Descargar la presentación
La descarga está en progreso. Por favor, espere

1
FACULTAD DE CIENCIAS BIOLÓGICAS
E.A.P MICROBIOLOGÍA Y PARASITOLOGÍA DOCENTE: Dr. Steban Ilich Zerpa INTEGRANTES Jara Carranza, Jacqueline Luján Sifuentes, Carlos Julca León, Gina Mantilla Germán, Fiorella Mendoza Mendoza, Gabriela León Matos, Ana Isabel Micha Burgos, Jorge Luis Leytón Móstiga, Olenka Lucas Segura, Natali Mundaca Rafael, Jorge

2
PROTEÍNAS

3
Definición: Son las macromoléculas más versátiles que se conocen.
Son polímeros lineales constituidos a partir de monómeros llamados aminoácidos.

4
Características: Contienen un amplio surtido de grupos funcionales, los cuales son responsables del amplio espectro de funciones que presentan las proteínas. Se encuentran formadas únicamente por aminoácidos L. Pueden intercambiar entre sí y con otras macromoléculas biológicas para formar asociaciones complejas. Algunas proteínas son muy rígidas, funcionando como elementos estructurales; otras manifiestan una flexibilidad limitada, facilitándoles el ensamblaje y la formación de unidades complejas.

5
Funciones: La función de una proteína es directamente dependiente de su estructura tridimensional. Pudiendo funcionar como: Catalizadores. Moléculas transportadoras y de almacén de otras moléculas. Brindan apoyo mecánico. Brindan protección inmunológica. Generan movimiento. Transmiten impulsos nerviosos. Controlan el crecimiento y la diferenciación.

6
Estructura: Estructura Primaria
Se encuentra formada por una secuencia de aminoácidos enlazados mediante la unión del grupo -carboxilo de un aminoácido al grupo -amino de otro aminoácido por medio de un enlace peptídico. Es importante porque nos permite conocer el mecanismo de acción de una proteína, así como determinar su estructura tridimensional.

7
Estructura Secundaria
Se aprecia en el plegamiento de las cadenas polipeptídicas en estructuras regulares como la hélice alfa, la hoja plegada beta, los giros y los bucles . Los responsables de estos plegamientos en las proteínas son los enlaces puentes de hidrógeno que se forman entre los grupos R de los diversos aminoácidos.

8
EstructuraTerciaria Es una característica de las proteínas solubles en agua. Es una estructura compacta con un núcleo no polar. Las cadenas polipeptídicas se pliegan, de modo que sus cadenas hidrofóbicas laterales estén en el interior y sus cadenas polares, cargadas, estén en la superficie.

9
Estructura Cuaternaria
Se refiere al ordenamiento espacial de las subunidades y la naturaleza de sus interacciones. Las cadenas polipeptídicas se pueden ensamblar en estructuras de múltiples subunidades, formando complejos moleculares.

10
LOS ENLACES QUÍMICOS ENLACES COVALENTES:
Los enlaces químicos permiten la interacción entre los átomos. Se clasifican en: ENLACES COVALENTES: Son los enlaces más fuertes que están presentes en los compuestos bioquímicos. Consiste en un par de electrones que se comparten entre átomos adyacentes. Se debe gastar una cantidad considerable de energía para romper los enlaces covalentes. Se puede compartir más de un par de electrones entre dos átomos para formar un enlace covalente múltiple.

11
ENLACES NO COVALENTES:
Son más débiles que los covalentes, pero son esenciales para los procesos bioquímicos, tales como la formación del doble helicoide. Los tipos fundamentales de enlaces covalentes son: INTERACCIONES ELECTROSTÁTICAS: Un grupo cargado de una molécula puede atraer a otro grupo con carga opuesta de otra molécula. La energía de una interacción electrostática viene dada por la Ley de Coulomb: Por convención, se considera que la interacción atrayente tiene una energía negativa. E= kq1q2/Dr2

12
PUENTES DE HIDRÓGENO: Son los responsables de la formación de los pares de bases específicos de la doble hélice del ADN. El átomo de hidrógeno está parcialmente compartido entre dos átomos relativamente electronegativos. Son mucho más débiles que los enlaces covalentes. Son responsables de muchas de las propiedades del agua. INTERACCIONES DE VAN DER WAALS: La distribución de cargas no es perfectamente simétrica. Las energías asociadas con las interacciones de van der Waals son bastante pequeñas.

