Descargar la presentación
La descarga está en progreso. Por favor, espere

1
SUPERCLASE GNATHOSTOMATA

4
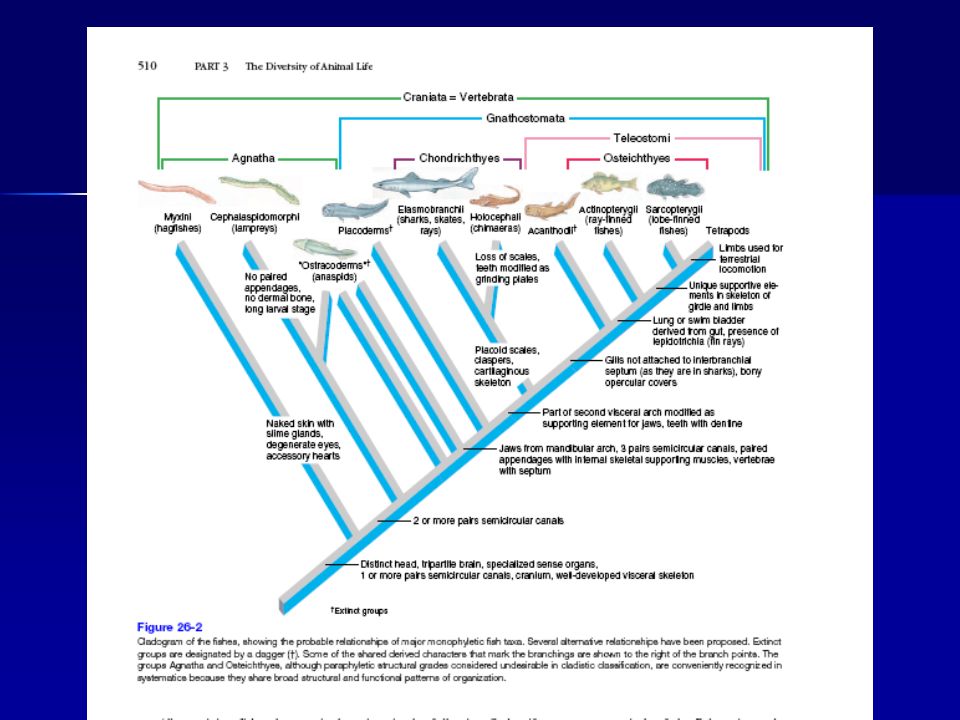
Fish Clades Gnathostomata Vertebrata “Agnathans” Chondrichthes
Osteichthyes “Cyclostomes” Cephalochordata Elasmobranchii Actinopterygii Sarcopterygii Conodonts Placoderms Holocephali Acanthodii Hagfishes Lampreys “Ostracoderms”

5
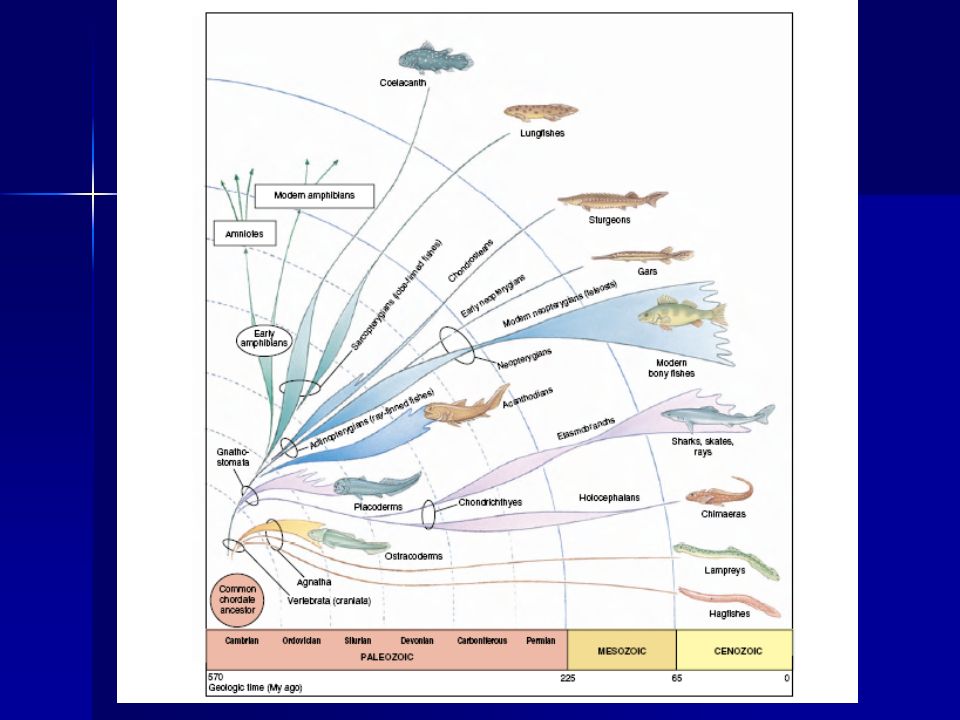
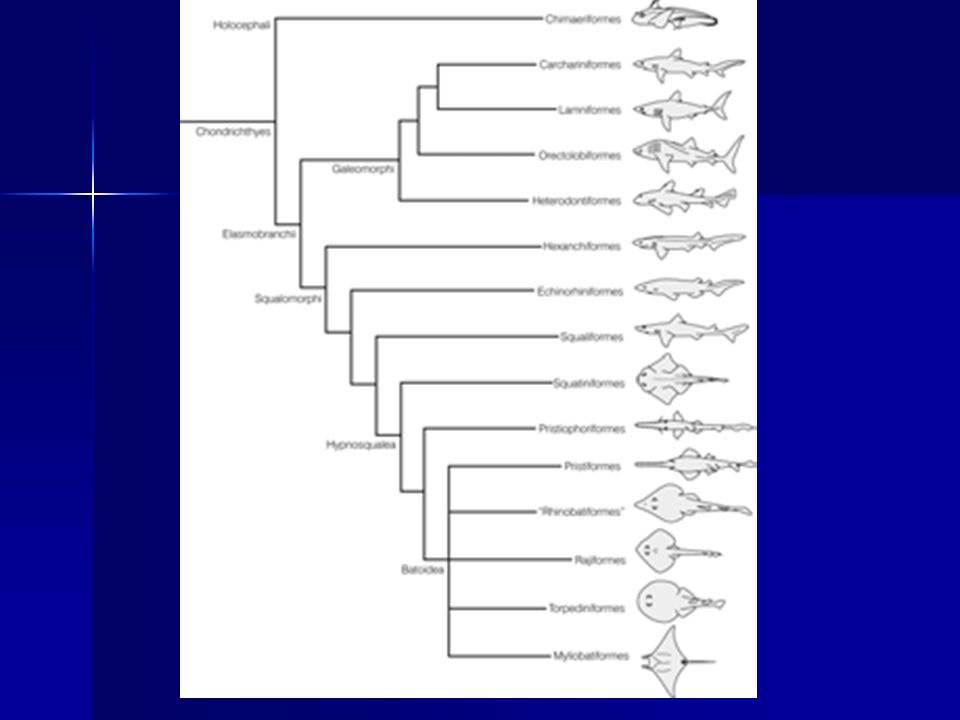
Filogenia de los peces

