Descargar la presentación
La descarga está en progreso. Por favor, espere

1
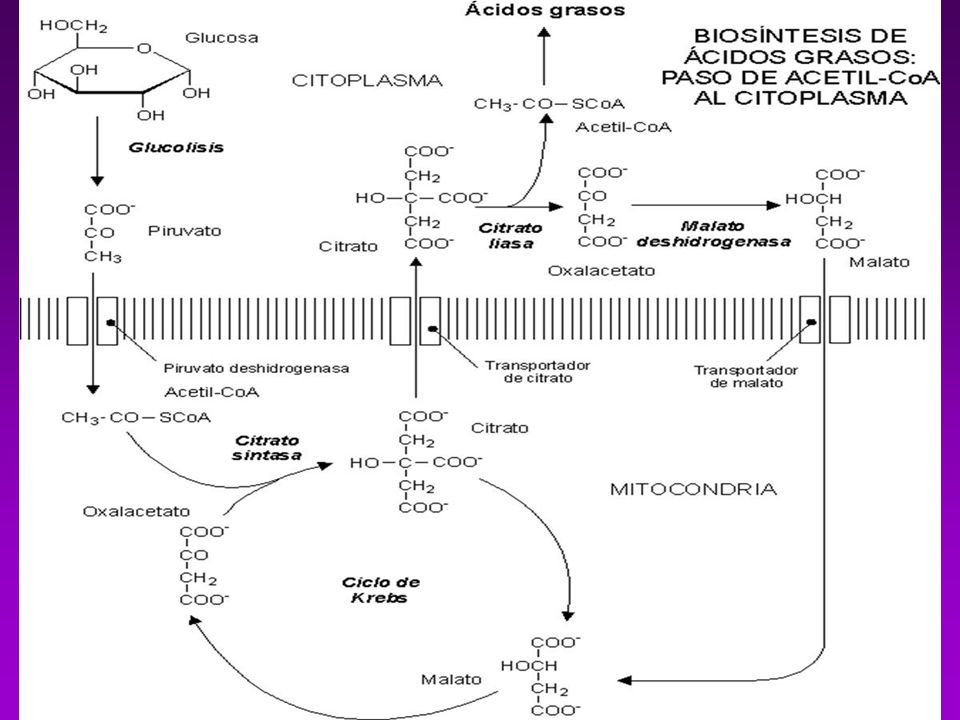
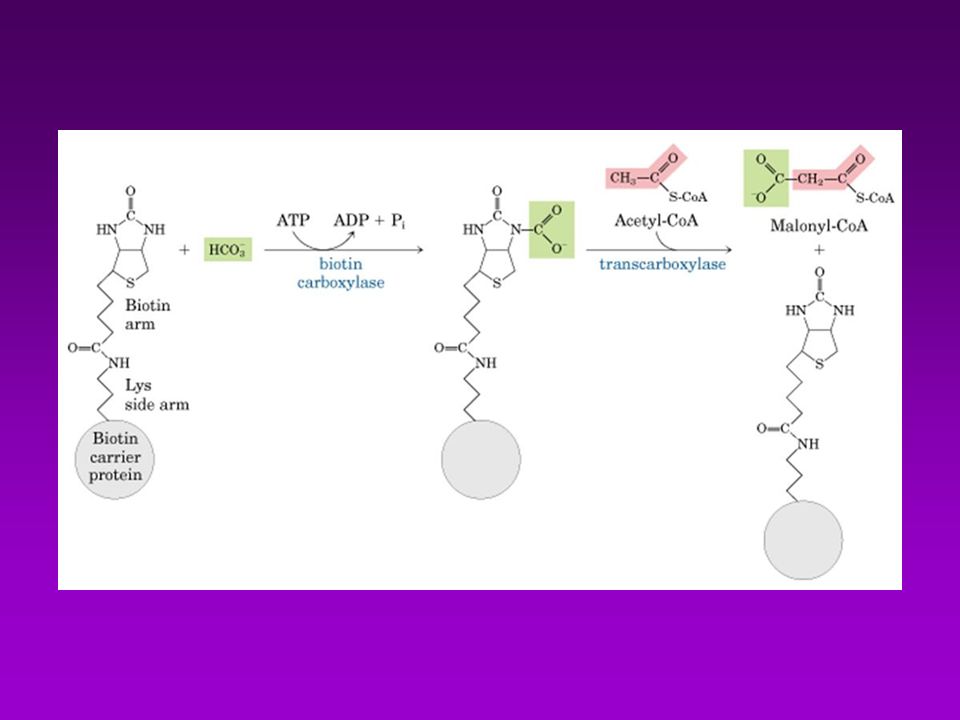
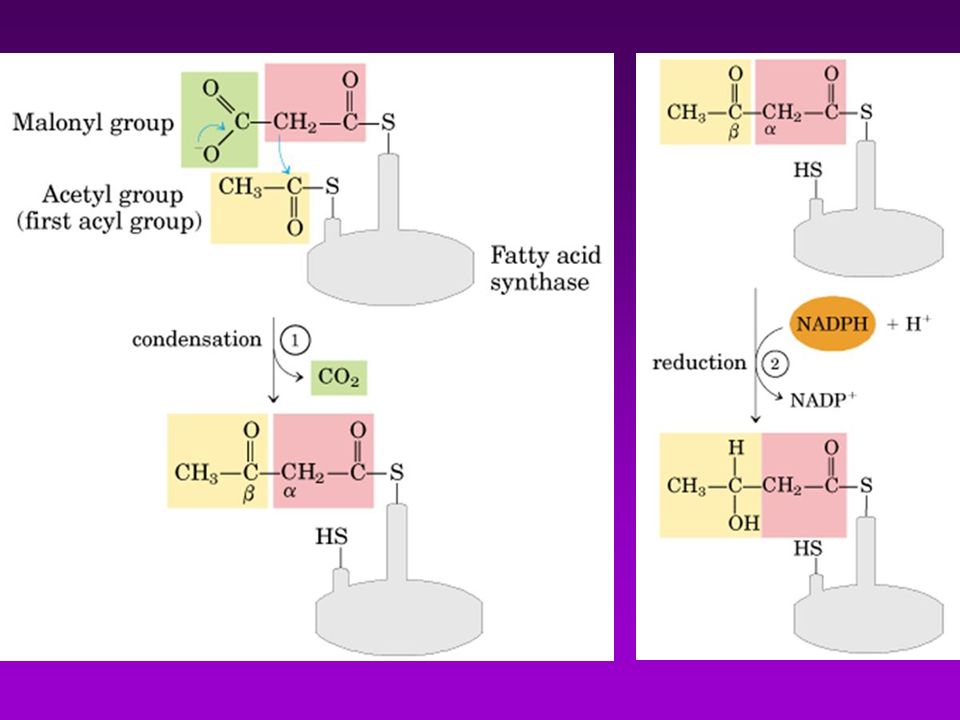
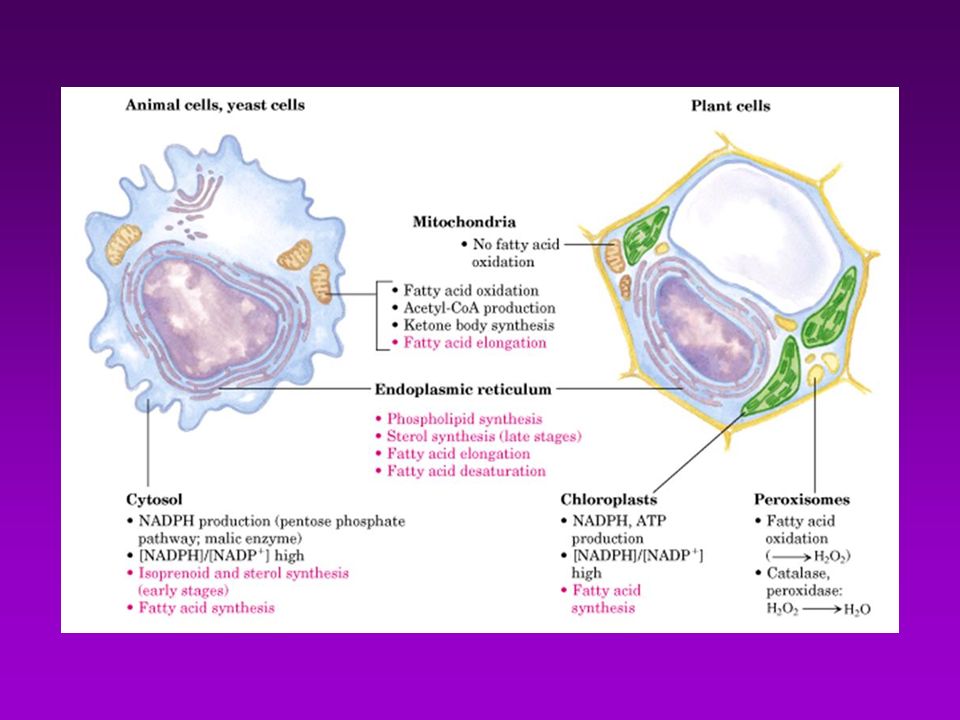
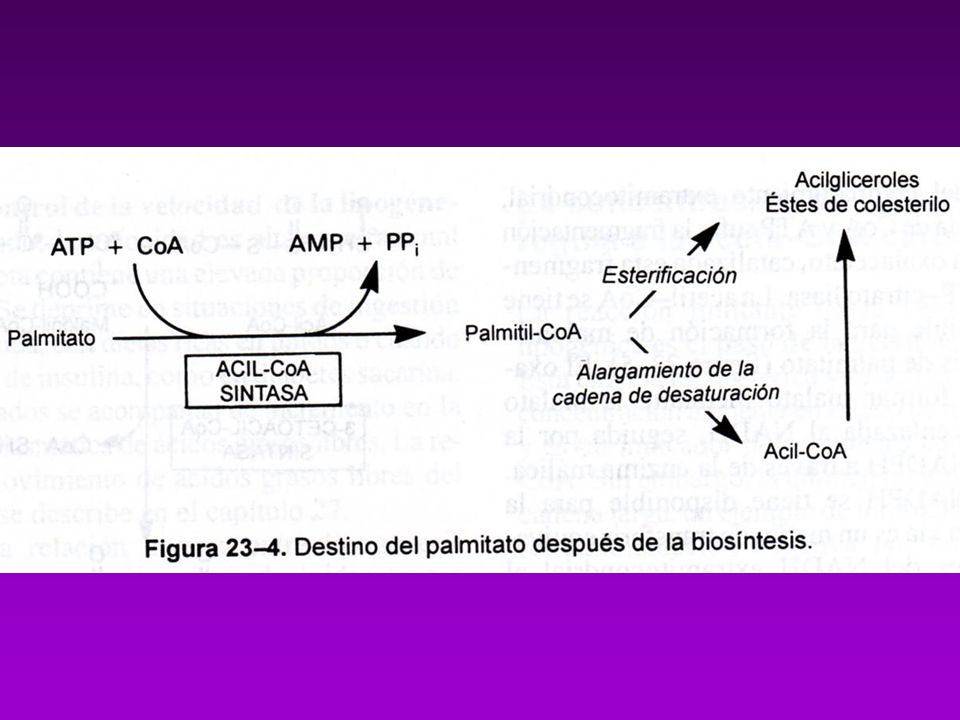
BIOSINTESÍS DE ACIDOS GRASOS
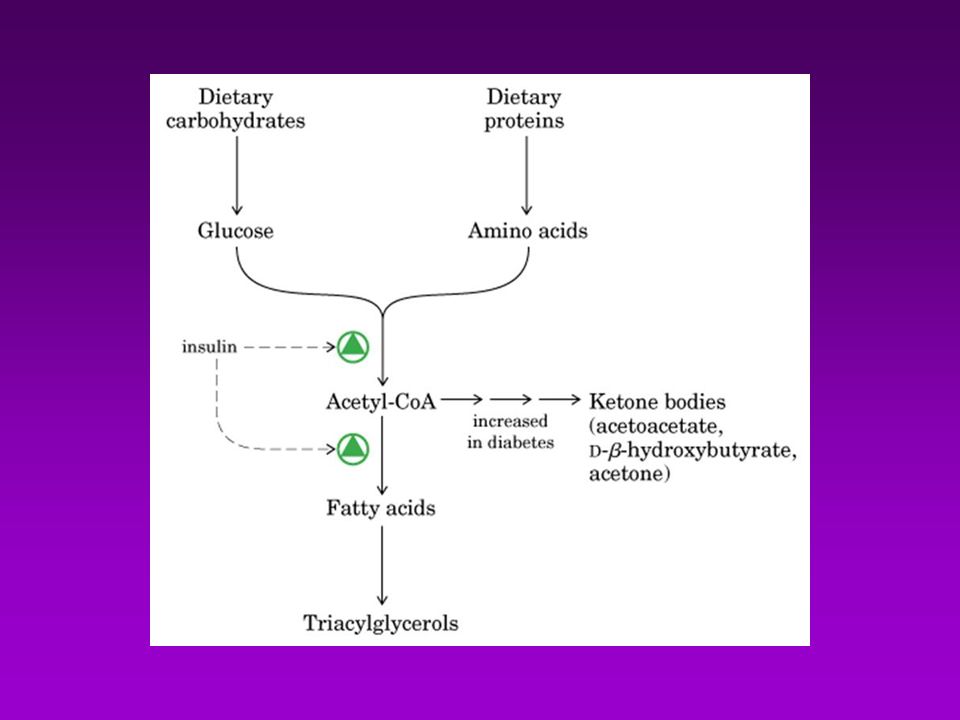
Acetil-CoA puede provenir de : Descarboxilación oxidativa de piruvato Beta oxidación de acidos grasos Degradación de las cadenas carbonadas de algunos aa

2
GLUCAGÓN GLUCOSA AMINOÁCIDOS PROTEÍNAS AG CO2 + H2O

7
AT =acetil-CoA ACP-transacetilasa (1)
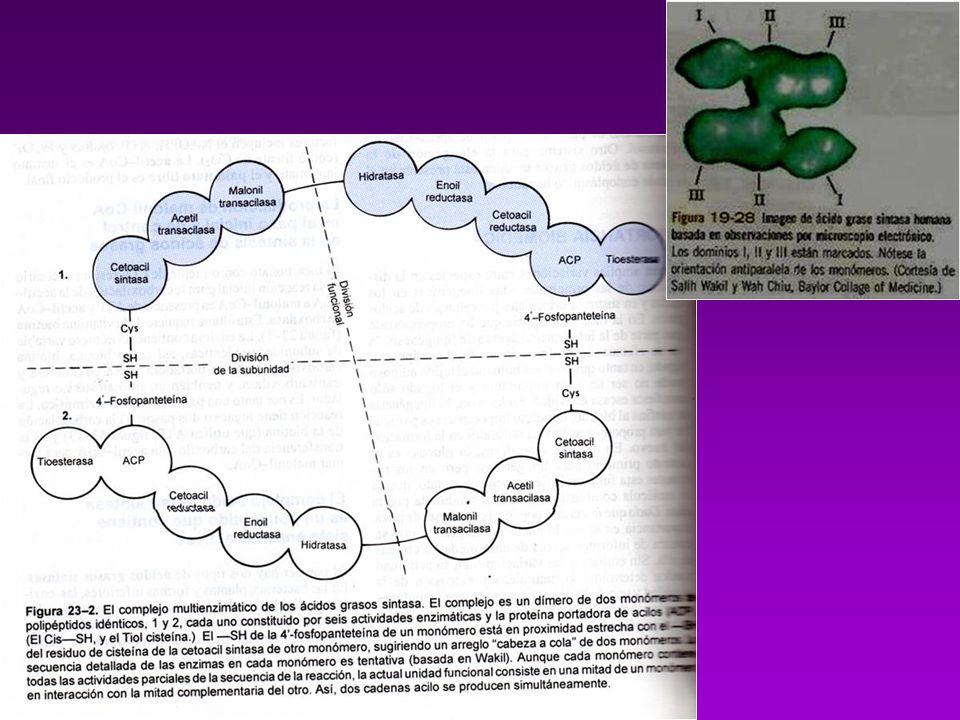
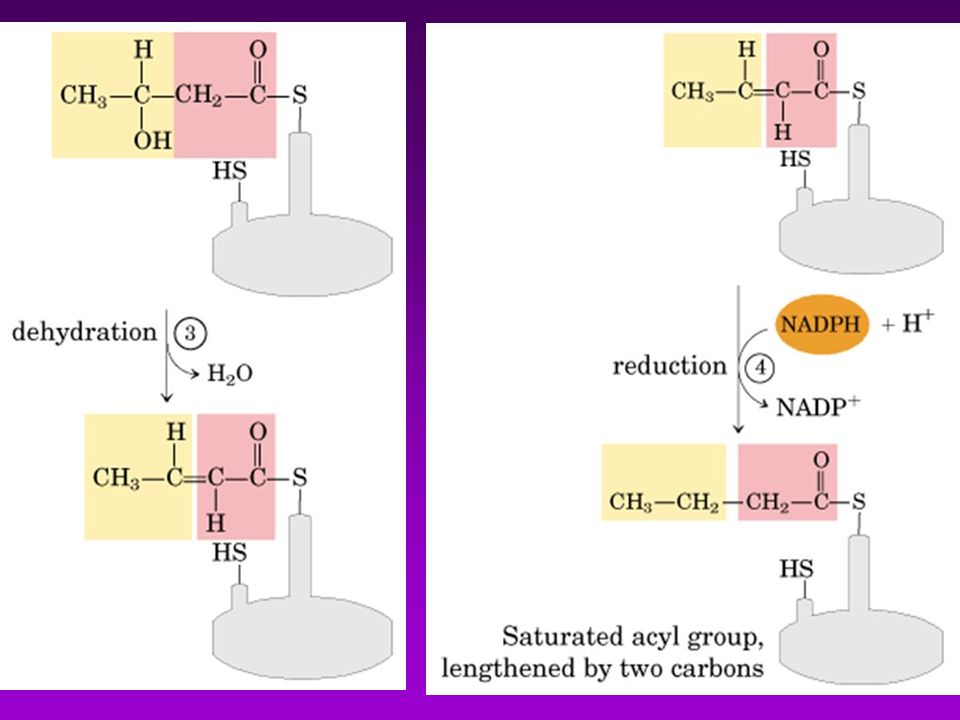
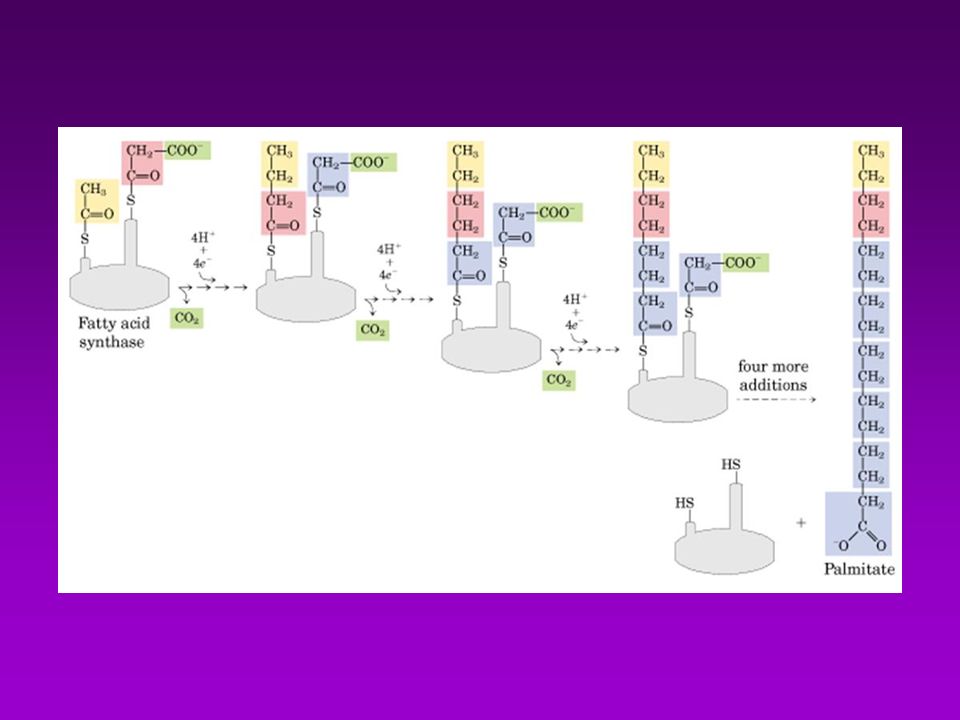
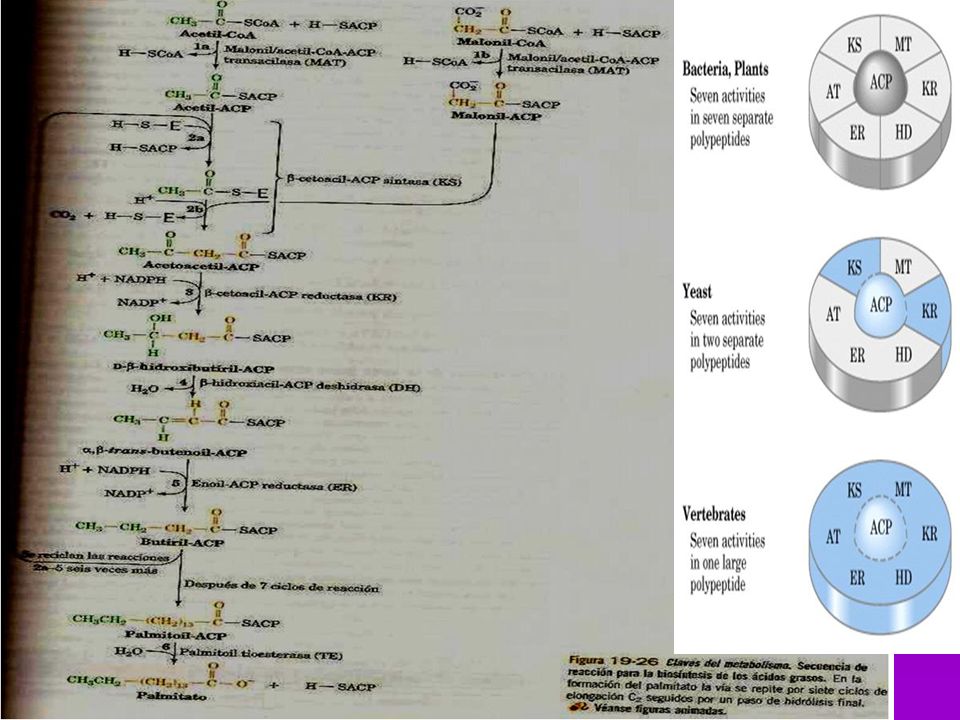
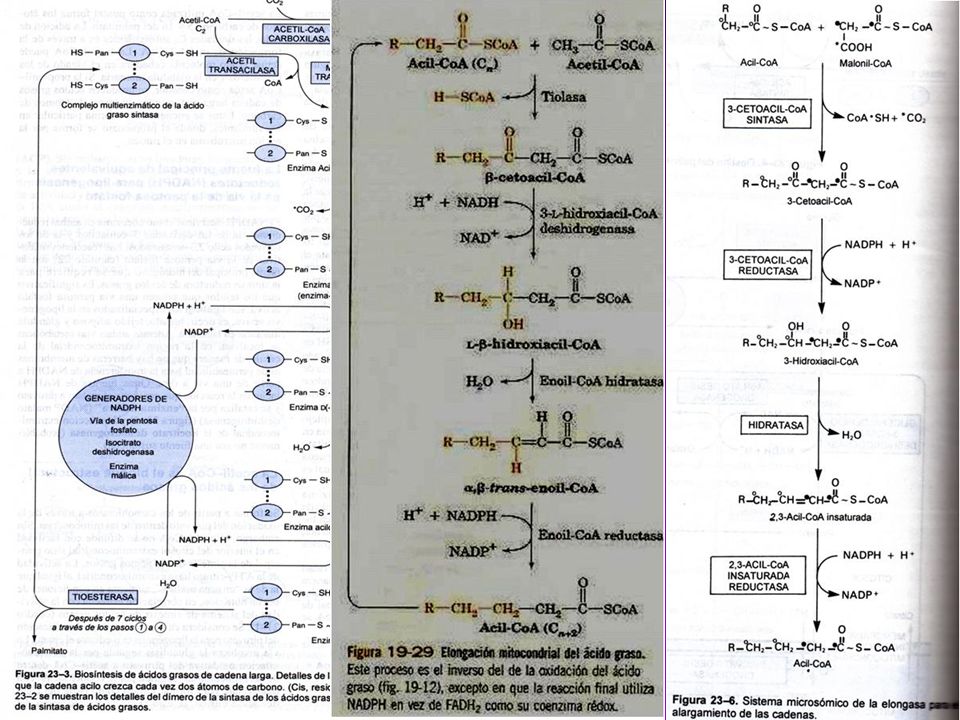
