incio,comienza el ciclo en el que el sitio A del ribosoma es ocupado por un aminoacil-tRNA y el sitio P es ocupado por un peptidil-tRNA. La entrada de los aminoacil-tRNA es mediada por un factor de elongación (EF-Tu). El estado, activo o inactivo, de dicho factor EF-Tu depende de la presencia de GTP. Cuando este nucleótido de guanidina está en su forma GTP, EF-Tu está activo, cuando hay GDP EF-Tu está inactivo. Cuando el complejo ternario aminoacil-tRNA/EF-Tu/GTP se forma, entonces el aminoacil-tRNA puede ocupar el sitiio A del ribosoma siempre y cuando el sitio P esté ocupado por un peptidil-tRNA.
. El estado, activo o inactivo, de dicho factor EF-Tu depende de la presencia de GTP. Cuando este nucleótido de guanidina está en su forma GTP, EF-Tu está activo, cuando hay GDP EF-Tu está inactivo. Cuando el complejo ternario aminoacil-tRNA/EF-Tu/GTP se forma, entonces el aminoacil-tRNA puede ocupar el sitiio A del ribosoma siempre y cuando el sitio P esté ocupado por un peptidil-tRNA.")
60
Estructura del factor de iniciación de la traducción EF-Tu unido a GDP y a GDPNP

63
Orientación de las moléculas de tRNA unidas a los sitios E-, A- y P-
anticodones

64
Algunos antibióticos que funcionan como inhibidores de la traducción en bacterias

66
Rescate de ribosomas bacterianos por tmRNAs
Fig.1. Secondary structure diagram of tmRNA. Base pairs are linked by lines, whereas GU pairs are represented by dots. The TLD, the four pseudoknots (PK1 to PK4), and helices (numbered from 1 to 12) are shown in the colors that are used to represent these modules in subsequent figures. The nucleotides within the ORF are shown in a larger font. T. thermophilus sequence numbers are shown, with the corresponding E. coli numbering in parentheses. The figure is adapted from the diagram for E. coli on the tmRDB Web site (29, 41).
, and helices (numbered from 1 to 12) are shown in the colors that are used to represent. these modules in subsequent figures. The nucleotides within the ORF are shown in a larger font. T. thermophilus sequence numbers are shown, with the corresponding E. coli numbering in parentheses. The figure is adapted from the diagram for E. coli on the tmRDB Web site (29, 41).")
67
The trans-translation model. (a) tmRNA,
complexed with SmpB, is charged with alanine (represented by “A” in a blue box) by AlaRS, the alanyl tRNA-synthetase. EF-Tu, the ribosomal protein S1, and then (b) PrsA, SAF, RNAse R and tRNAala join the tmRNAala–SmpB complex and (c) enter a stalled ribosome. The alanine from tmRNA is transferred to the nascent peptide chain, followed by release of the original mRNA template. (d) tmRNA then becomes the template for translation, with the associated tRNAala probably being the first amino acid donor of the tag sequence. The remainder of the tag sequence is translated, and (e) the translational complex is dissociated upon reaching the stop codon encoded by tmRNA. The released peptide, tagged by tmRNA, is recognized by SspB and is degraded by ClpXP or other proteases.
 by AlaRS, the. alanyl tRNA-synthetase. EF-Tu, the ribosomal. protein S1, and then (b) PrsA, SAF, RNAse R. and tRNAala join the tmRNAala–SmpB complex. and (c) enter a stalled ribosome. The alanine. from tmRNA is transferred to the nascent. peptide chain, followed by release of the original. mRNA template. (d) tmRNA then becomes the. template for translation, with the associated. tRNAala probably being the first amino acid. donor of the tag sequence. The remainder of. the tag sequence is translated, and (e) the. translational complex is dissociated upon. reaching the stop codon encoded by tmRNA. The released peptide, tagged by tmRNA, is. recognized by SspB and is degraded by ClpXP. or other proteases.")
