Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Bioquímica del Vanadio, Cromo y Níquel
Maria Magdalena Moya y Beatriz Rayo Grado de química 2013/2014

2
INDICE VANADIO CROMO NÍQUEL Abundancia En los tunicados Amavadina
Haloperoxidasas de vanadio Diabetes CROMO Abundancia en la tierra Bioquímica Esencialidad del Cromo en los mamíferos. Consumo Alimentos ricos en Cromo Cromodulina Función biológica de la cromodulina NÍQUEL Bioquímica Ureasas Estructura Mecanismo Hidrogenasas CO deshidrogenasas Acettil CoA sintetasa Metil-CoM reductasa

3
VANADIO Abundancia. No existe concentrada en los minerales, pero esta en casi todos lo suelos En agua se encuentra en muy bajas concentraciones El ión más común: VO43-, vanadato con EO= 5 reducible a 4 y 3. Esencial para los humanos Tóxico: interfiere en bioquimica del ión PO43- La reducción del vanadato EO= 5 a 4 es facil, pero para la reduccion de 5 a 3 ya es más dificil. Se sabe que el vanadio es esencial para los humanos pero no se conocen cuales son las especies biologicamente activas ni sus funciones.

4
En los tunicados. Se acumulan en los vacuolas en forma de V(III):
[V(SO4)(H2O)5]+ [V(H2O)6]3+ En H2O el V (V) se reduce a V (III) dificilmente mediante reductores energéticos (tunicromo) V (III) forma complejos con tunicromo: se cree que el [V(H2O)6] es el dador electronico terminal de una cadena de transporte electronico
(H2O)5]+ [V(H2O)6]3+ En H2O el V (V) se reduce a V (III) dificilmente mediante reductores energéticos (tunicromo) V (III) forma complejos con tunicromo: se cree que el [V(H2O)6] es el dador electronico terminal de una cadena de transporte electronico.")
5
Amavadina. Complejo V (IV)[VL2]2-. IC= 8. L, actúa como tetradentado y se une a V con: Carboxilato N imino O de OH- (desprotonado) En las setas amanita Complejo muy estable hongo extrae V con facilidad del suelo selectivamente Función similar a oxidasas de cobre Las oxidasas azules de obre cataliza la reacción de oxidación de alcoholes primarios a aldehídos, usando O2 reduciendo a H2O2.
![Amavadina. Complejo V (IV)[VL2]2-. IC= 8. L, actúa como tetradentado y se une a V con: Carboxilato.](http://slideplayer.es/slide/3280171/11/images/5/Amavadina.+Complejo+V+%28IV%29%EF%83%A0%5BVL2%5D2-.+IC%3D+8.+L%2C+act%C3%BAa+como+tetradentado+y+se+une+a+V+con%3A+Carboxilato..jpg "N imino. O de OH- (desprotonado) En las setas amanita. Complejo muy estable hongo extrae V con facilidad del suelo selectivamente. Función similar a oxidasas de cobre. Las oxidasas azules de obre cataliza la reacción de oxidación de alcoholes primarios a aldehídos, usando O2 reduciendo a H2O2.")
6
Amavadina. Estructura del complejo bis [N- hidroxi- imino- diacetato] vanadio (IV).
![Amavadina. Estructura del complejo bis [N- hidroxi- imino- diacetato] vanadio (IV).](http://slideplayer.es/slide/3280171/11/images/6/Amavadina.+Estructura+del+complejo+bis+%5BN-+hidroxi-+imino-+diacetato%5D+vanadio+%28IV%29..jpg "Amavadina. Estructura del complejo bis [N- hidroxi- imino- diacetato] vanadio (IV).")
7
Haloperoxidasas de vanadio.
Catalizan reacción de oxidación de iones haluro con agua para dar ácido hipoaloso: H2O2 + X- + H+ H2O + HOX En algas marinas y hongos En todo este ciclo catalítico es V (V), IC= 5 Bipiramide triangular, VO3-. La carga se neutraliza mediante enlaes de hidrógeno entre oxígenos y restos de Aá.
, IC= 5 Bipiramide triangular, VO3-. La carga se neutraliza mediante enlaes de hidrógeno entre oxígenos y restos de Aá.")
8
Estructura haloperoxidasas de vanadio.
Posición ecuatorial: 3 O de los oxo no protéicos Posición axial: N imidazol de histidina. 1.- Representación de tipo cinta de la estructura molecular de la cloroperoxidasa de vanadio. 2.- Representación centro activo.

9
Mecanismo haloperoxidasas de vanadio
Centro activo de la cloroperoxidasa de vanadio: azido complejo, forma nativa. La formación del complejo con el inhibidor implica la sustitución del ligando OH- por N3-. Por lo tanto, es muy probable que durante el mecanismo de la reación el grupo peroxo ocupe el mismo sitio. El siguiente paso es el ataque nucleofilo del cloruro sobre el peroxo coordinado.

10
Diabetes. La deficiencia de insulina o resistencia celular a su función conduce a diabetes. Mimetizadores de insulina: Na3VO4, VOSO4·H2O, VOSO4·3H2O. Se ha demostrado que el vanadio representa un elemento asociado a la regulación del metabolismo de la glucosa y mejora la sensibilidad del receptor a la insulina. También se conoce que ciertos complejos de Vanadio reducen los niveles de glucemia en diabéticos 2. Tomando en consideración que la proteína tirosin fosfatasa se considera como un regulador negativo de la vía de señalización de la insulina, se sugiere que el vanadio puede acentuar este evento gracias a la capacidad de inhibir la acción de estas fosfatasas e incrementar la fosforilación de la tirosina. La insulina es una hormona esencial para el metabolismo de carbohidratos i grasas. Segregada en pancreas en respuesta a altos niveles de glucosa en sangre. Promove la captura de glucosa por el higado o tejido muscular. Mimetizar= imitar La glucemia es la medida de concentración de glucosa

