Descargar la presentación
La descarga está en progreso. Por favor, espere

1
La secuencia de aminoácidos de la proteína no siempre refleja
la secuencia contínua de nucleótidos en el genoma splicing de pre-mRNA (eliminación de intrones) splicing alternativo (varios polipéptidos a partir de la misma secuencia de DNA) corrimiento del marco de lectura (translational frameshifting) edición del mRNA splicing de proteínas (eliminación de inteínas) procesamiento proteolítico de polipéptidos (en algunos casos se obtienen productos alternativos en diferentes tipos celulares) Glicosilación, fosforilación y otras modificaciones covalentes de proteínas
 splicing alternativo (varios polipéptidos a partir de la misma secuencia de DNA) corrimiento del marco de lectura (translational frameshifting) edición del mRNA. splicing de proteínas (eliminación de inteínas) procesamiento proteolítico de polipéptidos (en algunos casos se obtienen productos alternativos en diferentes tipos celulares) Glicosilación, fosforilación y otras modificaciones covalentes de proteínas.")
2
los RNAs pueden procesarse de diferente manera eliminando parte del producto de transcripción original

4
splicing alternativo del gen de calcitonina

5
RNA editing

6
Edición de mRNA

7
Translational Frameshift

8
terminación de la traducción Frameshift
UGA C UGA GAC RF2 RF2 Pausa Unión de RF2 terminación de la traducción Frameshift (UGAC se lee como GAC y sigue +1)
")
9
Corrimiento del marco de lectura Translational frameshifting
HIV The group antigens form the viral core structure, RNA genome binding proteins, and are the major proteins comprising the nucleoprotein core particle. Reverse transcriptase is the essential enzyme that carries out the reverse transcription process that take the RNA genome to a double-stranded DNA preintegrate form

10
p55 proteasa integrasa RNAsa p10 Transcriptasa reversa p50
Proteasa p10 p55 proteasa

11
Modificación de proteínas
1. las proteínas sufren modificaciones post-traducionales (y co-traduccionales) 2. la actividad biológica puede depender de las modificaciones 3. la espectrometría de masas es uno de los mejores métodos para identificar las modificaciones
 2. la actividad biológica puede depender de las modificaciones. 3. la espectrometría de masas es uno de los mejores métodos para identificar las modificaciones.")
12
Protein Post-translational Modifications
1. Folding and Processing of Proteins -During translation proteins fold as they exit ribosome -Some proteins can assume native 3D structure spontaneously -Other proteins may require chaperones

13
Protein Post-translational Modifications
2. Amino-terminal and carboxyterminal modifications -Cleavage of f-Met from bacterial proteins or Met from eukaryotic proteins. Other amino acids may be trimmed as well. -Acetylation of Met or other N-terminal amino acids -Removal of signal peptide for secreted or membrane proteins -Removal of C-terminal amino acids.

14
Protein Post-translational Modifications
3. Modification of Individual Amino Acids a. Phosphorylation -Enzymatic reaction by specific kinases - Usually on Ser, Thr, Tyr

15
Protein Post-translational Modifications
3. Modification of Individual Amino Acids b. Carboxylation Addition of extra carboxyl groups to Asp and Glu c. Methylation Addition of methyl groups to Lys and Glu

16
Protein Post-translational Modifications
3. Modification of Individual Amino Acids d. Isoprenylation -Addition of an isoprenyl group to a protein at either the C-terminus or the N-terminus -Derived from pyrophosphate intermediate in cholesterol biosynthesis

17
Protein Post-translational Modifications
3. Modification of Individual Amino Acids e. Addition of prosthetic groups Covalently bound prosthetic group – required for activity Example: Cytochrome C -- Heme group

18
Protein Post-translational Modifications
3. Modification of Individual Amino Acids f. Proteolytic Processing Some types of proteins are synthesized as a larger, inactive precursor protein and must be cleaved for activity g. Formation of disulfide bonds -Spontaneous cross-linking at Cys residues -Brought into proximity by folding -Helps to stabilize 3D structure

19
Protein Post-translational Modifications
3. Modification of Individual Amino Acids h. Glycosylation -Addition of oligosaccharides to proteins -Usually at Asn -Sugars are transferred from dolichol-P -Present in ER