68
The ribosomal rescue model for tmRNA function. (a) When a ribosome
becomes stalled (as in the grey ribosome in the figure), tmRNAala enters the stalled ribosome and its alanine is transferred to the nascent peptide. (b) This ribosome is then released from the mRNA to continue with trans-translation. The tmRNA tag is translated by the released ribosome and the tagged protein is later degraded. Release of the stalled ribosome allows the other ribosomes on that mRNA to continue translation, and returns the stalled ribosome to the available pool upon completion of trans-translation. (c) If no tmRNA activity is present, the stalled ribosome is not efficiently removed from the mRNA, and this blocks further translation of that mRNA and prevents all the ribsomes engaged upon that mRNA from translating any other mRNA.
, tmRNAala. enters the stalled ribosome and its alanine is transferred to the nascent. peptide. (b) This ribosome is then released from the mRNA to continue. with trans-translation. The tmRNA tag is translated by the released. ribosome and the tagged protein is later degraded. Release of the. stalled ribosome allows the other ribosomes on that mRNA to continue. translation, and returns the stalled ribosome to the available pool upon. completion of trans-translation. (c) If no tmRNA activity is present, the. stalled ribosome is not efficiently removed from the mRNA, and this. blocks further translation of that mRNA and prevents all the ribsomes. engaged upon that mRNA from translating any other mRNA.")
69
Fig.2. Model for tmRNA, EF-Tu, and SmpB in the cryo-EM map of 70S ribosome in complex with
tmRNA. Cryo-EM map obtained for 70S•tRNA•EF-Tu•tmRNA•SmpB in the presence of GTP and kirromycin (A and B). The 50S subunit is depicted in blue, the 30S subunit in yellow, and the P-site tRNA in green. Semitransparent ribosomal subunits in (B) show the relative position between EF-Tu•tmRNA•SmpB and P-site tRNA. Landmarks on the 50S subunit: L1, stalk of protein L1; CP, central protuberance; L7/L12, stalk of the proteins L7/L12; and GAC, GTPase-associated center. Landmarks on the 30S subunit: h, head; b, beak; dc, decoding center; and ch, entrance of mRNA channel. The density attributable to the EF-Tu•tmRNA•SmpB complex is colored in red and depicted semitransparent in the stereo pair of (C) (orientation as in panel 2B). The docked atomic coordinates are shown in a ribbons representation generated with the programs RIBBONS or Insight. The .tting of the coordinates for EF-Tu•GTP analog [Protein Data Bank (PDB) code 1TTT] (25) and the 3D model for tmRNA (29) did not account for the whole density. The coordinates for SmpB (PDB code 1K8H) (26) were satisfactorily docked in the unexplained region. The ribbon colors for elements of tmRNA model are the same as in the other .gures: TLD, burgundy; loop within helix H2, light green; helix H2b-d, dark green; PK1, orange; PK4, dark blue; PK3, red; PK2, light blue; and helix H5, magenta.
. The 50S subunit is depicted in blue, the 30S subunit in yellow, and the P-site. tRNA in green. Semitransparent ribosomal subunits in (B) show the relative position between. EF-Tu•tmRNA•SmpB and P-site tRNA. Landmarks on the 50S subunit: L1, stalk of protein L1; CP, central protuberance; L7/L12, stalk of the proteins L7/L12; and GAC, GTPase-associated center. Landmarks on the 30S subunit: h, head; b, beak; dc, decoding center; and ch, entrance of mRNA. channel. The density attributable to the EF-Tu•tmRNA•SmpB complex is colored in red and depicted. semitransparent in the stereo pair of (C) (orientation as in panel 2B). The docked atomic. coordinates are shown in a ribbons representation generated with the programs RIBBONS or. Insight. The .tting of the coordinates for EF-Tu•GTP analog [Protein Data Bank (PDB) code 1TTT] (25) and the 3D model for tmRNA (29) did not account for the whole density. The coordinates for. SmpB (PDB code 1K8H) (26) were satisfactorily docked in the unexplained region. The ribbon colors. for elements of tmRNA model are the same as in the other .gures: TLD, burgundy; loop within helix. H2, light green; helix H2b-d, dark green; PK1, orange; PK4, dark blue; PK3, red; PK2, light blue; and. helix H5, magenta.")