13
EL AGUA EN REACCIONES ENZIMATICAS
El esta reacción, el equilibrio esta mas desplazada hacia la hidrólisis que hace a síntesis, por ello la biosíntesis de los enlaces peptídicos requeridos un aporte de energía libre. Lo cual los enlaces peptídicos son muy estables cinéticamente, la vida de un enlace peptídicos en disolución acuosa en ausencia de un catalizador es cercana a los 1000 años.

14
ENZIMAS -Son catalizadores biológicos
-Moléculas de gran interés que determinan la pauta de las transformaciones químicas. -Intervienen en la transformación de un tipo de energía otra.

15
CARACTERÍSTICAS DE LAS ENZIMAS
Poder catalítico: Tiene lugar en el centro activo de la enzima, muy eficaces en catalizar muchas reacciones químicas Especificidad: Capacidad de unirse específicamente a un gran numero de moléculas.

16
APOENZIMA ENZIMA SIN COFACTOR = APOENZIMA
Enzima que no puede llevar a cabo su acción catalítica desprovista de los cofactores necesarios Esta enzima esta catalíticamente inactiva, hasta que se le une el cofactor adecuado. ENZIMA SIN COFACTOR = APOENZIMA

17
HOLOENZIMAS -Es la enzima completamente activa catalíticamente.
APOENZIMA + COFACTOR = HOLOENZIMA *Cofactor: Moléculas indispensables en la actividad catalítica de la enzima, tipos: -Iones metálicos -Coenzimas

18
ENERGÍA LIBRE

19
Hay dos conceptos básicos sobre la energía libre:
La diferencia de energía libre ( G); relación entre los reactantes y los productos que determina si la reacción será espontánea. La energía requerida para iniciar la reacción, esta propiedad determina la velocidad.
; relación entre los reactantes y los productos que determina si la reacción será espontánea. La energía requerida para iniciar la reacción, esta propiedad determina la velocidad.")
20
VARIACIÓN DEL “G” Si “G” es negativo Si “G” es nulo
La reacción puede tener lugar espontáneamente. Reacción exergónica. La reacción no puede tener lugar espontáneamente. Reacción endergónica. Si “G” es nulo El sistema está en equilibrio.

21
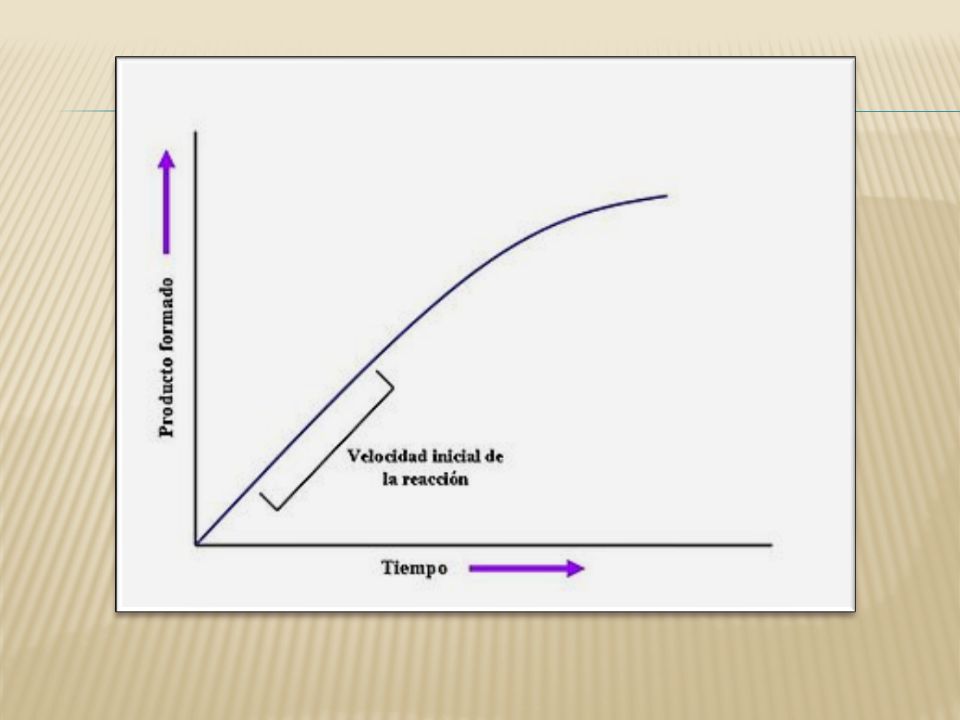
CINÉTICA ENZIMÁTICA Estudia la velocidad de reacción catalizadas por enzimas. Las dos propiedades cinéticas más importantes de una enzima son: el tiempo que tarda en saturarse con un sustrato en particular y la máxima velocidad de reacción que pueda alcanzar.

