Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Tema 10. El código genético

2
Tema 10. El código genético
en tripletes, no solapado, inicio y fin de mensaje, dirección de lectura • supresión intergénica • descifre del código genético: homopolímeros, heteropolímeros, prueba de unión al triplete, copolímeros con repetición • hipótesis del tambaleo • el código genético es universal • características del código genético

3
el código genético? 60’s! 1 N (41) -> 4 aa 2 N (42) -> 16 aa
secuencia de 4 nucleótidos -> secuencia de 20 aminoácidos 1 N (41) -> 4 aa 2 N (42) -> 16 aa 3 N (43) -> 64 aa 4 N (44) -> 256 aa DOS VEN MAS QUE UNO DOS VEN MAS QUE UNO -1 DOS VNM ASQ UEU NO DOS VNM ASQ UEU NO 3 adiciones o 3 deleciones restauran la fase, pauta o marco de lectura DOS VMA SQU EUN O DOS VMA SQU EUN O DOS VAS QUE UNO
 -> 4 aa. 2 N (42) -> 16 aa. 3 N (43) -> 64 aa. 4 N (44) -> 256 aa. DOS VEN MAS QUE UNO. DOS VEN MAS QUE UNO -1. DOS VNM ASQ UEU NO. DOS VNM ASQ UEU NO adiciones o 3 deleciones restauran la. fase, pauta o marco. de lectura. DOS VMA SQU EUN O. DOS VMA SQU EUN O -3. DOS VAS QUE UNO.")
4
el código genético ‘se lee’ en tripletes, Brenner y Crick 1961
Figure: 13-02 Frameshift Mutations The effect of frameshift mutations on a DNA sequence with the repeating triplet sequence GAG. (a) The insertion of a single nucleotide shifts all subsequent triplet reading frames. (b) The insertion of three nucleotides changes only two triplets, but the frame of reading is then reestablished to the original sequence. Crick y Brenner trabajaron analizando mutantes rII de T4 (silvestre: calvas de lisis normales; mutantes: calvas de lisis más grandes) y mutagenizando con proflavina (agente mutagénico intercalante) que añade o elimina un nucleótido. Esto mismo se demostró posteriormente con mutantes de la lisozima, proteína de secuencia conocida (Streisinger). Francois Jacob and Jaques Monod discovered mRNA in 1961, but work of others paved the way Brenner (early 60’s) confirmed degeneracy, triplet, nonoverlapping nature of code
 The insertion of a single nucleotide shifts all subsequent triplet reading frames. (b) The insertion of three nucleotides changes only two triplets, but the frame of reading is then reestablished to the original sequence. Crick y Brenner trabajaron analizando mutantes rII de T4 (silvestre: calvas de lisis normales; mutantes: calvas de lisis más grandes) y mutagenizando con proflavina (agente mutagénico intercalante) que añade o elimina un nucleótido. Esto mismo se demostró posteriormente con mutantes de la lisozima, proteína de secuencia conocida (Streisinger). Francois Jacob and Jaques Monod discovered mRNA in 1961, but work of others paved the way. Brenner (early 60’s) confirmed degeneracy, triplet, nonoverlapping nature of code.")
5
el código genético no es solapado
Fig Suzuki Duferencia entre un código solapado y uno no solapado. EL ejemplo es un código de tripletes (el real). Ojo: el numero que acompaña a ‘aa’ sólo hace referencia a la posición que ocupa el aa en la cadena polipeptídica, no a su naturaleza química. Datos para NO solapado: no hay restricción en la disposición de aa contiguos un cambio de un nucleótido sólo afecta a un aa Messages in mRNA Shown are the results of mRNA messages read in (a) overlapping and (b) nonoverlapping three-letter codes. In an overlapping code, each letter is part of three different three-letter words (as indicated for the letter G in blue); in a nonoverlapping code, each letter is part of only one three-letter word. Each "word" in mRNA uses a code of three bases together (triplet or codon). The non-overlapping nature of the mRNA code was deciphered in the 1960's.
. Ojo: el numero que acompaña a ‘aa’ sólo hace referencia a la posición que ocupa el aa en la cadena polipeptídica, no a su naturaleza química. Datos para NO solapado: no hay restricción en la disposición de aa contiguos. un cambio de un nucleótido sólo afecta a un aa. Messages in mRNA. Shown are the results of mRNA messages read in (a) overlapping and (b) nonoverlapping three-letter codes. In an overlapping code, each letter is part of three different three-letter words (as indicated for the letter G in blue); in a nonoverlapping code, each letter is part of only one three-letter word. Each word in mRNA uses a code of three bases together (triplet or codon). The non-overlapping nature of the mRNA code was deciphered in the 1960 s.")
6
genes solapados The code is nonoverlapping, but mRNA may have different initation points (viruses like phage X174, SV40, phage G4 Problem: single mutation may affect multiple products