11
CROMO Abundancia en la tierra.
Elemento abundante en la corteza terrestre (110ppm). Puesto 24 Por delante de: Cu y Zn. Cr(III) ESENCIAL Estado de oxidación más común y estable Presente en muchos silicatos Única mena importante la cromita FeCr2O4 Elemento escaso en el agua de mar (10-4 ppm) Cr(VI) TOXICO Ion cromato CrO4 2- El cromo es un elemento relativamente abundante en la corteza terrestre. Con un contenido de unos 110 oom, ocupa el puesto 24 en abundancia, por delante de metales tan importantes en lo sistemas biologicos como el cobre o el cinc. Su estado de oxidaciion mas estable y comun el (III). El Cr(III) se encuentra como un constituyente minoritario de muchos silicatos y su única mena importante es la cromita, un oxido mixto con estructura de espinela. Por otro lado, el cromo es un elemento escaso en el agua de mar, tendremos concentraciones de 10^-4 ppm y solo se encuentra como ion cromato, cuyo estado de oxidación del Cr es Cr(VI) muy tóxico.
. Puesto 24. Por delante de: Cu y Zn. Cr(III) ESENCIAL. Estado de oxidación más común y estable. Presente en muchos silicatos. Única mena importante la cromita FeCr2O4. Elemento escaso en el agua de mar (10-4 ppm) Cr(VI) TOXICO. Ion cromato CrO4 2- El cromo es un elemento relativamente abundante en la corteza terrestre. Con un contenido de unos 110 oom, ocupa el puesto 24 en abundancia, por delante de metales tan importantes en lo sistemas biologicos como el cobre o el cinc. Su estado de oxidaciion mas estable y comun el (III). El Cr(III) se encuentra como un constituyente minoritario de muchos silicatos y su única mena importante es la cromita, un oxido mixto con estructura de espinela. Por otro lado, el cromo es un elemento escaso en el agua de mar, tendremos concentraciones de 10^-4 ppm y solo se encuentra como ion cromato, cuyo estado de oxidación del Cr es Cr(VI) muy tóxico.")
12
Estabilidad relativa de los estados de oxidación
Bioquímica de Cromo. 1959 Se establece la esencialidad del cromo para a vida. Bioquímica muy limitada. Ha empezado a conocerse en los últimos años. Comparación con el Mo. Mo Cr Bioquímica amplia Bioquímica limitada Poco abundante Abundante Estabilidad relativa de los estados de oxidación Mo(VI) EO más estable Cr(VI) muy oxidante (acción mutagénica y cancerígena) Mo(V) y Mo(IV) EO importantes Cr (V) y Cr(IV) no forma especies estables en disolución Cr (III) común y estable Cr (II) fuerte reductor Bioquímica de Mo química redox Mo(VI) Mo(IV) Bioquímica del Cr NO química redox NO química acido-base La esencialidad del cromo para la vida de los mamíferos, empezó a establecerse en No obstante su bioquímica es muy limitada y solo a empezado a conocerse en los últimos años. En la siguiente tabla vamos a comparar la bioquímica de Mo (elemento situado en el mismo grupo que el Cr) con la del cromo. El Mo tienen una bioquímica mucho mas amplia que el Cromo, a pesar de ser un metal mucho menos abundante, lo que podría parecer extraño ya que los seres vivos se adaptan a la necesidad de elementos abundantes en su entorno. Las causas de estas situación se encuentran en la estabilidad relativa de los diferentes EO. (comentar EO tabla) Una vez conocida la estabilidad de los diferentes estados de oxidación, podemos entender que la bioquímica del molibdeno se base en reacciones red-ox entre Mo(VI) y( Mo(IV). Mientras que para el cromo, no parece posible una bioquímica redox. Tampoco se llevarán a cabo reacciones ácido-base en complejos de Cr(III) dado el carácter inerte de este a la sustitución de los ligandos.
 EO más estable. Cr(VI) muy oxidante (acción mutagénica y cancerígena) Mo(V) y Mo(IV) EO importantes. Cr (V) y Cr(IV) no forma especies estables en disolución. Cr (III) común y estable. Cr (II) fuerte reductor. Bioquímica de Mo química redox. Mo(VI) Mo(IV) Bioquímica del Cr. NO química redox. NO química acido-base. La esencialidad del cromo para la vida de los mamíferos, empezó a establecerse en No obstante su bioquímica es muy limitada y solo a empezado a conocerse en los últimos años. En la siguiente tabla vamos a comparar la bioquímica de Mo (elemento situado en el mismo grupo que el Cr) con la del cromo. El Mo tienen una bioquímica mucho mas amplia que el Cromo, a pesar de ser un metal mucho menos abundante, lo que podría parecer extraño ya que los seres vivos se adaptan a la necesidad de elementos abundantes en su entorno. Las causas de estas situación se encuentran en la estabilidad relativa de los diferentes EO. (comentar EO tabla) Una vez conocida la estabilidad de los diferentes estados de oxidación, podemos entender que la bioquímica del molibdeno se base en reacciones red-ox entre Mo(VI) y( Mo(IV). Mientras que para el cromo, no parece posible una bioquímica redox. Tampoco se llevarán a cabo reacciones ácido-base en complejos de Cr(III) dado el carácter inerte de este a la sustitución de los ligandos.")
13
Bioquímica del Cromo. Una única función biológica.
Tipo de función semejante al Ca Cr mucho más limitado. APLIFICACIÓN DE LA SEÑAL DE LA INSULINA INSULINA Hormona importante en la regulación del metabolismo de los carbohidratos y los lípidos. Su función es la de favorecer la incorporación de glucosa de la sangre hacia las células