23
Constante de Michaelis – Menten (Km)
Corresponde a la concentración de sustrato con la cual la velocidad de reacción enzimática alcanza un valor igual a la mitad de la velocidad máxima. Es independiente a la concentración de enzima y de sustrato.

25
FACTORES QUE INFLUYEN EN LA ACTIVIDAD ENZIMÁTICA
TIEMPO DE INCUBACIÓN Velocidad puede disminuir: -por agotamiento de substrato - desnaturalización de la enzima

26
CONCENTRACIÓN DE ENZIMA
Cuando el substrato se encuentra en exceso, a concentraciones saturantes, la velocidad de reacción en proporcional a la concentración de la enzima.

27
CONCENTRACIÓN DEL SUBSTRATO (C.S)
Efecto de la concentración del substrato sobre la velocidad de la reacción enzimática.

28
EFECTO DEL PH Curvas de la actividad de las fosfatasas con las variaciones del PH. La curva de la actividad de las enzimas con el pH presenta generalmente una forma a campanada. EFECTO DE LA TEMPERATURA La temperatura óptima es el resultado de dos procesos: el incremento de la velocidad de reacción, con la temperatura y el incremento de la desnaturalización térmica de la enzima. La mayor parte de la enzima se inactiva a temperaturas superiores 55-60º. Efecto de la temperatura sobre la actividad enzimática.

29
MODELO DE MICHAELIS MENTEN
Relaciona la velocidad de catálisis con la concentración de sustrato. Michaelis y Menten propusieron que las reacciones catalizadas enzimáticamente ocurren en dos etapas: En la primera etapa se forma el complejo enzima-sustrato En la segunda, el complejo enzima-sustrato da lugar a la formación del producto, liberando el enzima libre:

30
ECUACIÓN DE MICHAELIS MENTEN
Modelo de la Reacción v1 = k1 [E] [S] v2 = k2 [ES] v3 = k3 [ES] Se obtiene la ecuación de Michaelis Menten:

31
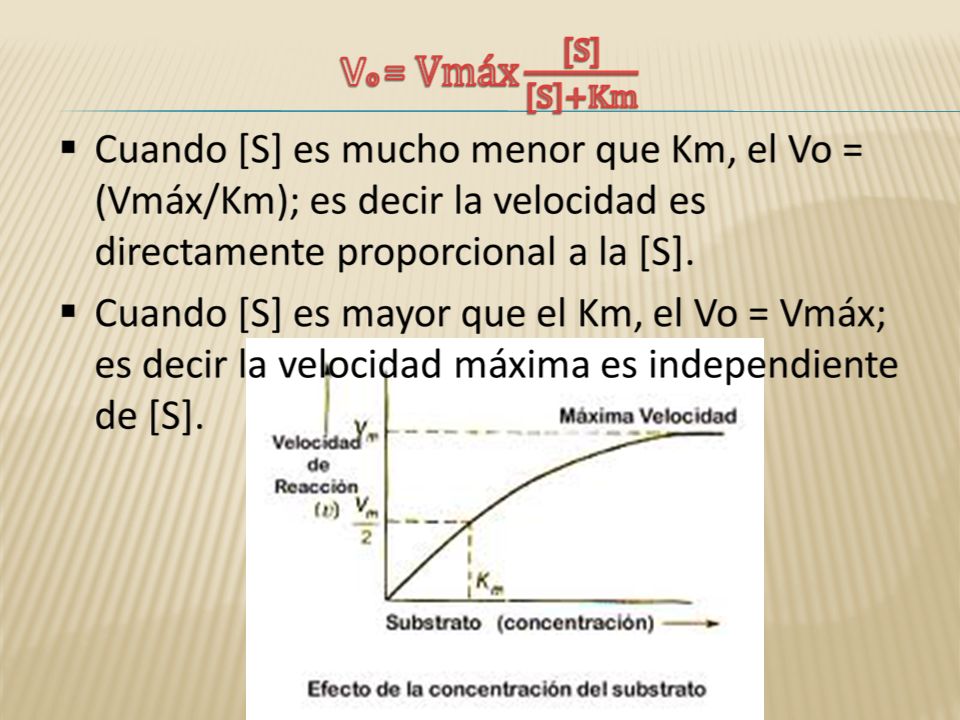
Está ecuación explica:
Sustrato mucho menor que Km la velocidad es directamente proporcional a la concentración del substrato. Sustrato mucho mayor que Km V=Vmax