70
Fig. 3. Interactions of SmpB•tmRNA with the 70S ribosome
Fig.3. Interactions of SmpB•tmRNA with the 70S ribosome. Two different ribosomal orientations are used to illustrate the interactions of SmpB and tmRNA with the 50S subunit (A) and the 30S subunit (B). The orientations are defined by ribosomal thumbnails on the left. Landmarks on the 50S subunit (A): L1, L1 stalk; L7/L12, stalk of proteins L7/L12; GAC, GTPase- associated center; H69, H71, and H89, helices of the 23S RNA. Labeling on the 30S subunit (B): ch, entrance of the mRNA channel; dc, decoding site; b, beak; loop, RNA loop within helix H2 of tmRNA; TLD, tRNA-like domain of tmRNA; S12, ribosomal protein S12; h44 and h38, the position of those helixes from 16S rRNA. Color coding for the tmRNA is as in Fig. 2C. For clarity, EF-Tu is not shown.
 and the 30S subunit (B). The orientations are defined by ribosomal thumbnails on the left. Landmarks on the 50S subunit (A): L1, L1 stalk; L7/L12, stalk of proteins L7/L12; GAC, GTPase- associated center; H69, H71, and H89, helices of the 23S RNA. Labeling on the 30S subunit (B): ch, entrance of the mRNA channel; dc, decoding site; b, beak; loop, RNA loop within helix H2 of tmRNA; TLD, tRNA-like domain of tmRNA; S12, ribosomal protein S12; h44 and h38, the position of those helixes from 16S rRNA. Color coding for the tmRNA is as in Fig. 2C. For clarity, EF-Tu is not shown.")
71
Fig. 4. Comparison of maps of tmRNA complexes with and without S1
Fig.4. Comparison of maps of tmRNA complexes with and without S1. Cryo-EM maps for the 70S•tRNA•EFTu• tmRNA•SmpB complex in the presence (A) and absence (B) of ribosomal protein S1. Different colors for the density attributable to EF-Tu•tmRNA•SmpB are used to allow a comparison in (C), where the purple semitransparent map from (B) is depicted together with the solid red from (A). In the absence of S1 (B) an additional connection is seen between PK1 and helix H5 of the tmRNA (D) that reveals the presence of the folded ORF.
 and absence (B) of ribosomal protein S1. Different colors for the density attributable to EF-Tu•tmRNA•SmpB are used to allow a comparison in (C), where the purple semitransparent map from (B) is depicted together with the solid red from (A). In the. absence of S1 (B) an additional connection is seen between PK1 and helix H5 of the tmRNA (D) that reveals. the presence of the folded ORF.")
73
Modificación de los rRNA y funcionalidad del ribosoma
Fig. 1. Major types of rRNA modification. (a) 2'- O-methylation (Nm). (b) Isomerization of uridine to pseudouridine (.), ‘the fifth nucleoside’. (c) Most other modifications involve base methylation at various positions (arrows). This includes .– a few are acetylated or otherwise modified at such positions [47].
 2 - O-methylation (Nm). (b) Isomerization of uridine to pseudouridine (.), ‘the fifth nucleoside’. (c) Most other modifications involve base methylation at various. positions (arrows). This includes .– a few are acetylated or otherwise modified at such positions [47].")
74
Fig. 2. Distribution of Escherichia colirRNA modifications in the ribosome. Nucleotides known to be modified in E. coliare highlighted in the E. colisecondary structure and in crystal structures of the small and large ribosomal subunits (SSU and LSU), derived for Thermus thermophilusand Haloarcula marismortui, respectively. (a) The E. colisecondary structure was adapted from the Gutell Laboratory Comparative RNA site ( There is a total of 35 modified nucleotides [48]), here divided into three groups and highlighted with different colours. The groupings include: (1) 11 pseudouridines and modified pseudouridines (., red triangles); (2) four 2'- O-methylations (Nm, green circles); and (3) ‘other’, consisting of 19 base methylations (mN, different types) plus one LSU dihydrouridine (orange squares). Their distributions are (SSU and LSU): ., 1/10; Nm, 1/3; ‘other’, 9/11. (b) SSU ( T. thermophilus; PDB entry 1fjf). The E. coliSSU contains 11 modified sites. Helix 44 is indicated in cyan. Morphological features (head, neck and body) are shown. (c) LSU ( H. marismortui; PDB entries 1ffk and 1ffz). The transition-state analogue is shown bound (magenta). The E. coliLSU contains 24 modified sites. Three .s in helix 69 are not shown because of disorder in parts of the current crystal structure [shaded areas in (a)]. Functional regions are indicated for each subunit. (b,c) The modified nucleotides are distinguished by showing full atomic volume (van der Waals radii), whereas a backbone representation is used for the rRNA (grey) and protein chains (blue for SSU, maroon for LSU), and a skeleton representation for unmodified nucleotides (grey).