20
Protein Post-translational Modifications
4. Methods to Discover Modifications a. Enzymatic methods Phosphatases – remove phosphate groups Glycosylases – remove carbohydrates b. Physical methods 1. Hydrolysis in 6N HCl – amino acid composition 2. Digestion with specific protease or chemical agent 3. Sequence of peptide by Edman Chemistry 4. Mass Spectrometry (MALDI-TOF)
")
21
procesamiento proteolítico
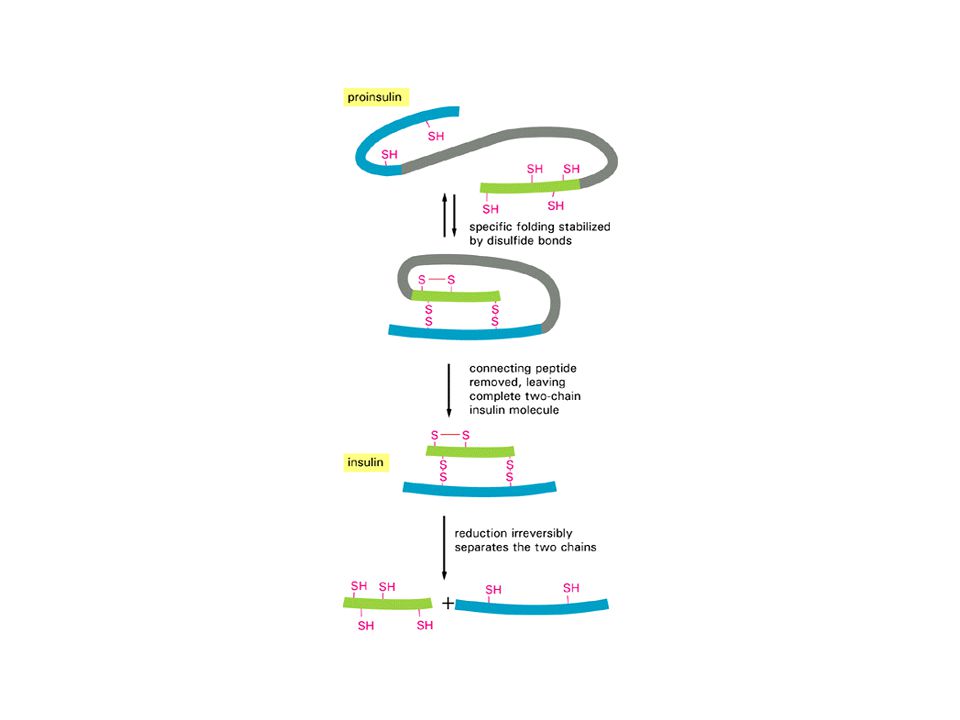
insulina: sobre-simplificación

23
Rutas de procesamiento alternativas de la prohormone pro-opiocortina
Alternative processing pathways of the prohormone pro-opiocortin. The initial cleavages are made by membrane-bound proteases that cut next to pairs of positively charged amino acid residues (Lys-Arg, Lys-Lys, Arg-Lys, or Arg-Arg pairs), and trimming reactions then produce the final secreted products. Different cell types contain different processing enzymes, so that the same prohormone precursor can be used to produce different peptide hormones. In the anterior lobe of the pituitary gland, for example, only corticotropin (ACTH) and b-lipotropin are produced from pro-opiocortin, whereas in the intermediate lobe of the pituitary, mainly a-MSH, g-lipotropin, b-MSH, and b-endorphin are produced.
, and trimming reactions then produce the final secreted products. Different cell types contain different processing enzymes, so that the same prohormone precursor can be used to produce different peptide hormones. In the anterior lobe of the pituitary gland, for example, only corticotropin (ACTH) and b-lipotropin are produced from pro-opiocortin, whereas in the intermediate lobe of the pituitary, mainly a-MSH, g-lipotropin, b-MSH, and b-endorphin are produced.")
24
Protein Splicing T. littoralis is the source of vent DNA polymerase, used extensively in PCR. When scientists at New England Biolabs cloned the gene for the enzyme, they were surprised to find that it contains an extra sequence of non-DNA-polymerase. They assumed it was an intron, but found that the mRNA isn't spliced and the extra sequence is translated into a novel domain in the enzyme. This polypeptide domain is a peptidyltransferase that specifically splices itself out of the DNA polymerase, rejoining the 2 parts of the DNA polymerase as it leaves. This protein-splicing reaction is remarkably analogous to RNA-splicing by introns, and so it's called an 'intein' (intervening protein). Once the intein has removed itself from the DNA polymerase, it has another activity - it is a transposase. This enzyme cleaves DNA specifically at the ends of the intein-encoding sequence and directs a DNA repair process that results in the insertion of the intein DNA into other protein-encoding genes - in other words, the intein DNA is also a transposon.
. Once the intein has removed itself from the DNA polymerase, it has another activity - it is a transposase. This enzyme cleaves DNA specifically at the ends of the intein-encoding sequence and directs a DNA repair process that results in the insertion of the intein DNA into other protein-encoding genes - in other words, the intein DNA is also a transposon.")
25
Protein splicing