7
signos de puntuación no comas no espacios
- sí principio y fin de mensaje NO signos de puntuación porque: longitud total del mRNA de la proteína de la cápsida del VMT se corresponde con el número de aa de la proteína

8
señales de inicio y terminación
AUG: codón de iniciación N-formil-Metionina (fMet) en bacterias metionina en eucariotas también codifica para metioninas internas UAG, UGA, UAA: codones de fin de mensaje, de parada, de terminación o codones sin sentido en el interior generan una mutación sin sentido mutaciones llamadas: ámbar, ópalo y ocre, respectivam. hay mutantes que suprimen terminaciones prematuras: mutante supresor mutaciones llamadas: ámbar, ópalo y ocre producen paradas internas en las proteínas
 en bacterias. metionina en eucariotas. también codifica para metioninas internas. UAG, UGA, UAA: codones de fin de mensaje, de parada, de terminación o codones sin sentido. en el interior generan una mutación sin sentido. mutaciones llamadas: ámbar, ópalo y ocre, respectivam. hay mutantes que suprimen terminaciones prematuras: mutante supresor. mutaciones llamadas: ámbar, ópalo y ocre producen paradas internas en las proteínas.")
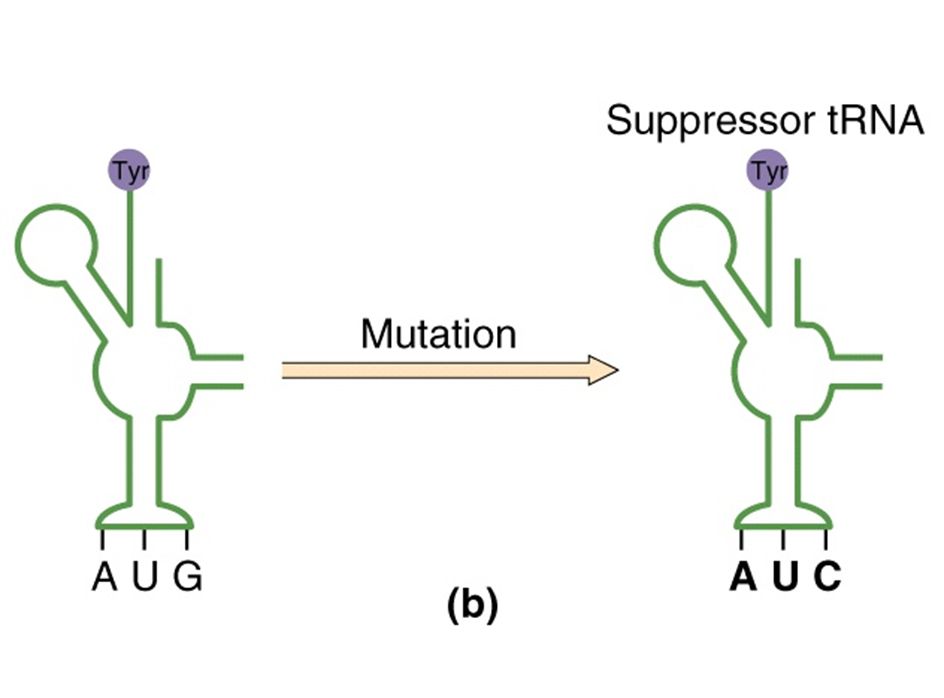
9
supresión intergénica
mutación sin sentido silvestre tRNA supresor tRNA supresores de mutaciones sin sentido, descubiertos por Brenner y col. Suppressor mutation Occur at sites different from the original mutation and mask or compensate for the initial mutation without reversing it. Intragenic suppressors occur on the same codon; e.g., nearby addition restores a deletion Intergenic suppressors occur on a different gene. Intergenic suppressor genes: Many function in mRNA translation. Each suppressor gene works on only one type of nonsense, missense, of frameshift mutation. Suppressor genes often encode tRNAs, which possess anti-codons that recognize stop codons and insert an amino acid. Three classes of tRNA nonsense suppressors, one for each stop codon (UAG, UAA, UGA). tRNA suppressor genes coexist with wild type tRNAS. tRNA suppressors compete with release factors, which are important for proper amino acid chain termination. Small number of read-through polypeptides are produced; tandem stop codons (UAGUAG) are responsible for correct translation termination.
. tRNA suppressor genes coexist with wild type tRNAS. tRNA suppressors compete with release factors, which are important for proper amino acid chain termination. Small number of read-through polypeptides are produced; tandem stop codons (UAGUAG) are responsible for correct translation termination.")
11
dirección lectura S. Ochoa 1965
20 A + 1C -> AAAAAAAAAAAAAAAAAAAACAAAAAAAAAAAAAAAAAACAAAAAAA - ribonucleasa pancreática -> 5’-AAAAAAAAAAAAAAAAAAAAC- 3’ síntesis de proteínas -> NH2 -Lys-Lys-Lys-Lys-Lys-Lys-Asn- COOH lectura polarizada ribonucleasa pancreática: corta tras pirimidinas en dirección 5’->3’