![Fig. 2. Distribution of Escherichia colirRNA modifications in the ribosome. Nucleotides known to be modified in E. coliare highlighted in the E. colisecondary structure and in crystal structures of the small and large ribosomal subunits (SSU and LSU), derived for Thermus thermophilusand Haloarcula marismortui, respectively. (a) The E. colisecondary structure was adapted from the Gutell Laboratory Comparative RNA site ( There is a total of 35 modified nucleotides [48]), here divided into three groups and highlighted with different colours. The groupings include: (1) 11 pseudouridines and modified pseudouridines (., red triangles);](http://slideplayer.es/slide/4188460/13/images/74/Fig.+2.+Distribution+of+Escherichia+colirRNA+modifications+in+the+ribosome.+Nucleotides+known+to+be+modified+in+E.+coliare+highlighted+in+the+E.+colisecondary+structure+and+in+crystal+structures+of+the+small+and+large+ribosomal+subunits+%28SSU+and+LSU%29%2C+derived+for+Thermus+thermophilusand+Haloarcula+marismortui%2C+respectively.+%28a%29+The+E.+colisecondary+structure+was+adapted+from+the+Gutell+Laboratory+Comparative+RNA+site+%28++There+is+a+total+of+35+modified+nucleotides+++%5B48%5D%29%2C+here+divided+into+three+groups+and+highlighted+with+different+colours.+The+groupings+include%3A+%281%29+11+pseudouridines+and+modified+pseudouridines+%28.%2C+red+triangles%29%3B.jpg "(2) four 2 - O-methylations (Nm, green circles); and (3) ‘other’, consisting of 19 base methylations (mN, different types) plus one LSU dihydrouridine (orange squares). Their distributions are (SSU and LSU): ., 1/10; Nm, 1/3; ‘other’, 9/11. (b) SSU ( T. thermophilus; PDB entry 1fjf). The E. coliSSU contains 11 modified sites. Helix 44 is indicated in cyan. Morphological features (head, neck and body) are shown. (c) LSU ( H. marismortui; PDB entries 1ffk and 1ffz). The transition-state analogue is shown bound (magenta). The E. coliLSU contains 24 modified sites. Three .s in helix 69 are not shown because of disorder in parts of the current crystal structure [shaded areas in (a)]. Functional regions are indicated for each subunit. (b,c) The modified nucleotides are distinguished by showing full atomic volume (van der Waals radii), whereas a backbone representation is used for the rRNA (grey) and protein chains (blue for SSU, maroon for LSU), and a skeleton representation for unmodified nucleotides (grey).")
75
Fig. 2. Distribution of Escherichia colirRNA modifications in the ribosome. Nucleotides known to be modified in E. coliare highlighted in the E. colisecondary structure and in crystal structures of the small and large ribosomal subunits (SSU and LSU), derived for Thermus thermophilusand Haloarcula marismortui, respectively. (a) The E. colisecondary structure was adapted from the Gutell Laboratory Comparative RNA site ( There is a total of 35 modified nucleotides [48]), here divided into three groups and highlighted with different colours. The groupings include: (1) 11 pseudouridines and modified pseudouridines (., red triangles); (2) four 2'- O-methylations (Nm, green circles); and (3) ‘other’, consisting of 19 base methylations (mN, different types) plus one LSU dihydrouridine (orange squares). Their distributions are (SSU and LSU): ., 1/10; Nm, 1/3; ‘other’, 9/11. (b) SSU ( T. thermophilus; PDB entry 1fjf). The E. coliSSU contains 11 modified sites. Helix 44 is indicated in cyan. Morphological features (head, neck and body) are shown. (c) LSU ( H. marismortui; PDB entries 1ffk and 1ffz). The transition-state analogue is shown bound (magenta). The E. coliLSU contains 24 modified sites. Three .s in helix 69 are not shown because of disorder in parts of the current crystal structure [shaded areas in (a)]. Functional regions are indicated for each subunit. (b,c) The modified nucleotides are distinguished by showing full atomic volume (van der Waals radii), whereas a backbone representation is used for the rRNA (grey) and protein chains (blue for SSU, maroon for LSU), and a skeleton representation for unmodified nucleotides (grey).
