Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Mutación Universidad Nacional del Santa Facultad de Ciencias
Departamento de Biología, Microbiología y Biotecnología Mutación Blga. Pesq. Eliana Zelada Mázmela Blgo. Acuic. Carmen Yzásiga Barrera

2
Recombinación independiente
Evolución variantes genéticas mutación entrecruzamiento Recombinación independiente Una de las fuentes usuales de biodiversidad son las “mutaciones”. Estas consisten en cambios en la secuencia de nucléotidos del ADN. Si la mutación se produce en un sector no codificante o regulador del genoma, esta no tendrá trascendencia Si afecta en cambio un codón de una secuencia codificante, podrá traducirse en el cambio de un AA de una CHON, lo que puede significar un cambio estructural y/o funcional de la misma, el que puede afectar en forma negativa o positiva, su funcionalidad y competencia.

3
Consecuencias de los cambios genéticos
Agentes físicos Cambios en el material genético del núcleo celular Agentes químicos Instancias de la vida 1) Mutaciones 2) Alteraciones cromosómicas 3) Recombinación meiótica Reordenamientos que se producen regularmente durante las divisiones meióticas en la formación de los gametos. Si los cambios se producen en las células somáticas, se perderán con la muerte de la célula portadora o del individuo.
 Mutaciones. 2) Alteraciones cromosómicas. 3) Recombinación meiótica Reordenamientos que se producen regularmente durante las divisiones meióticas en la formación de los gametos. Si los cambios se producen en las células somáticas, se perderán con la muerte de la célula portadora o del individuo.")
5
Sólo si la alteración se hubiera producido en una célula que participara en la formación de un gameto, tendrá posibilidades de que el cambio se transmita a un descendiente, y por tanto pueda incidir primero en la población de esa especie y eventualmente en su futuro. Cambio genómico No tienen importancia para el individuo Supondrá desventajas para el individuo Supondrá ventajas para el individuo Es decir son deletéreas, aparecen con menor frecuencia debido a la acción de la SN

6
La mayoría de los genes son relativamente estables, las mutaciones son eventos relativamente extraños Tasa de mutación: 1 x 10-5 a 1 x 10-6 , es decir de cada 1/ a 1/ gameto puede contener una mutación en un locus determinado En un organismo superior con genes, se espera que 1/10 gametos a 1/100 gametos contenga al menos una mutación La tasa de mutación de un gene es característica del gene como lo es su expresión fenotípica Depende del genotipo residual, es decir existen algunos genes que se dedican a incrementar la frecuencia de mutación de otros genes: genes mutadores

7
Ejm: ojo W 60% viabilidad W+ W x W Y 50 s : 50 % b
La mutación ocasiona desviaciones de las leyes de Mendel, ya que estas suponen igualdad de supervivencia y/o capacidad reproductiva de los diferentes genotipos Ejm: ojo W % viabilidad W+ W x W Y 50 s : 50 % b 50 x 0,6 = 30 b : 50 s

8
Mutante: Individuo que expresa el cambio fenotípico causado
Mutación Proceso por el cual un gene sufre un cambio estructural Gene modificado producido por el cambio mutacional Cambio fenotípico causado por un gene modificado Puede ser de adelante hacia atrás o viceversa (reververancia) Mutante: Individuo que expresa el cambio fenotípico causado Cuando ocurre la mutación, la nueva forma del gene “mutante” se hereda de manera estable, de igual manera que la forma previa o “silvestre”, que es el término que se usa para describir el fenotipo y genotipo. La mutación varía desde ser indetectable, es decir las ausencia del péptido no parece ocasionar ningún problema, hasta ser letal, es decir el organismo no puede vivir sin el polipéptido
 Mutante: Individuo que expresa el cambio fenotípico causado. Cuando ocurre la mutación, la nueva forma del gene mutante se hereda de manera estable, de igual manera que la forma previa o silvestre , que es el término que se usa para describir el fenotipo y genotipo. La mutación varía desde ser indetectable, es decir las ausencia del péptido no parece ocasionar ningún problema, hasta ser letal, es decir el organismo no puede vivir sin el polipéptido.")
9
Historia El primero en utilizar el término mutación fue Hugo de Vries en 1901, pero él lo aplicó a cambios bruscos en los caracteres de una especie, al observar cómo inesperadamente entre la descendencia de una planta llamada Oenothera lamarckiana había individuos gigantes. Las investigaciones de Thomas Hunt Morgan en Drosophila mostraron que existen numerosas mutaciones que pueden provocar cambios tan pequeños que son difícilmente apreciables. Desde entonces, el concepto de mutación no se restringe a los cambios bruscos, sino a cualquier cambio heredable del fenotipo. En concreto, las variaciones alélicas de los genes (color de los guisantes, forma de las crestas de las gallinas, color del pelo del ratón, etc.), son mutaciones del gen más primitivo en la especie, que suele llamarse gene silvestre
, son mutaciones del gen más primitivo en la especie, que suele llamarse gene silvestre.")
10
Definiciones iniciales
Desde punto de vista fenotípico, mutación es la aparición brusca y espontánea de una variación fenotípica de un individuo, la cual se transmite hereditariamente a la progenie. Pero tras el descubrimiento del ADN como material de la herencia, podemos plasmar una definición más ajustada, desde el punto de vista genotípico: Es cualquier alteración de la secuencia de bases de un segmento de ADN correspondiente a un gen o a un locus (sea éste transcribible o no), aún cuando esta alteración no se refleje en forma de cambio fenotípico observable o detectable.
, aún cuando esta alteración no se refleje en forma de cambio fenotípico observable o detectable.")
11
Clasificación de las mutaciones
Según el mecanismo que ha provocado el cambio en el material genético, se suele hablar de tres tipos de mutaciones: Mutaciones cariotípicas o genómicas. Mutaciones cromosómicas Mutaciones génicas o moleculares Hay una tendencia actual a considerar como mutaciones en sentido estricto, solamente las génicas, mientras que los otros tipos entrarían en el término de aberraciones cromosómicas.

12
Mutaciones génicas o moleculares
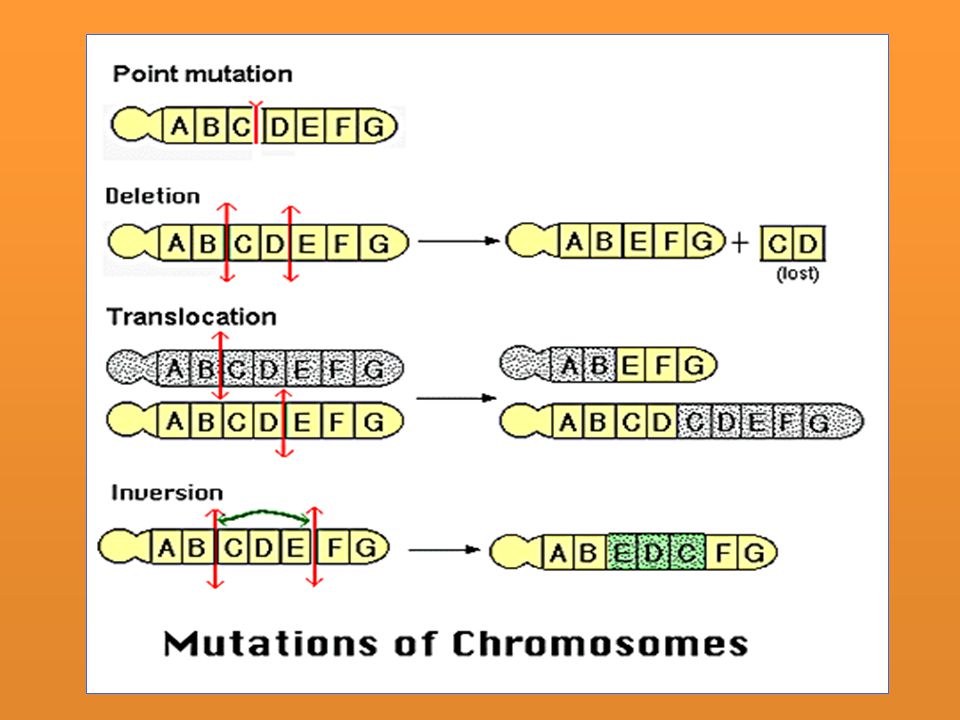
Son la mutaciones que ocurren al alterar la secuencia de nucleótidos del ADN. Entre las mutaciones puntuales podemos distinguir: Mutación por sustitución de bases: Se producen al cambiar la posición de un nucleótido por otro, por ejemplo, donde debería haber un nucleótido de citosina, se inserta uno de timina. Mutación por pérdida de nucleótidos o delección: En la secuencia de nucleótidos se pierde uno y no se sustituye por nada. Mutación por inserción de nuevos nucleótidos: Dentro de la secuencia del ADN se introducen nucleótidos que no deberían estar. Mutación por inversión de nucleótidos: Cuando ocurre un cambio en la orientación de un segmento de DNA Mutación por traslocación de pares de nucleótidos complementarios:

15
Mutaciones cromosómicas
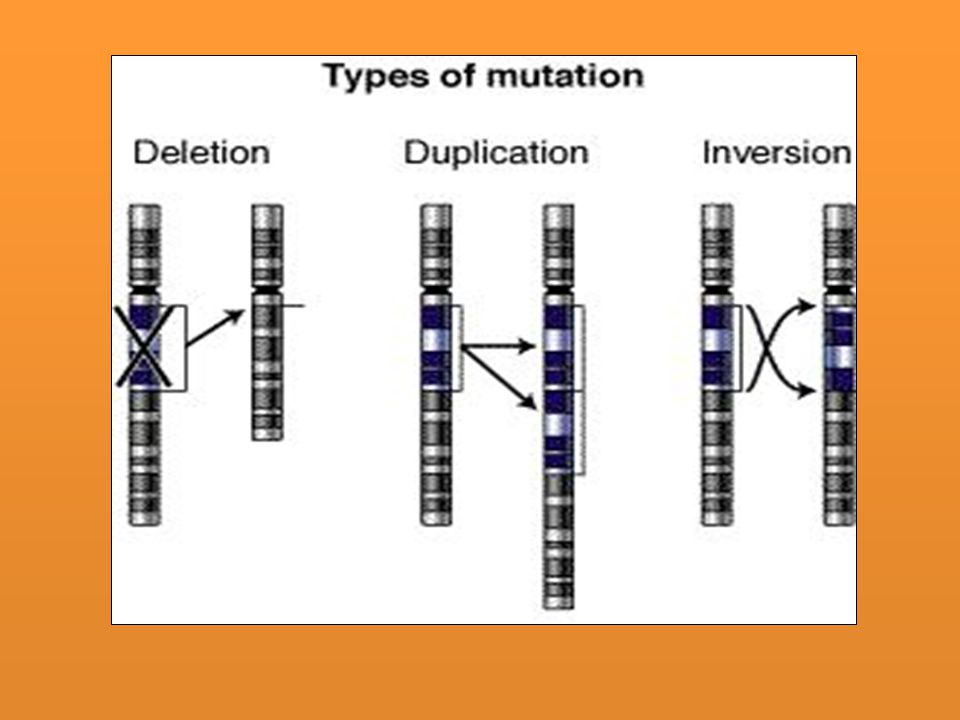
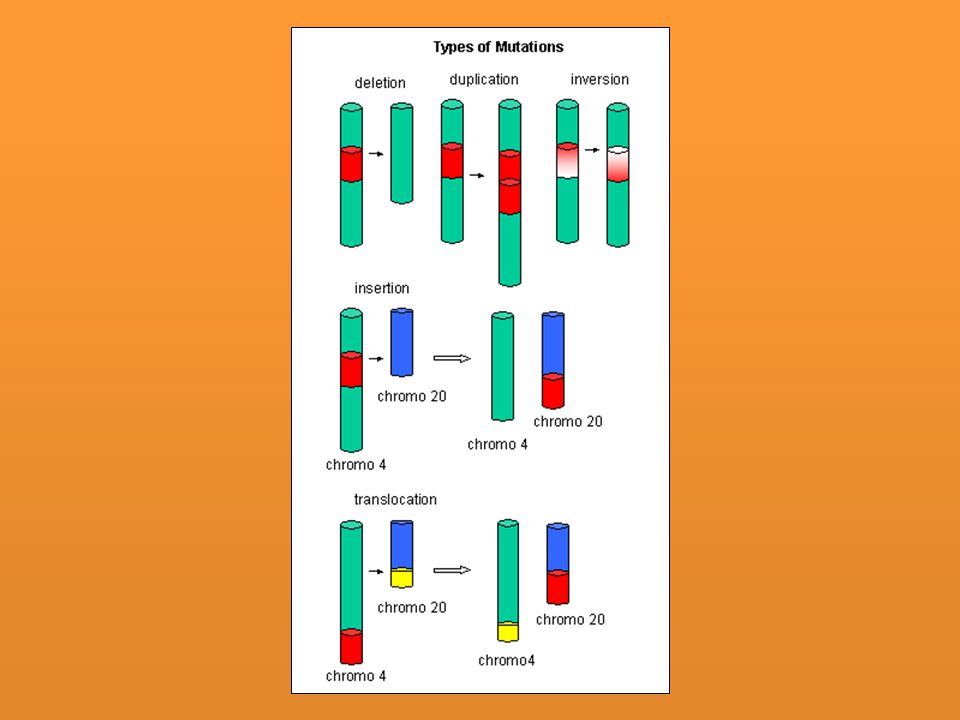
Afectan la secuencia de los hipotéticos fragmentos en que podría subdividirse transversalmente un cromosoma. Muchas de ellas son apreciables al microscopio gracias a la “técnica de bandas” con la que se confecciona el cariotipo: Mutación por inversión de un fragmento cromosómico. Mutación por deleción o pérdida de un fragmento cromosómico. Mutación por duplicación de un fragmento cromosómico. Suelen estar asociadas casi siempre con delecciones en otro cromosoma. Mutación por translocación de un fragmento cromosómico, es decir por un cambio en la posición de un fragmento cromosómico. La translocación puede ocurrir en un solo cromosoma, entre cromosomas homólogos o entre cromosomas diferentes

17
Mutaciones cariotípicas o genómicas
Son las mutaciones que afectan al número de cromosomas o todo el genoma. Poliploidía: Es la mutación que consiste en el aumento del número normal de “juegos de cromosomas” . Los seres poliploides pueden ser autopoliploides, si todos los juegos proceden de la misma especie, o alopoliploides, si proceden de la hibridación, es decir, del cruce de dos especies diferentes. Haploidía: Son las mutaciones que provocan una disminución en el número de juegos de cromosomas.

18
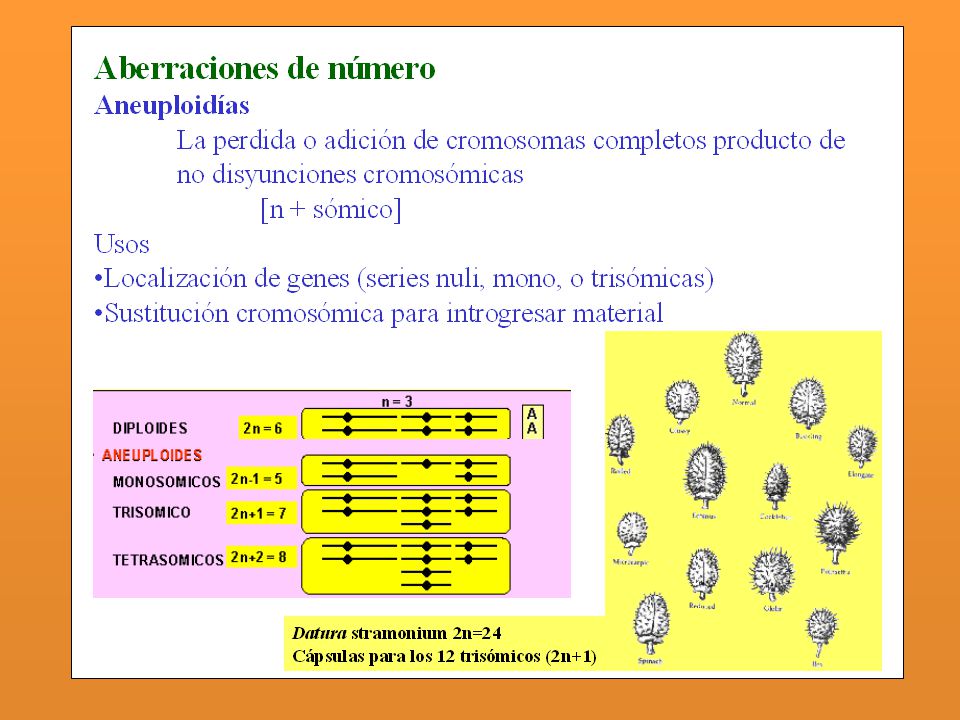
Aneuploidía: Son las mutaciones que afectan sólo a un número de ejemplares de un cromosoma o más, pero sin llegar a afectar al juego completo. Las aneuploidías pueden ser monosomías, trisomías, tetrasomías, etc, cuando en lugar de dos ejemplares de cada tipo de cromosomas, que es lo normal, hay o sólo uno, o tres, o cuatro, etc. Entre las aneuplodías podemos encontrar diferentes tipos de trastornos genéticos en humanos como pueden ser: Trisomía 21 o Síndrome de Down o mongolismo que tienen 47 cromosomas. Trisomía 18 o Síndrome de Edward. También tienen 47 cromosomas Monosomía X0 o Síndrome de Turner Trisomía sexual XXX Trisomía sexual XXY o Klinefelter Trisomía sexual XYY