6
SUPERCLASE GNATHOSTOMATA
CARACTERÍSTICAS: La presencia de mandíbulas generalmente provista de dientes, en que la mandíbula superior está firmemente unida al neurocráneo o que la suspensión son más o menos libre. La presencia de dientes es un carácter primario que se conserva en todas las clases de vertebrados, sin embargo, estos dientes pueden estar ausentes o modificados como manifestaciones secundarias

7
Las mandíbulas en general, partiendo de las estructuras tan particulares de los miembros de la superclase, presentan transformaciones que tienden primero hacia la sustitución de los cartílagos fundamentales el pterigocuadrado y el de Meckel, por huesos de origen dérmico y posteriormente, hacia la disminución en el número de elementos esqueléticos hasta que en los mamíferos la mandíbula inferior está formado sólo por un hueso, cuyo origen se atribuye al tercero de los arcos viscerales, puesto que los anteriores desaparecen y los posteriores quedan libres, con funciones branquiales en los animales con respiración acuática y como vestigios o ausentes en los vertebrados pulmonados.

8
Todos los mandibulados, presentan como carácter primario, miembros pares que son los pterigios en los vertebrados acuáticos y quiridios en los terrestres. En ambos casos, se presentan formas que carecen de uno o de ambos pares o los tienen modificados para el vuelo u otra función diferente a la peculiar de esas estructuras anatómicas. Los orificios nasales son siempre dos, aunque secundariamente puede dividirse cada una y dar apariencia de duplicación. En la mayoría con respiración branquial, las fosas nasales no tienen comunicación con la parte inicial del aparato digestivo, pero en los terrestres, los anfibios, los dipnoos y los crosopterigios, hay conductos entre las fosas nasales y la cavidad bucal, llamados coanas (coanados).
.")
9
Carecen de bolsas branquiales, de endostilo en cualquiera de sus etapas ontogénicas o de algún otro aparato para la concentración de alimento como el mencionado. Oído interno con tres canales semicirculares Lo más común es que la notocorda sea sustituida por vértebras bien definidas y que el aparato digestivo presente órganos más especializados. Los representantes fósiles, los placodermos, presentan una coraza formada por placas.