12
descifre del código genético
one of Nirenberg's many laboratory notebooks Descifre: Acción y efecto de descifrar. one of Nirenberg's many laboratory notebooks

13
M. Grunberg-Manago y S. Ochoa, 1955
Pasos previos para el desciframiento del código genético Polynucleotide Phosphorylase The reaction catalyzed by the enzyme polynucleotide phosphorylase. Note that the equilibrium of the reaction favors the degradation of RNA but can be “forced” in the direction favoring synthesis. Polynucleotide Phosphorylase: In 1955, Marianne Grunberg-Manago and Severo Ochoa reported the isolation of an enzyme that catalyzed the synthesis of RNA. Their work built upon the earlier work of Jerard Hurwitz & J.J. Furth who performed experiments to see if isolated E. coli protein fractions could polymerize radioactively labeled nucleotides. Later Grunberg-Manago & Ochoa tested a protein fraction that could make RNA. For this work, Ochoa shared the 1959 Nobel Prize in Medicine with Arthur Kornberg (who received the Prize for his work on DNA polymerase I). This enzyme could convert ribonucleoside diphosphates into RNA: (RNA)n + NDP --> (RNA)n+1 + Pi However, the enzyme had a number of unsettling properties. It did not need a template; and, it could use as little as 1 NDP or as many as 4 NDPs as substrate. In fact, the sequence of the product RNA depended entirely on the number and concentration of substrate NDPs. These are not the properties of an enzyme that must faithfully copy the genetic material for expression! We now know that Grunberg-Manago and Ochoa had isolated the enzyme polynucleotide phosphorylase which usually catalyzes the breakdown of RNA - not its synthesis! i.e., its a ribonuclease. Experiment drew on methods devised by Paul Zamecnik a few years earlier. Zamecnik had developed a cell-free in vitro translation system which was capable of directing the synthesis of radioactively labelled protein. This system consisted of a membrane-free cell supernatant, ATP, GTP, radioactively labelled amino acids, and RNA. * The cell-free suspension was obtained by gently braking E. coli cells by grinding them with finely powdered alumina. Cell "sap" remains. "Sap"= DNA, mRNA, ribosomes, cellular enzymes, and Adenosine Triphosphate (ATP). ATP should provide adequate energy for the incorporation of amino acids into protein. Paul Zamecnik proved that the ribosome was the site of protein synthesis and developed a cell-free in vitro translation system, which was capable of directing the synthesis of radioactively labeled protein. Nirenberg used this method in his experiments. As early as the 1950s, researchers realized that protein biosynthesis does not require the integrity of the cell and can continue after cell disruption. Eventually, Zamecnik and his colleagues (Littlefield et al., 1955) made a real cell-free ribosomal system of protein synthesis (translation) based on mitochondria-free cytoplasmic extracts of animal cells (rat liver). Soon after, cell-free translation systems based on bacterial (Escherichia coli) extracts were made by German and American groups (Schachtschabel and Zillig, 1959; Lamborg and Zamecnik, 1960; Tissiéres et al., 1960). Ribosomes in all of those systems, however, were programmed with endogenous mRNA; they were simply reading the messages to which they had been already attached at the time of cell disruption. sistema libre de células para la síntesis de proteínas, P. Zamecnik 1955