20
Síndrome de Klinefelter

23
Trisomía del 21 (47, XY, +21)
")
24
Síndrome de Patau (47, XX, +13)
newborn male with full trisomy 13 (Patau syndrome). this baby has a cleft palate, atrial septal defect, inguinal hernia, and postaxial polydactyly of the left hand. Síndrome de Patau (47, XX, +13)
. this baby has a cleft palate, atrial septal defect, inguinal hernia, and postaxial polydactyly of the left hand. Síndrome de Patau (47, XX, +13)")
25
Trisomía del 18 o Síndrome de Edwars (47, XY, +18)
")
26
Síndrome del duplo Y (47, XYY, +23)
")
27
Síndrome de Turner

29
Clasificación de las mutaciones
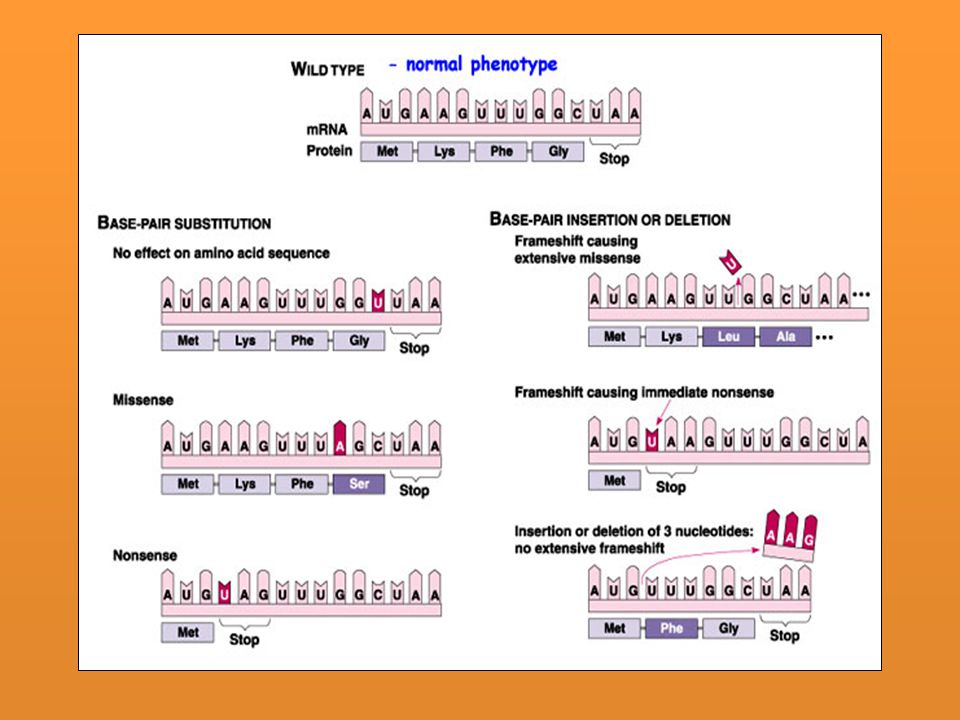
I. TAMAÑO A) M. Puntual: Un cambio en un segmento muy pequeño del ADN: 1 ó 2 nuc Con igual sentido: Cambio en un codón (3a posición) que no altera el AA especificado. Se considera mutación silenciosa cualquier cambio en la secuencia de nucleótidos del ADN que no produce cambio en el fenotipo estudiado: El fenotipo analizado es simplemente la función de una enzima, es evidente que existen mutaciones en la secuencia de nucleótidos del ADN que no producen cambios en la secuencia de AA, pero también hay mutaciones en el ADN que producen cambios en la secuencia de aminoácidos que no alteran la función de la enzima.
 M. Puntual: Un cambio en un segmento muy pequeño del ADN: 1 ó 2 nuc. Con igual sentido: Cambio en un codón (3a posición) que no altera el AA especificado. Se considera mutación silenciosa cualquier cambio en la secuencia de nucleótidos del ADN que no produce cambio en el fenotipo estudiado: El fenotipo analizado es simplemente la función de una enzima, es evidente que existen mutaciones en la secuencia de nucleótidos del ADN que no producen cambios en la secuencia de AA, pero también hay mutaciones en el ADN que producen cambios en la secuencia de aminoácidos que no alteran la función de la enzima.")
30
Purina por otra o pirimidina por otra
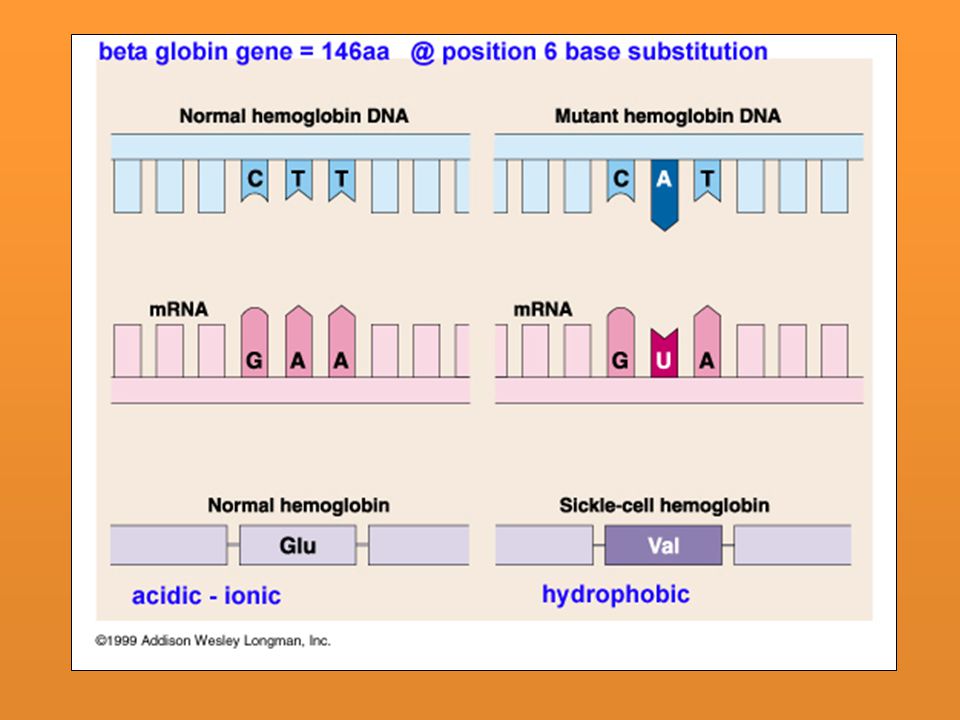
b) Sin sentido: Término de producto proteico por un signo de terminación. c) Con sentido equivocado: Cambio en la secuencia de AA por el que el AA equivocado ocupa una posición dada en la cadena peptídica.: Hb d) Por corrimiento en el marco de lectura: Crea codones equivocados o sin sentido a los largo del cistrón. Transición Purina por otra o pirimidina por otra Transversión Purina por pirimidina y viceversa
 Sin sentido: Término de producto proteico por un signo de terminación. c) Con sentido equivocado: Cambio en la secuencia de AA por el que el AA equivocado ocupa una posición dada en la cadena peptídica.: Hb. d) Por corrimiento en el marco de lectura: Crea codones equivocados o sin sentido a los largo del cistrón. Transición. Purina por otra o pirimidina por otra. Transversión. Purina por pirimidina y viceversa.")
38
Corrimiento en el marco de lectura