14
Esencialidad del Cromo en los mamíferos.
Experimento. Ratas alimentadas con una dieta con déficit de cromo incapaces de responder a un aumento de glucosa en sangre. Alteración del metabolismo de glucosa Administración de sales de Cr(III) DESAPARECE LA PATOLOGÍA IMP: Evitar el contacto del anima con objetos de acero inoxidable. La demostración de la necesidad de cromo en los mamíferos, se demostró hace más de cuarenta años mediante experimentos clásicos conanimales de laboratorio. En el experimento se demostró que ratas alimentadas con una dieta deficiente de este elemento desarrollaban una patología concreta, eran incapaces de responder de manera eficaz a un aumento de glucosa en sangre, (tenían el metabolismo de la glucosa alterado). No obstante, esta patología desaparecía cuando se les administraba simples sales de cromo (III). Acero inoxidable Aleación de hierro Carbono 0,03-1,075% Cromo 10-12%
 DESAPARECE LA PATOLOGÍA. IMP: Evitar el contacto del anima con objetos de acero inoxidable. La demostración de la necesidad de cromo en los mamíferos, se demostró hace más de cuarenta años mediante experimentos clásicos conanimales de laboratorio. En el experimento se demostró que ratas alimentadas con una dieta deficiente de este elemento desarrollaban una patología concreta, eran incapaces de responder de manera eficaz a un aumento de glucosa en sangre, (tenían el metabolismo de la glucosa alterado). No obstante, esta patología desaparecía cuando se les administraba simples sales de cromo (III). Acero inoxidable. Aleación de hierro. Carbono. 0,03-1,075% Cromo %")
15
Tris(picolinato)cromo(III)
Consumo de Cromo Seres vivos [Cr(III)] muy baja Humanos. [Cr(III)] = 120nM Dieta normal Ingestión diaria de entre 0,01 y 1,2 mg Cr(III) Absorción muy pequeña alrededor del 0,5% Las necesidades diarias de este elementos son mínimas. Difícil encontrar personas con deficiencia de Cr(III). Cantidades superiores a 200mg tiene efectos tóxicos. Complejos metálicos de cromo mejor absorción (más eficaz) Tris(picolinato)cromo(III) Absorbe entre 2% y un 5% de Cr(III) no tiene actividad biológica. Solo es una fuente de Cr(III) Se comercializa como suplemento alimenticio. PBL: efectos perjudiciales La mayor parte de los seres vivos contienen cromo, pero generalmente en muy bajas concentraciones. Existen organismos como la levadura de cerveza que es capaz de acumular el cromo en sus células, pero se desconoce en que forma química se encuentra y cuales son sus funciones. En los seres humanos las concentraciones de cromo significativas fisiológicamente son de 120 nM. Una dieta normal permite la ingestión diaria de 0,01 y 1,2 mg Cr(III) del que se absorbe alrededor del 0,5%. Estos datos indican que las necesidades diarias de cromo son muy pequeñas, por lo que no es común encontrar personas con un déficit de este. Solo, se ha observado en personas alimentadas por exclusivamente por vía intravenosa, en las que han aparecido sintomas parecidos a los de la diabetes no dependiente de la insulina. Estos síntomas desaparecen al administrar pequeñas cantidades de compuestos de cromo (III). Aunque el cromo en la dieta se absorbe muy mal, algunos complejos metálicos son mucho más eficaces. El [Cr(pic)3] llego a creerse que tenia actividad biológica, es un compuesto presente en suplementos alimenticios. No obstante, estudios recientes cuestionan su utilidad porque la cesión del cromo del complejo alas biomoléculas, necesita de una reducción previa del cromo a Cr(II), que al oxidarse rapidamente por la aciión del oxigeno molecular puede inducir la formación de los peligrosos radicales hidroxilo. (Pic = 2-carboxipiridina)
] muy baja. Humanos. [Cr(III)] = 120nM. Dieta normal Ingestión diaria de entre 0,01 y 1,2 mg Cr(III) Absorción muy pequeña alrededor del 0,5% Las necesidades diarias de este elementos son mínimas. Difícil encontrar personas con deficiencia de Cr(III). Cantidades superiores a 200mg tiene efectos tóxicos. Complejos metálicos de cromo mejor absorción (más eficaz) Tris(picolinato)cromo(III) Absorbe entre 2% y un 5% de Cr(III) no tiene actividad biológica. Solo es una fuente de Cr(III) Se comercializa como suplemento alimenticio. PBL: efectos perjudiciales. La mayor parte de los seres vivos contienen cromo, pero generalmente en muy bajas concentraciones. Existen organismos como la levadura de cerveza que es capaz de acumular el cromo en sus células, pero se desconoce en que forma química se encuentra y cuales son sus funciones. En los seres humanos las concentraciones de cromo significativas fisiológicamente son de 120 nM. Una dieta normal permite la ingestión diaria de 0,01 y 1,2 mg Cr(III) del que se absorbe alrededor del 0,5%. Estos datos indican que las necesidades diarias de cromo son muy pequeñas, por lo que no es común encontrar personas con un déficit de este. Solo, se ha observado en personas alimentadas por exclusivamente por vía intravenosa, en las que han aparecido sintomas parecidos a los de la diabetes no dependiente de la insulina. Estos síntomas desaparecen al administrar pequeñas cantidades de compuestos de cromo (III). Aunque el cromo en la dieta se absorbe muy mal, algunos complejos metálicos son mucho más eficaces. El [Cr(pic)3] llego a creerse que tenia actividad biológica, es un compuesto presente en suplementos alimenticios. No obstante, estudios recientes cuestionan su utilidad porque la cesión del cromo del complejo alas biomoléculas, necesita de una reducción previa del cromo a Cr(II), que al oxidarse rapidamente por la aciión del oxigeno molecular puede inducir la formación de los peligrosos radicales hidroxilo. (Pic = 2-carboxipiridina)")
16
Alimentos ricos en Cromo.
Los alimentos más ricos en cromo son, junto ala levadura de cerveza, las carnes, quesos, algunos hogos, el zumo de remolacha, pimienta negra, nueces y espárragos. En menor proporción el ajo y las alcachofas. Además, también se ha demostrado que cuanto más elaborado es el producto, menores es la cantidad de cromo. Son el ejemplo del trigo, la harina o el pan blanco.

