Descargar la presentación
La descarga está en progreso. Por favor, espere

1
INHIBICIÓN DE LA ACTIVIDAD ENZIMÁTICA
Esteban Osorio Cadavid, PhD. Curso de Biología Celular y Bioquímica I

2
La inhibición de la actividad enzimática
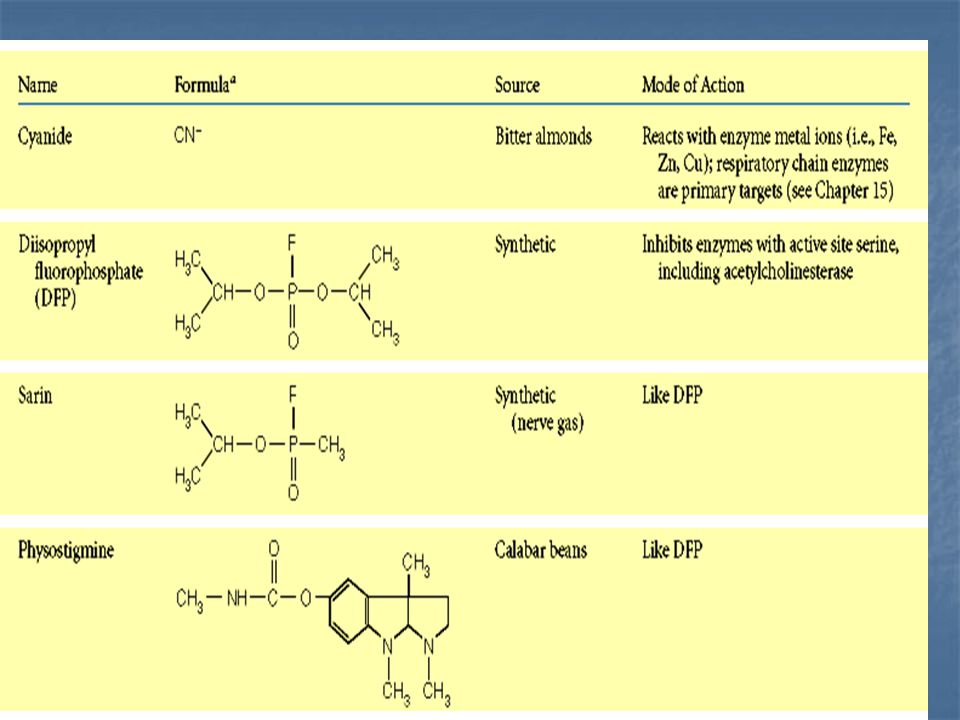
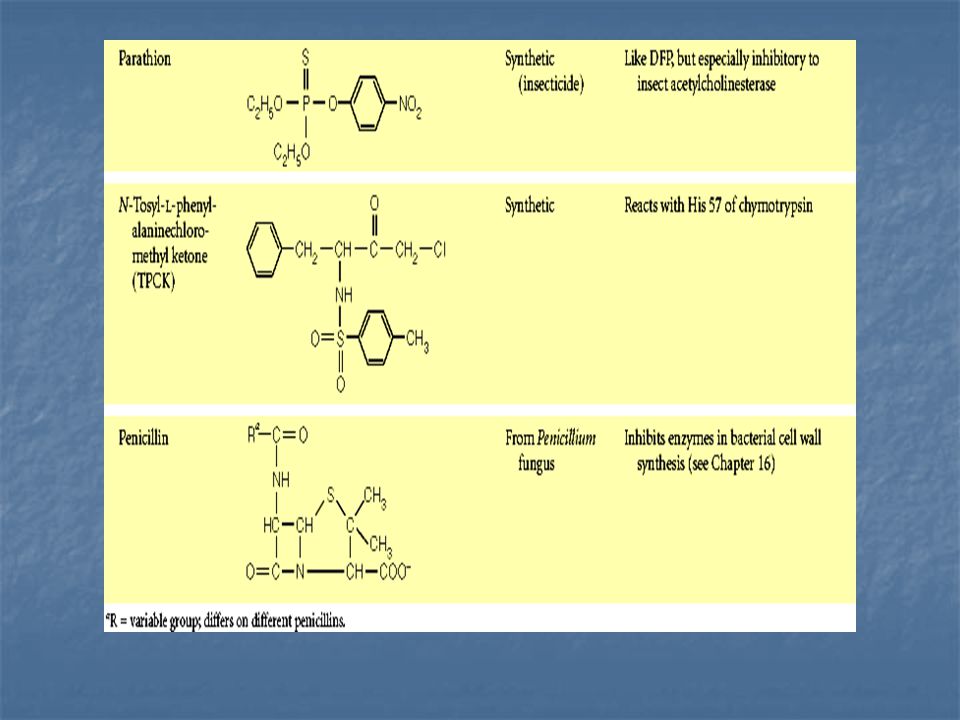
Inhibición irreversible: El inhibidor queda covalentemente unido al enzima o ligado tan fuertemente a él, que su disociación es muy lenta.