39
Tipos de mutaciones génicas
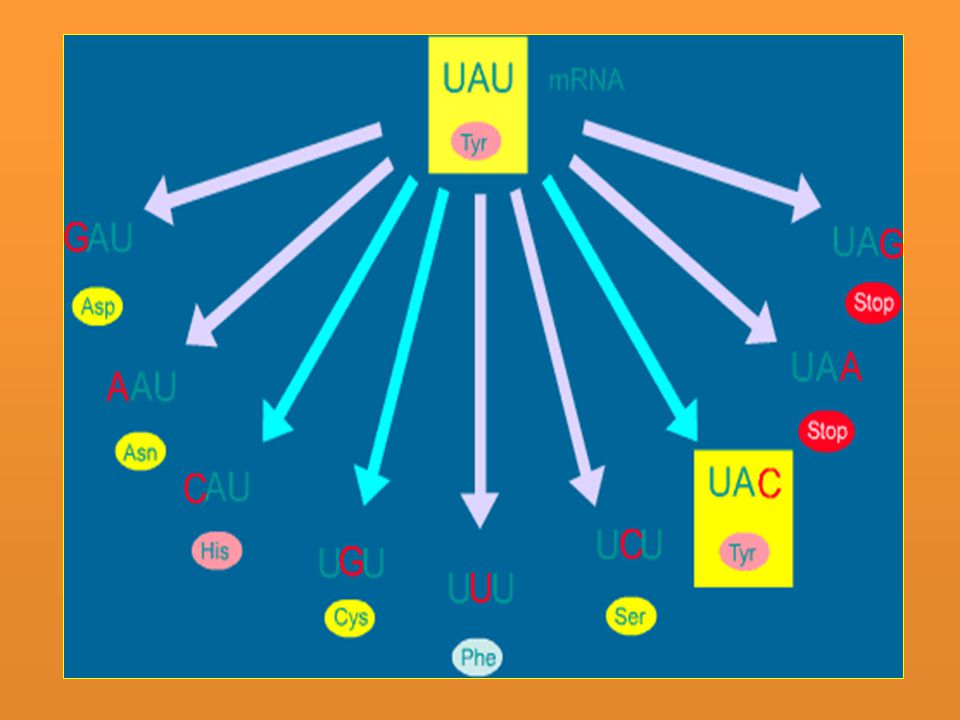
Resultados y ejemplos En el ADN Transiciones Pu→Pu o Pi→Pi: AT→GC, GC→AT, CG→TA y TA→CG Transversiones Pu→Pi o Pi→Pu: AT→CG, AT→TA, GC→TA, GC→CG, TA→GC, TA→AT, CG→AT y CG→GC En la proteína Mutación silenciosa Tripletes que codifican para el mismo aminoácido: AAG(arg)→CGG(arg) Mutación neutra Tripletes que codifican para aminoácidos equivalentes distintos. AAA(lys)→AGA(arg). Ambos son aminoácidos básicos Mutación cambio de sentido Aparece un nuevo triplete que codifica para un aminoácido de distinto tipo. La proteína pierde su función. Mutación sin sentido Aparece un triplete de terminación o FIN: CAG(gln)→UAG(FIN) Mutación cambio de fase o pauta de lectura Adición o deleción de un único par de nucleótidos o de varios pares de nucleótidos, siempre que no sean múltiplo de tres. Es importante aclarar el concepto de mutación silenciosa, una mutación silenciosa
→CGG(arg) Mutación neutra. Tripletes que codifican para aminoácidos equivalentes distintos. AAA(lys)→AGA(arg). Ambos son aminoácidos básicos. Mutación cambio de sentido. Aparece un nuevo triplete que codifica para un aminoácido de distinto tipo. La proteína pierde su función. Mutación sin sentido. Aparece un triplete de terminación o FIN: CAG(gln)→UAG(FIN) Mutación cambio de fase o pauta de lectura. Adición o deleción de un único par de nucleótidos o de varios pares de nucleótidos, siempre que no sean múltiplo de tres. Es importante aclarar el concepto de mutación silenciosa, una mutación silenciosa.")
40
B) Mutaciones Gruesas: Involucran más de un par de nucleótidos, genes completos, cromosomas completos, juegos de cromosomas Alteraciones cromosómicas numéricas: Fusión céntrica: Compromete a cromosomas con centrómeros terminales. Fisión céntrica: Compromete a cromosomas metacéntricos, submetacéntricas e inclusive acrocéntricos 3 2 2/ 3 1 1a b

41
- Euplodía: Variación en el número de juegos cromosómicos
- Euplodía: Variación en el número de juegos cromosómicos. Frecuente en vegetales donde se estima que 1/3 de las angiospermas lo presentan. Se aplica a organismos que presentan múltiplos del número básico n -) Monoploide: En el núcleo sólo se encuentra un juego cromosómico y en organismos superiores donde generalmente el n es más pequeño y menos vigoroso que el 2n. Pocos viven y generalmente son estériles.
 Monoploide: En el núcleo sólo se encuentra un juego cromosómico y en. organismos superiores donde generalmente el n es más pequeño y menos vigoroso que el 2n. Pocos viven y generalmente son estériles.")
42
-) Triploide: Se originan tres juegos por la fusión de un n con un 2n
-) Triploide: Se originan tres juegos por la fusión de un n con un 2n. El juego extra se distribuye en varias combinaciones en las células germinales resultando gametos desbalanceados genéticamente: Estériles y no se encuentran comúnmente en poblaciones naturales. Ofrece la oportunidad para estudiar efecto de dosis: como dos o más alelos de un locus se comportan en presencia de una sóla dosis del alelo alternativo: Si la sóla dosis enmascara a la doble: xenia sobre el 2º Endospermo floculento S SSS, SSs, Sss; sss Endospermo azucarado s f a
 Triploide: Se originan tres juegos por la fusión de un n con un 2n. El juego extra se distribuye en varias combinaciones en las células germinales resultando gametos desbalanceados genéticamente: Estériles y no se encuentran comúnmente en poblaciones naturales. Ofrece la oportunidad para estudiar efecto de dosis: como dos o más alelos de un locus se comportan en presencia de una sóla dosis del alelo alternativo: Si la sóla dosis enmascara a la doble: xenia sobre el 2º. Endospermo floculento S. SSS, SSs, Sss; sss. Endospermo azucarado s. f a.")
43
Dos sps vegetales 2n diferentes pueden hibridarse
-) Tetraploide: Con cuatro juegos de cromosomas (4n) por duplicación somática del Nº de cromosomas. Espontánea o inducida. . Autotetraploide: La ploidía incluye sólo complementos de cromosomas homólogos. Pueden dar gametos viables si la disyunción produce gametos genéticamente balanceados, es decir de los cuatro juegos dos se van a un polo y los otros dos al otro. Piede ser por suplicación somática o unión de gametos no reducidos. . Alotetraploide: Dos sps vegetales 2n diferentes pueden hibridarse Gametos no reducidos de especies diferentes F1 2n y estéril Una nueva sp Resulta del fracaso de c/complemento cromosómico para logrtar suficiente homología genética y permitir el apareamiento
 Tetraploide: Con cuatro juegos de cromosomas (4n) por duplicación somática del Nº de cromosomas. Espontánea o inducida. . Autotetraploide: La ploidía incluye sólo complementos de cromosomas homólogos. Pueden dar gametos viables si la disyunción produce gametos genéticamente balanceados, es decir de los cuatro juegos dos se van a un polo y los otros dos al otro. Piede ser por suplicación somática o unión de gametos no reducidos. . Alotetraploide: Dos sps vegetales 2n diferentes pueden hibridarse. Gametos no reducidos de especies diferentes. F1 2n y estéril. Una nueva sp. Resulta del fracaso de c/complemento cromosómico para logrtar suficiente homología genética y permitir el apareamiento.")
44
Duplicación de cromosomas
Sea AA el complemento diploide de cromosomas de una especie y BB de la otra especie AA x BB Híbrido estéril AB Duplicación de cromosomas AA BB Fértil Anfidiploide -) Poliploidía: Cualquier célula con más de dos juegos Trigo de pan: 6n (3n es haploide). En humanos se da en hepatocitos
 Poliploidía: Cualquier célula con más de dos juegos. Trigo de pan: 6n (3n es haploide). En humanos se da en hepatocitos.")
48
Tetrasomía: 2n + 2 (cuatro del mismo)
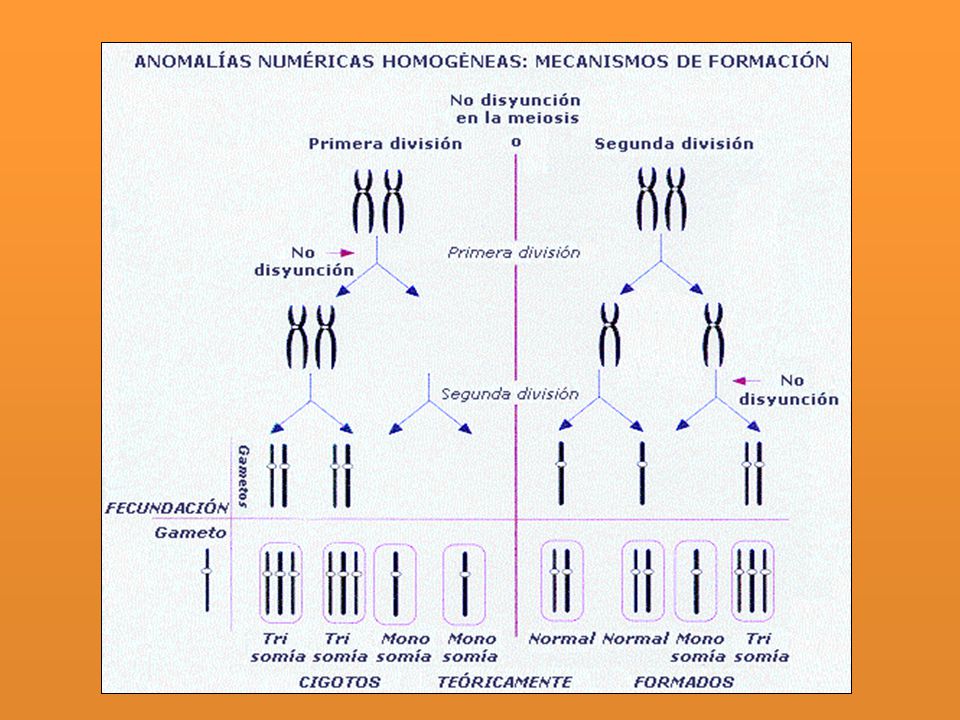
- Aneuploidía: Cuando se presentan variaciones en el Nº cromosómico pero que no incluye el juego completo. Se usa el sufijo sómico Monosomía: 2n - 1 Trisomía: 2n + 1 Tetrasomía: 2n + 2 (cuatro del mismo) Trisomía doble: 2n Nulisomía: 2n - 2 ( Ha perdido todo un par)
 Trisomía doble: 2n Nulisomía: 2n - 2 ( Ha perdido todo un par)")
50
2. Alteraciones cromosómicas estructurales
Duplicación: Un cromsoma gana parte de su homólogo. Su presencia no es tan dañina como la deficiencia. Delección: Pérdida de una región. Nunca retromutan. La extensión genética de la delección se define por su incapacidad de combinar con su homólogo. Cuando se pierde una pequeña parte del alelo dominante el recesivo se puede expresar en heterocigosis. Cuando es un segmento grande el que se pierde generalmente es letal Translocación: El segmento se va a otro que no es su homólogo. Inversión: La región se invierte en el mismo cromosoma - Por sustitución de bases:

51
Robertsonian translocations are a special case of 'almost balanced' translocations. any two out of chromosomes 13, 14, 15, 21 and 22. These chromosomes are all acrocentric, that is, the centromere is very close to one end. The short arms contain few, if any, genes except for many tandemly repeated copies of the ribosomal RNA genes. A Robertsonian translocation is a fusion between the centromeres of two of these chromosomes with loss of the short arms forming a chromosome with two long arms, one derived from each chromosome. The loss of the short arms does not matter, each cell still has eight copies of the rRNA gene block and that, apparently, is enough. In the same Every diploid cell thus contains 10 copies of the block of repeated genes. way as balanced reciprocal translocation carriers have difficulties at meiosis in ensuring the correct segregation of the chromosomes to make a balanced set, so too do Robertsonian translocation carriers .21 possess a special problem of their own, one of the possible unbalanced gametes will contain effectively two copies of chromosome 21 (when the translocation chromosome and chromosome 21 segregate to the same pole). They are thus at risk of producing a baby with Down syndrome.
. They are thus at risk of producing a baby with Down syndrome.")
54
Crudi chat delecion de una pequeña fraccion del 5 (46,XY, +p)
")
55
Translocación

63
B) Control Genético: Influenciada por genes mutadores
II. POR SU ORIGEN Espontáneas: De origen desconocido, “mutación basal” Puede ser provocado por el metabolismo, edad, predisposición genética. Aparecen con una frecuencia relativamente baja. B) Control Genético: Influenciada por genes mutadores - Específicas: Efecto limitado a un locus - No específicas: Efecto limitado a varios loci C) Inducidas o artificiales: Por exposición a ambientes anormales Radiaciones ionizantes: Producen cambios en la valencia química por la expulsión de electrones. Producido por protones, neutrones, rayos X, alfa, beta gamma.. La mutabilidad está en proporción directa a la cantidad.
 Control Genético: Influenciada por genes mutadores. - Específicas: Efecto limitado a un locus. - No específicas: Efecto limitado a varios loci. C) Inducidas o artificiales: Por exposición a ambientes anormales. Radiaciones ionizantes: Producen cambios en la valencia química por la expulsión de electrones. Producido por protones, neutrones, rayos X, alfa, beta gamma.. La mutabilidad está en proporción directa a la cantidad.")
64
- Radiaciones no ionizantes: Elevan los niveles de energía átomos: UV, calor. Producen los dímeros de timina ( enlaces T-T en la misma hebra) - Mutagénicos químicos: Sustancias químicas que incrementan la mutabilidad de los genes.

65
Dímeros de timina

66
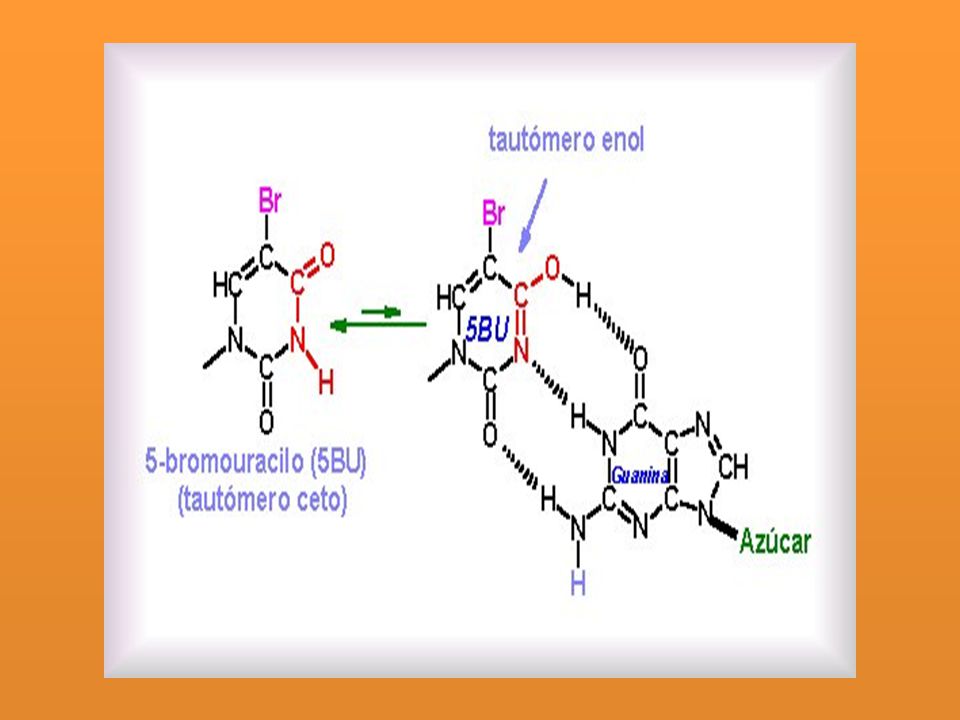
Errores de copia: Surgen después de la replicación del ADN: Análogos de bases: Bases similares a las normales. Son compuestos químicos que pueden remplazar a una base determinada. 5BU análogo T 5BU A BU empareja con G

68
Cuando el 5BU se incorpora en el ADN en lugar de una timina, lo que usualmente sucede es que induce una transición A·T G·C.

69
Luego induce una transición de G - C A - T

70
El análogo de adenina 2-aminopurina (2AP), normalmente se une con la timina
Pero a veces forma un par de bases desordenado con la citosina

71
b) Cambio directo de los genes: En ADN no replicativo:
- HNO2 causa desaminación directa: A hipoxantina (Hipoxantina es análoga de G) Causa entonces una transición A-T G-C
 Causa entonces una transición A-T G-C.")
72
Convierte también la C en U

73
La hidroxilamina (NH2OH) también induce la transición G·C A·T
La hidroxilamina (NH2OH) también induce la transición G·C A·T. Específicamente reacciona con citosina convirtiéndola en un compuesto que hace par de bases con la adenina.
 también induce la transición G·C A·T. Específicamente reacciona con citosina convirtiéndola en un compuesto que hace par de bases con la adenina.")
74
- Agentes alquilantes: como el EMS que añade radicales etilo y produce transiciones GC→AT.

75
- Aflatoxina B1: se une a la G modificándola de manera que la G modificada se separa del azúcar al que estaba unida produciendo una sede apurínica. El sistema SOS pone habitualmente en la sede apurínica una Adenina (A). Es un potente carcinógeno.
. Es un potente carcinógeno..")
76
Agentes de tipo intercalante como la Proflavina, Naranja de acridina y Compuestos ICR: Son compuestos con estructuras planas que se meten o intercalan entre las bases nitrogenadas del ADN produciendo adiciones o deleciones de un solo par de nucleótidos.