17
Cromodulina. Único complejo metálico de cromo que presenta actividad biológica. LMWCr = sustancia de baja masa molecular que se une al cromo Cromodulina, Por su similitud con la calmodulina (bioquímica del Ca; ) Cadena polipeptídica. Formada: mayoritariamente por restos aspartato y glutamato. Cuatro centros Cr(III) Complejo bastante estable Efecto cooperativo No se conoce la estructura Estudios espectroscópicos y químicos sugieren que la estructura es: Complejo tetranuclear Ligandos puente oxo o hidroxo entre los iones metálicos La cromodulina es el único compuesto de cromo que se ha aislado de los seres vivos y que presenta actividad biológica , llamado en un principio LMWCr. Es un complejo metálico de un péptido pequeño, extraido el hígado de conejo y vacas. Más de la mitad del total de restos de aminoacidos de la cadena polipetidica de la cromodulina son restos aspartato y gutamato, aunque también resenta glicina y cisteina. A pesar de ser un peptido pequeño, es capaz de unirse a cuatro Cr(III), formando un complejo bastante estable. La unión de los iones metálicos de Cr(III) presenta un acusado efecto cooperativo, de forma que las únicas especies existentes en disolución en cantidades significativas son el ligando libre (apocromodulina) y la especie totalmente metalada (holocromodulina). Aún no se conoce la estructura de la cromodulina pero una serie de estudios químicos y espectroscópicos sugieren que la estructura es de complejos tetranucleares con ligandos puente
 Cadena polipeptídica. Formada: mayoritariamente por restos aspartato y glutamato. Cuatro centros Cr(III) Complejo bastante estable. Efecto cooperativo. No se conoce la estructura. Estudios espectroscópicos y químicos sugieren. que la estructura es: Complejo tetranuclear. Ligandos puente oxo o hidroxo entre los. iones metálicos. La cromodulina es el único compuesto de cromo que se ha aislado de los seres vivos y que presenta actividad biológica , llamado en un principio LMWCr. Es un complejo metálico de un péptido pequeño, extraido el hígado de conejo y vacas. Más de la mitad del total de restos de aminoacidos de la cadena polipetidica de la cromodulina son restos aspartato y gutamato, aunque también resenta glicina y cisteina. A pesar de ser un peptido pequeño, es capaz de unirse a cuatro Cr(III), formando un complejo bastante estable. La unión de los iones metálicos de Cr(III) presenta un acusado efecto cooperativo, de forma que las únicas especies existentes en disolución en cantidades significativas son el ligando libre (apocromodulina) y la especie totalmente metalada (holocromodulina). Aún no se conoce la estructura de la cromodulina pero una serie de estudios químicos y espectroscópicos sugieren que la estructura es de complejos tetranucleares con ligandos puente.")
18
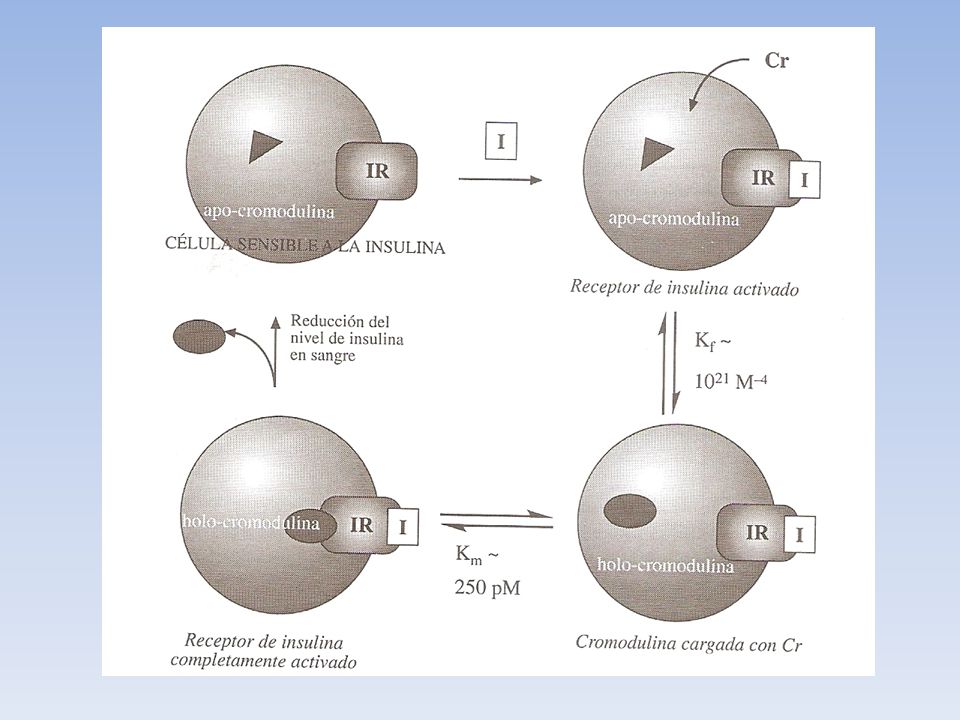
Función biológica de la cromodulina.
Modo de acción. Estudios in vivo han permitido identificar la función biológica de la cromodulina y proponer un modelo detallado de su modo de acción. La cromodulina incrementa la capacidad de las células para metabolizar glucosa cuando son estimuladas por la insulina. Las células son capaces de almacenar el oligopéptido en su forma de ligando libre, sin metal. Cuando se produce un aumento de glucosa en sangre, las células del páncreas sintetizan insulina que se vierte en el corriente sanguíneo. La insulina se une a los receptores específicos (IR) de la membrana celular, provoca un cambio estructural (que activa al receptor, que se transforma en un aproteina quinasa que transmite la señal de la insulina) y favorece la entrada de cromo en el interior de la célula que se une a la apocromodulina. Una vez se ha incorporado el sufriente cromo como para formar la holocromodulina , esta se une al receptor de la insulina y lo mantiene en su forma activa, amplificando su actividad como quinasa (modificación de proteinas por fosofrilación) Cuando desaparece la señal, se reducen los niveles de insulina en la sangre, se relaja la conformación del receptor y la cromodulina es expulsada de la célula, siendo excretada finalmete a traves de la orina.
 de la membrana celular, provoca un cambio estructural (que activa al receptor, que se transforma en un aproteina quinasa que transmite la señal de la insulina) y favorece la entrada de cromo en el interior de la célula que se une a la apocromodulina. Una vez se ha incorporado el sufriente cromo como para formar la holocromodulina , esta se une al receptor de la insulina y lo mantiene en su forma activa, amplificando su actividad como quinasa (modificación de proteinas por fosofrilación) Cuando desaparece la señal, se reducen los niveles de insulina en la sangre, se relaja la conformación del receptor y la cromodulina es expulsada de la célula, siendo excretada finalmete a traves de la orina.")
19
NIQUEL EO: 0,I,II,III y IV más común y con gran tendencia a formar complejos Oh EO=II. Finales de los 70. esencialidad del Ni es sistemas bacterianos. 5 Tipos de enzimas que contienen Níquel. Enzima Centro Ni Reacción Ureasas Dímero de Ni(II), alto espín Urea NH3 + CO2 Hidrogenasa Centro monomérico 2H+ + 2e- ↔H2 Metil-CoM reductasa F-430 CH3-CoM CH4 Acettil CoA sintetasa NiFe3S4 (?) CO + [CH3] + CoA + 2e- CH3COCoA CO deshidrogenasa ? CO + [aceptor] CO2 El níquel, cuyo nombre es de origrn alemán y significa cobre del diablo, se puede encontrar en sus compuestos en los EO=0,I,II,III,IV. Entre ellos, el estado de oxidación II es mucho más abitual y muestra una gran tendencia a formar comlejos Oh, aunque tambien encontramos estructuras bipiramides triangular o cuadradas. A finales de la década de los 70, pudo establecerse que el níquel era un elemento traza esencial para diferentes sistemas bacterianos. Hasta el presente, se han identificado 5 tipos diferentes de enzimas que presentan níquel. Estas enzimas de níquel, están especialmente presentes en el metabolismo de bacterias anaeróbicas, como por ejemplo, las llamadas bacterias metanogénicas, que pueden crecer en presencia de una mezcla H2 y CO2, siendo los únicos organismos capaces de producir metano como producto final de su metabolismo. En la tabla podemos ver que las diferentes enzimas no presentan aspectos comunes, ni desde el punto de vista de los complejos de níquel involucrados ni por el tipo de reacción que catalizan. El níquel muestra, por ejemplo, química redox en las hidrogenasas pero no en las ureasas. Por ello mas adelante, explicaremos las funciones del níqule separadamente. Predominantes en el metabolismo de bacterias anaeróbicas Bacterias metanogénicas Las diferentes enzimas, no presentan aspectos comunes: Complejos de Ni involucrados, distintos. Reacciones catalizadas distintas.
, alto espín. Urea NH3 + CO2. Hidrogenasa. Centro monomérico. 2H+ + 2e- ↔H2. Metil-CoM reductasa. F-430. CH3-CoM CH4. Acettil CoA sintetasa. NiFe3S4 ( ) CO + [CH3] + CoA + 2e- CH3COCoA. CO deshidrogenasa. CO + [aceptor] CO2. El níquel, cuyo nombre es de origrn alemán y significa cobre del diablo, se puede encontrar en sus compuestos en los EO=0,I,II,III,IV. Entre ellos, el estado de oxidación II es mucho más abitual y muestra una gran tendencia a formar comlejos Oh, aunque tambien encontramos estructuras bipiramides triangular o cuadradas. A finales de la década de los 70, pudo establecerse que el níquel era un elemento traza esencial para diferentes sistemas bacterianos. Hasta el presente, se han identificado 5 tipos diferentes de enzimas que presentan níquel. Estas enzimas de níquel, están especialmente presentes en el metabolismo de bacterias anaeróbicas, como por ejemplo, las llamadas bacterias metanogénicas, que pueden crecer en presencia de una mezcla H2 y CO2, siendo los únicos organismos capaces de producir metano como producto final de su metabolismo. En la tabla podemos ver que las diferentes enzimas no presentan aspectos comunes, ni desde el punto de vista de los complejos de níquel involucrados ni por el tipo de reacción que catalizan. El níquel muestra, por ejemplo, química redox en las hidrogenasas pero no en las ureasas. Por ello mas adelante, explicaremos las funciones del níqule separadamente. Predominantes en el metabolismo de bacterias anaeróbicas. Bacterias metanogénicas. Las diferentes enzimas, no presentan aspectos comunes: Complejos de Ni involucrados, distintos. Reacciones catalizadas distintas.")
20
Ureasas. Presentes en algas, hongos y bacterias.
Cataliza la hidrólisis de urea. Hidrólisis espontanea Hidrólisis catalizada (velocidad mayor) La ureasa es una enzima que se conoce desde hace mas de 100 años, se encuentra en alngunas plantas, hoongos y bacterias cuyo metabolismo requiere hidrolizar urea. La descomposición no catalítica de urea implica una eliminación directa del amoníaco, mientras que la hidrólisis catalizada, forma en una primer etapa carbamato, que posteriormente se degrada de manera espontanea a bicarbonato y amonio. Aumentando así la velocidad de la reacción catorce ordenes de magnitud.
 La ureasa es una enzima que se conoce desde hace mas de 100 años, se encuentra en alngunas plantas, hoongos y bacterias cuyo metabolismo requiere hidrolizar urea. La descomposición no catalítica de urea implica una eliminación directa del amoníaco, mientras que la hidrólisis catalizada, forma en una primer etapa carbamato, que posteriormente se degrada de manera espontanea a bicarbonato y amonio. Aumentando así la velocidad de la reacción catorce ordenes de magnitud.")
21
Estructura ureasas. Constituida por 6 subunidades idénticas
Con dos iones Ni(II) cada una. Esfera de coordinación del Ni(II) Propuesta a base de estudios espectroscópicos. Constituida por O, N pero no S. Geometría: Oh distorsionada
 cada una. Esfera de coordinación del Ni(II) Propuesta a base de estudios espectroscópicos. Constituida por O, N pero no S. Geometría: Oh distorsionada.")
22
Mecanismo de la ureasa. El mecanismo implica en primer lugar la formación de un enlace entre el grupo carbonilo de la urea y el Ni(1). Seguido de la formación simultanea del enlace Ni(2)-NH2 y un ataque nucleofilo del OH puente al átomo de carbono de la urea. De este modo, el enlace Ni –OH-Ni se debilita activando el ataque nucleofílico del OH al cabonilo de la urea y al Ni del complejo. (adición –Eliminación) Eliminamos el primer grupo amino de la urea y formando amonio.
. Seguido de la formación simultanea del enlace Ni(2)-NH2 y un ataque nucleofilo del OH puente al átomo de carbono de la urea. De este modo, el enlace Ni –OH-Ni se debilita activando el ataque nucleofílico del OH al cabonilo de la urea y al Ni del complejo. (adición –Eliminación) Eliminamos el primer grupo amino de la urea y formando amonio.")
23