![Fig. 2. Distribution of Escherichia colirRNA modifications in the ribosome. Nucleotides known to be modified in E. coliare highlighted in the E. colisecondary structure and in crystal structures of the small and large ribosomal subunits (SSU and LSU), derived for Thermus thermophilusand Haloarcula marismortui, respectively. (a) The E. colisecondary structure was adapted from the Gutell Laboratory Comparative RNA site ( There is a total of 35 modified nucleotides [48]), here divided into three groups and highlighted with different colours. The groupings include: (1) 11 pseudouridines and modified pseudouridines (., red triangles);](http://slideplayer.es/slide/4188460/13/images/75/Fig.+2.+Distribution+of+Escherichia+colirRNA+modifications+in+the+ribosome.+Nucleotides+known+to+be+modified+in+E.+coliare+highlighted+in+the+E.+colisecondary+structure+and+in+crystal+structures+of+the+small+and+large+ribosomal+subunits+%28SSU+and+LSU%29%2C+derived+for+Thermus+thermophilusand+Haloarcula+marismortui%2C+respectively.+%28a%29+The+E.+colisecondary+structure+was+adapted+from+the+Gutell+Laboratory+Comparative+RNA+site+%28++There+is+a+total+of+35+modified+nucleotides+++%5B48%5D%29%2C+here+divided+into+three+groups+and+highlighted+with+different+colours.+The+groupings+include%3A+%281%29+11+pseudouridines+and+modified+pseudouridines+%28.%2C+red+triangles%29%3B.jpg "(2) four 2 - O-methylations (Nm, green circles); and (3) ‘other’, consisting of 19 base methylations (mN, different types) plus one LSU dihydrouridine (orange squares). Their distributions are (SSU and LSU): ., 1/10; Nm, 1/3; ‘other’, 9/11. (b) SSU ( T. thermophilus; PDB entry 1fjf). The E. coliSSU contains 11 modified sites. Helix 44 is indicated in cyan. Morphological features (head, neck and body) are shown. (c) LSU ( H. marismortui; PDB entries 1ffk and 1ffz). The transition-state analogue is shown bound (magenta). The E. coliLSU contains 24 modified sites. Three .s in helix 69 are not shown because of disorder in parts of the current crystal structure [shaded areas in (a)]. Functional regions are indicated for each subunit. (b,c) The modified nucleotides are distinguished by showing full atomic volume (van der Waals radii), whereas a backbone representation is used for the rRNA (grey) and protein chains (blue for SSU, maroon for LSU), and a skeleton representation for unmodified nucleotides (grey).")
76
Fig. 3. Sites of yeast rRNA modification in the ribosome
Fig. 3. Sites of yeast rRNA modification in the ribosome. The rRNA of Saccharomyces cerevisiaeis estimated to contain 109 modifications, and nucleotide positions are known for 108 of these. Predicted sites are shown in the three-dimensional ribosomal subunits for all 44 pseudouridines (.), 54 (of 55) 2'- O-methylations (Nm) and 10 (of ~10) base methylations (mN). The distribution of these modifications within the small subunit (SSU) and large subunit (LSU) are as follows: .(red), 14/30; mN (orange), 3/7; Nm (green), 17/37. (a) Secondary structure maps showing modification sites for the 18S (left) and 25S–5.8S (right) rRNAs. RNA domains are identified with Roman numerals; shaded areas indicate substantial segments of RNA present in both the yeast and Haloarcula marismortui sequences but absent in the present LSU crystal structure. Blue dots mark modifications located in 18S rRNA segments with no equivalent in archaeal or eubacterial organisms, which are thus absent from the three-dimensional structure of the SSU. (b,c) Positions of yeast modifications in two views of the SSU. Helix 44 is shown in cyan. (d,e) Sites of yeast modification in two views of the LSU. (b–e) The colour scheme is the same as in (a) and modified nucleotides are distinguished by showing full atomic volume (van der Waals radii), whereas a backbone representation is used for the rRNA (grey) and protein chains (blue for SSU, maroon for LSU), and a skeleton representation for unmodified nucleotides (grey); in (e), the proteins are hidden and the RNA is de-emphasized.