. This enzyme could convert ribonucleoside diphosphates into RNA: (RNA)n + NDP --> (RNA)n+1 + Pi. However, the enzyme had a number of unsettling properties. It did not need a template; and, it could use as little as 1 NDP or as many as 4 NDPs as substrate. In fact, the sequence of the product RNA depended entirely on the number and concentration of substrate NDPs. These are not the properties of an enzyme that must faithfully copy the genetic material for expression! We now know that Grunberg-Manago and Ochoa had isolated the enzyme polynucleotide phosphorylase which usually catalyzes the breakdown of RNA - not its synthesis! i.e., its a ribonuclease. Experiment drew on methods devised by Paul Zamecnik a few years earlier. Zamecnik had developed a cell-free in vitro translation system which was capable of directing the synthesis of radioactively labelled protein. This system consisted of a membrane-free cell supernatant, ATP, GTP, radioactively labelled amino acids, and RNA. * The cell-free suspension was obtained by gently braking E. coli cells by grinding them with finely powdered alumina. Cell sap remains. Sap = DNA, mRNA, ribosomes, cellular enzymes, and Adenosine Triphosphate (ATP). ATP should provide adequate energy for the incorporation of amino acids into protein. Paul Zamecnik proved that the ribosome was the site of protein synthesis and developed a cell-free in vitro translation system, which was capable of directing the synthesis of radioactively labeled protein. Nirenberg used this method in his experiments. As early as the 1950s, researchers realized that protein biosynthesis does not require the integrity of the cell and can continue after cell disruption. Eventually, Zamecnik and his colleagues (Littlefield et al., 1955) made a real cell-free ribosomal system of protein synthesis (translation) based on mitochondria-free cytoplasmic extracts of animal cells (rat liver). Soon after, cell-free translation systems based on bacterial (Escherichia coli) extracts were made by German and American groups (Schachtschabel and Zillig, 1959; Lamborg and Zamecnik, 1960; Tissiéres et al., 1960). Ribosomes in all of those systems, however, were programmed with endogenous mRNA; they were simply reading the messages to which they had been already attached at the time of cell disruption. sistema libre de células. para la síntesis de proteínas, P. Zamecnik")
14
poly U -> Phe-Phe-Phe... => UUU = Phe Nirenberg y Matthaei 1961
Figure: 13-T01 Incorporation of 14C-Phenylalanine into Protein

15
Nirenberg y Matthaei, 1961 Método desarrollado por Nirenberg y Matthaei para identificar el aminoácido especificado por un homopolímero. Tubo con sistema de traducción libre de células con 1 aa marcado radiactivamente y 19 aa no marcados.