5
Inhibición irreversible mediante la formación de un aducto covalente entre DFP y serina

6
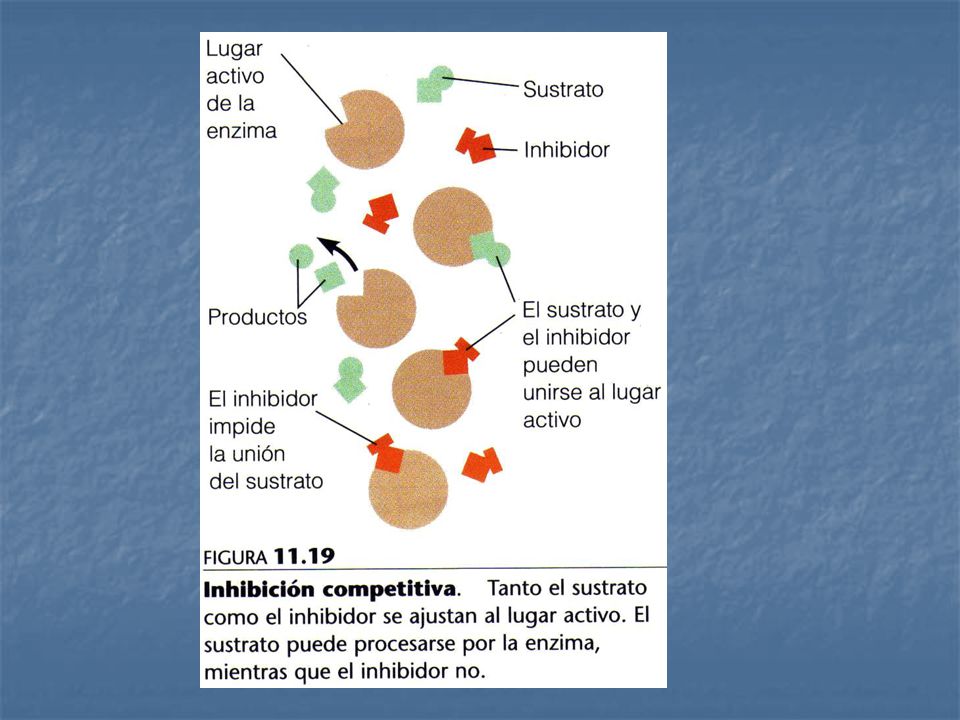
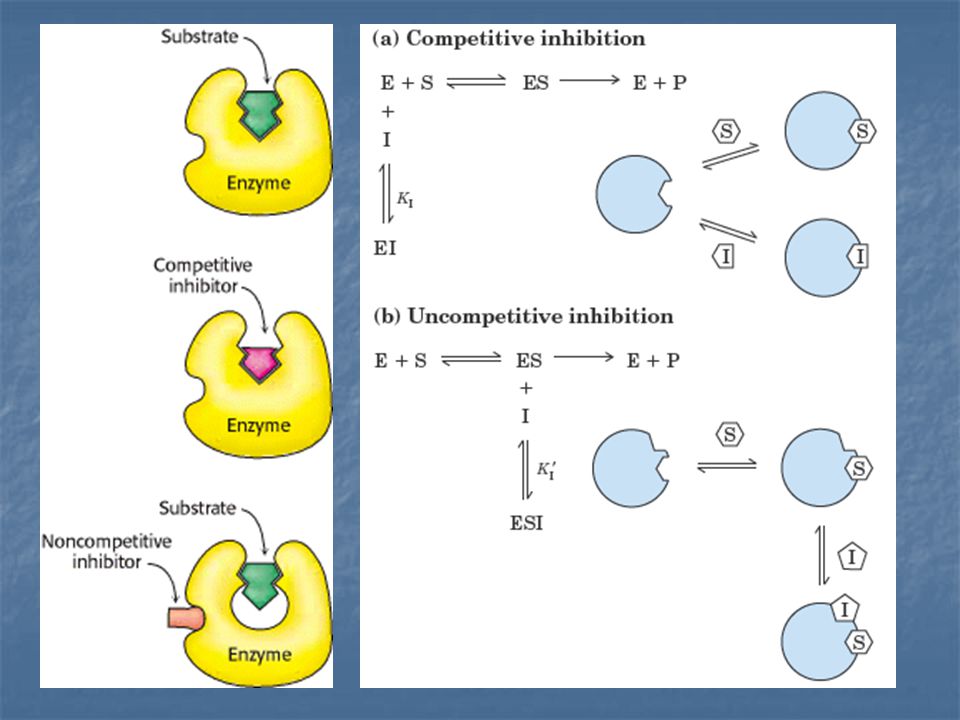
2. Inhibición reversible: se caracteriza por un rápido equilibrio entre el inhibidor y la enzima, puede ser de dos tipos: 2.1 Inhibición reversible competitiva: El inhibidor se parece al sustrato y se une al centro activo de la enzima. La unión del sustrato y el inhibidor competitivo son acontecimientos mutuamente excluyentes. Un inhibidor competitivo disminuye la velocidad de catálisis reduciendo la proporción de moléculas de enzima que quedan ligadas al sustrato.

8
Es frecuente que el producto final de una serie de reacciones enzimáticas, en una vía metabólica, sea un inhibidor del sustrato a causa de su semejanza estructural. Principio de la Inhibición por retroalimentación o “feed back”.

9
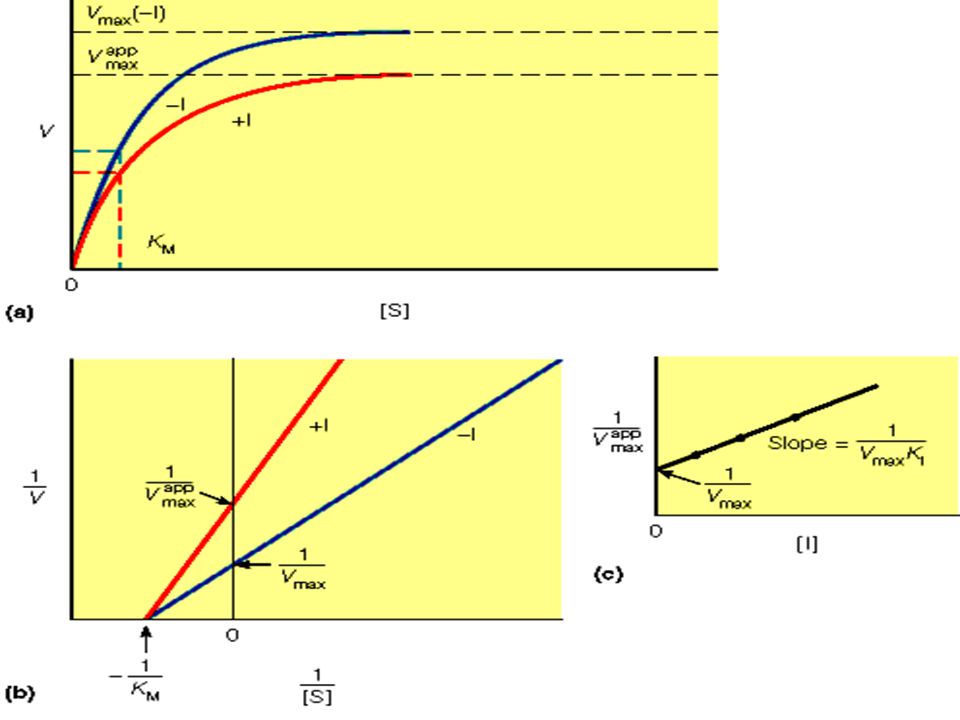
2.2 Inhibición reversible no competitiva: tanto el inhibidor como el sustrato pueden unirse simultáneamente a una molécula de enzima. Sus sitios de unión no se solapan. Un inhibidor no competitivo actúa disminuyendo el número de recambio del enzima, en vez de disminuir la proporción de moléculas enzimáticas que se han ligado al sustrato.