, 54 (of 55) 2 - O-methylations (Nm) and 10 (of ~10) base methylations (mN). The distribution of these. modifications within the small subunit (SSU) and large subunit (LSU) are as follows: .(red), 14/30; mN (orange), 3/7; Nm (green), 17/37. (a) Secondary structure maps showing modification sites for the. 18S (left) and 25S–5.8S (right) rRNAs. RNA domains are identified with Roman numerals; shaded. areas indicate substantial segments of RNA present in both the yeast and Haloarcula marismortui. sequences but absent in the present LSU crystal structure. Blue dots mark modifications located in. 18S rRNA segments with no equivalent in archaeal or eubacterial organisms, which are thus absent. from the three-dimensional structure of the SSU. (b,c) Positions of yeast modifications in two views of. the SSU. Helix 44 is shown in cyan. (d,e) Sites of yeast modification in two views of the LSU. (b–e) The colour scheme is the same as in (a) and modified nucleotides are distinguished by showing. full atomic volume (van der Waals radii), whereas a backbone representation is used for the rRNA. (grey) and protein chains (blue for SSU, maroon for LSU), and a skeleton representation for. unmodified nucleotides (grey); in (e), the proteins are hidden and the RNA is de-emphasized.")
77
Fig. 3. Sites of yeast rRNA modification in the ribosome
Fig. 3. Sites of yeast rRNA modification in the ribosome. The rRNA of Saccharomyces cerevisiaeis estimated to contain 109 modifications, and nucleotide positions are known for 108 of these. Predicted sites are shown in the three-dimensional ribosomal subunits for all 44 pseudouridines (.), 54 (of 55) 2'- O-methylations (Nm) and 10 (of ~10) base methylations (mN). The distribution of these modifications within the small subunit (SSU) and large subunit (LSU) are as follows: .(red), 14/30; mN (orange), 3/7; Nm (green), 17/37. (a) Secondary structure maps showing modification sites for the 18S (left) and 25S–5.8S (right) rRNAs. RNA domains are identified with Roman numerals; shaded areas indicate substantial segments of RNA present in both the yeast and Haloarcula marismortui sequences but absent in the present LSU crystal structure. Blue dots mark modifications located in 18S rRNA segments with no equivalent in archaeal or eubacterial organisms, which are thus absent from the three-dimensional structure of the SSU. (b,c) Positions of yeast modifications in two views of the SSU. Helix 44 is shown in cyan. (d,e) Sites of yeast modification in two views of the LSU. (b–e) The colour scheme is the same as in (a) and modified nucleotides are distinguished by showing full atomic volume (van der Waals radii), whereas a backbone representation is used for the rRNA (grey) and protein chains (blue for SSU, maroon for LSU), and a skeleton representation for unmodified nucleotides (grey); in (e), the proteins are hidden and the RNA is de-emphasized.
, 54 (of 55) 2 - O-methylations (Nm) and 10 (of ~10) base methylations (mN). The distribution of these. modifications within the small subunit (SSU) and large subunit (LSU) are as follows: .(red), 14/30; mN (orange), 3/7; Nm (green), 17/37. (a) Secondary structure maps showing modification sites for the. 18S (left) and 25S–5.8S (right) rRNAs. RNA domains are identified with Roman numerals; shaded. areas indicate substantial segments of RNA present in both the yeast and Haloarcula marismortui. sequences but absent in the present LSU crystal structure. Blue dots mark modifications located in. 18S rRNA segments with no equivalent in archaeal or eubacterial organisms, which are thus absent. from the three-dimensional structure of the SSU. (b,c) Positions of yeast modifications in two views of. the SSU. Helix 44 is shown in cyan. (d,e) Sites of yeast modification in two views of the LSU. (b–e) The colour scheme is the same as in (a) and modified nucleotides are distinguished by showing. full atomic volume (van der Waals radii), whereas a backbone representation is used for the rRNA. (grey) and protein chains (blue for SSU, maroon for LSU), and a skeleton representation for. unmodified nucleotides (grey); in (e), the proteins are hidden and the RNA is de-emphasized.")