10
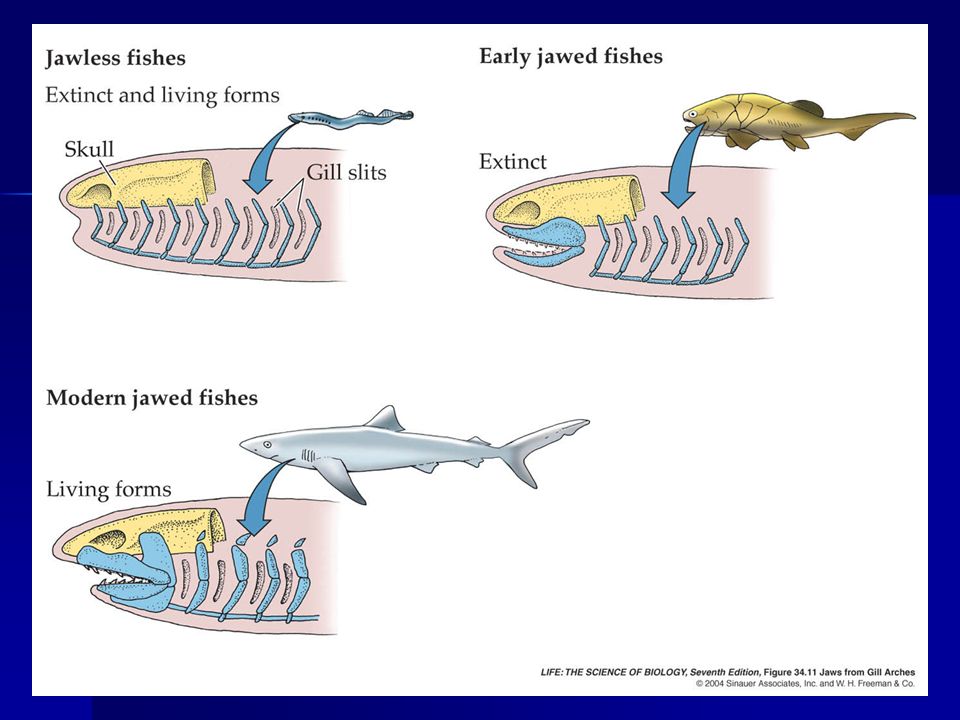
Evolución de las mandibulas

11
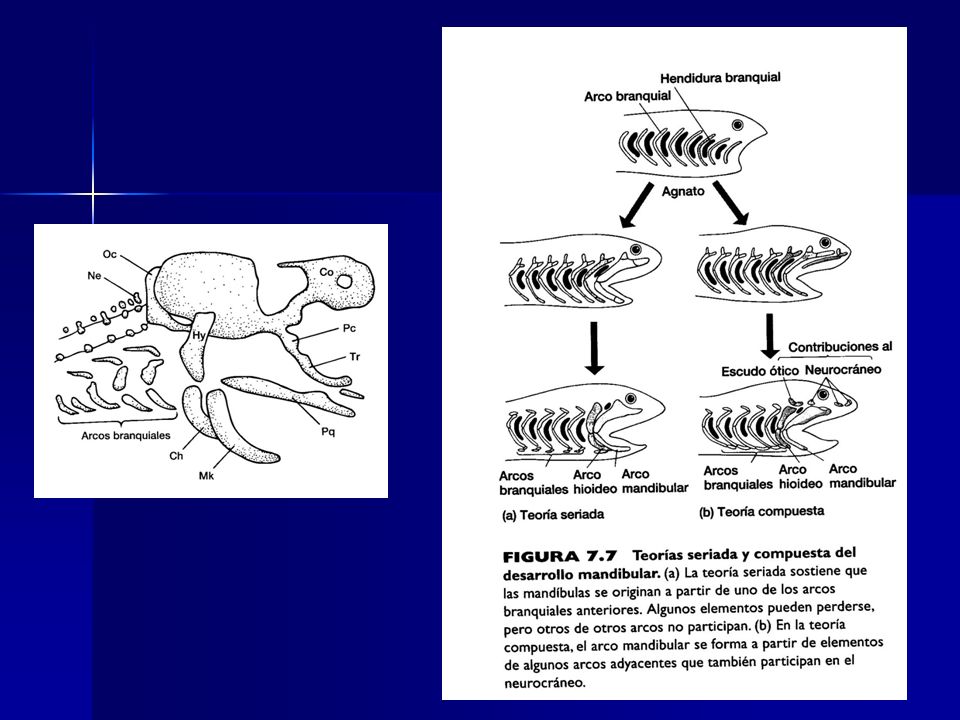
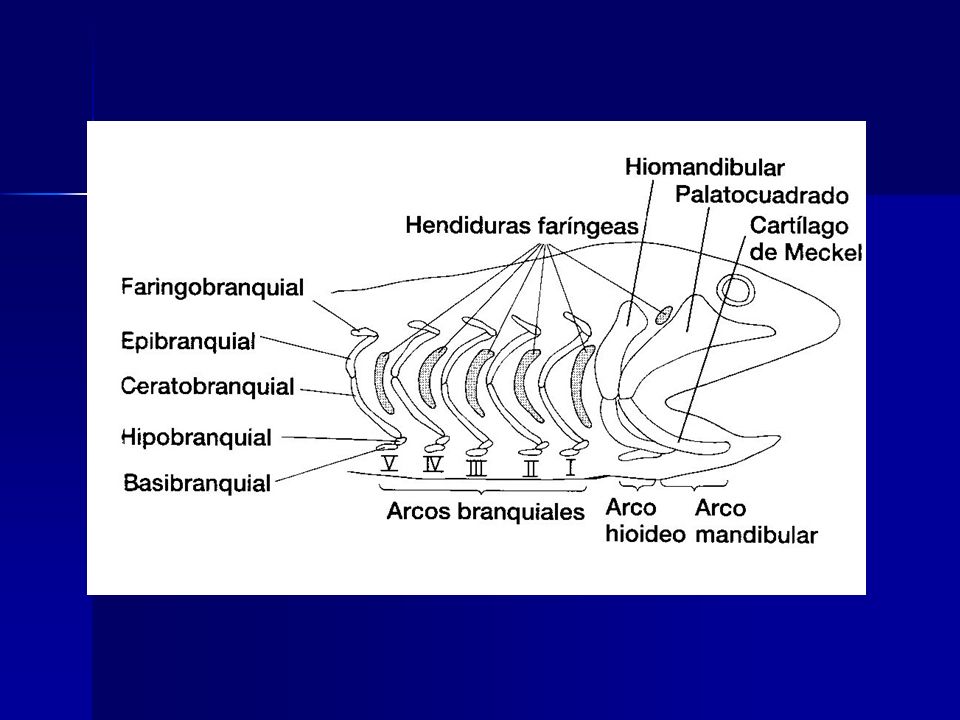
Evolución de la mandíbula
Arco branquial superior Arco branquial inferior Mandibula Superior : cartílago palatocuadrado Mandibula Inferior: cartílago de Meckel