16
homopolímeros, Nirenberg y Matthaei 1962
UUUUUUU.. -> Phe-Phe-Phe-Phe... CCCCCCC > Pro-Pro-Pro-Pro... AAAAAAA... -> Lys-Lys-Lys-Lys... GGGGGGG... -> Gly-Gly-Gly-Gly... UUU Phe CCC Pro AAA Lys GGG Gly

17
heteropolímeros, Nirenberg, Matthaei y Ochoa
proporción utilizada 1A:5C (1/6A:5/6C) Figure: 13-04 A Mixed Copolymer Experiment Results and interpretation of a mixed copolymer experiment where a ratio of 1A:5C is used (1/6A:5/6C).
 Figure: A Mixed Copolymer Experiment. Results and interpretation of a mixed copolymer experiment where a ratio of 1A:5C is used (1/6A:5/6C).")
18
prueba de unión al triplete Nirenberg y Leder, 1964
Figure: 13-05 Triplet Binding Assay An example of the triplet binding assay. The UUU triplet acts as a codon, attracting the complementary tRNAphe anticodon AAA. Nirenberg and Leder (1964)- Triplet assay with radioactively labeled amino acids led to assigment of 50 codons reacción en presencia de alta concentración de MgCl para que las subunidades ribosomicas se ensamblen en presencia de tripletes
- Triplet assay with radioactively labeled amino acids led to assigment of 50 codons. reacción en presencia de alta concentración de MgCl para que las subunidades ribosomicas se ensamblen en presencia de tripletes.")
19
Nirenberg y Leder, 1964 Figure 15.9 Nirenberg and Leder used ribosome-bound tRNAs to provide additional information about the genetic code.

20
Prueba de unión al triplete

21
Figure: 13-T02 Amino Acid Assignments to Specific Trinucleotides Derived from the Triplet Binding Assay se identifican así 50 de los 64 codones

22
copolímeros con repetición, Khorana 1964
Figure: 13-06 The Conversion of di-, tri-, and Tetranucleotides into Repeating Copolymers The conversion of di-, tri-, and tetranucleotides into repeating copolymers. The triplet codons that are produced in each case are shown. Khorana- Repeating Copolymers Note that neither GAUA and GUAA resulted in any amino acids being incorporated

23
Figure: 13-T03 Amino Acids Incorporated Using Repeated Synthetic Copolymers of RNA aquí ya se identifican los 64 codones