(7)TE= tioesterasa AT KR ACP MT DH TE KS ER (6)ER ER TE KS DH (5)DH MT ACP KR AT C (4)KR (3) KS (2) MT AT =acetil-CoA ACP-transacetilasa (1) MT =malonil-CoA ACP-transacilasa (2) KS =B-cetoacil ACP-sintasa (3) KR = B-cetoacil ACP-reductasa (4) DH =deshidrasa (5) ER = enoil-reductasa (6) TE = tioesterasa (palmitil deacilasa) (7) ACP = prot. Transp. de grupos acilo (c/4-P-Pantoteína) Acetil-CoA Transacetilasa
TE= tioesterasa. AT. KR. ACP. MT. DH. TE. KS. ER. (6)ER. ER. TE. KS. DH. (5)DH. MT. ACP. KR. AT. C. (4)KR. (3) KS. (2) MT. AT =acetil-CoA ACP-transacetilasa (1) MT =malonil-CoA ACP-transacilasa (2) KS =B-cetoacil ACP-sintasa (3) KR = B-cetoacil ACP-reductasa (4) DH =deshidrasa (5) ER = enoil-reductasa (6) TE = tioesterasa (palmitil deacilasa) (7) ACP = prot. Transp. de grupos acilo (c/4-P-Pantoteína) Acetil-CoA. Transacetilasa.")
18
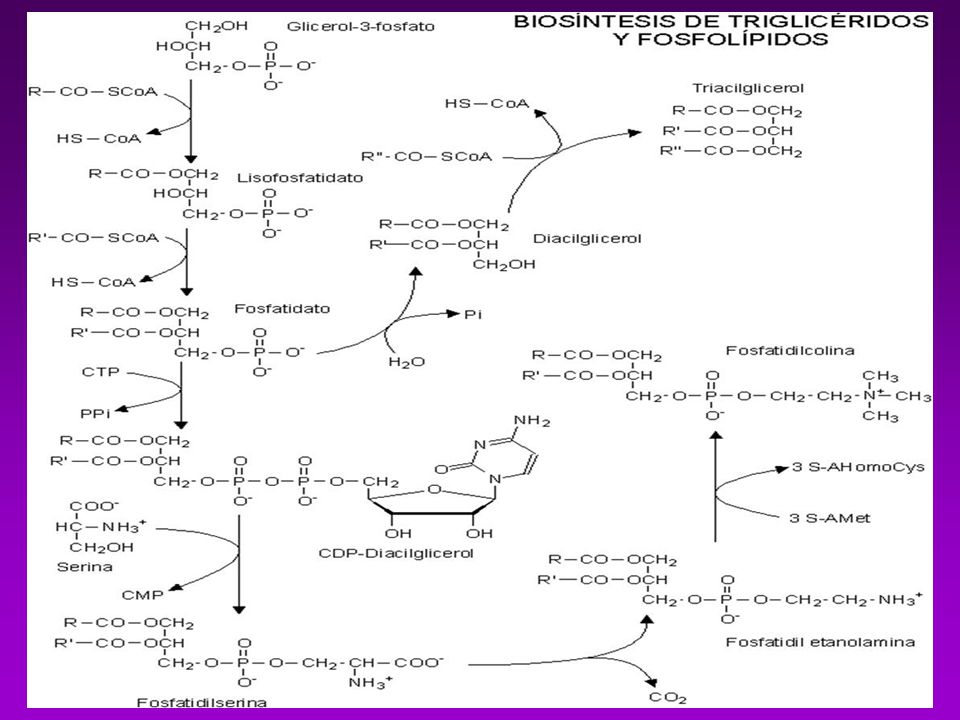
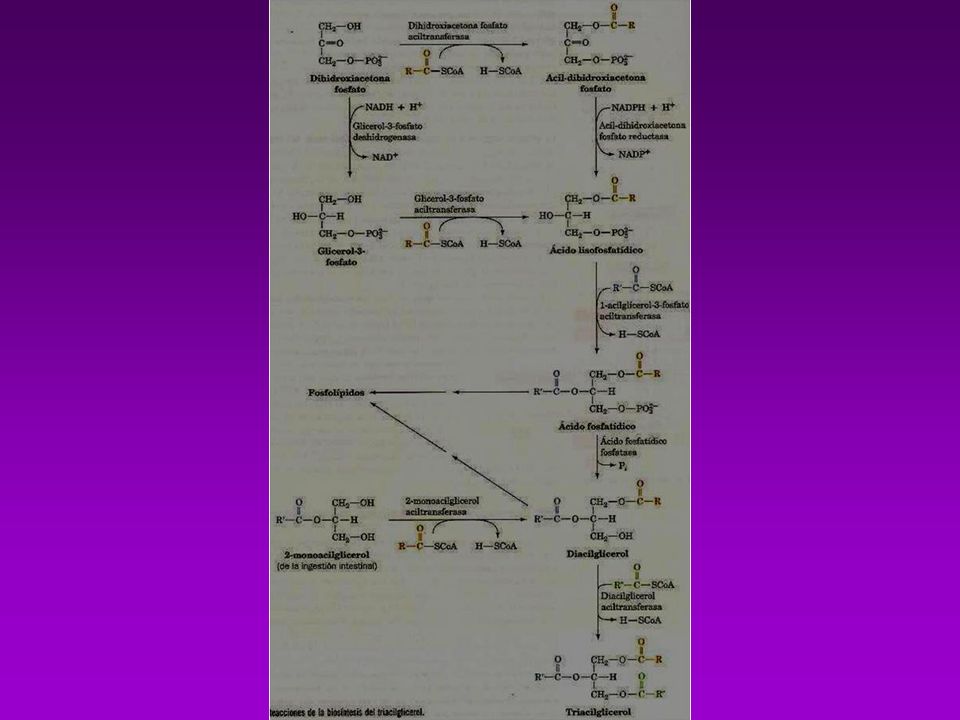
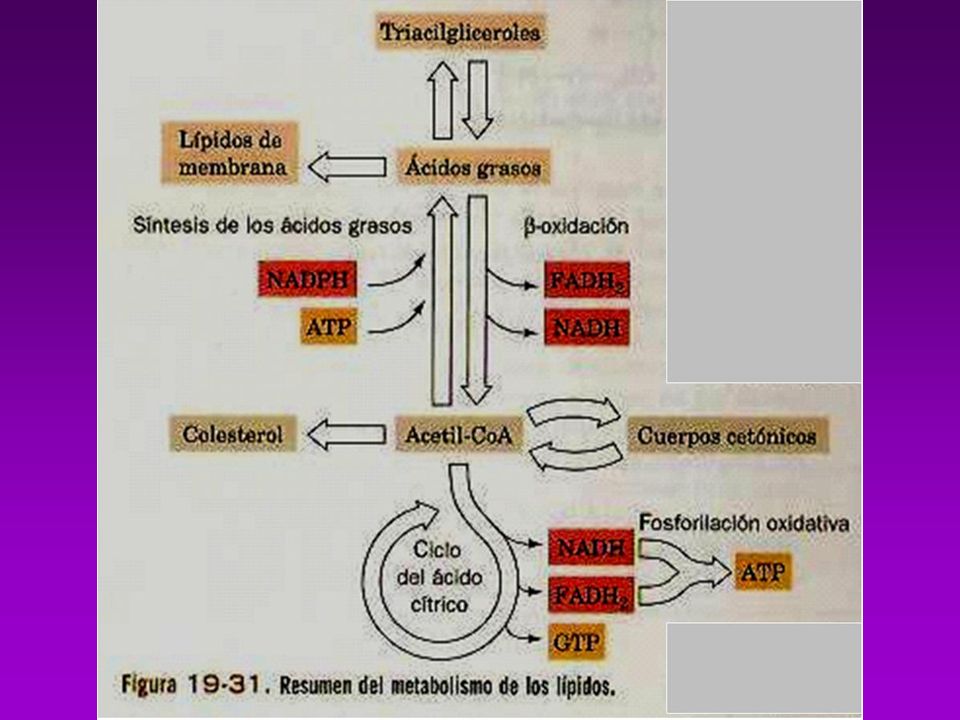
BIOSINTESIS DE TRIACILGLICEROLES