12
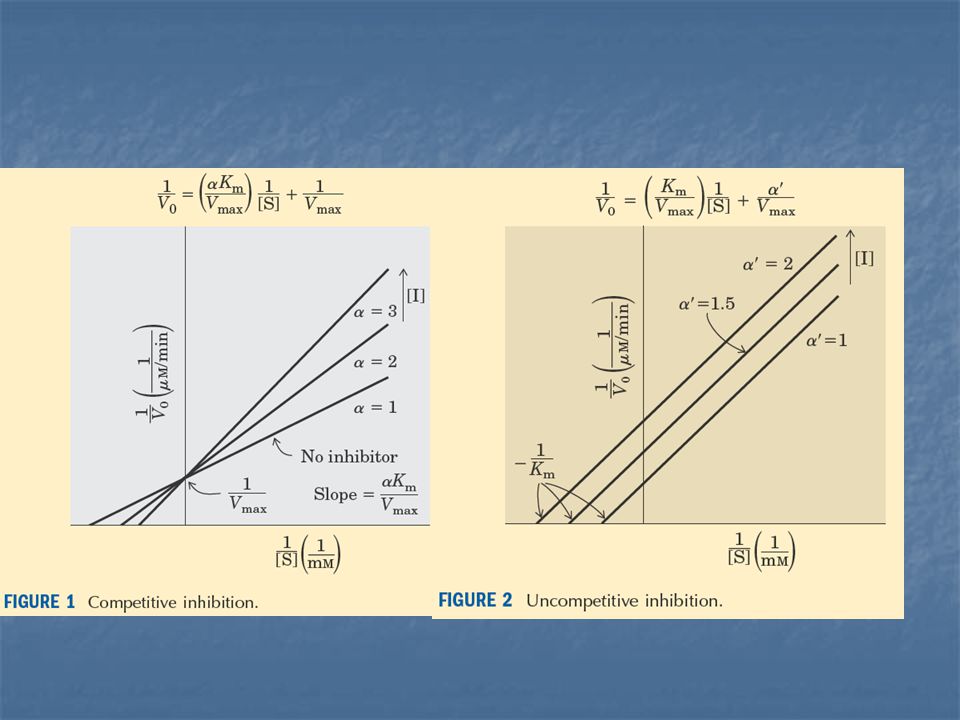
La inhibición competitiva y no competitiva son distinguibles cinéticamente.
Las medidas de velocidad de catálisis, a diferentes concentraciones de sustrato e inhibidor, sirven para distinguir entre la inhibición competitiva y la no competitiva.

14
En la inhibición competitiva, la intersección de la representación de 1/V frente a 1/S es la misma, en presencia y en ausencia del inhibidor, aunque la pendiente sea distinta.

15
Vmáxima no se altera por el inhibidor competitivo.

16
La característica fundamental de la inhibición competitiva, es que ésta puede ser superada a concentraciones altas de sustrato. El aumento en la pendiente de 1/V frente a 1/[S] indica la fuerza de unión del inhibidor competitivo.

17
En presencia de un inhibidor competitivo, la ecuación (16) puede reemplazarse por:
 puede reemplazarse por:")
18
donde E + I EI [I] = concentración del inhibidor y
Ki = constante de disociación del complejo: E + I EI
![donde E + I EI [I] = concentración del inhibidor y](http://slideplayer.es/slide/4868081/15/images/18/donde+E+%2B+I+EI+%5BI%5D+%3D+concentraci%C3%B3n+del+inhibidor+y.jpg "Ki = constante de disociación del complejo: E + I EI.")
19
La pendiente de la gráfica está aumentada por el factor en presencia de un inhibidor competitivo.

20
En la inhibición no competitiva: La velocidad máxima disminuye y la intersección con el eje Y está aumentada. La pendiente, que es igual a está aumentada por el mismo factor. En contraste con la velocidad máxima (Vmax), KM no está afectada por esta clase de inhibición. La inhibición no competitiva no puede superarse aumentando la concentración del sustrato.

21
La velocidad máxima en presencia de un inhibidor no competitivo (Vmax) , viene dada por
 , viene dada por")
23
Tratamiento del envenenamiento por Etilenglicol
El etilenglicol es un constituyente de los anticongelantes para automóviles. El perjuicio es debido al ácido oxálico, un producto de oxidación del etilenglicol por alcohol deshidrogenasa. El etanol es un sustrato competitivo y así bloquea la oxidación del etilenglicol hasta productos de tipo aldehido . El etilenglicol se excreta.

24
FALTA

25
Distinción entre inhibidores competitivos y no competitivos
Complejo enzima – sustrato El inhibidor competitivo se une al sitio catalítico de la enzima impidiendo la unión del sustrato. El inhibidor no competitivo se une a un sitio diferente, permitiendo que el sustrato se una a la enzima; sin embargo, esta unión no culmina en la transformación del sustrato en producto.

27
Curvas de actividad en función del pH de algunas enzimas

28
Efectos de la temperatura sobre la actividad de las enzimas
Las enzimas varían con respecto a su estabilidad térmica La porción descendiente de la curva se debe a la desnaturalización térmica.

29
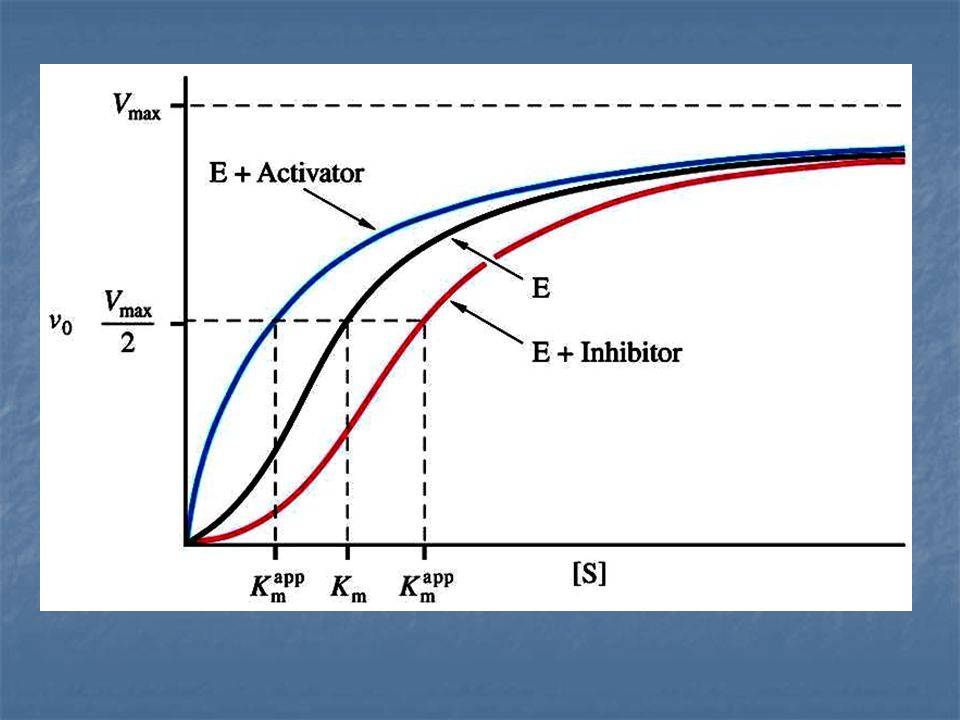
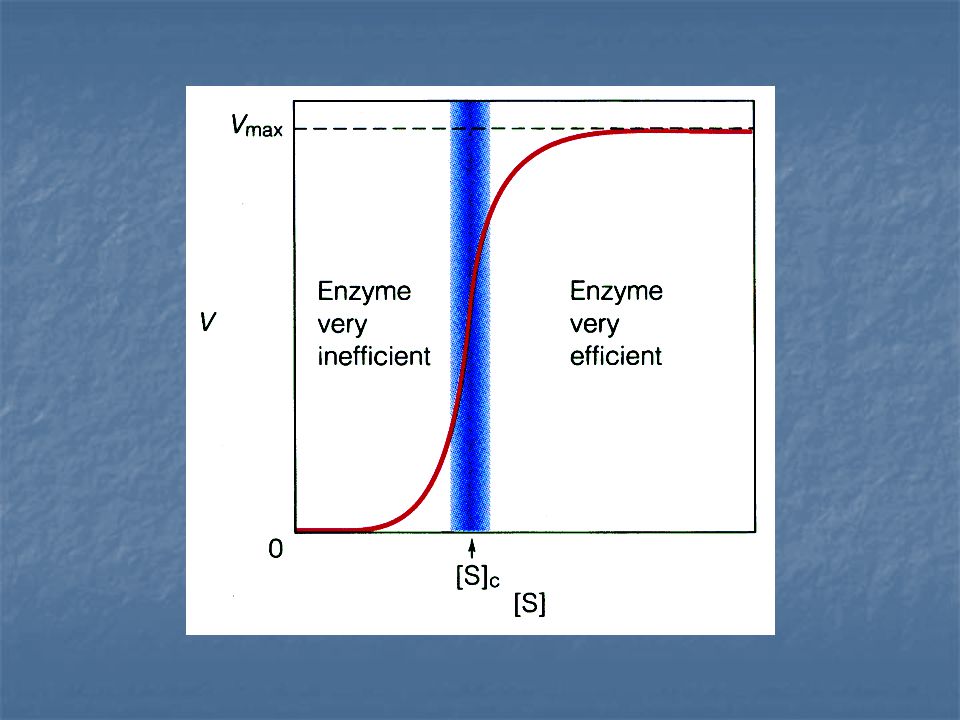
Enzimas alostéricas Está formada por varias subunidades polipeptídicas y cada una de ellas tiene un sitio activo. Su gráfica de V vs. [S] es