32
La velocidad máxima (Vmáx):
Es la velocidad cuando todos los centros activos están ocupados con sustrato. Se la considera como una velocidad teórica, pues nunca es alcanzada en la realidad.

33
A [S] bajas ([S]<<KM) la mayor parte de la enzima se encuentra libre ([ET] ≅[E]) y entonces𝑉= 𝑘 𝑐𝑎𝑡 𝐾 𝑀 𝐸 𝑆 con lo que kcat/KM pasa a ser una constante de velocidad para la reacción entre el sustrato y la enzima libre. Esta relación es importante ya queproporciona una medida directa de la eficiencia y la especificidad de la enzima. Muestralo que una enzima y el sustrato pueden realizar cuando se dispone de lugaresenzimáticos abundantes, y permite una comparación directa de la eficacia de unaenzima respecto a distintos sustratos. kcat/KM tiene un valor límite dado por la teoría de la difusión de 108 a 109 (mol/L)-1s-1. Permite comparar eficiencia de una enzima con diferentes sustratos Un ejemplo de esto es la triosa fosfato isomerasa con kcat/KM=2,4x108 (mol/L)-1s-1. Kcat/Km es una medida de la eficacia catalítica, por lo que permite comparar enzimas. A mayor valor de Kcat/Km, más eficaz será el enzima
 la mayor parte de la enzima se encuentra libre ([ET] ≅[E]) y entonces𝑉= 𝑘 𝑐𝑎𝑡 𝐾 𝑀 𝐸 𝑆 con lo que kcat/KM pasa a ser una constante de velocidad para la reacción entre el sustrato y la enzima libre. Esta relación es importante ya queproporciona una medida directa de la eficiencia y la especificidad de la enzima. Muestralo que una enzima y el sustrato pueden realizar cuando se dispone de lugaresenzimáticos abundantes, y permite una comparación directa de la eficacia de unaenzima respecto a distintos sustratos. kcat/KM tiene un valor límite dado por la teoría de la difusión de 108 a 109 (mol/L)-1s-1. Permite comparar eficiencia de una enzima con diferentes sustratos. Un ejemplo de esto es la triosa fosfato isomerasa con kcat/KM=2,4x108 (mol/L)-1s-1. Kcat/Km es una medida de la eficacia catalítica, por lo que permite comparar enzimas. A mayor valor de Kcat/Km, más eficaz será el enzima.")
34
SUSTRATOS MÚLTIPLES EN LAS REACCIONES BIOQUÍMICAS
La mayor parte de las reacciones en los sistemas biológicos incluyen normalmente dos sustratos y dos productos y se representa mediante la reacción bisustrato: 𝐴+𝐵⇌𝑃+𝑄 Las reacciones de múltiples sustratos se dividen en dos clases: DESPLAZAMIENTO SECUENCIAL: Todos los sustratos se unen al enzima antes de que se libere cualquier producto. consecuentemente, se forma un complejo ternario entre la enzima y ambos sustratos. MECANISMO SECUENCIAL ORDENADO: entra el primer sustrato y luego el segundo; sale el primer producto y luego el otro. E EA EAB EPQ EQ A catálisis B P Q b) SECUENCIAL AL AZAR: cualquiera de los 2 sustratos puede entrar primero y cualquiera de los 2 productos puede salir primero. A E B EA EB EAB P Q EQ EP EPQ II)REACION DE DOBLE DESPLAZAMIENTO (Ping-pong):uno o mas productos se liberan antes de que todos los sustratos se unan al enzima. La característica definitiva de estas reacciones es la existencia de un intermediario del enzima sustituido, en el cual la enzima se modifica temporalmente. A E EA EP P B EB EQ Q Mecanismo ping-pong: la enzima reacciona con el primer sustrato entrega el primer producto, toma el segundo sustrato y entrega el segundo producto.
 SECUENCIAL AL AZAR: cualquiera de los 2 sustratos puede entrar primero y cualquiera de los 2 productos puede salir primero. A. E. B. EA. EB. EAB. P. Q. EQ. EP. EPQ. II)REACION DE DOBLE DESPLAZAMIENTO (Ping-pong):uno o mas productos se liberan antes de que todos los sustratos se unan al enzima. La característica definitiva de estas reacciones es la existencia de un intermediario del enzima sustituido, en el cual la enzima se modifica temporalmente. A. E. EA. EP. P. B. EB. EQ. Q. Mecanismo ping-pong: la enzima reacciona con el primer sustrato entrega el primer producto, toma el segundo sustrato y entrega el segundo producto.")
35
INHIBICIÓN ENZIMATICA
Es la disminución de la actividad de un enzima por la presencia de un inhibidor. Inhibidor: Son moléculas o iones que se unen a la enzima y la hacen disminuir su velocidad.