Hidrogenasas. Catalizan la generación o consumo de hidrógeno gaseoso. 2H+ + 2e- ↔ H2 Papel importante en: la fijación del N2 Procesos de fermentación de biomasa. Presente en microorganismos aeróbicos y anaeróbicos Tipos de hidrogenasas. (Ferredoxinas 4:4 Fe:S) [Fe]-hidrogenasas [Fe/Ni]-hidrogenasas [Fe/Ni/Se]-hidrogenasas Pueden catalizar la reacción en una única dirección o en ambas. Inhibidores de hidrogenasas: moléculas gaseosas O2, CO, NO, acetileno H2 FUENTE DE ENERGÍA PRODUCTO FINAL DE PROCESOS REDUCTIVOS Son enzimas que catalizan la generación o el consumo de hidrógeno gaseoso, con la participación de donadores o aceptores electrónicos. Esta reacción juega un papel importante en la fijación de N2 así como durante la fermentación de biomasa con producción final de metano. Tanto organismos aeróbicos como anaeróbicos presentan estas hidrogenasa. La importancia de esta reaccion se debe a que el H2 puede utilizarse como fuente de energía o aparecer como producto final en procesos productivos. En general todas las hidrogenasas contienen cúmulos de Fe/S del tipo de las ferredoxinas 4:4. Algunas no necesitan ningún otro sistema metálico, [Fe]-hidrogenasas, para su funcionamiento. Otras contienen Ni y otras Ni y Se (en forma de seleniocisteina).
 [Fe]-hidrogenasas. [Fe/Ni]-hidrogenasas. [Fe/Ni/Se]-hidrogenasas. Pueden catalizar la reacción en una única dirección o en ambas. Inhibidores de hidrogenasas: moléculas gaseosas O2, CO, NO, acetileno. H2 FUENTE DE ENERGÍA PRODUCTO FINAL DE PROCESOS REDUCTIVOS. Son enzimas que catalizan la generación o el consumo de hidrógeno gaseoso, con la participación de donadores o aceptores electrónicos. Esta reacción juega un papel importante en la fijación de N2 así como durante la fermentación de biomasa con producción final de metano. Tanto organismos aeróbicos como anaeróbicos presentan estas hidrogenasa. La importancia de esta reaccion se debe a que el H2 puede utilizarse como fuente de energía o aparecer como producto final en procesos productivos. En general todas las hidrogenasas contienen cúmulos de Fe/S del tipo de las ferredoxinas 4:4. Algunas no necesitan ningún otro sistema metálico, [Fe]-hidrogenasas, para su funcionamiento. Otras contienen Ni y otras Ni y Se (en forma de seleniocisteina).")
24
Forma activa (más reducida)
Estructura hidrogenasas Ni-Fe. Resuelta por difracción de rayos X Estructura de la hidrogenasa Ni-Fe de Desulfovibrio gigas (forma inactiva) Dos subunidades distintas Centro activo Fe-Ni Diferentes cluster [4Fe-4S] y [3Fe-4S] Centro activo Forma inactiva Fe (II) d6 de alto espín. Ni(III), d7 Ligando Cys68 protonado Forma activa (más reducida) Ni(II) , d8 - perdida del ligando puente hidroxo Fe(II) pentacoordinado; N(II) geometría cuadrada Mediante difracción de rayos X se ha resuelto la estrcutura de la hidrogenasa desulfovibrio gigas, aislada aerobcamente en una forma inactiva. Se trata de una proteina heterodimérica que consta de dos subunidades. Una con el centro activo Ni-Fe y la otra subunidad con una serie de clusters de Fe-S. A partir de estos estudios se ha podido determinar la estructura del centro activo. El centro activo está constituido por un complejo heteribinuclear formado por un átomo de níquel y uno de hierro. El entorno de coordinación del Fe presenta geometría Oh distorsionada. El Fe esta coordinado por dos grupos CN- y un ligando CO y dos grupos S-Cys puente que enlazan el Fe con el Ni. Para completar la esfera de coordinación tenemos un tercer ligando puente X = hidroxo o un oxo. El Ni tiene una geometría de piramide cuadrada que se completa, a parte de col los ligando puente, con la presencia de por dos S-Cis. La estructura de la forma activa e inactiva presenta una serie de diferencias: (diapo)
 Dos subunidades distintas. Centro activo Fe-Ni. Diferentes cluster [4Fe-4S] y [3Fe-4S] Centro activo. Forma inactiva. Fe (II) d6 de alto espín. Ni(III), d7. Ligando Cys68 protonado. Forma activa (más reducida) Ni(II) , d8. - perdida del ligando puente hidroxo. Fe(II) pentacoordinado; N(II) geometría cuadrada. Mediante difracción de rayos X se ha resuelto la estrcutura de la hidrogenasa desulfovibrio gigas, aislada aerobcamente en una forma inactiva. Se trata de una proteina heterodimérica que consta de dos subunidades. Una con el centro activo Ni-Fe y la otra subunidad con una serie de clusters de Fe-S. A partir de estos estudios se ha podido determinar la estructura del centro activo. El centro activo está constituido por un complejo heteribinuclear formado por un átomo de níquel y uno de hierro. El entorno de coordinación del Fe presenta geometría Oh distorsionada. El Fe esta coordinado por dos grupos CN- y un ligando CO y dos grupos S-Cys puente que enlazan el Fe con el Ni. Para completar la esfera de coordinación tenemos un tercer ligando puente X = hidroxo o un oxo. El Ni tiene una geometría de piramide cuadrada que se completa, a parte de col los ligando puente, con la presencia de por dos S-Cis. La estructura de la forma activa e inactiva presenta una serie de diferencias: (diapo)")
25
Mecanismo catalítico de las hidrogenasas Ni-Fe.
El mecanismo de este proceso catalítico es aun hoy en día un tema de debate. La reacción se inicia con la coordinación de la molécula de hidrógeno al ion Fe(II) de a forma activa (NiSi 1934 ( numero: corresponde a la frecuencia de vibración de tensión del enlace CO). Que el hidrogeno se fije primero al Fe demuestra la existencia de que haya hidrogenasas que solo prefente Fe. El dihidrogen resultante,Ni-R1940, pierde un electrón. Como resultado el Ni(II) se oxida a Ni(III) y el ligando tiolato Cys350 ataca nucleoficamente al H2 coordinado. Se produce la rotura heterolitica del H-H La cisteina se protona y el ligando hidruro unido al Fe se convierte en un ligando puente. En el siguiente paso, la Cys 530 se vuelve a protonar, dando ugar a la especie intermedia NiC1952, lo que induce una transformación redox interna en el complejo. El ligandi hidruro reduce el Ni(III) a Ni (I) y el proton resultante se vuelve a fjar a la Cys530. Por último, la cesión de un segundo electrón a través de un cluster Fe-S y la perdida simultanes del protón de la Cys 530 conlleva a la regeneración de la especie inicial .
 de a forma activa (NiSi 1934 ( numero: corresponde a la frecuencia de vibración de tensión del enlace CO). Que el hidrogeno se fije primero al Fe demuestra la existencia de que haya hidrogenasas que solo prefente Fe. El dihidrogen resultante,Ni-R1940, pierde un electrón. Como resultado el Ni(II) se oxida a Ni(III) y el ligando tiolato Cys350 ataca nucleoficamente al H2 coordinado. Se produce la rotura heterolitica del H-H La cisteina se protona y el ligando hidruro unido al Fe se convierte en un ligando puente. En el siguiente paso, la Cys 530 se vuelve a protonar, dando ugar a la especie intermedia NiC1952, lo que induce una transformación redox interna en el complejo. El ligandi hidruro reduce el Ni(III) a Ni (I) y el proton resultante se vuelve a fjar a la Cys530. Por último, la cesión de un segundo electrón a través de un cluster Fe-S y la perdida simultanes del protón de la Cys 530 conlleva a la regeneración de la especie inicial .")
26
CO deshidrogenasa(CODH)/Acetilcoenzima Sintasa(ACS).
Sensibles al O2 atm. Presente en microbios anaeróbicos CODH: cataliza procesos redox CO+ H2O CO2+ H++ 2 e- ACS: activan condensaciones no redox de metilo, carbonilo y tiol MeB12+ CO+ HS-CoA CH3(CO)SCoA+ H++ B12r CODH+ ACS= complejo Papel importante: Química medioambiental: microorganismos metabolizan CO bajan nivel CO (gas tóxico) Química biomédica: en tracto gastrointestinal bacterias hidrogenotróficas transforman H2 en otros gases La CODH y la ACS son enzimas sensibles al oxigeno atmosferico que se encuentran en microbios anaeróbicos. Se trata de dos enzimas diferentes que estan asociadas formando un complejo. En el complejo, los centros activos de ambos enzimas están conectados através de un tunel que el CO puede pasar del uno al otro. Las CODH y ACS tienen importancia en quimica medioambiental ya que sus funciones acopladas permiten que los microorganismos metabolicen CO ayudando así a mantener la concentracion de este gas toxico por debajo de niveles peligrosos. Las CODH y ACS desempeñan papeles biomedicos fundamentales en el tracto intestinal de algunos animales rumiantes, así como en el de los seres humanos. La biodegradación de materia orgánica en ambientes como el colon de los rumiantes y el colon de los humanos genera cantidades de hidrogeno que pueden inhibir su metabolismo. Las bacterias hidrogenotroficas usan el excedente de hidrogeno para sintetizar metano, acido acetico o sulfuro de hidrogeno.
SCoA+ H++ B12r. CODH+ ACS= complejo. Papel importante: Química medioambiental: microorganismos metabolizan CO bajan nivel CO (gas tóxico) Química biomédica: en tracto gastrointestinal bacterias hidrogenotróficas transforman H2 en otros gases. La CODH y la ACS son enzimas sensibles al oxigeno atmosferico que se encuentran en microbios anaeróbicos. Se trata de dos enzimas diferentes que estan asociadas formando un complejo. En el complejo, los centros activos de ambos enzimas están conectados através de un tunel que el CO puede pasar del uno al otro. Las CODH y ACS tienen importancia en quimica medioambiental ya que sus funciones acopladas permiten que los microorganismos metabolicen CO ayudando así a mantener la concentracion de este gas toxico por debajo de niveles peligrosos. Las CODH y ACS desempeñan papeles biomedicos fundamentales en el tracto intestinal de algunos animales rumiantes, así como en el de los seres humanos. La biodegradación de materia orgánica en ambientes como el colon de los rumiantes y el colon de los humanos genera cantidades de hidrogeno que pueden inhibir su metabolismo. Las bacterias hidrogenotroficas usan el excedente de hidrogeno para sintetizar metano, acido acetico o sulfuro de hidrogeno.")
27
Estructura CODH/ ACS. 2 unidades enlazadas convalentemente con 5 clústers metálicos: Clúster C: Ni- 4 Fe- 5 S Clúster B: 4 Fe- 4 S Clúster D: adicional entre subunidades, 4 Fe- 4 S Cluster C: centro activo para oxidacion CO. Metales se unen a la proteína por 5 Cys y 1 His Fe: coordinacion Td Ni (II): geometria de coordinacion cuadrada distorsionada, bajo espin, diamagnético. Centros activos de ambos enzimas conectados por un tunel para que pase el CO (oxidación) donde se consume H2O El enzima dimerico contiene cinco clusters metalicos, cada subunidad contiene un cluster mixto compuesto de Ni, fe y S (cluster C) y un cluster de tipo cubano 4Fe- 4S (cluster B). Un cluster adicional 4Fe- 4S (cluster D) situdado en la interfase entre las dos subunidades. Los cluster mixto Ni- Fe (cluster C) son los centros activos para la oxidacion del CO.
: geometria de coordinacion cuadrada distorsionada, bajo espin, diamagnético. Centros activos de ambos enzimas conectados por un tunel para que pase el CO (oxidación) donde se consume H2O. El enzima dimerico contiene cinco clusters metalicos, cada subunidad contiene un cluster mixto compuesto de Ni, fe y S (cluster C) y un cluster de tipo cubano 4Fe- 4S (cluster B). Un cluster adicional 4Fe- 4S (cluster D) situdado en la interfase entre las dos subunidades. Los cluster mixto Ni- Fe (cluster C) son los centros activos para la oxidacion del CO.")
28
Estructura CODH/ ACS.