30
Enzimas alostéricas La actividad de estas enzimas no sigue la cinética clasica de Michaelis-Menten, en lugar de dar curvas hiperbólicas de saturación de V frente a [S], dan curvas sigmoides.

31
Los activadores y los inhibidores de las enzimas alostéricas se denominan moduladores o efectores.
Un inhibidor de la actividad enzimática se conoce como modulador negativo y un activador se conoce como modulador positivo.

32
2. En las enzimas alostéricas, un centro en una molécula enzimática puede efectuar a otro centro activo de la misma molécula de enzima, resultando de ésta interacción entre subunidades que la unión del sustrato se transforma en cooperativa, lo que daría una representación sigmoidea de V frente a [S].

33
3. Además, la actividad de las enzimas alostéricas puede ser alterada por moléculas reguladoras que están ligadas a centros diferentes a los catalíticos.

35
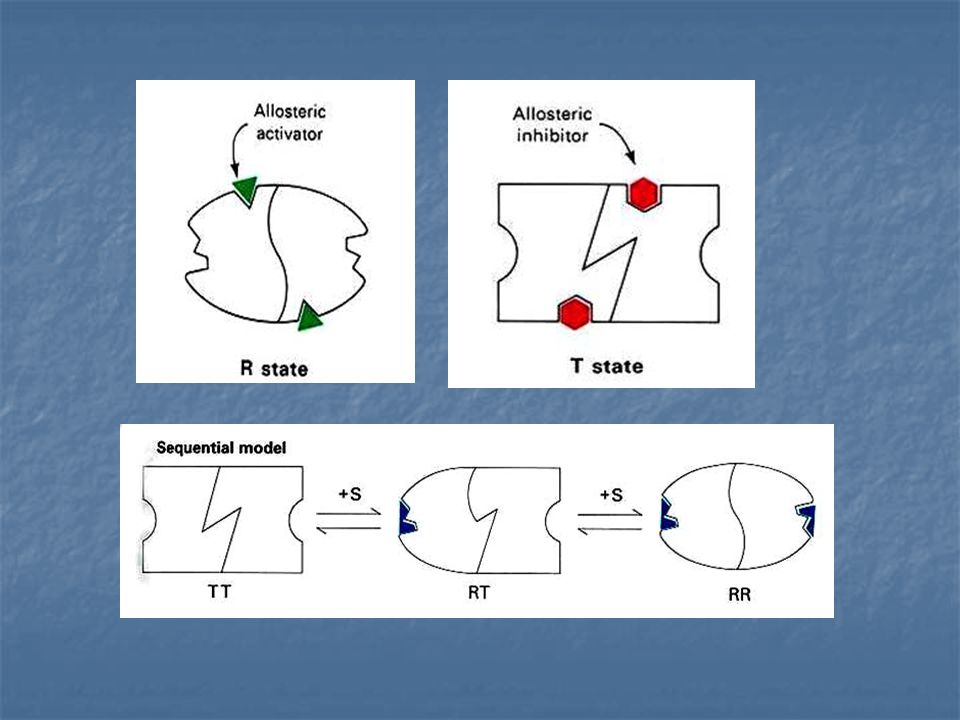
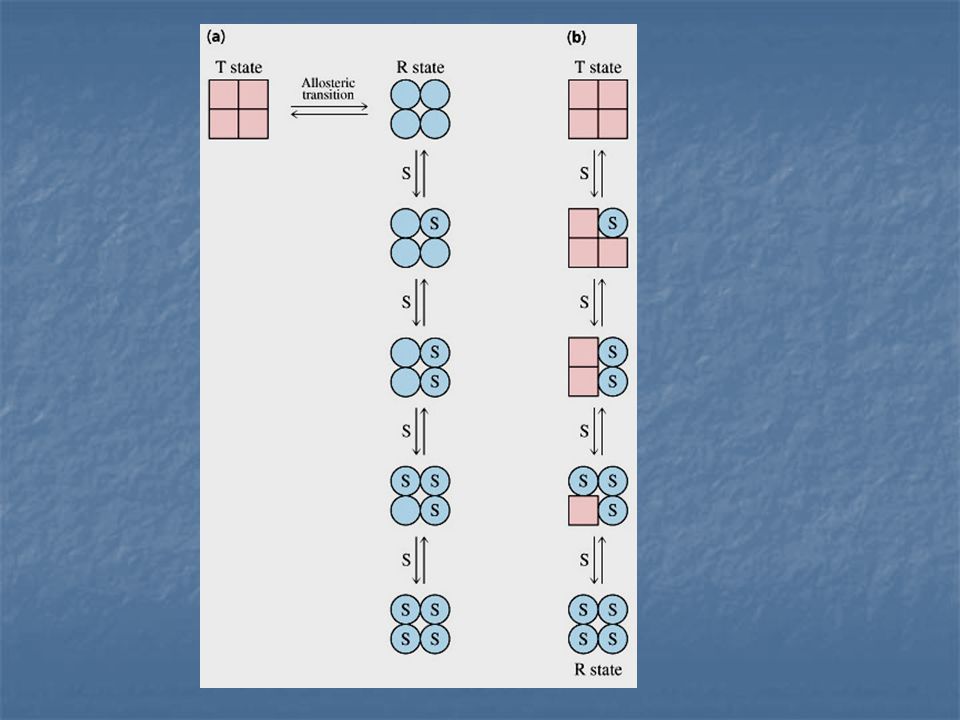
Modelo concertado de Interacciones alostéricas:
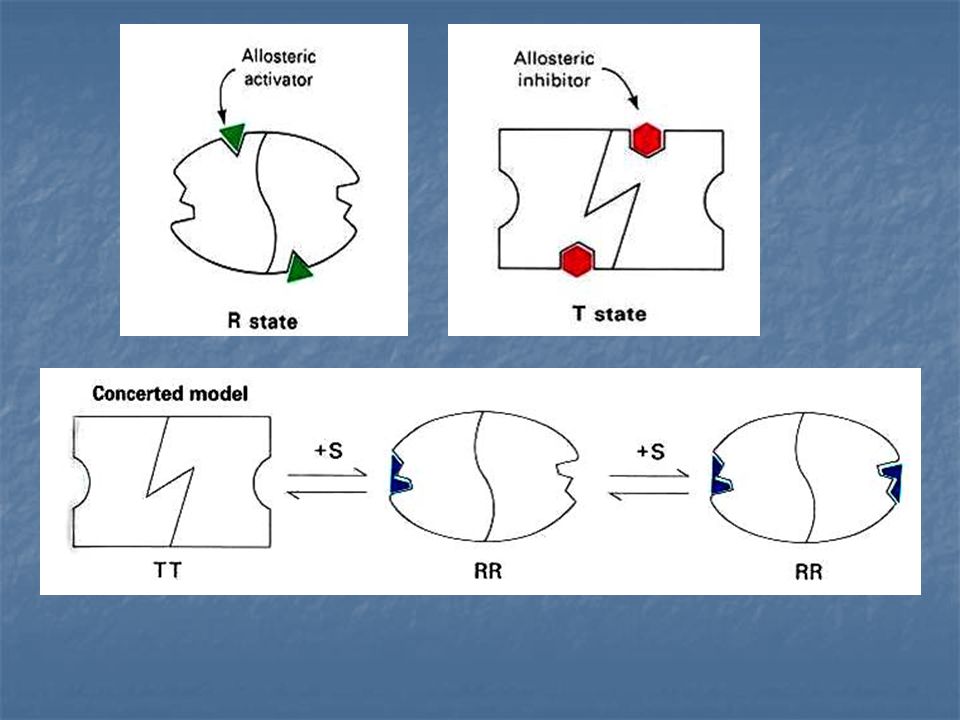
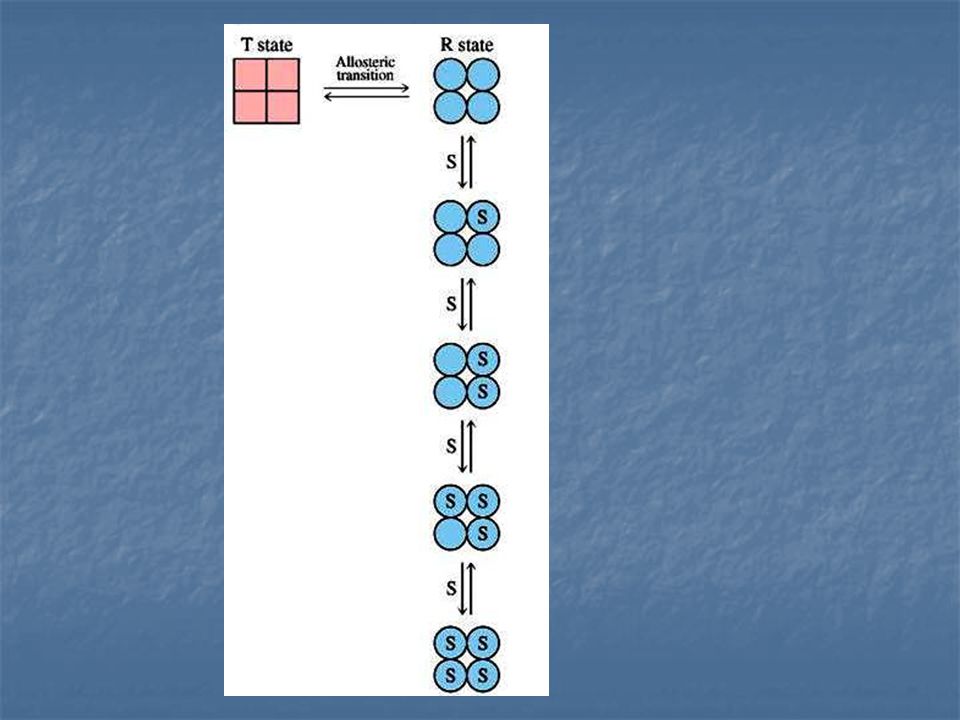
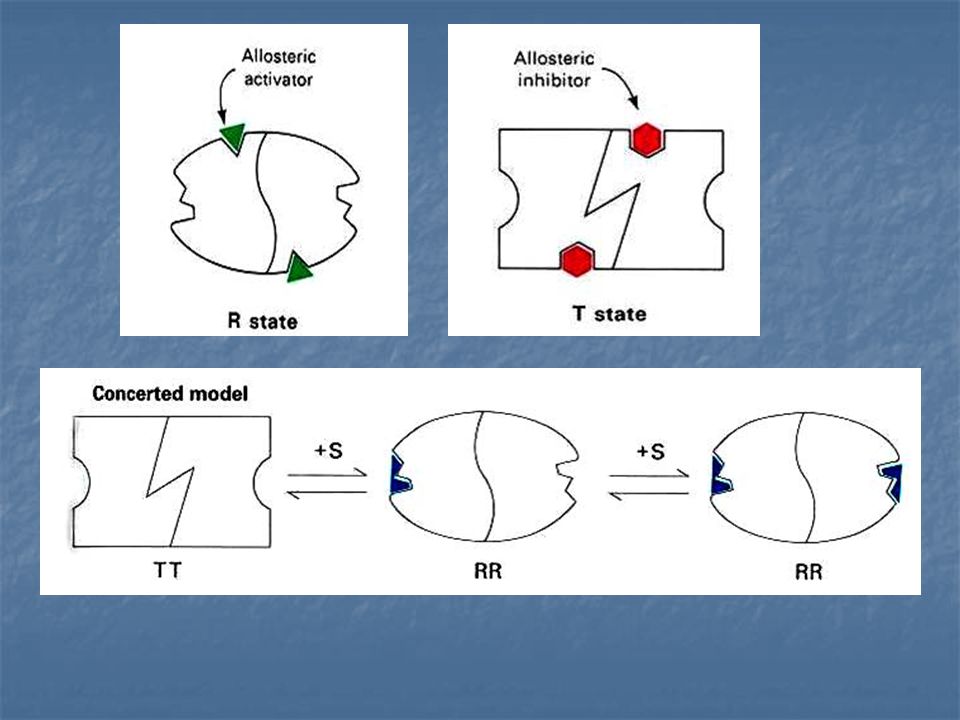
Jacques Manod, Jeffries Wyman y Jean –Pierre Changeux. Enzima Alostérica compuesta de dos subunidades idénticas, cada una con un centro activo. Cada subunidad puede exístir en cualquiera de las dos conformaciones, llamadas R y T.