36
TIPOS DE INHIBICIÓN INHIBICION REVERSIBLE:
Hay una rápida disociación del complejo enzima_inhibidor. Se caracterizan por su constante de equilibrio (Ki). Hay 2 tipos : Competitiva No competitiva
. Hay 2 tipos : Competitiva. No competitiva.")
37
INHIBICION REVERSIBLE COMPETITIVA
El inhibidor compite por el centro activo con el sustrato. Aumenta el valor de [I], incrementa el valor del Km. Alcanza la Vmax , al incrementar la cantidad de sustrato que llega a superar la inhibición.

38
INHIBICION REVERSIBLE NO COMPETITIVA
El sustrato se puede unir al complejo enzima_inhibidor. No cambia el valor del Km . Vmax disminuye. No se puede superar si se incrementa la concentración del sustrato.

39
2. INHIBICIÓN IRREVERSIBLE
El inhibidor queda unido fuertemente a la enzima. Algunos son importantes fármacos como: la penicilina y la aspirina. Los inhibidores irreversibles se dividen en 3 categorías: Reactivos específicos de grupo Análogos de sustrato Inhibidores suicidas

40
Reactivos Específicos:
Reaccionan con un grupo R específicos de aminoacidos. El DIPF modifica 1 de los 28 aminoacidos “tripsina”

41
Marcadores de afinidad:
Son moléculas que tiene parecida estructura al sustrato, que modifican los residuos del centro activo de la enzima. Sustratos Suicidas: Modifican al centro activo de la enzima. El inhibidor se une a la enzima y se transforma. N, N –dimetilpropargilamina es un ejemplo de ello.

42
LA PENICILINA Es un antibiótico que consta de un anillo de tiazolidina fundido a un anillo ß-lactámico (muy lábil), al cual se une un grupo R variable, mediante enlace peptídico.
, al cual se une un grupo R variable, mediante enlace peptídico.")
43
¿ Como inhibe la penicilina el crecimento bacteriano?
El peptidoglicano de la pared celular consta de cadenas de polisacáridos lineales entrecruzados con péptidos cortos. La penicilina inhibe la transpeptidasa responsable del entrecruzamiento bloqueándolo. Entonces la transpeptidasa se inhibe irreversiblemente y no hay síntesis de pared celular. La penicilina actúa como inhibidor suicida.