12
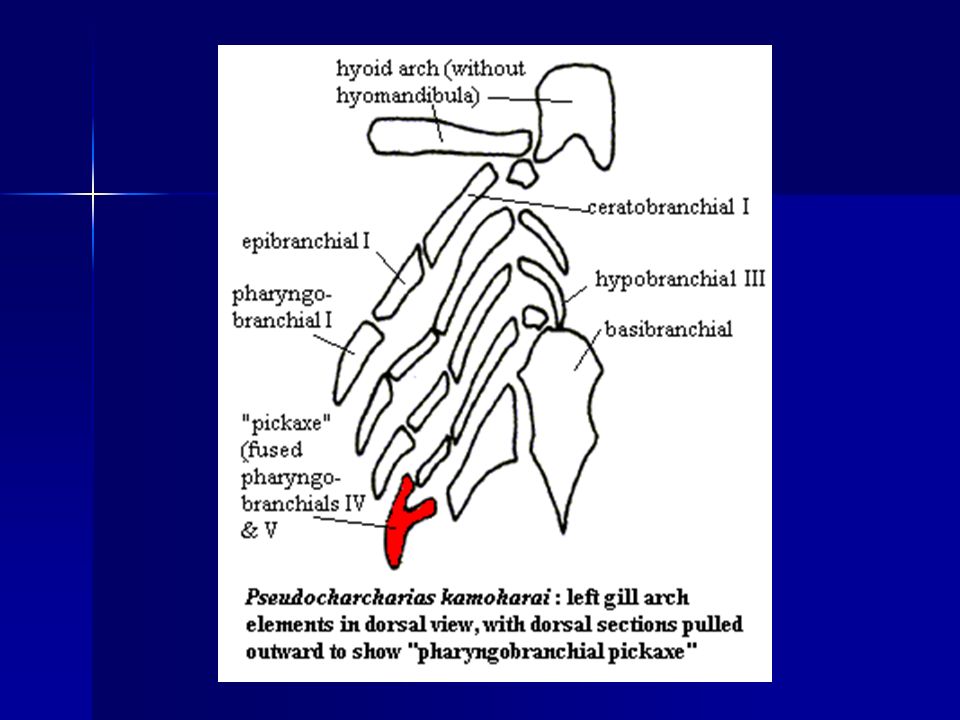
Evolución de la mandíbula

17
Mandibular arch - 1st branchial arch
Mandibulas condrocráneo Mandibular arch - 1st branchial arch modified as jaws Remaining branchial arches support gills

18
Hyoid Arch: Branchial Arches: Jaws: 17 - palatoquadrate cartilage 9 - hyomandibular 6 - epibranchials 5 - ceratohyal 11 - Meckle’s cartilage 6 - ceratobranchials

21
The Evolution of Jaws From pharyngeal support elements
Jaws and branchial arches are derived from embryonic neural crest cells Enlargement of the first branchial arch (mandibular arch) was probably in association with the need for greater/stronger pumping of water over gills Later, became modified as jaws
 was probably in association with the need for greater/stronger pumping of water over gills. Later, became modified as jaws.")
22
Advantages of Jaws Ability to grasp prey
Teeth bring the ability to chew and grind/tear prey Pick up objects for nest building Grasp a mate during courtship Grab young during parental care

24
Representantes fósiles:
SUPERCLASE GNATHOSTOMATA Representantes fósiles: Espinosos y placodermos. Clase Acanthodii “espinosos” Clase Pterichthyes Clase Coccostei

25
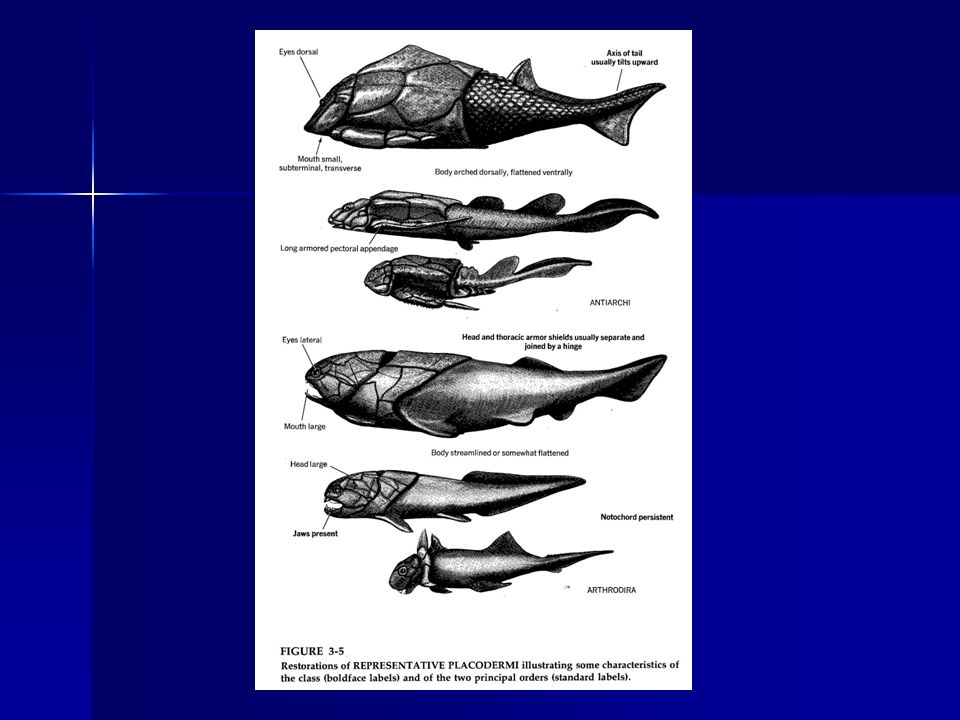
PLACODERMI (Placodermos)
Las principales características son: 1. Gruesas placas óseas cubrían la mitad anterior o un tercio del cuerpo. La mitad posterior estaba desnuda o cubierta con pequeñas escamas, permitiendo así la movilidad.