37
La forma R (relajada) tiene elevada afinidad por el sustrato, la forma T (tensa o contraída) tiene afinidad baja. Una suposición importante de este modelo es la de que ambas subunidades deben esta en el mismo estado conformacional, de modo que se mantenga la simetría del dimero.

38
RR y TT son conformaciones permitidas; RT no es permitida

40
Supongamos que el sustrato no se une al estado T
Supongamos que el sustrato no se une al estado T. el estado T del dímero puede unir una o dos moléculas de sustrato, que vienen representadas por R1 y R2 respectivamente.

41
FALTA

42
Constante de disociación para un único sitio
En este modelo, la constante de disociación microscópica (KR) es igual, para el enlace de la primera y segunda molécula del sustrato a la forma R de la molécula enzimática dimérica. La fracción de Saturación (Y): o la fracción de centros activos, que tienen sustratos ligados en función de la concentración de sustrato.
 es igual, para el enlace de la primera y segunda molécula del sustrato a la forma R de la molécula enzimática dimérica. La fracción de Saturación (Y): o la fracción de centros activos, que tienen sustratos ligados en función de la concentración de sustrato.")
43
Sustituyendo las ecuaciones desde (1) a (5) en la ecuación (6) se obtiene la expresión deseada para Y:
 a (5) en la ecuación (6) se obtiene la expresión deseada para Y:")
44
Si el número de recambio de un centro activo es el mismo para los complejos ES en R1 y R2 entonces la gráfica de reacción de V vs. [S] será también sigmoidal?

45
En ausencia de sustrato, casi todas las moléculas de la enzima están en la forma T. la adicción de sustrato cambia este equilibrio conformación o favor de la forma R (el sustrato se una únicamente a la forma R). La proporción de moléculas enzimáticas en la forma R aumenta progresivamente cada vez que se añade más sustrato y por ello la unión del sustrato es cooperativa. Cuando los centros activos están completamente saturados, toda la enzima está en la forma R.
. La proporción de moléculas enzimáticas en la forma R aumenta progresivamente cada vez que se añade más sustrato y por ello la unión del sustrato es cooperativa. Cuando los centros activos están completamente saturados, toda la enzima está en la forma R..")
48
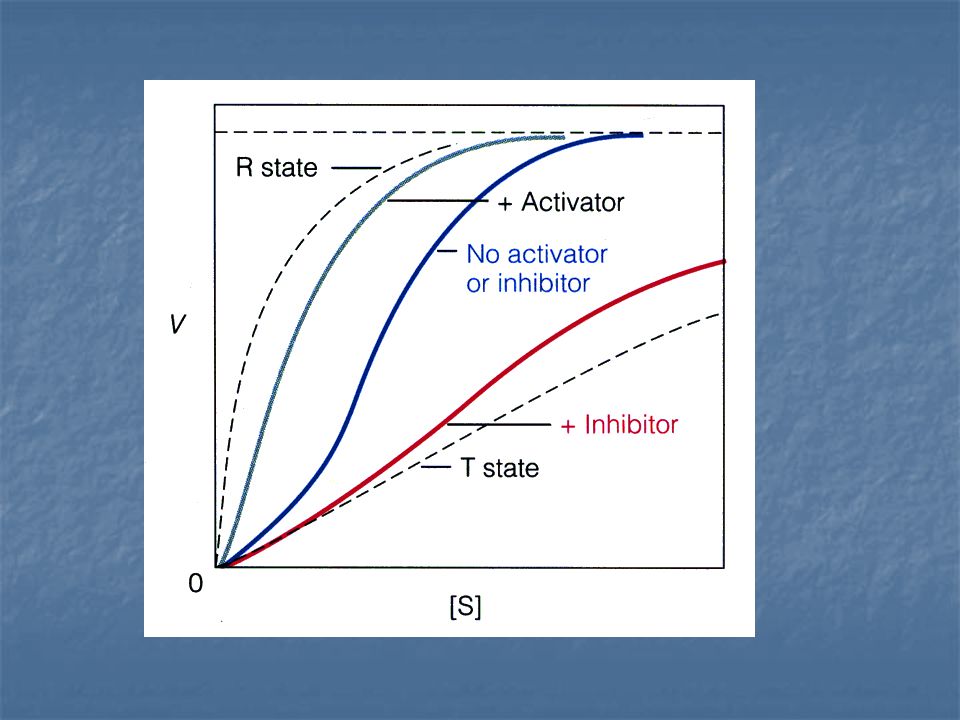
Un inhibidor alostérico se une preferentemente a la forma T.
Un activador alostérico se une con preferencia a la forma R. Un inhibidor alostérrico cambia el equilibrio conformacional R ---T hacia T, mientras que un activado alostérico lo cambia hacia R. L = constante alostérica de equilibrio

49
Un inhibidor alostérico aumenta L, un activador alostérico hace decrecer L. La fracción de saturación Y a cualquier valor del sustrato disminuye por la presencia del inhibidor y aumenta por la presencia del activador.

50
Efectos homotrópicos: Se refieren a aquellas interaciones alostéricas entre ligandos idénticos (moléculas o iones enlazados). Ej. Unión cooperativa del sustrato a la enzima.

52
Efectos heterotrópicos: Se refieren a las interacciones entre ligandos diferentes.
Ej. El efecto de un activador o inhibidor sobre la unión del sustrato porque supone las interacciones entre diferentes clases de moléculas. En el modelo concentrado de interacciones alostéricas, los efectos homotrópicos son mas (cooperativos) y los heterotrópicos (+ ó -).
 y los heterotrópicos (+ ó -).")
54
Modelo Secuencial de interacciones alostéricas.
Hay solamente dos estados conformacionales (R y T) accesibles a cada una de las subunidades. La unión del sustrato cambia la forma de la subunidad a la que está ligado. Sin embargo, la conformación de la otra subunidad no queda alterada de forma apreciable.
 accesibles a cada una de las subunidades. La unión del sustrato cambia la forma de la subunidad a la que está ligado. Sin embargo, la conformación de la otra subunidad no queda alterada de forma apreciable.")
55
3. El cambio conformacional producido por la unión del sustrato en una subunidad puede aumentar o disminuir la afinidad de unión al sustrato de las otras subunidades en la misma molécula enzimática.

57
El modelo secuencial simple difiere del modelo concertado en varios aspectos:
No se supone un equilibrio entre las formas R y T en ausencia del sustrato. La transición conformacional desde T hasta R está inducida por la unión del sustrato. El cambio conformacional de T hasta R en las diferentes subunidades de una molécula enzimática es secuencial, no concertado la especie hibrida RT es importante en el modelo secuencial, pero está excluida en el modelo concertado.

58
3. El modelo concertado supone que la simetría es esencial para la interacción de las unidades en proteínas oligoméricas y por ello requiere que sea conservada en las transiciones alostéricas. El modelo secuencial admite la interacción entre las subunidades aún cuando estén en diferente estado conformacional. 4. Las interacciones homotrópicas son necesariamente positivas en el modelo concertado, pero pueden ser positivas o negativas en el modelo secuencial.