44
LAS VITAMINAS SON CON FRECUENCIA PRECURSORAS DE LOS COENZIMAS
Son moléculas orgánicas necesarias en pequeñas cantidades en la dieta. Su deficiencia puede generar enfermedades. Se clasifican en: * vitaminas hidrosolubles * vitaminas liposolubles

45
VITAMINAS HIDROSOLUBLES
COENZIMA TIPO DE REACCION TIPICA CONSECUENCIA DE LA DEFICIENCIA Tiamina (B1) Tiamina pirofosfato Transferencia de aldehido Beriberi(perdida de peso, problemas del corazon, disfunciones neurologicas) Rivoflavina (B2) Flavina ina dinucleotido (FAD) Oxidacion-reduccion Queliosis y estomatiti angular (lesiones de la boca), dermatitis Piridoxina (B6) Piridoxal fosfato Transferencia de grupo hacia o desde los aminoácidos Depresión, confusión, convulsiones Ácido nicotínico (niacina) Nicotinamida adenina dinucleótido (NAD+) Pelagra (dermatitis, depresión, diarrea ) Ácido pantotenico Coenzima A Transferencia de grupos acilo Hipertensión Biotina Complejos biotina-lisina (biocitina) Carboxilacion dependiente de ATP y transferencia del grupo carboxilo Salpullido en las cejas, dolor muscular, fatiga (poco común) Ácido fólico Tetrahirofolato Transferencia de compuestos de un carbono, síntesis de timina Anemia, defectos en el tubo neural (en el desarrollo) B12 5’-Desoxiadenosil-cobalamina Transferencia de grupos metilo, reestructuraciones intramoleculares Anemia perniciosa, anemia, acidosis metilmalonica C (ácido ascórbico) Antioxidante Escorbuto (encías inflamadas y sangrantes, hemorragias subdermicas)
 Tiamina pirofosfato. Transferencia de aldehido. Beriberi(perdida de peso, problemas del corazon, disfunciones neurologicas) Rivoflavina (B2) Flavina ina dinucleotido (FAD) Oxidacion-reduccion. Queliosis y estomatiti angular (lesiones de la boca), dermatitis. Piridoxina (B6) Piridoxal fosfato. Transferencia de grupo hacia o desde los aminoácidos. Depresión, confusión, convulsiones. Ácido nicotínico (niacina) Nicotinamida adenina dinucleótido (NAD+) Pelagra (dermatitis, depresión, diarrea ) Ácido pantotenico. Coenzima A. Transferencia de grupos acilo. Hipertensión. Biotina. Complejos biotina-lisina (biocitina) Carboxilacion dependiente de ATP y transferencia del grupo carboxilo. Salpullido en las cejas, dolor muscular, fatiga (poco común) Ácido fólico. Tetrahirofolato. Transferencia de compuestos de un carbono, síntesis de timina. Anemia, defectos en el tubo neural (en el desarrollo) B12. 5’-Desoxiadenosil-cobalamina. Transferencia de grupos metilo, reestructuraciones intramoleculares. Anemia perniciosa, anemia, acidosis metilmalonica. C (ácido ascórbico) Antioxidante. Escorbuto (encías inflamadas y sangrantes, hemorragias subdermicas)")
46
VITAMINAS LIPOSOLUBLES
FUNCION DEFICIENCIA A En la visión, crecimiento, reproducción Ceguera nocturna, lesión en la cornea, daños en el tubo digestivo y aparato respiratorio D Regulación del metabolismo del calcio y del fosfato Raquitismo (niños), deformaciones en el esqueleto, crecimiento retrasado Osteomalacia(adultos), huesos blandos y curvados E Antioxidante Inhibición de la producción de esperma, lesiones en músculos y nervios (poco común) K Coagulación sanguínea Hemorragias subdermicas
, deformaciones en el esqueleto, crecimiento retrasado. Osteomalacia(adultos), huesos blandos y curvados. E. Antioxidante. Inhibición de la producción de esperma, lesiones en músculos y nervios (poco común) K. Coagulación sanguínea. Hemorragias subdermicas.")
Presentaciones similares













![1 KM 1 1 Vo= Vmax [S] + Vmax Y = m x + b = m = b.](/2/161459/big_thumb.jpg "1 KM 1 1 Vo= Vmax [S] + Vmax Y = m x + b = m = b.>")