78
Fig. 3. Sites of yeast rRNA modification in the ribosome
Fig. 3. Sites of yeast rRNA modification in the ribosome. The rRNA of Saccharomyces cerevisiaeis estimated to contain 109 modifications, and nucleotide positions are known for 108 of these. Predicted sites are shown in the three-dimensional ribosomal subunits for all 44 pseudouridines (.), 54 (of 55) 2'- O-methylations (Nm) and 10 (of ~10) base methylations (mN). The distribution of these modifications within the small subunit (SSU) and large subunit (LSU) are as follows: .(red), 14/30; mN (orange), 3/7; Nm (green), 17/37. (a) Secondary structure maps showing modification sites for the 18S (left) and 25S–5.8S (right) rRNAs. RNA domains are identified with Roman numerals; shaded areas indicate substantial segments of RNA present in both the yeast and Haloarcula marismortui sequences but absent in the present LSU crystal structure. Blue dots mark modifications located in 18S rRNA segments with no equivalent in archaeal or eubacterial organisms, which are thus absent from the three-dimensional structure of the SSU. (b,c) Positions of yeast modifications in two views of the SSU. Helix 44 is shown in cyan. (d,e) Sites of yeast modification in two views of the LSU. (b–e) The colour scheme is the same as in (a) and modified nucleotides are distinguished by showing full atomic volume (van der Waals radii), whereas a backbone representation is used for the rRNA (grey) and protein chains (blue for SSU, maroon for LSU), and a skeleton representation for unmodified nucleotides (grey); in (e), the proteins are hidden and the RNA is de-emphasized.
, 54 (of 55) 2 - O-methylations (Nm) and 10 (of ~10) base methylations (mN). The distribution of these. modifications within the small subunit (SSU) and large subunit (LSU) are as follows: .(red), 14/30; mN (orange), 3/7; Nm (green), 17/37. (a) Secondary structure maps showing modification sites for the. 18S (left) and 25S–5.8S (right) rRNAs. RNA domains are identified with Roman numerals; shaded. areas indicate substantial segments of RNA present in both the yeast and Haloarcula marismortui. sequences but absent in the present LSU crystal structure. Blue dots mark modifications located in. 18S rRNA segments with no equivalent in archaeal or eubacterial organisms, which are thus absent. from the three-dimensional structure of the SSU. (b,c) Positions of yeast modifications in two views of. the SSU. Helix 44 is shown in cyan. (d,e) Sites of yeast modification in two views of the LSU. (b–e) The colour scheme is the same as in (a) and modified nucleotides are distinguished by showing. full atomic volume (van der Waals radii), whereas a backbone representation is used for the rRNA. (grey) and protein chains (blue for SSU, maroon for LSU), and a skeleton representation for. unmodified nucleotides (grey); in (e), the proteins are hidden and the RNA is de-emphasized.")
79
Fig. 4. Pattern of yeast nucleotide modification in the large subunit (LSU) active site region of the peptidyl transferase centre (PTC). (a) The secondary structure of the active-site region of yeast domain V [17], showing six pseudouridines (.; red), seven 2'- O-methylations (Nm; green) and one base methylation (mN; orange). (b) Predicted locations of yeast ., Nm and mN nucleotides [colour coded as in (a)] in the three-dimensional structure of the active site. The transition-state analogue at the reaction centre is shown in magenta. (c,d) Base pairings of modified nucleotides involved in tRNA binding through the CCA-acceptor end, derived from the three-dimensional structures (PDB entries 1ffz and 1fg0). (c) Pairing predicted to occur at the A site, between C75 in tRNA and Gm2918 in yeast rRNA (G2588 in Haloarcula marismortuiand G2553 in Escherichia coli). (d) Pairing at the P site between C75 in tRNA and a universally conserved Gm nucleotide [Gm2618 in yeast, G2284 in H. marismortui(in which methylation has not been examined) and Gm2251 in E. coli].