Es necesario que estén activados, el Glicerol y los ácidos grasos -Los acidos grasos se activan a acil-Coa (tioquinasa con ATP y CoA.) -Se esterifica y forma el Ácido Fosfatídico (glicerofosfato aciltransferasa) -Se hidroliza y se convierte en 1,2 diacilglicerol (fosfatasa) -Se une otra molécula de Acil CoA y forma el triacilglicerol
 -Se esterifica y forma el Ácido Fosfatídico (glicerofosfato aciltransferasa) -Se hidroliza y se convierte en 1,2 diacilglicerol (fosfatasa) -Se une otra molécula de Acil CoA y forma el triacilglicerol.")
22
ADIPOCITO

26
Metabolismo del colesterol

27
Funciones del colesterol

30
Órganos importantes en el metabolismo: intestino e hígado.
Mamíferos: sintetizan 3 ó 4 veces más colesterol que el obtenido en la dieta. Órganos importantes en el metabolismo: intestino e hígado. Aterosclerosis es un tipo de lipoidosis. Los mamíferos carecen del sistema necesario p/degradar colesterol, se excreta en la bilis junto a los ácidos biliares hacia la MF, como sus derivados (dehidrocolesterol prod. por varios tejidos y el coprosterol, del colesterol biliar por la actividad bacteriana intestinal). La hipercolesterolemia resulta de una sobreproducción y/o subutilización de LDL. La deficiencia de receptores para LDL (genética o inducida por la dieta), eleva los niveles de LDL por dos mecanismos: incrementando la producción de LDL (decreciendo la absorción IDL) y reduciendo la absorción de LDL. Además de un bajo consumo de colesterol en la dieta, se puede: 1.- Ingerir resinas que unen ácidos biliares, lo cual previene la absorción intestinal. Pero esto ocasiona un aumento en la síntesis de HMG-CoA reductasa, la cual incrementa la síntesis de colesterol. 2.- Tratar c/inhibidores competit. de la HMG-CoA reductasa, como los derivados de productos de hongos : compactina y lovastina (también llamada mevinolina). Estos decrecen la velocidad de la síntesis de colesterol. La combinación de estos tratamientos resulta en un decremento de 50-60% del colesterol en sangre.
. La hipercolesterolemia resulta de una sobreproducción y/o subutilización de LDL. La deficiencia de receptores para LDL (genética o inducida por la dieta), eleva los niveles de LDL por dos mecanismos: incrementando la producción de LDL (decreciendo la absorción IDL) y reduciendo la absorción de LDL. Además de un bajo consumo de colesterol en la dieta, se puede: 1.- Ingerir resinas que unen ácidos biliares, lo cual previene la absorción intestinal. Pero esto ocasiona un aumento en la síntesis de HMG-CoA reductasa, la cual incrementa la síntesis de colesterol. 2.- Tratar c/inhibidores competit. de la HMG-CoA reductasa, como los derivados de productos de hongos : compactina y lovastina (también llamada mevinolina). Estos decrecen la velocidad de la síntesis de colesterol. La combinación de estos tratamientos resulta en un decremento de 50-60% del colesterol en sangre.")