26
Class Placodermi

27
. 2. Las escamas y placas eran de tipo cosmoidea, constituidas por un estrato profundo de densas láminas óseas, un estrato medio del hueso vascularizado y una capa externa de material similar a la dentina, la cosmina, que en 2 los placodermos más evolucionados formaba dentículos dermales semejantes a dientes. Se cree que la retención de los dentículos y la pérdida del resto de placas y escamas ha sido el origen del diente de los tiburones y formas afines, sí es que éstos descienden de los placodermos. 3. Presentan unos dientes falsos que se disponen sobre los huesos dermales que cubrian las mandíbulas cartilaginosas proporcionando una superficie afilada y cortante y un borde aserrado desgarrante, muy efectivo en la depredación, pero no renovable por reposición como a menudo ocurre en el diente. En algunos, estas “placas dentales” formaban una superficie sólida, machacadora, útil en la depredación de moluscos.

28
†Clase Placodermi

29
All Long Extinct †Class Placodermi

30
†Clase Placodermi Rhenaniformes

31
†Clase Placodermi Antiarchiformes

32
mandíbulas+ aletas pares bien desarrolladas y cinturas

33
4. Una sutura transversal separaba el escudo de la cabeza del tronco, permitiendo que la cabeza pudiera inclinarse hacia arriba elevando la mandíbula superior (que estaba unida sólidamente al cráneo mediante huesos dermales). 5. Aletas pectorales y pelvianas bien desarrolladas. Las primeras presentaban una considerable diversificación adaptativa.

34
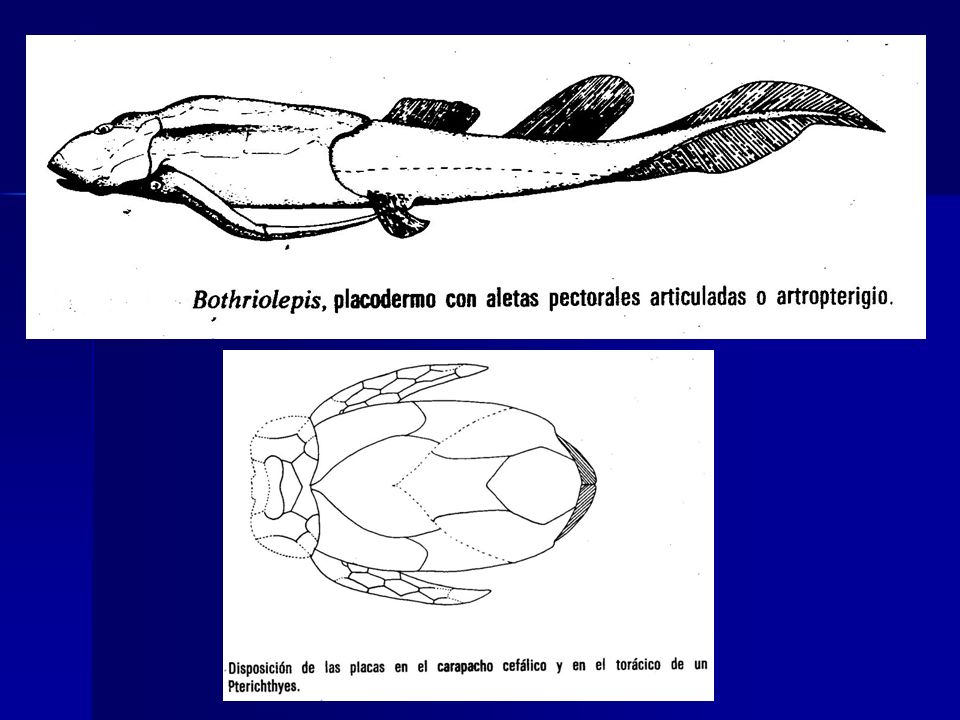
En algunos placodermos las aletas pectorales llagaron a estar revestidas de tubos óseos. Formando un exoesqueleto articulado (artropterigios). En otros, como las rayas, aumentaron de tamaño y se ensancharon (aunque menos flexibles). En ciertos grupos las aletas pelvianas llegaron a ser sexualmente dimórficas, indicando su empleo durante la cópula (como el “órgano de la cópula” de los elasmobranquios).
. En otros, como las rayas, aumentaron de tamaño y se ensancharon (aunque menos flexibles). En ciertos grupos las aletas pelvianas llegaron a ser sexualmente dimórficas, indicando su empleo durante la cópula (como el órgano de la cópula de los elasmobranquios)..")
35
Placodermos Coccosteus Arctolepis Lunaspis Phyllolepis Rhamphodopsis
Gemuendina Coccosteus Lunaspis Rhamphodopsis Bothriolepis

36
Placodermos

39
Placodermi “Plate skinned” fishes
With much armor plating, but reduced compared to the Ostracoderms Bony skeleton Paired pelvic and pectoral fins Fins allow for greater swimming ability In order to capture prey In order to escape being preyed upon Prevention of roll, pitch, and yaw More stream-lined body shape Better swimming ability

40
Gnathostomata: Acanthodii
Climatius

41
Acanthodi

42
Acanthodi

44
Placoderm “improvements”
Possessed “lungs” Some sort of accessory structure off the esophagus Lived in freshwater Modified dermal bones which served as “teeth” Could be replaced if broken May have been an evolutionary “dead-end” May have led to the Chondrichthyes May have led to the Osteichthyes Taxonomic affinities are uncertain Extinct by the middle of Carboniferous period