78
ERRORES EN LA REPLICACIÓN
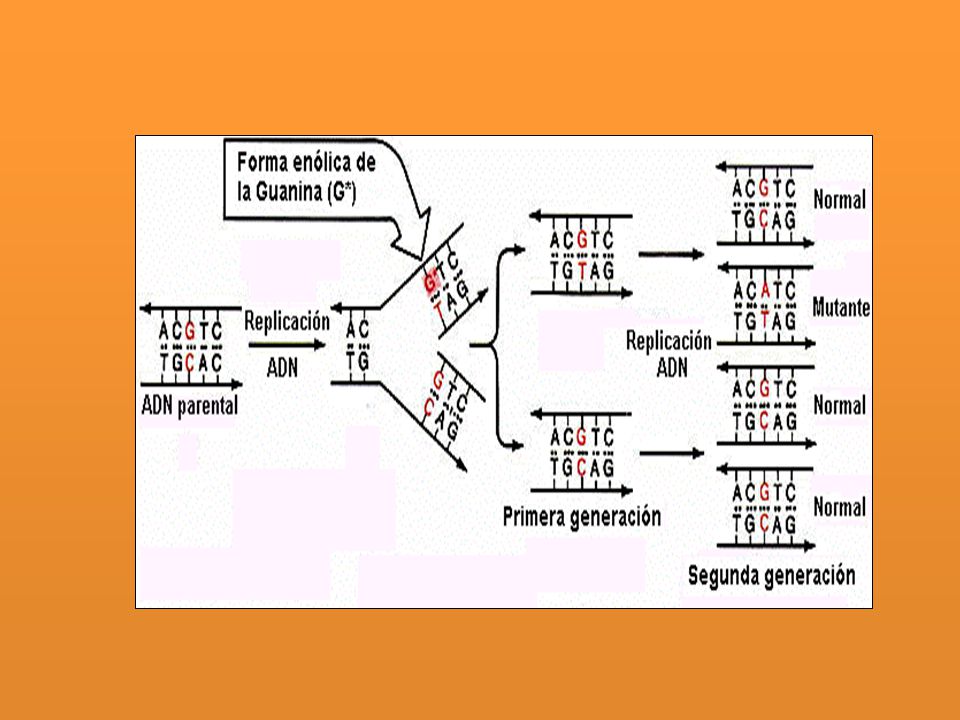
La tautomería: las bases nitrogenadas se encuentran habitualmente en su forma cetónica y con menos frecuencia aparecen en su forma tautomérica enólica o imino. Las formas tautoméricas o enólicas de las bases nitrogenadas (A*, T*, G* y C*) muestran relaciones de apareamiento distintas: A*-C, T*-G, G*-T y C*-A. El cambio de la forma normal cetónica a la forma enólica produce transiciones. Los errores en el apareamiento incorrecto de las bases nitrogenadas pueden ser detectados por la función correctora de pruebas de la ADN polimerasa III.
 muestran relaciones de apareamiento distintas: A*-C, T*-G, G*-T y C*-A. El cambio de la forma normal cetónica a la forma enólica produce transiciones. Los errores en el apareamiento incorrecto de las bases nitrogenadas pueden ser detectados por la función correctora de pruebas de la ADN polimerasa III.")
81
Metal Efecto genotóxico
Arsénico Cáncer de piel, efectos sobre la reproducción, mutagénico. Cadmio Mutagénico, sarcomas. Cromo Cáncer pulmonar, mutagénico. Níquel Carcinoma nasal, mutagénico. Plomo Anemia, linfomas, carcinomas y sarcomas renales, mutagénico.

82
Mecanismo Resultado en el ADN
Mutágeno Mecanismo Resultado en el ADN Agentes alquilantes (nitrosourea,nitrosoguanidina) Se une covalentemente y forma sitios apurínicos Transición y transversión Agentes desaminantes (ácido nitroso) Adenina-hipoxantina y citosina-uracilo Transición Base análoga (2-aminopurina) Substitución durante la replicación del ADN Agente intercalante (antridinas, antraciclinas) Inserción o eliminación de pares de bases Cambio de cuadro Fraccionadores de las cadenas (radiaciones ionisantes) Translocación cromosomal Cambio de una o más bases
 Se une covalentemente y forma sitios apurínicos. Transición y transversión. Agentes desaminantes (ácido nitroso) Adenina-hipoxantina y citosina-uracilo. Transición. Base análoga (2-aminopurina) Substitución durante la replicación del ADN. Agente intercalante (antridinas, antraciclinas) Inserción o eliminación de pares de bases. Cambio de cuadro. Fraccionadores de las cadenas (radiaciones ionisantes) Translocación cromosomal. Cambio de una o más bases.")
84
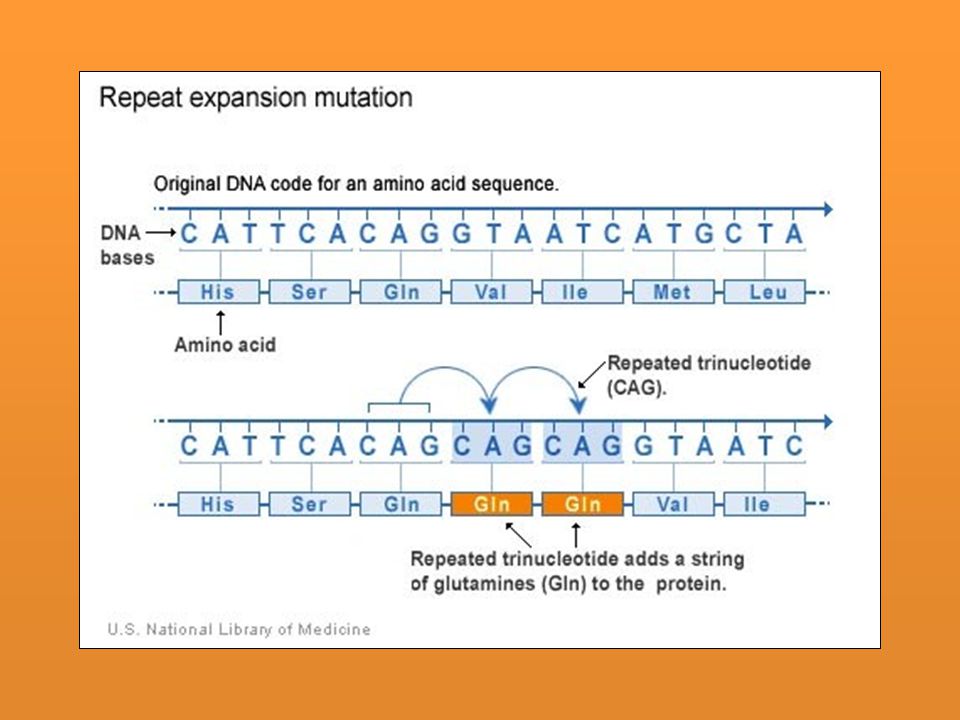
MUTAGENOS BIOLOGICOS La disposición exacta de los genes en los cromosomas no está necesariamente fijada. En ciertas condiciones algunos genes son capaces de cambiar de posición. Este proceso se denomina trasnposición, el cual típicamente es un proceso raro (10-5 a por generación). Necesaria la presencia de un gene que codifica para una enzima transposasa
. Necesaria la presencia de un gene que codifica para una enzima transposasa.")
86
III. POR EL TIPO CELULAR - Somática Si la mutación ocurre en una célula que desarrolla un tejido somático, dará origen a una población de células mutantes idénticas clones. Si la mutación es dominante se expresará en el fenotipo de aquellos organismos híbridos. Si es recesiva no se expresará ya que quedará enmascarada por el alelo salvaje (dominante), una segunda mutación puede crear una mutación homocigota recesiva, pero es un evento raro. ¿Son heredables?: por definición NO SE HEREDAN, ya que las células somáticas son aquellas que no origina progenie. Sin embargo hay que recordar que porciones de plantas (con mutaciones somáticas) pueden originar nuevos individuos con su eventual línea germinal. En vegetales se heredan por injerto.
, una segunda mutación puede crear una mutación homocigota recesiva, pero es un evento raro. ¿Son heredables : por definición NO SE HEREDAN, ya que las células somáticas son aquellas que no origina progenie. Sin embargo hay que recordar que porciones de plantas (con mutaciones somáticas) pueden originar nuevos individuos con su eventual línea germinal. En vegetales se heredan por injerto.")
87
- Gamética o de la línea germinal
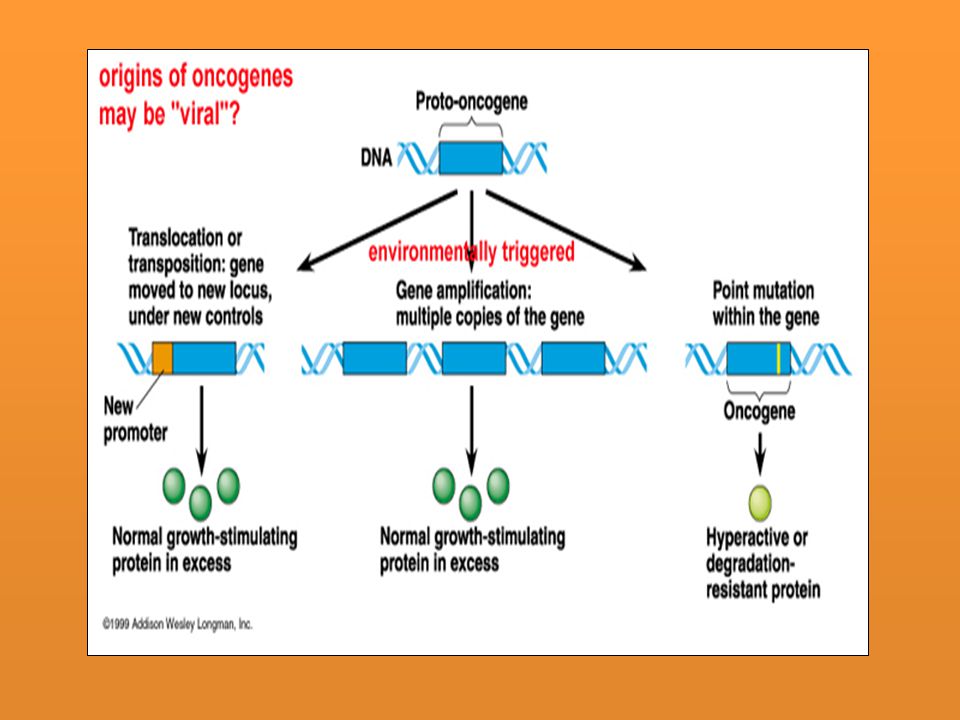
Consecuencias en un organismo multicelular: un ejemplo de mutaciones somáticas son las células que origina el cáncer. cuando ciertos genes (proto-oncogenes) sufren una mutación las células inician una secuencia descontrolada de divisiones celulares resultando en una masa de células denominada tumor. - Gamética o de la línea germinal Si la mutación se produce en una de las células sexuales inevitablemente pasará a la descendencia (en tanto ésta participe de la fecundación). Un fenotipo completamente normal puede tener células mutantes en su línea germinal y sólo se detectará en su descendencia. Casos especiales: Algunas mutaciones pueden afectar al cromosoma X y éste encontrarse inactivado al azar en las hembras de mamíferos, por lo que pueden no expresar el fenotipo. Ej: hemofilia ligada al cromosoma X.
 sufren una mutación las células inician una secuencia descontrolada de divisiones celulares resultando en una masa de células denominada tumor. - Gamética o de la línea germinal. Si la mutación se produce en una de las células sexuales inevitablemente pasará a la descendencia (en tanto ésta participe de la fecundación). Un fenotipo completamente normal puede tener células mutantes en su línea germinal y sólo se detectará en su descendencia. Casos especiales: Algunas mutaciones pueden afectar al cromosoma X y éste encontrarse inactivado al azar en las hembras de mamíferos, por lo que pueden no expresar el fenotipo. Ej: hemofilia ligada al cromosoma X.")
88
IV. POR SU DIRECCION Hacia delante: Silvestre normal Hacia atrás: Normal silvestre Las mutaciones puntuales son reversibles. Se llaman revervientes a las cepas en la que aparece restaurado el fenotipo silvestre que se perdió en el mutante Consiste en recuperar la secuencia exacta de nucleótidos en el ADN, un ejemplo de esta situación sería: AAA (lys)→GAA(glu)→AAA(lys). A veces se habla de retromutación en sentido funcional, de manera, que se recupera la actividad enzimática normal pero no se recupera la secuencia original de nucleótidos en el ADN, por ejemplo: UCC(ser)→UGC(cys)→AGA(ser). Sin embargo, el concepto de retromutación funcional no se diferencia del concepto de mutación supresora y puede inducir a error.
→GAA(glu)→AAA(lys). A veces se habla de retromutación en sentido funcional, de manera, que se recupera la actividad enzimática normal pero no se recupera la secuencia original de nucleótidos en el ADN, por ejemplo: UCC(ser)→UGC(cys)→AGA(ser). Sin embargo, el concepto de retromutación funcional no se diferencia del concepto de mutación supresora y puede inducir a error.")
89
Magnitud del efecto fenotípico
Cambio en la tasa de mutación: Algunos alelos sólo pueden distinguirse por la frecuencia con la que mutan. Isoalelos: Producen fenotipos idénticos en combinaciones homócigas o heterócigas de unos con otros, pero se distinguen cuando se combinan con alelos diferentes. Mutantes que afectan la viabilidad: Subvital: La viabiliadad relativa es > 10% pero < 100% Semiletales: Causan más del 90% pero < del 100% de la mortalidad Letales: Matan a todos antes de la edad adulta Cuando en una población < del 1% tiene el gene alterado se considera mutación (no recurrencia), cuando es mayor se considera polimorfismo (recurrencia)
, cuando es mayor se considera polimorfismo (recurrencia)")
90
Efecto a nivel individual
Neutra: No altera la viabilidad: color de ojos, polidactilia Facultativa: El efecto a nivel individual depende de los factores ecológicos o ambientales: alopecia Letal: Nace muerto o muere al poco tiempo: vegetales sin clorofila D) Beneficiosos: Las mutaciones pueden inducir cambios que adaptan los seres vivos al medio ambiente . Las tasas de mutación se consideran que ocurre entre 1 de cada gametos o 1 de cada A pesar de que la incidencia de las mutaciones es relativamente grande en relación con el número de organismos de cada especie,
 Beneficiosos: Las mutaciones pueden inducir cambios que adaptan los seres vivos al medio ambiente . Las tasas de mutación se consideran que ocurre entre 1 de cada gametos o 1 de cada A pesar de que la incidencia de las mutaciones es relativamente grande en relación con el número de organismos de cada especie,")
91
la evolución no depende ni mucho menos de las mutaciones que surgen en cada generación, sino de la acumulación de toda la variabilidad durante la evolución de las especies.