31
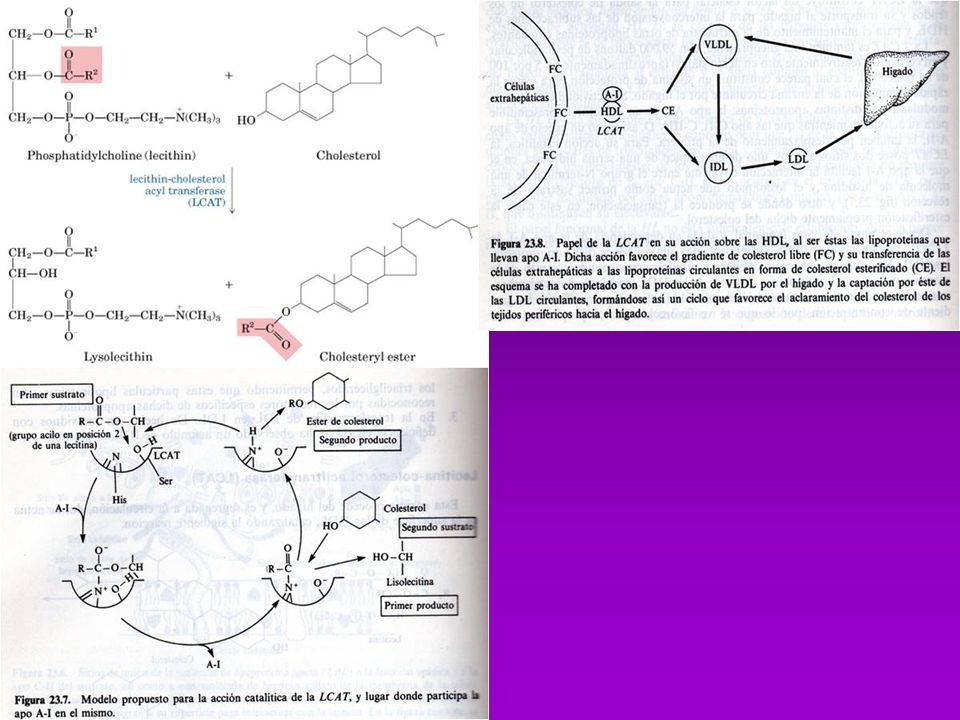
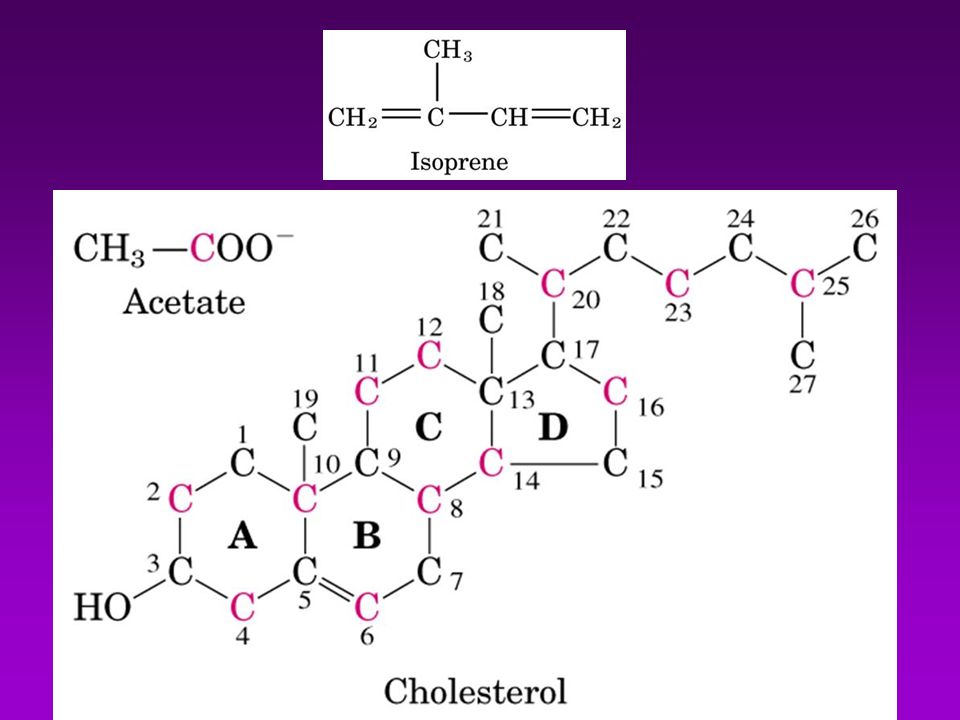
METABOLISMO DE COLESTEROL
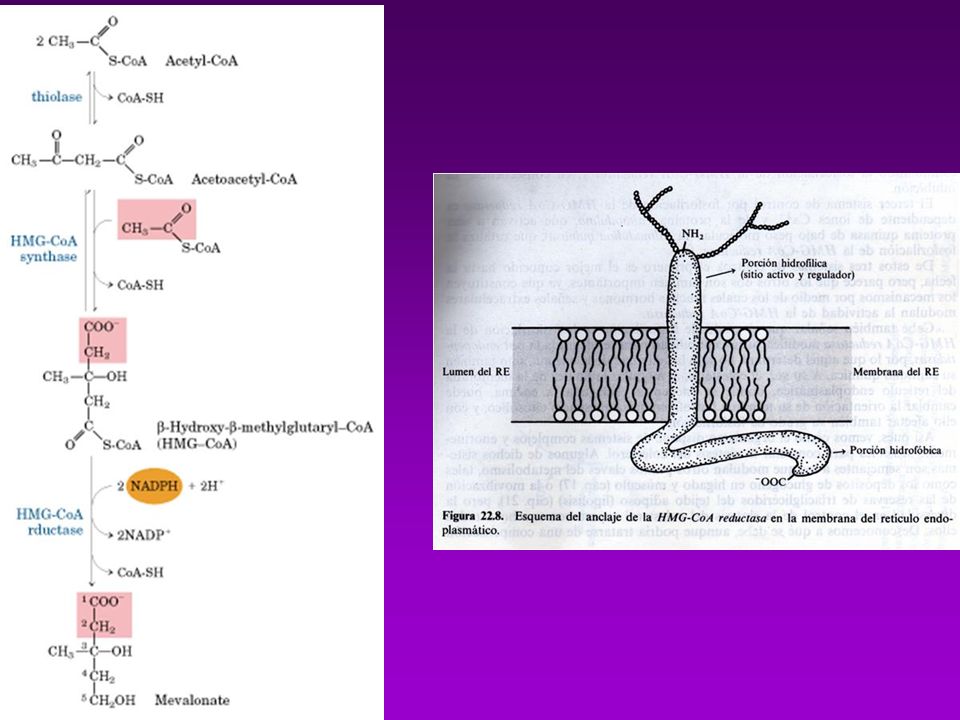
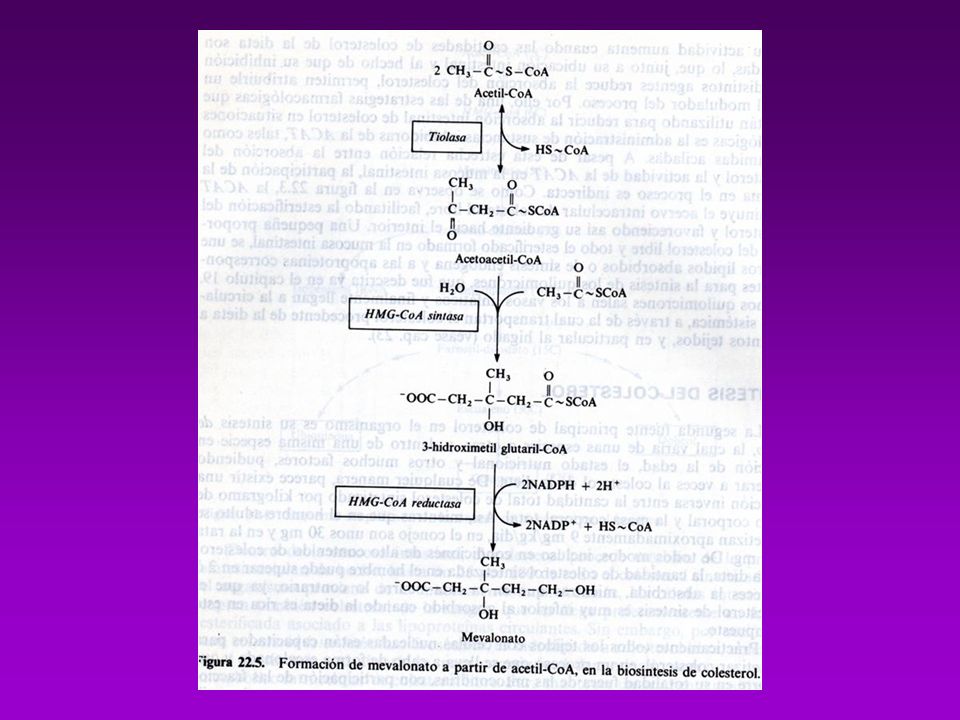
Se absorbe en intestino y se esterifica con ácido oleico (catalizada por ACAT=Acil-Coa-Colesterol-Acil-Transferasa) Los esteres se incorporan a los QUILOMICRONES, en el Hígado ocurre su degradación final BIOSINTESÍS Comprende 3 fases: A)Conversiónde acetatos en mevalonato B) Conversión de mevalonato en escualeno C) Conversión de escualeno en Colesterol A.- La formación de isopentenil pirofosfato se lleva a cabo en cuatro reacciones entre las que se cuenta la sínt. de Mevalonato B.- Formación del escualeno. C.- El escualeno es ciclizado para formar el esqueleto esteroide tetracíclico en dos pasos D.- La transformación del lanosterol a colesterol
 Los esteres se incorporan a los QUILOMICRONES, en el Hígado ocurre su degradación final. BIOSINTESÍS. Comprende 3 fases: A)Conversiónde acetatos en mevalonato. B) Conversión de mevalonato en escualeno. C) Conversión de escualeno en Colesterol. A.- La formación de isopentenil pirofosfato se lleva a cabo en cuatro reacciones. entre las que se cuenta la sínt. de Mevalonato. B.- Formación del escualeno. C.- El escualeno es ciclizado para formar el esqueleto esteroide tetracíclico en dos pasos. D.- La transformación del lanosterol a colesterol.")
33
A.- La formación de isopentenil pirofosfato se lleva a cabo en cuatro reacciones
entre las que se cuenta la sínt. de Mevalonato B.- Formación del escualeno.

34
C.- El escualeno es ciclizado para formar el esqueleto esteroide tetracíclico en dos pasos
D.- La transformación del lanosterol a colesterol

39
Síntesis de colesterol: se regula en general mediante tres vías:
1.- Modulando la actividad de la HMG-CoA reductasa, enz paso limitante en la síntesis de novo: la inhibición de corto plazo puede ser a.- competitiva. b.- por efectos alostéricos o c.- por modificaciones covalentes (fosforilación reversible vía cAMP). La regulación a largo plazo, se da regulando la síntesis y degradación de las enzimas. Este es el camino principal de regulación de la HMG-CoA reductasa, controlando la cantidad de enzima en la célula. Al disminuir LDL-colesterol o mevalonato disminuyen la canditad de HMG-CoA puede aumentar (↑síntesis y ↓degradación), cuando los niveles de LDL-colesterol o de mevalonato son regenerados el efecto es el contrario. La HMG-CoA puede ser regulada también