45
CLASE ELASMOBRANCHII

48
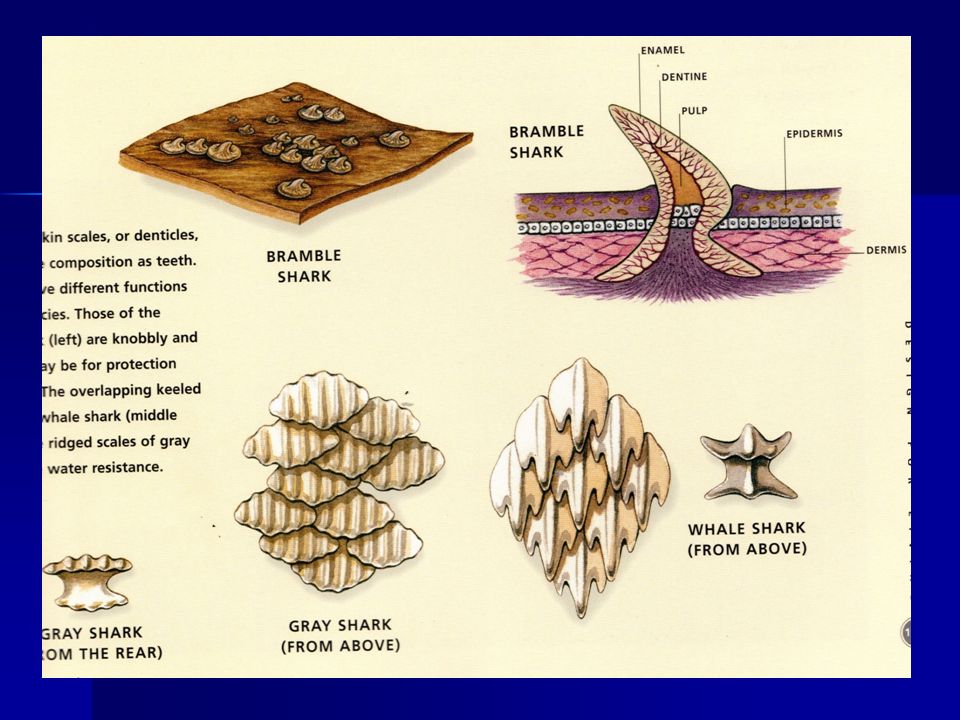
CLASE ELASMOBRANCHII Los elasmobranquios difieren de los peces con espinas en las siguientes características: Carecen de tejido óseo y el esqueleto está formado por cartílago que puede ser calcificado por endurecimiento y reducirse por flotabilidad. Piel con escamas placoideas, como diminutos dientes curvados que sobresalen a través de la epidermis y reducen la resistencia de avance al nadar La piel es muy dura, tan estimada como el cuero, o puede tener dentículos como una zapa para pulir. Los espiráculos y cinco de los siete pares de hendiduras branquiales están descubiertos, es decir, no están protegidos por un opérculo

50
Escamas en los peces

51
Peces cartilaginosos

52
Las aletas pares tienen una base ancha y no pueden ser maniobradas lo suficiente para poder frenar, permanecer inmóvil o retroceder. La cintura escapular está constituida por barras unidas ventralmente, y la cintura pectoral no está articulado con el cráneo. El aplanamiento dorsoventral afecta a la cabeza y a veces también al tronco. En los tiburones, el lóbulo dorsal de la aleta caudal es más larga: heterocerca, dentro del cual se prolonga la columna vertebral. Esto hace que cuando nadan la cola sobresalga y la cabeza se incline hacia abajo, compensándose el efecto plano ascendente de la aplanada cabeza. La boca y el par de sacos nasales se sitúan en la superficie ventral de la cabeza