29
Mecanismo CODH/ ACS. Oxidación CO: aporta 2 e- a centro activo formacion intermedio Ni(I) o desolcalizacion de e- d en los orbitales de ligandos y Fe Centros activos C y C’: actuan ndependientemente por su separacion, son el punto de partida de 2 cadenas de transporte que se unen en cluster D. Cluster D: facilita paso de los e- a aceptores externos La oxidacion de Co aporta dos electrones al centro activo lo que puede llevar a dos cosas: formacion de un intermedio de niquel (I) o la incorporacion de niquel en el cluster polinuclear con los ligandos S permitiria una importante deslocalizacion de los electrones d del metal en los orbitales de los ligandos y de los atomos de hierro adyacentes.
 o desolcalizacion de e- d en los orbitales de ligandos y Fe. Centros activos C y C’: actuan ndependientemente por su separacion, son el punto de partida de 2 cadenas de transporte que se unen en cluster D. Cluster D: facilita paso de los e- a aceptores externos. La oxidacion de Co aporta dos electrones al centro activo lo que puede llevar a dos cosas: formacion de un intermedio de niquel (I) o la incorporacion de niquel en el cluster polinuclear con los ligandos S permitiria una importante deslocalizacion de los electrones d del metal en los orbitales de los ligandos y de los atomos de hierro adyacentes.")
30
Mecanismo CODH/ ACS. Mientras que la distancia entre los clusters C y B o entre C’’ y B’’, y C y D o C’ y D’ son demasiado largas para permitir un transporte efectivo de electrones, la que se encuentra entre los clusters C y B’ o C’ y B es mas corta, permitiendo un transporte electronico efectivo. La orientacion del cluster hacia C hacia B’ y la del C’ hacia B tambien parecen apoyar esta ruta de transferenci electronica. El mecanismo implica la insercion del Co generado en la subunidad CODH entre el metilo aportado por una metilcobalamina y el tiol del coenzima A.