60
Enlaces electrostáticos de Hidrógeno y de Van der Waals en los complejos Enzima-Sustrato.
Las interacciones moleculares reversibles en los sistemas biológicos vienen condicionadas por 3 clases de fuerzas diferentes: Enlaces electrostáticos Enlaces de Hidrógeno Enlaces de Van der Waals.

61
Los sustratos cargados pueden unirse a los grupos con cargas de signo opuesto de las enzimas:
La fuerza de tal interacción electrostática viene dada por la Ley de Coulomb: q1, q2, cargas de los dos grupos r distancia entre ellas D constante dielectrica del medio

62
Una interacción electrostática es mayor en el vacío (D=1) y es más débil en un medio como agua (D= 80).
 y es más débil en un medio como agua (D= 80).")
64
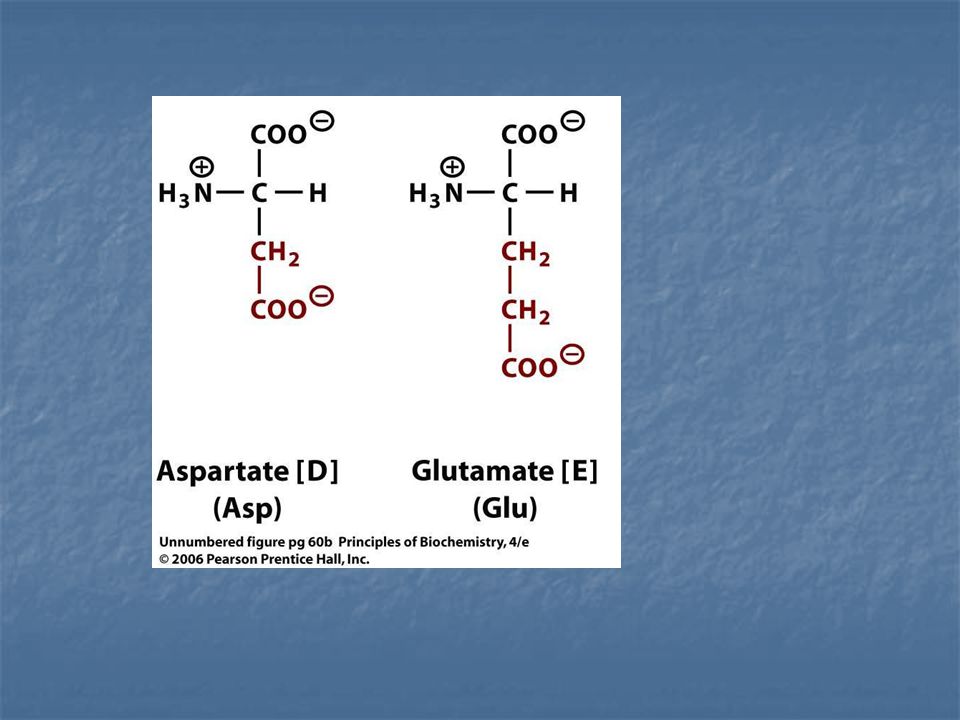
*Un sustrato Cadena lateral de lisina y arginina.
cargado Grupo imidazol de una histidina. Negativamente Grupo amino terminal de la cadena. *Un sustrato Grupos carboxílicos del aspártico y cargado glutámico Positivamente Grupo carboxido terminal de la cadena

65
2. Los sustratos se unen a las enzimas mediante enlaces de hidrógeno dirigidos muy precisamente.
Muchos sustratos no están cargados, sin embargo se unen a las enzimas con una alta afinidad y especificidad mediante puentes de Hidrogeno. En un puente de hidrógeno, un átomo de hidrogeno queda compartido por otros dos átomos: donador de Hidrógeno, al cual el hidrogeno está unido más fuertemente (covalentemente) y aceptor de hidrogeno el otro.
 y aceptor de hidrogeno el otro.")
67
Los enlaces de hidrógeno son más fuertes que los enlaces de Van der Waals, pero mucho más débiles que los enlaces covalentes. La longitud de un enlace de hidrógeno es intermedia entre la de un enlace covalente y la de un enlace de Van der Waals.

68
Un importante aspecto es que los enlaces de H son altamente direccionales.
Interacciones de los puentes de hidrógeno en la unión de un sustrato a la ribonucleasa.

69
Las proteínas son ricas en potencialidad para formar puentes de Hidrógeno.
11 de los 20 aminoácidos fundamentales pueden formar enlaces de Hidrógeno a través de sus cadenas laterales.

70
Las cadenas laterales de Triptófano y Arginina pueden servir solamente como dadores de Hidrógeno para el enlace.

71
La cadena lateral de la Asparragina, Glutamina, Serina y Treonina pueden servir a la vez como donadores y aceptores de enlaces. Lo mismo ocurre en el grupo peptídico.

72
Las capacidades para formar puente de Hidrógeno de la lisina (y el grupo amino terminal), del aspártico y glutámico (y el grupo carboxilo terminal), la tirosina y la histidina varían con el pH. La manera de enlazar hidrogeno de estos residuos ionizables es dependiente del pH.

74
Las interacciones de Van der Waals son importantes cuando existe complementariedad estérica.

75
Fuerza de atracción no específica entre dos átomos cualesquiera cuando se encuentran separados alrededor de 3-4 A (enlace de Van der Waals). La atracción de un par de átomos aumenta a medida que se acercan, hasta que están separados por una distancia de contacto de Van der Waals. b. A distancia menor, las fuerzas fuertemente repulsivas se hacen dominantes a causa que las nubes electrónicas se solapan. La energía del enlace de Van der Waals para un par de átomos, es de aproximadamente 1Kcal/mol.

76
Radios de contacto de Van der Waals de los átomos.
ATOMO RADIO (A) H C N O S P
 H 1.2. C 2.0. N 1.5. O 1.4. S P 1.9.")
77
Las fuerzas de Van der Waals desaparecen rápidamente cuando la distancia de un par de átomos es mas grande que sus distancia de contacto. Las fuerzas de Van der Waals se hacen significativas en los enlaces, sólo cuando numerosos átomos de sustrato pueden acercarse simultánemente a numerosos átomos de la enzima.

78
Numerosos átomos de sustrato pueden interaccionar con muchos átomos del enzima solamente si sus formas son adecuadas. Una interacción efectiva de Van der Waals entre el sustrato y un enzima puede darse solamente si son estéricamente complementarias

79
Aunque no existe virtualmente especificidad en una sola interacción de Van der Waals, la especificidad se produce cuando existe la oportunidad de producir un gran número de enlaces de Van der Waals de manera simultánea.