53
No poseen pulmones ni vejiga gaseosa.
Los dientes se encuentran sobre las mandíbulas y son constantemente reemplazados por dientes nuevos que ocupan la posición en las bandas de tejido conectivo. Los elasmobranquios (“branquias planas”) se denominan así debido a que sus filamentos branquiales se originan a partir de las paredes de unos tabiques planos (septos interbranquiales) que separan las cámaras branquiales adyacentes. La única glándula rectal de la sal, en posición anterior a la cloaca, secreta los excesos de sales de la corrientes sanguínea al intestino
 se denominan así debido a que sus filamentos branquiales se originan a partir de las paredes de unos tabiques planos (septos interbranquiales) que separan las cámaras branquiales adyacentes. La única glándula rectal de la sal, en posición anterior a la cloaca, secreta los excesos de sales de la corrientes sanguínea al intestino.")
54
14. A excepción de las rayas de agua dulce, los elasmobranquios efectúan la osmorregulación reteniendo urea, almacenando concentraciones de este desecho nitrogenado tóxico que afectarían a cualquier otro animal de sangre fría. En efecto, sus líquidos corporales contiene una elevada cantidad de solutos (3,9 por 100), siendo levemente hiperosmótica con respecto al agua de mar, de manera que producen abundante orina diluida ( los tiburones típicos producen una cantidad de orina a razón de 1 ml/kg/h mientras que los peces óseos marinos sólo 0.3 ml/kg/h.). Este método de osmorregulación limita a la mayoría de los elasmobranquios a vivir en el mar. Si embargo, el cosmopolita tiburón “cabeza de toro” se introduce en los ríos aguas arriba en cientos de kilómetros, con el contenido de urea y NaCl de su cuerpo enormemente disminuido durante estas incursiones. Esta especie incluso sobrevive en las aguas del lago Nicaragua, un lago interior.
, siendo levemente hiperosmótica con respecto al agua de mar, de manera que producen abundante orina diluida ( los tiburones típicos producen una cantidad de orina a razón de 1 ml/kg/h mientras que los peces óseos marinos sólo 0.3 ml/kg/h.). Este método de osmorregulación limita a la mayoría de los elasmobranquios a vivir en el mar. Si embargo, el cosmopolita tiburón cabeza de toro se introduce en los ríos aguas arriba en cientos de kilómetros, con el contenido de urea y NaCl de su cuerpo enormemente disminuido durante estas incursiones. Esta especie incluso sobrevive en las aguas del lago Nicaragua, un lago interior..")
56
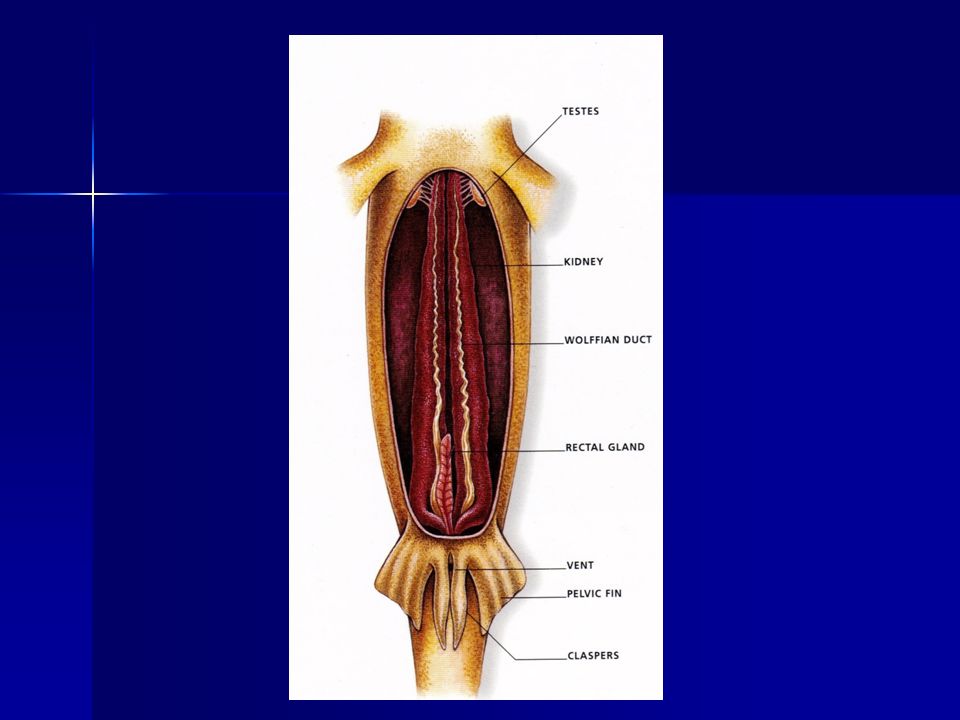
Claspers

57
15. Todos poseen un corazón de dos cámaras principales y dos accesorias (Aurícula, ventrículo, seno venoso y cono arterial). 16. Los órganos copuladores de los machos (clasper, pterigopodio), modificaciones de las aletas pélvicas, pueden flexionarse formando ángulos rectos con respecto al cuerpo mientras el semen fluye a lo largo de un canal en la superficie dorsal del órgano
, modificaciones de las aletas pélvicas, pueden flexionarse formando ángulos rectos con respecto al cuerpo mientras el semen fluye a lo largo de un canal en la superficie dorsal del órgano.")
61
17. Lo huevos tienen longitud de unos 1º cm, y en las especies ovíparas y ovovivíparas poseen una gran cantidad de vitelo. En los ovíparos, los huevos están protegidos por una fuerte cáscara queratinosa que es secretada por la región anterior del oviducto. En los ovovivíparos, los huevos permanecen en el útero hasta la eclosión, y la incubación puede durara hasta dos años en algunos tiburones. Algunos tiburones son realmente vivíparos: los huevos de estas especies contienen substancialmente menos cantidad de vitelo, y ya que el peso seco de un huevo es menor que el de una cría recién nacida, es evidente que el feto recibe alimento a partir de la pared uterina a través de una placenta simple. Al nacer, las crías de los elasmobranquios son muy largas si se les compara con las crías de los peces óseos, y están dotadas para defenderse de sus depredadores.

62
OTRAS CARACTERISTICAS DE LOS ELASMOBRANQUIOS
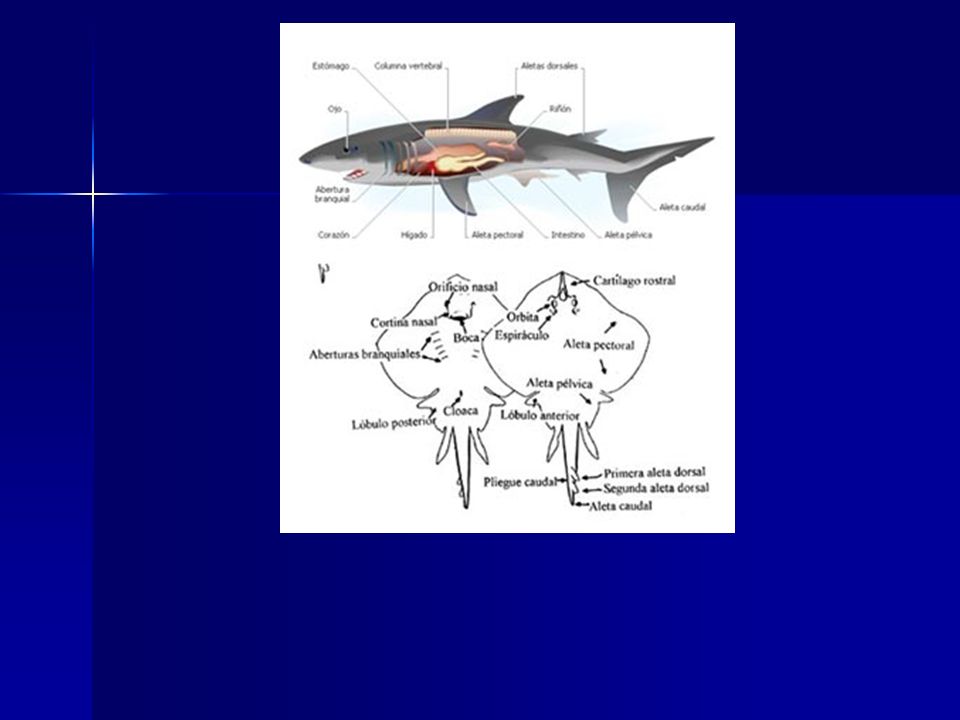
1. El tubo digestivo está formado por una boca, una cavidad bucal, una faringe, un esófago corto, un estómago en forma de J con un esfínter pilórico cerrando su extremo anterior, un intestino con válvula espiral que aumenta la superficie funcional del intestino, una cloaca y un ano. Los conductos del páncreas y del hígado desembocan independientemente justo debajo del estómago. El hígado, bilobulado, es muy rico en aceites (siendo una de las principales fuentes de vitaminas A y D), constituye alrededor del 20 % del peso total del cuerpo y proporciona flotabilidad.
, constituye alrededor del 20 % del peso total del cuerpo y proporciona flotabilidad.")
63
2. Como en los peces óseos, las branquias están destinadas al intercambio gaseoso. La corriente de agua pasa por los filamentos en dirección opuesta a la corriente sanguínea en los capilares branquiales, debido a que los flujos se mueven en direcciones opuestas, las gradientes de concentración de los gases diluidos no se igualarán, como sería en el caso de que ambas corrientes fluyeran en la misma dirección.