31
Metil-coenzima M reductasa.
Enzima que cataliza la reduccion de CO2 hasta CH4 En bacterias metanogénicas Sustratos: Metil coenzima M: 2- [metiltio] etanosulfonato (CH3-S-CoM) Coenzima B: 7- tioheptanoil- treonina fosfato (CoB-S-H) Las bacterias metanogenicas catalizan reacciones de reducción del CO2 hasta CH4 a través de unos procesos intermedios. Esta reaccion produce anualmente 109 toneladas de metano, el cual es un gas de efecto invernadero. Imagen: forma en que se unen los sustratos.
 Coenzima B: 7- tioheptanoil- treonina fosfato (CoB-S-H) Las bacterias metanogenicas catalizan reacciones de reducción del CO2 hasta CH4 a través de unos procesos intermedios. Esta reaccion produce anualmente 109 toneladas de metano, el cual es un gas de efecto invernadero. Imagen: forma en que se unen los sustratos.")
32
Estructura MCR. Inactiva: Activa: Ni (I)+ cofactor F430
Proteina formada por 3 subunidades: α, β, γ Ni(II): Geometria Oh Complejado por cofactor P430 (ecuatorial). Situado en el fondo del canal Unido a la proteina por O glutámico (axial) Unido a CoM- S- CH3 por S (axial). S interacciona con H2O. H2O: nexo de union de CoB CoB: situado en parte estrecha del canal. Bloquea el paso a muchas moleculas (H2O también) CoM: situado paralelamente a cofactor F430 Cavidad: proteina con subunidad α con Ni (II) con: cofactor F430+ CoM-S-CH3+ CoB-S-H o cofactor F430+ CoM-S-S-CoB Activa: Ni (I)+ cofactor F430
: Geometria Oh. Complejado por cofactor P430 (ecuatorial). Situado en el fondo del canal. Unido a la proteina por O glutámico (axial) Unido a CoM- S- CH3 por S (axial). S interacciona con H2O. H2O: nexo de union de CoB. CoB: situado en parte estrecha del canal. Bloquea el paso a muchas moleculas (H2O también) CoM: situado paralelamente a cofactor F430. Cavidad: proteina con subunidad α con Ni (II) con: cofactor F430+ CoM-S-CH3+ CoB-S-H o cofactor F430+ CoM-S-S-CoB. Activa: Ni (I)+ cofactor F430.")
33
Estructura MCR. Imagen 1: cofactor 430 Imagen 2: estructura