a corto plazo, por fosforilación reversible (al igual que la glucógeno sintasa, piruvato deshidrogenasa, acetil-CoA carboxilasa y otras enzimas). La forma fosforilada es menos activa, esta reacción es catalizada por la HGM-CoA reductasa cinasa (RK); esta enzima es idéntica a la proteína cinasa dependiente de AMP (AMPK) que hace la misma reacción en la acetil-CoA carboxilasa. 2.- Regulando la síntesis de los receptores de LDL (mecanismo similar al de VLDL). El aumento en la concentración intracelular de colesterol, inhibe la síntesis del receptor de LDL y viceversa. La concentración sérica de lDL depende de la velocidad con que el hígado remueve IDL (apolipoproteínas que se unen específicamente a los receptores de LDL) de la circulación, lo cual a su vez depende del número de receptores para LDL funcionales en la superficie de los hepatocitos. 3.- Regulando la velocidad de esterificación y por tanto la liberación. La ACAT es regulada por fosforilaciones reversibles y por control a largo plazo.
. La regulación a largo plazo, se da regulando la síntesis y degradación de las enzimas. Este es el camino principal de regulación de la HMG-CoA reductasa, controlando la cantidad de enzima en la célula. Al disminuir LDL-colesterol o mevalonato disminuyen la canditad de HMG-CoA puede aumentar (↑síntesis y ↓degradación), cuando los niveles de LDL-colesterol o de mevalonato son regenerados el efecto es el contrario. La HMG-CoA puede ser regulada también a corto plazo, por fosforilación reversible (al igual que la glucógeno sintasa, piruvato deshidrogenasa, acetil-CoA carboxilasa y otras enzimas). La forma fosforilada es menos activa, esta reacción es catalizada por la HGM-CoA reductasa cinasa (RK); esta enzima es idéntica a la proteína cinasa dependiente de AMP (AMPK) que hace la misma reacción en la acetil-CoA carboxilasa. 2.- Regulando la síntesis de los receptores de LDL (mecanismo similar al de VLDL). El aumento en la concentración intracelular de colesterol, inhibe la síntesis del receptor de LDL y viceversa. La concentración sérica de lDL depende de la velocidad con que el hígado remueve IDL (apolipoproteínas que se unen específicamente a los receptores de LDL) de la circulación, lo cual a su vez depende del número de receptores para LDL funcionales en la superficie de los hepatocitos. 3.- Regulando la velocidad de esterificación y por tanto la liberación. La ACAT es regulada por fosforilaciones reversibles y por control a largo plazo.")