92
Reparación del ADN El ADN es modificado: agentes, endonucleasas.
Como contrapartida los organismos han desarrollado mecanismos para reparar las alteraciones sufridas en su ADN y garantizar así la estabilidad de la molécula . Como primera medida se produce un bloqueo de la replicación y se ponen en marcha un sistema de emergencia denominado abreviadamente SOS para reparar los daños y permitir que la célula se replique y pueda seguir viviendo. El sistema consta de al menos tres genes denominados recA, umuC y umuD que produce un relajamiento de la especificidad de apareamiento de la ADN pol III de E. coli. Los sistemas de reparación del ADN, principal herramienta de las células para controlar estas alteraciones del material genético, van perdiendo eficiencia con el proceso de envejecimiento, contribuyendo progresivamente con la edad y generando un efecto acumulativo en las células.

93
Los mecanismos de reparación pueden funcionar antes o después de la replicación del ADN. Su eficiencia varía, ya que pueden reparar eficientemente, es decir, sin errores, situación que se presenta cuando la exposición a agentes genotóxicos es baja; o bien reparar de manera ineficiente, promoviendo errores en el ADN, lo que depende de la saturación del primer mecanismo y que generalmente ocurre cuando hay exposiciones altas. Ambos mecanismos se ven afectados por numerosas variables además de la exposición. Dependen también de la estructura química del mutágeno, del tipo de producto formado y de la cantidad de daño inducido.

94
SISTEMAS QUE EVITAN LOS ERRORES ANTES DE QUE OCURRAN
Superóxido dismutasa: este enzima convierte los radicales superóxido en peróxido de hidrógeno. Catalasa: este enzima convierte el peróxido de hidrógeno en agua. Gen mutT: este gen codifica para un enzima que impide la incorporación de la 8-oxo-G al ADN. Este enzima hidroliza el trifosfato de la 8-oxo-G a la forma monofosfato.

95
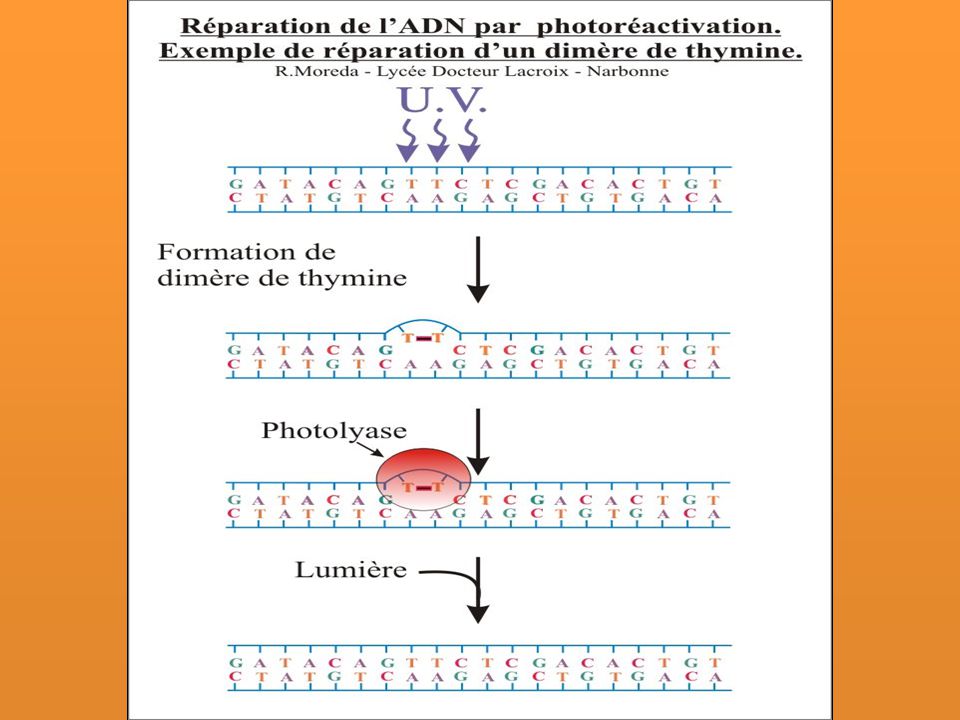
Fotorreactivación Enzimática
Es in situ, es decir es un sistema de reparación directa de los daños producidos por la luz UV. La enzima Fotoliasa codificada por el gen phr reconoce en la oscuridad los dímeros de Timina y se une a ellos, y cuando se expone a la luz (mediante un fotón) deshace el dímero de Timinas. Descubierto por Kelner cuando observó un incremento en la tasa de supervivencia de bacterias expuestas a UV
 deshace el dímero de Timinas. Descubierto por Kelner cuando observó un incremento en la tasa de supervivencia de bacterias expuestas a UV.")
96
Xeroderma pigmentosum