34
Mecanismo MCR Centro activo: accesible para moléculas con un tamaño < 6 Å Molécula H2o: desplazada por CoM-S-CH3 CoM-S-CH3: se une a CoB y da CH4 (en zona hidrofóbica) Pasos: Ataque nucleófilo de Ni (I) al -CH3 del CH3-S-CoM formación enlace organometálico Ni (III)- CH3 Ni (III) (oxidante fuerte) arranca 1 e- al –SH formación radical (ácido) disociación El –CH3 se protona formación enlace –S-S- generación 1 e- reducción Ni (II) a Ni (I) Eliminación de CH4 y disulfuro entrada de nuevos sustratos reiniciación ciclo catalítico
 Pasos: Ataque nucleófilo de Ni (I) al -CH3 del CH3-S-CoM formación enlace organometálico Ni (III)- CH3. Ni (III) (oxidante fuerte) arranca 1 e- al –SH formación radical (ácido) disociación. El –CH3 se protona formación enlace –S-S- generación 1 e- reducción Ni (II) a Ni (I) Eliminación de CH4 y disulfuro entrada de nuevos sustratos reiniciación ciclo catalítico.")
35
Mecanismo MCR

36
Mecanismo MCR

37
BIBLIOGRAFIA Enrique J. Baran. Química bioinorgánica.McGraw-Hill (1995) VVAA. Itroducción a la química bioinorgánica.Editorial sintesis (2003) Recursos electroicos: PROTEIN DATA BANK
 VVAA. Itroducción a la química bioinorgánica.Editorial sintesis (2003) Recursos electroicos: PROTEIN DATA BANK")
38
BIBLIOGRAFIA Libros Páginas:
Conceptos y modelos de química inorgánica / Bodie E. Douglas, Darl H. McDaniel, John J. Alexander ; versión española de José Gálvez Flores, Gregorio López López y Vicente Iranzo Rubio. Barcelona : Reverte, a. ed. Química inorgánica básica / F. Albert Cotton, Geoffrey Wilkinson. México : Limusa, 1980. Páginas:

39
RESUMEN Tunicados: en vacuolas, V(III)
VANADIO Abundancia: No existe concentrada en minerales, si todos lo suelos En agua: muy bajas concentraciones Ión más común: VO43- EO= 5 reducible a 4 y 3. Interfiere en bioquímica de PO43- Tunicados: en vacuolas, V(III) En H2O: V (V) se reduce a V (III) difícilmente reductores energéticos (tunicromo, complejo v3++ tunicromo [V(H2O)6]: dador electrónico terminal de cadena de transporte electrónico
 En H2O: V (V) se reduce a V (III) difícilmente reductores energéticos (tunicromo, complejo v3++ tunicromo. [V(H2O)6]: dador electrónico terminal de cadena de transporte electrónico.")
40
RESUMEN Amavadina: V (IV)[VL2]2-. IC= 8.
Hongo extrae V con facilidad del suelo selectivamente Oxidación de alcoholes primarios a aldehídos, usando O2 reduciendo a H2O2 H aloperoxidasas de vanadio: V (V), IC= 5 Bipiramide triangular, VO3-. Carga neutralizada con enlaces de hidrógeno entre oxígenos y restos de Aá Algas marinas y hongos Oxidación de iones haluro con agua para dar ácido hipoaloso: H2O2 + X- + H+ H2O + HOX
![RESUMEN Amavadina: V (IV)[VL2]2-. IC= 8.](http://slideplayer.es/slide/3280171/11/images/40/RESUMEN+Amavadina%3A+V+%28IV%29%EF%83%A0%5BVL2%5D2-.+IC%3D+8..jpg "Hongo extrae V con facilidad del suelo selectivamente. Oxidación de alcoholes primarios a aldehídos, usando O2 reduciendo a H2O2. H aloperoxidasas de vanadio: V (V), IC= 5 Bipiramide triangular, VO3-. Carga neutralizada con enlaces de hidrógeno entre oxígenos y restos de Aá. Algas marinas y hongos. Oxidación de iones haluro con agua para dar ácido hipoaloso: H2O2 + X- + H+ H2O + HOX.")
41
RESUMEN Diabetes: deficiencia de insulina o resistencia celular a su función conduce a diabetes. Se ha demostrado que el vanadio representa un elemento asociado a la regulación del metabolismo de la glucosa y mejora la sensibilidad del receptor a la insulina.

42
RESUMEN CROMO Cr (III) Elemento abundante y esencial
Bioquímica Amplificación de la señal de la insulina Consumo (0,01 y 1,2 mg Cr(III) diarios) levadura de cerveza. Difícil encontrar personas con déficit de Cr. Complejo metálico con actividad biológica CROMODULINA Estructura desconocida 4 centros Cr(III) Efecto cooperativo
 diarios) levadura de cerveza. Difícil encontrar personas con déficit de Cr. Complejo metálico con actividad biológica. CROMODULINA. Estructura desconocida. 4 centros Cr(III) Efecto cooperativo.")
43
RESUMEN Modo de acción Células capaces de acumular apo-cromodulina
↑[glucosa] Insulina Insulina se une a un receptor especifico Cambio estructural incorporación de Cr Holo-cromodulina se une al receptor Mantiene forma activa amplificando su actividad Desaparece la señal se relaja la conformación del receptor Cromodulina expulsada de la célula, excretada a traves de la orina.

45
RESUMEN NIQUEL Elemento traza esencial sistemas bacterianos
EO (II) complejos Oh Bioquímica 5 tipos de enzimas Catalizan, distintas reacciones.
 complejos Oh. Bioquímica 5 tipos de enzimas. Catalizan, distintas reacciones.")
46
RESUMEN Reacción catalizada Enzima Centro Ni Reacción UREASAS
Dímero de Ni(II) Urea NH3 + CO2 Reacción catalizada HIDROGENASAS 2H+ + 2e- ↔H2 IMP Clusters [4Fe-4S] y [3Fe-4S]
 Urea NH3 + CO2. Reacción catalizada. HIDROGENASAS. 2H+ + 2e- ↔H2. IMP. Clusters. [4Fe-4S] y [3Fe-4S]")
47
RESUMEN Mecanismo ureasas

48
RESUMEN Mecanismo hidrogenasas

49
MeB12+ CO+ HS-CoA CH3(CO)SCoA+ H++ B12r
Metil-CoM reductasa F-430 CH3S-CoM CH4 Acettil CoA sintetasa/CO deshidrogenasa Ni- 4 Fe- 5 S CO deshidrogenasas: CO+ H2O CO2+ H++ 2 e- Acetil CoA sintasa: MeB12+ CO+ HS-CoA CH3(CO)SCoA+ H++ B12r
SCoA+ H++ B12r.")
50
RESUMEN Mecanismo CO deshidrogenasa(CODH)/Acetilcoenzima Sintasa(ACS).
/Acetilcoenzima Sintasa(ACS).")
51
RESUMEN Mecanismo Metil-coenzima M reductasa.

Presentaciones similares