40
Regulación de síntesis del colesterol
La biosíntesis y el transporte de colesterol son altamente regulados en los mamíferos. La síntesis aumenta al incrementarse la concentración de colesterol, de estrógenos o sales biliares. La insulina y algunos esteroides incrementan la actividad de la HMG-CoA reductasa, por el contrario, el glucagon, los glucocorticoides y el ayuno, disminuyen su actividad. Los ácidos grasos de la dieta aumentan el colesterol sérico, en general los ácidos grasos saturados favorecen el proceso, por el contrario, los insaturados no lo estimulan. Regulación de síntesis del colesterol

41
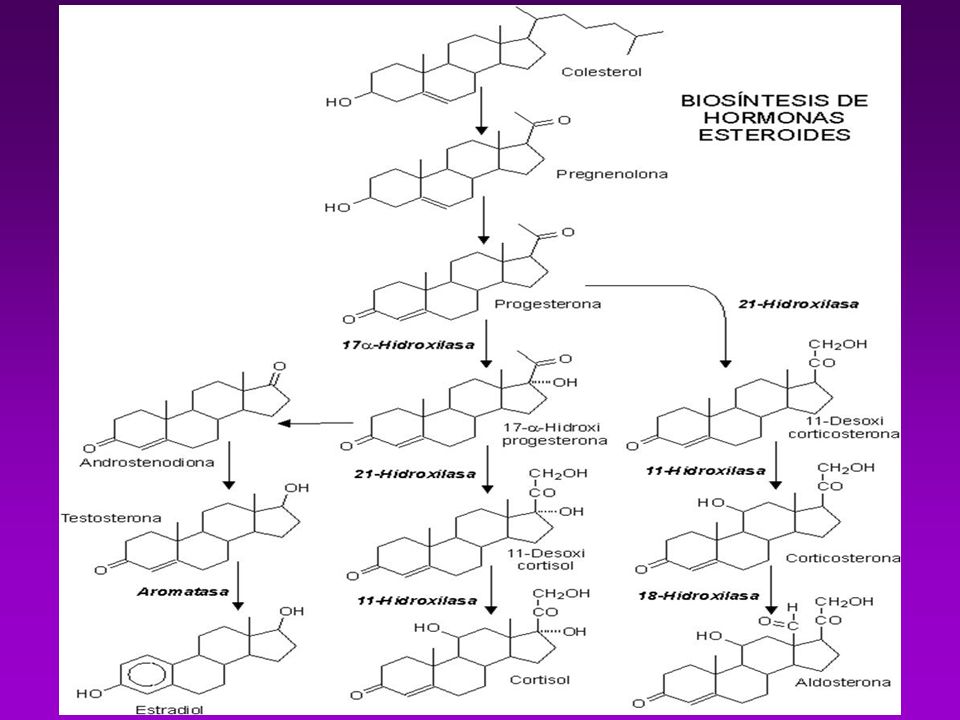
En la biosíntesis de los esteroides hay siete pasos y ocho enzimas que se listan a continuación:
Enzima que corta la cadena lateral del colesterol Esteroide C17 hidroxilasa Esteroide C17,C20 liasa Esteroide C21 hidroxilasa Esteroide 11 beta-hidroxilasa Esteroide C18 hidroxilasa 18-hidroxiesteroide oxidasa Aromatasa

44
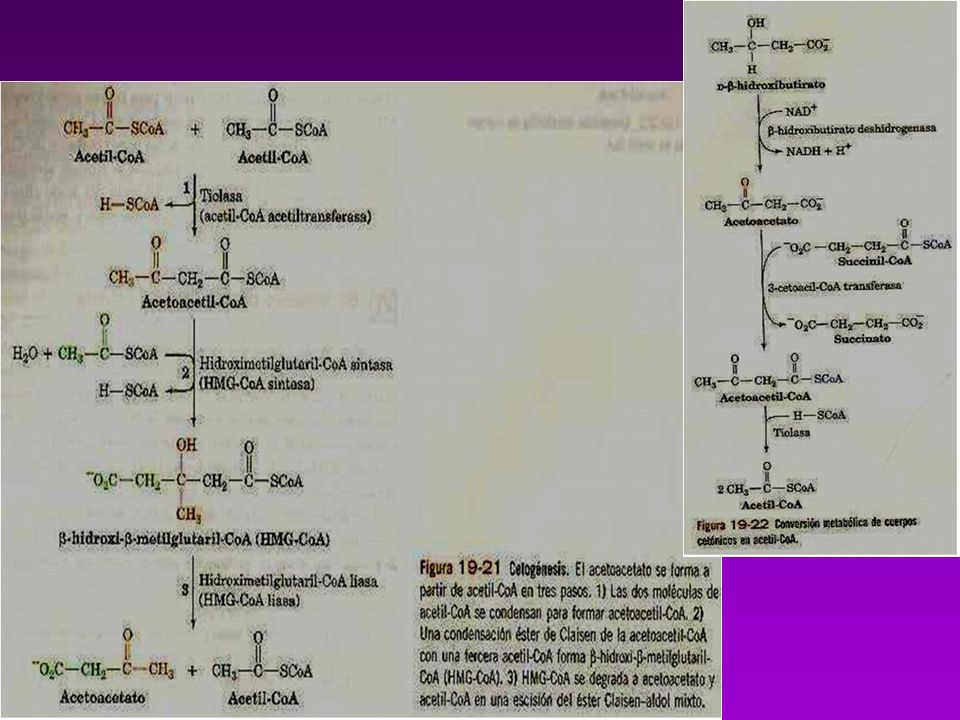
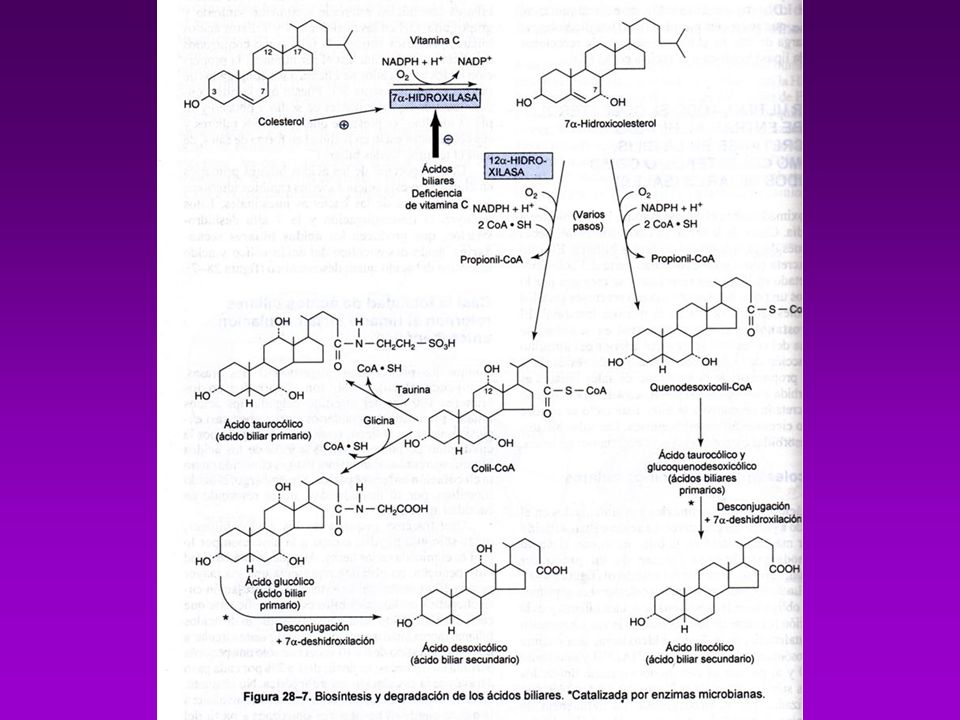
CATABOLISMO DE COLESTEROL
El organismo carece de enzimas para degradar al ciclopentanoperhidrofenantreno Se elimina a través del Hígado una parte se transforma en ácidos biliares. En el intestino son en parte reabsorbidos y vuelven al Hígado, los que no se reabsorben sufren la acción de bacterias de la flora normal se convierte el Colesterol en Coprostanol Existe equilibrio entre las cantidades sintetizadas y excretadas.

Presentaciones similares