64
Ciertos tiburones ( por ejemplo, el gran tiburón blanco), así como algunas especies de peces óseos ( por ejemplo el atún) son de sangre caliente. El calor generado en el músculo rojo profundo es retenido en el cuerpo en lugar de perderse a través de las branquias, debido a la contracorriente de cambio de calor entre la sangre que sale y entra en los vasos sanguíneos en una rete mirabile. Así, sí el agua está a una temperatura de 21°C, los tejidos internos tendrán una temperatura de 27°C. Mientras tiburones y rayas (junto con algunos peces óseos) localizan a sus presas detectando pequeñísimas cantidades de bioenergía que emana del cualquier cuerpo vivo. Los electrorreceptores de los elasmoibranquios, está localizados en unas depresiones cutáneas (ampollas de Lorenzini) situados en el hocico. Estas, incluso pueden ayudar en la orientación captando la diferencia de voltaje asociadas a las corrientes oceánicas o captando el campo gravitacional terrestre o ambos.
 localizan a sus presas detectando pequeñísimas cantidades de bioenergía que emana del cualquier cuerpo vivo. Los electrorreceptores de los elasmoibranquios, está localizados en unas depresiones cutáneas (ampollas de Lorenzini) situados en el hocico. Estas, incluso pueden ayudar en la orientación captando la diferencia de voltaje asociadas a las corrientes oceánicas o captando el campo gravitacional terrestre o ambos.")
65
AMPOLLAS DE LORENZINI Muchos tiburones poseen numerosos poros superficiales, sobre todo en la parte cefálica, que esconden una complicada estructura sensorial: están conectados pro medio de canales (bien aislado y llenos de una sustancia conductora de la electricidad) a una pequeña cámara interior (la ampolla) tapizada de células pilíferas y electrorreceptoras. Las ampollas de Lorenzini suelen estar agrupadas y constituyen el sistema electrosensorial -el sentido más mistorioso- capaz de detectar los campos eléctricos de bajísima intensidad que poseen los seres vivos

66
5. Presentan sexos separados y parece que no existe inversión de sexo
5. Presentan sexos separados y parece que no existe inversión de sexo. Las gónadas pares están conectadas a un par de conductos genitales. Como todos los anamniotas gnatostomos, el esperma se transporta a través del conducto del riñón opistonéfrico. En las hembras, un conducto separado, el oviducto tubular, comienza como un embudo ciliado cerca de los ovarios, y en las especies vivíparas se ensancha en la región posterior para formar un útero que desemboca en la cloaca.

67
Invasión de aguas dulces
Hace 400 millones de años los peces con quijadas empezaron a invadir los estuarios y las aguas dulces Necesitan desarrollar una regulación osmótica efectiva para regular la entrada de agua y la pérdida de sales

68
Soluciones al problema
La pared del cuerpo es bastante impermeable al agua El agua que entra al cuerpo pasa a los riñones que producen una orina diluida Las agallas tienen unas células especiales que absorben sales (iones de Na y Cl, por transporte activo) del agua y los pasan a la sangre
 del agua y los pasan a la sangre.")
69
Regreso de los peces al mar
A lo largo de millones de años los peces de agua dulce establecieron su concentración de iones en los fluidos del cuerpo equivalente a 1/3 parte a la del agua salada Cuando regresaron al mar tuvieron problemas ya que sus fluidos tenían una concentración de sales menor al del agua salada (hipo-osmóticos) Problema: ganan sal y pierden agua
 Problema: ganan sal y pierden agua.")
70
Soluciones al problema
Los peces beben agua de mar para adquirir agua y la absorben en el intestino El NaCl va a las agallas (por la sangre) donde células especializadas lo tranportan de nuevo al mar Los demás iones salen con las heces o por los riñones Sólo toman la cantidad necesaria de agua para sustituir la perdida.
 donde células especializadas lo tranportan de nuevo al mar. Los demás iones salen con las heces o por los riñones. Sólo toman la cantidad necesaria de agua para sustituir la perdida.")
71
Placodermos Dinichthys chasing Cladoselache

72
Elasmobranchii

Presentaciones similares






















>")