26
proteínas de secreción: la vía secretoria secretory pathway

28
SÍNTESIS DE PROTEÍNAS EN EUCARIOTES: SEÑALES DE LAS PROTEÍNAS QUE
DETERMINAN SU DIRECCIONAMIENTO CELULAR.

29
The Nobel Prize in Physiology or Medicine 1999
The Nobel Prize in Physiology or Medicine 1999 "for the discovery that proteins have intrinsic signals that govern their transport and localization in the cell" Günter Blobel USA Rockefeller University New York, NY, USA; Howard Hughes Medical Institute

30
Signal hypothesis 1971: “Las proteínas secretadas al espacio extracelular contienen una señal intrínseca que las dirige hacia y a través de las membranas”

31
Milstein, C. et al., Nature New Biology 239: 117-120, 1972
Signal hypothesis La traducción de poly(A) mRNA de células de mieloma (principalmente mRNA de IgG) en un sistema libre de células carente de vesículas microsomales genera una proteína 2-3kDa mayor. El mapa peptídico indica que la extensión se encuentra en el amino terminal Milstein, C. et al., Nature New Biology 239: , 1972
 mRNA de células de mieloma (principalmente mRNA de IgG) en un sistema libre de células carente de vesículas microsomales genera una proteína 2-3kDa mayor. El mapa peptídico indica que la extensión se encuentra en el amino terminal. Milstein, C. et al., Nature New Biology 239: ,")
32
preparación de microsomas

33
Experimentos utilizando microsomas
Las proteínas del lumen de los microsomas no son atacadas por proteasas

34
Descubriendo cómo trabaja la secuencia señal…

35
Dobberstein and Blobel, 1975

36
Descubriendo cómo trabaja la secuencia señal…
La secuencia señal es hidrolizada en el lumen del ER La secuencia señal dirige la proteína al microsoma. Este proceso es una translocación co-traduccional

37
“La proteína atraviesa la membrana mediante un canal”
Signal hypothesis 1975: “La señal consiste en una secuencia de aminoácidos que forma parte integral de la proteína” “La proteína atraviesa la membrana mediante un canal”

38
Ruta secretoria Las proteínas destinadas para la secreción o incorporación al ER, Golgi, lisozoma o membrana plasmática son sintetizadas en ribosomas asociados a membrana y transferidas al RER durante su síntesis

39
¿Qué elementos son requeridos para entrar a la ruta secretoria?
Una señal en la proteína Un receptor que reconozca la señal y direccione la proteína a la membrana correcta Una maquinaria de translocación, un canal Energía para abrir la compuerta y translocar la proteína

41
Estructura de la partícula de reconocimiento de señal “signal recognition particle” (SRP)
")
42
¿Qué elementos son requeridos para entrar a la ruta secretoria?
Una señal en la proteína Un receptor que reconozca la señal y direccione la proteína a la membrana correcta Una maquinaria de translocación (un canal) Energía para abrir la compuerta y translocar la proteína
 Energía para abrir la compuerta y translocar la proteína.")
43
La hidrólisis de GTP potencia el transporte al ER