. (a) The secondary structure of the active-site region of yeast domain. V [17], showing six pseudouridines (.; red), seven 2 - O-methylations (Nm; green) and one base. methylation (mN; orange). (b) Predicted locations of yeast ., Nm and mN nucleotides [colour coded. as in (a)] in the three-dimensional structure of the active site. The transition-state analogue at the. reaction centre is shown in magenta. (c,d) Base pairings of modified nucleotides involved in tRNA. binding through the CCA-acceptor end, derived from the three-dimensional structures (PDB entries. 1ffz and 1fg0). (c) Pairing predicted to occur at the A site, between C75 in tRNA and Gm2918 in yeast. rRNA (G2588 in Haloarcula marismortuiand G2553 in Escherichia coli). (d) Pairing at the P site. between C75 in tRNA and a universally conserved Gm nucleotide [Gm2618 in yeast, G2284 in. H. marismortui(in which methylation has not been examined) and Gm2251 in E. coli].")
86
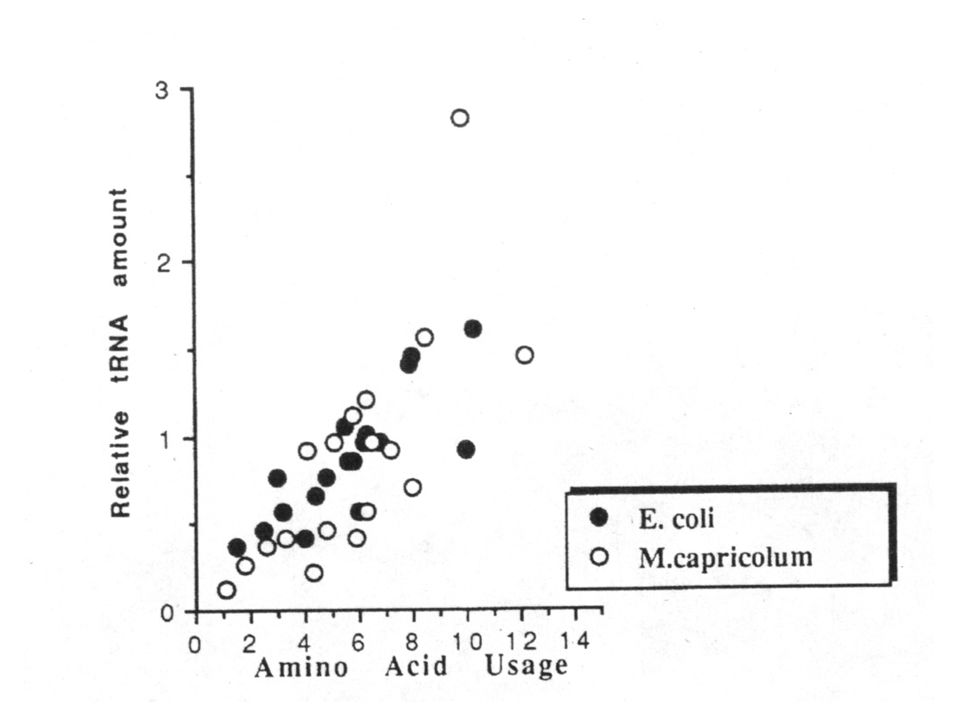
Muchos de los nucleótidos en posiciones invariantes son importantes en la estructura terciaria del tRNA. Por ejemplo, por cristalografía de rayos X se sabe que la interacción entre nucleótidos del brazo D y del brazo TpsiC dan la forma de L característica de la estructura terciaria. Las interacciones que mantienen esta estructura son básicamente puentes de H entre residuos no apareados en la estructura secundaria. Es importante que el brazo con el aminoácido se encuentre lo más lejos posible del brazo anticodón, de este modo el aminoácido está en contacto con la peptidiltransferasa en la subunidad grande del ribosoma, mientras que el brazo anticodón está cerca del ARNm en la subunidad pequeña del ribosoma.

Presentaciones similares






























>")