24
el código genético 5’-mRNA-3’ Standard genetic code.
Figure: 13-07 Triplet Coding Dictionary The coding dictionary. AUG encodes methionine, which initiates most polypeptide chains. All other amino acids except tryptophan, which is encoded only by UGG, are represented by two to six triplets. The triplets UAA, UAG, and UGA are termination signals and do not encode any amino acids. Standard genetic code. The standard genetic code is composed of 64 triplet codons. The left-hand column indicates the nucleotide found at the first (5') position of the codon; the top row indicates the nucleotide found at the second (middle) position of the codon; and the right column indicates the nucleotide found at the third (3') position of the codon. The codon AUG specifies methionine (Met) and is also used to initiate protein synthesis. STOP indicates a termination codon. The genetic code is not randomly assigned. If an amino acid is coded by several codons, they often share the same sequence in the first two positions and differ in the third position. Such assignment is accomplished by the design of wobble position, but "the evolutionary dynamic that shaped the code remains a mystery".
 position of the codon; the top row indicates the nucleotide found at the second (middle) position of the codon; and the right column indicates the nucleotide found at the third (3 ) position of the codon. The codon AUG specifies methionine (Met) and is also used to initiate protein synthesis. STOP indicates a termination codon. The genetic code is not randomly assigned. If an amino acid is coded by several codons, they often share the same sequence in the first two positions and differ in the third position. Such assignment is accomplished by the design of wobble position, but the evolutionary dynamic that shaped the code remains a mystery .")
26
hipótesis del tambaleo, Crick 1966
Fig Suzuki hipótesis del tambaleo la base tercera del codón (extremo 3’) puede emparejarse con varias de la primera posición (extremo 5’) del tRNA Ej: tRNASer puede reconocer dos cosdones en el mRNA: UCC y UCU -> explica parte de la redundancia del código genético (el resto es por la existencia de tRNA isoaceptores)
 puede emparejarse con varias de la primera posición (extremo 5’) del tRNA. Ej: tRNASer puede reconocer dos cosdones en el mRNA: UCC y UCU. -> explica parte de la redundancia del código genético (el resto es por la existencia de tRNA isoaceptores)")
27
Codon-Anticodon Base-Pairing Rules
Figure: 13-T04 Codon-Anticodon Base-Pairing Rules * un mismo tRNA puede llegar a leer hasta 3 codones (cuando la I esta en el 5’ del anticodón)! Degeneracy and the Wobble Hypothesis (Crick, 1966) Degeneracy occurs at 3rd position Hydrogen bond at 3rd position is not as constrained as positions1 & 2 Allows for minimum of 30 tRNA (actually for bacteria, up to 50 for plants and animals
! Degeneracy and the Wobble Hypothesis (Crick, 1966) Degeneracy occurs at 3rd position. Hydrogen bond at 3rd position is not as constrained as positions1 & 2. Allows for minimum of 30 tRNA (actually for bacteria, up to 50 for plants and animals.")
28
redundancia del código genético (> 1 codón / aa), se explica por:
existencia de > 1 tRNA / aa (50 tRNA y 20 aa) hipótesis del tambaleo
 hipótesis del tambaleo.")
29
el código genético es universal
Demostración de la universalidad del código genético: clonación de genes procarióticos en eucariontes y viceversa

30
universalidad del flujo de la información genética
Fig Suzuki traducción del mRNA de la hemoglobina de conejo en oocitos de rana (Xenopus) produce hemoglobina de conejo
 produce hemoglobina de conejo.")
31
The standard genetic code applies to most, but not all, cases.
Exceptions have been found in the mitochondrial DNA of many organisms and in the nuclear DNA of a few lower organisms. Some examples are given in the table. For more information, visit The Genetic Codes - From NCBI.

32
Figure: 13-T05 Exceptions to the Universal Code

33
Frecuencia de aa, número de codones y uso de letras

34
el código genético = la clave genética
se ha descifrado el genoma de una sp. se ha secuenciado el genoma de una sp.

35
el código genético lineal se lee en tripletes (codón-codones)
no solapado no puntuación interna sí señales de inicio y parada redundante (> 1 codón / aa) no ambiguo (1 codón -> 1 aa) ordenado universal (casi) 1. code is linear sequence of ribonucleotides derived from DNA template 2. code is comprised of triplet codons 3 ribonucleotides=1 amino acid 3. unambiguous- 1 a.a. per triplet codon 4. degenerate- one a.a. may be specified by multiple codons 5. ordered- can group related codons (see 4) together 6. certain codons are start or stop initiate and terminate -punctuation only at beginnings and ends of sentence 7. no internal punctuation 8. nonoverlapping- each ribonucleotide is part of only one triplet 9. code is universal (almost)
 no ambiguo (1 codón -> 1 aa) ordenado. universal (casi) 1. code is linear sequence of ribonucleotides derived from DNA template. 2. code is comprised of triplet codons 3 ribonucleotides=1 amino acid. 3. unambiguous- 1 a.a. per triplet codon. 4. degenerate- one a.a. may be specified by multiple codons. 5. ordered- can group related codons (see 4) together. 6. certain codons are start or stop initiate and terminate -punctuation only at beginnings and ends of sentence. 7. no internal punctuation. 8. nonoverlapping- each ribonucleotide is part of only one triplet. 9. code is universal (almost)")
36
traducción y código genético
problemas capítulo 2 traducción y código genético 2ª ed.: 2.6, 3ª ed.: 2.9, 1 h de clase: enero 2010 (junto a los problemas del tema 8 y 9)
")
Presentaciones similares