98
Transferasa de grupos alquilo (metilo o etilo):
Elimina los gupos alquilo producidos por el EMS o por NG. La enzima metiltransferasa transfiere el grupo metilo de la O-6-metilguanina a una cisteína (cys) de la enzima.Es un sistema de reparación directa.
 de la enzima.Es un sistema de reparación directa.")
99
Reparación por escisión de bases
Se pone en marcha cuando la lesión radioinducida origina dímeros de pirimidina (T-T, C-T y C-C). Cuando se reconoce el dímero, una glicosidasa endonucleasa Uvr ABC, utilizando ATP en la reacción, corta la hebra de ADN dañada a ambos lados de la lesión, a continuacón una helicasa elimina un fragmento con aproximadamente 12 nucleótidos entre los que se encuentran los que están dañados. Posteriormente, estos nucleótidos son nuevamente sintetizados por la polimerasa que utiliza la hebra complementaria intacta de molde. Finalmente una ligasa sella la unión
. Cuando se reconoce el dímero, una glicosidasa endonucleasa Uvr ABC, utilizando ATP en la reacción, corta la hebra de ADN dañada a ambos lados de la lesión, a continuacón una helicasa elimina un fragmento con aproximadamente 12 nucleótidos entre los que se encuentran los que están dañados. Posteriormente, estos nucleótidos son nuevamente sintetizados por la polimerasa que utiliza la hebra complementaria intacta de molde. Finalmente una ligasa sella la unión.")
104
Reparación AP El sitio apurinico o apirimidinico o sitio AP, es removido por una enzima endonucleasa AP, el hueco es llenado por las ADNpolimerasa I y la ligasa. Reparación de las sedes apurínicas o apirimidínicas. La llevan a cabo las Endonucleasas AP de la clase I que cortan por el extremo 3' y las de la clase II que cortan por el extremo 5'. Una exonucleasa elimina una pequeña región que contiene entre dos y 4 nucleótidos, la ADN polimerasa I rellena el hueco y la Ligasa sella los extremos.

105
Reparación mediante glucosidasas
Estas enzimas detectan las bases dañadas y las retiran rompiendo el enlace N-glucosídico con el azúcar. Como consecuencia se origina una sede AP que se repara de la forma indicada anteriormente (reparación AP). La Glucosidasa de Uracilo elimina el Uracilo (U) del ADN. La Glucoxidasa de Hipoxantina, elimina la Hipoxantina (H) del ADN. Además de estas dos glucosidasas existen otras diferentes
. La Glucosidasa de Uracilo elimina el Uracilo (U) del ADN. La Glucoxidasa de Hipoxantina, elimina la Hipoxantina (H) del ADN. Además de estas dos glucosidasas existen otras diferentes.")
106
Reparación de roturas simples de cadena
Utiliza el mecanismo de escisión de bases. La reparación de roturas simples de cadena es un proceso rápido, ya que el 50% de las mismas se reparan en aproximadamente 15 minutos. Uno de los genes implicados en este tipo de reparación es el que codifica la enzima nuclear PARP-1 que reconoce las roturas simples de cadena.

107
Reparación de roturas dobles de cadena: En este caso no existe una cadena intacta de ADN para ser utilizada de molde en el proceso de reparación. Las cadenas con rotura doble son reagrupadas entre 4 y 6 horas por la gran complejidad del proceso que casi siempre conduce a errores o mutaciones que conducen a la muerte celular, aunque existen células que soportan el daño, como las tumorales. Existen dos mecanimos de reparación que son: Reparación por recombinación de cromosomas homólogos: Están implicados al menos 5 genes: Ku 70, ku 80, DNA-PCKcx, ligasa IV, Xrcc4 Además existen otras dos proteínas como la ATM y la ATR que se activan al unirse a los extremos rotos del ADN originados por roturas dobles de cadena y comienzan la reparación. Algunas de estas proteínas intervienen en la interrupción del ciclo celular para que la célula tenga tiempo de reparar la lesión o inducir la apoptosis. También está involucrada el gen BCRA1 y BCRA2.

108
Reparación por unión de extremos no homólogos: Requiere un locus recíproco en la cromátida hermana o secuencias de ADN que posean gran homología con aquella que ha sido dañada. Se activa cuando la lesión originada conlleva pérdida de material genético. Es un mecanismo de reparación minoritario, dada la baja posibilidad de encontrar el locus recíproco dentro del genoma completo de la célula.

109
Reparación de apareamientos incorrectos
La reparación de apareamientos incorrectos posterior a la replicación requiere la existencia de un sistema que sea capaz de realizar las siguientes operaciones: Reconocer las bases mal apareadas. Determinar cual de las dos bases es la incorrecta. Eliminar la base incorrecta y sintetizar. Esta reparación la realizan los productos de los genes mutH, mutL, mutS y mutU. Además, para distinguir la hélice de nueva síntesis de la hélice molde y así saber eliminar la base incorrecta, el sistema consiste utiliza el hecho de que la hélice de nueva síntesis tarda un cierto tiempo en metilarse la Adenina (A) de la secuencia GATC, mientras que la A de la secuencia GATC de la hélice molde ya está metilada. El enzima que reconoce la secuencia GATC metilando la A que contiene es la Metilasa de Adenina
 de la secuencia GATC, mientras que la A de la secuencia GATC de la hélice molde ya está metilada. El enzima que reconoce la secuencia GATC metilando la A que contiene es la Metilasa de Adenina.")
112
Reparación post replicación
Mecanismo que quedaría como alternativa cuando a pesar de los mecanismos anteriores aún quedan lesiones en el ADN que tiene que iniciar su replicación. Cuando esto sucede, el aparato de replicación salta al llegar a tales lesiones y deja en la cadena que se está sintetizando el hueco correspondiente a la lesión de la cadena molde, continuando su actividad.. Los especios pueden ser 103 pb. Posteriormente el hueco por dos mecanismos: A través de la desoximetiltransferasa terminal se pueden dar la adición de bases. Esta enzima no necesita de una cadena molde, actúa añadiendo cualquier base en el extremo 3’ de la cadena portadora del hueco, pudiendo causar mutaciones como transición, transversión, adición, deleción, etc. Por recombinación

113
Per tal de determinar com actuen els mecanismes de reparació en el conjunt del genoma humà, els científics de la UAB han estudiat línies cel·lulars derivades de pell de persones sanes i de pacients deficients en la reparació de les lesions al DNA produïdes per la radiació solar, un desordre genètic anomenat xeroderma pigmentosa. En els afectats per aquest desordre (popularitzat pels personatges dels nens de la pel·lícula Los Otros), els mecanismes de reparació no actuen quan la llum ultraviolada incideix sobre les cèl·lules de la pell, la qual cosa els hi provoca una acumulació de mutacions i, per tant, una incidència altíssima de melanoma (càncer de pell). Han pogut observar com la reparació de les lesions causades pels raigs ultraviolats es concentra en les regions del genoma més riques en gens i, per tant, hi ha una reparació preferencial de la part més rellevant del genoma, anomenada transcriptoma. A tall d'exemple, el cromosoma 19, el de més densitat i activitat gènica, presenta elevats nivells de reparació, mentre que en el cromosoma 4, un dels més pobres en gens, pràcticament no hi ha reparació preferencial de les mutacions induïdes per la llum ultraviolada.
, els mecanismes de reparació no actuen quan la llum ultraviolada incideix sobre les cèl·lules de la pell, la qual cosa els hi provoca una acumulació de mutacions i, per tant, una incidència altíssima de melanoma (càncer de pell). Han pogut observar com la reparació de les lesions causades pels raigs ultraviolats es concentra en les regions del genoma més riques en gens i, per tant, hi ha una reparació preferencial de la part més rellevant del genoma, anomenada transcriptoma. A tall d exemple, el cromosoma 19, el de més densitat i activitat gènica, presenta elevats nivells de reparació, mentre que en el cromosoma 4, un dels més pobres en gens, pràcticament no hi ha reparació preferencial de les mutacions induïdes per la llum ultraviolada..")
114
Màquina hipotética de reparación del ADN

115
New research shows that a single gene controls the loss of bony plates (shown in red) when sticklebacks evolve from ocean (top) to freshwater (bottom) varieties found around the world.
 when sticklebacks evolve from ocean (top) to freshwater (bottom) varieties found around the world.")
117
Tumores causados por Salmonella typhimurium

Presentaciones similares