80
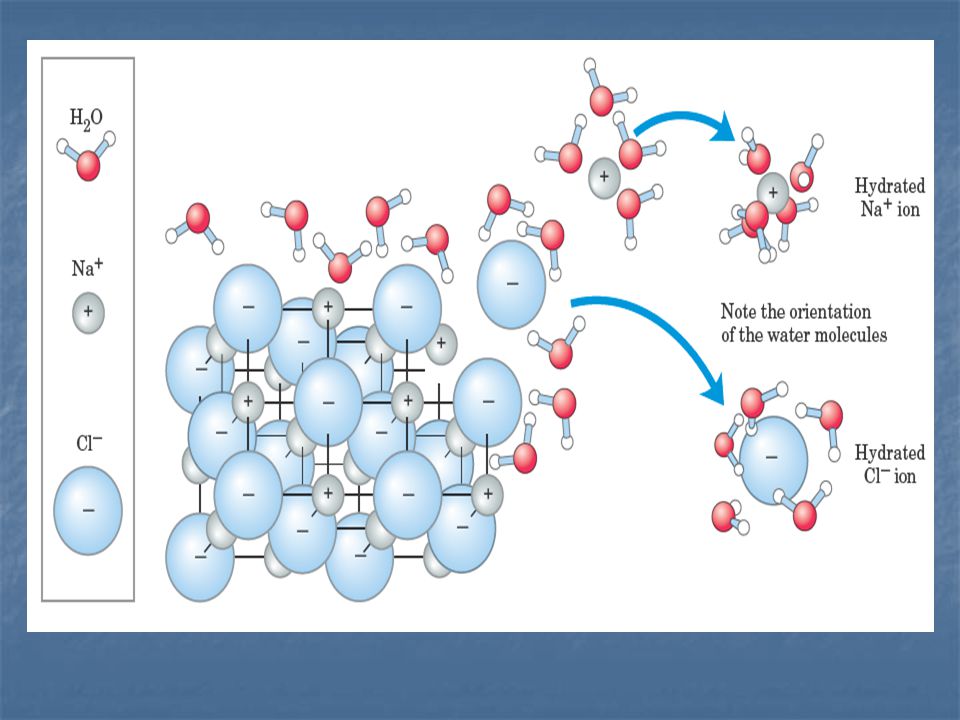
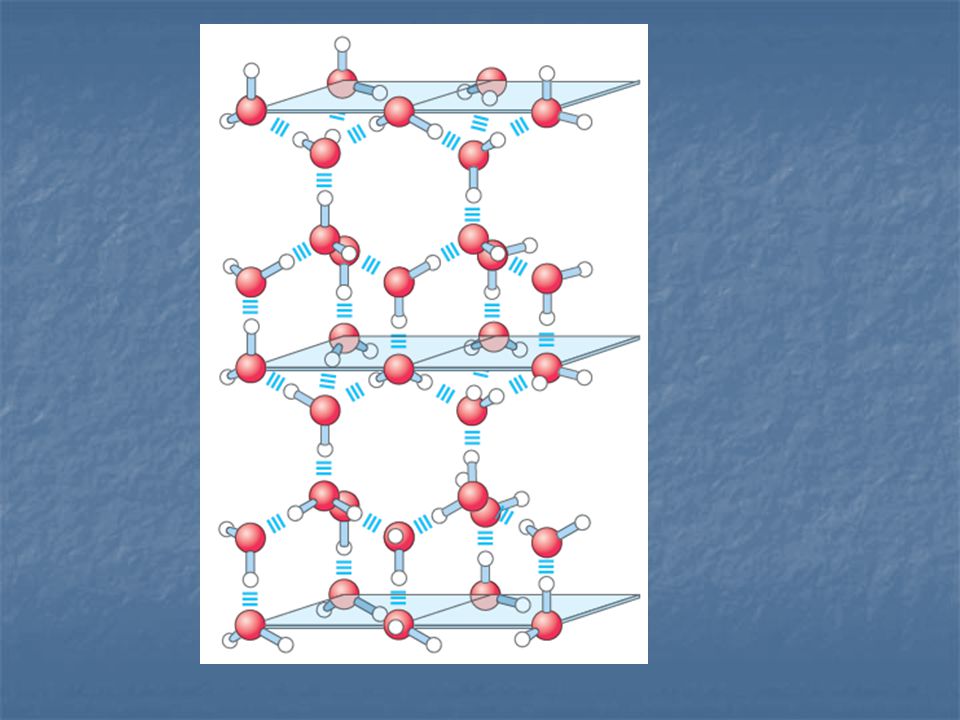
Cuál es el efecto del agua sobre las tres clases fundamentales de enlace?.
Dos propiedades del agua son especialmente importantes a este respecto: El agua es una molécula polar. Las moléculas de agua poseen entre si una alta afinidad. El agua es una molécula altamente cohesiva.

81
Por su polaridad y su capacidad de formar puentes de hidrógeno, el agua es una molécula altamente reactiva. El agua debilita las interacciones electrostáticas y los enlaces de H, entre otras moléculas.

82
Una interacción fuerte de enlaces de hidrógeno entre el grupo CO y NH sucede solamente si se excluye el agua.

83
El agua disminuye la fuerza de las interacciones electrostáticas unas 80 veces, respecto al vacío, debido a que el valor de su constante dieléctrica es 80.

84
Las moléculas o grupos apolares tienden a agruparse en el seno del agua. Las moléculas Apolares se reúnen conjuntamente en el seno del agua, no precisamente porque manifiesten una gran afinidad entre ellas mismas, sino porque las moléculas de agua se enlazan mutuamente con fuerza.

85
Las interacciones hidrofóbicas son la fuerza principal que dirige:
el plegado de las macromoléculas, la unión de sustratos a las enzimas y muchas mas interacciones moleculares en sistemas biológicos. La base de la interacción hidrofóbica radica en el aumento de entropía que resulta del incremento de libertad de las moléculas de agua libradas.

86
Consideremos la disolución de una sola molécula apolar, como el hexano En agua
Se crea una cavidad que transitivamente interrumpe algunos enlaces de hidrógeno entre molécula de agua. Las moléculas de agua dispersadas se reorientan y tienden a formar el máximo de nuevos enlaces de Hidrógeno.

87
3. Las posibilidades de formación de enlaces de Hidrógeno favorables en la cavidad de acuosa que rodea la molécula de hexano son mucho menores que en el agua pura. 4. Las moléculas de agua que rodean al hexano están mucho más ordenadas que en el resto de la disolución. La entropía de la disolución diminuye.

88
Consideremos la ordenación de dos moléculas de hexano en agua.
Se situarán en dos cavidades pequeñas o en una sola cavidad mayor? Las moléculas se reúnen y ocupan una cavidad mayor. Asociación favorecida por la liberación de parte de las moléculas de agua que rodeaban a los hexanos separados.

89
2- De este modo, la base de la interacción hidrofóbica radica en el aumento de entropía que resulta del incremento de libertad de la moléculas de agua liberadas.

90
FALTA DIAGRAMA

Presentaciones similares














![1 KM 1 1 Vo= Vmax [S] + Vmax Y = m x + b = m = b.](/2/161459/big_thumb.jpg "1 KM 1 1 Vo= Vmax [S] + Vmax Y = m x + b = m = b.>")