44
Direccionamiento al lumen del ER

45
Anclaje a membrana

46
Topología de proteínas integrales de membrana sintetizadas

47
Una secuencia interna topogénica dirige la insersión de algunas proteínas de transmembrana

48
Se requieren múltiples secuencias topogénicas para las proteínas de transmembrana de pasaje múltiple

49
Overview de la ruta secretoria
Espacio extracelular Overview de la ruta secretoria trans Golgi Cis Golgi RE rugoso

50
La N-glicosilación de las proteínas comienza en el ER

51
La N-glicosilación de las proteínas comienza en el ER

52
Direccionamiento de proteínas codificadas por el genoma nuclear

53
Las proteínas destinadas al citosol o a ser incorporadas en el núcleo, mitocondria, cloroplasto o peroxisoma son sintetizadas a partir de ribosomas libres POST TRADUCCIONALES!!

54
Direccionamiento a los diferentes compartimientos sub-mitocondriales

55
¿Qué elementos son requeridos para el direccionamiento mitocondrial?
Una o más señales en la proteína Un receptor que reconozca la señal y direccione la proteína a la membrana correcta Una maquinaria de translocación, un canal Energía para abrir la compuerta y translocar la proteína Chaperonas para desplegar la proteína ya sintetizada

56
emplea ATP en el citosol,
fuerza protón motriz a través de la membrana interna ATP en la matriz emplea ENERGIA CHAPERONAS citosólicas y Matriz mitocondrial

57
Se requieren múltiples rutas y señales para dirigir las proteínas a los distintos compartimientos submitocondriales

58
Intermembrane-Space Proteins

59
Se requieren múltiples rutas y señales para dirigir las proteínas a los distintos compartimientos submitocondriales

60
Direccionamiento de proteínas codificadas por el genoma nuclear

61
Direccionamiento a Cloroplasto

62
Importación de proteínas a cloroplasto

63
¿Qué elementos son requeridos para entrar al cloroplasto?
Una o más señales en la proteína Un receptor que reconozca la señal y direccione la proteína a la membrana correcta Una maquinaria de translocación, un canal Energía para abrir la compuerta y translocar la proteína Chaperonas para desplegar la proteína ya sintetizada

64
Peroxisoma Secuencias C- o N-terminal dirigen la entrada de proteínas plegadas a la matriz del peroxisoma

65
Direccionamiento al núcleo

66
Las proteinas con la señal de localización nuclear (NLS) son reconocidas por receptores y transportadas al núcleo Antígeno T de SV40

67
El complejo de poro nuclear (NPC)
The nuclear pore complex

68
Modelo de importación de proteínas citosólicas con NLS

69
¿Qué elementos son requeridos para la entrada al núcleo?
Una REGION SEÑAL en la proteína (no es clivada!!) Un receptor que reconozca la señal y direccione la proteína Una maquinaria de translocación, un canal Energía para translocar la proteína
 Un receptor que reconozca la señal y direccione la proteína. Una maquinaria de translocación, un canal. Energía para translocar la proteína.")
70
Características de las secuencias señal

71
After insertion into the ER membrane, some proteins are transferred to a GPI anchor

72
Anchoring of integral proteins to the plasma membrane by hydrocarbon chains

73
Post-translational modifications and quality control in the rough ER
Newly synthesized polypeptides in the membrane and lumen of the ER undergo five principal modifications Formation of disulfide bonds Proper folding Addition and processing of carbohydrates Specific proteolytic cleavages Assembly into multimeric proteins

74
Disulfide bonds are formed and rearranged in the ER lumen

75
ER-resident proteins often are retrieved from the cis-Golgi

76
Different structures characterize N- and O-linked oligosaccharides

77
The immediate precursors in the synthesis of oligosaccharides are nucleoside diphosphate or monophosphate sugars

78
Specific sugars are linked by specific glycosyltransferases

79
Sugar nucleotides and free nucleotides are exchanged by antiporters in the ER membrane

80
ABO blood type is determined by two glycosyltransferases

81
ABO blood groups

82
Mannose 6-phosphate residues target proteins to lysosomes

83
The mannose 6-phosphate (M6P) pathway
 pathway")
Presentaciones similares