Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Introducción a las enzimas

2
What is a catalyst? Example:
“A catalyst is a compound which enhances the rate of a chemical reaction without being destroyed or incorporated in the product” (IUPAC) Example:
 Example:")
3
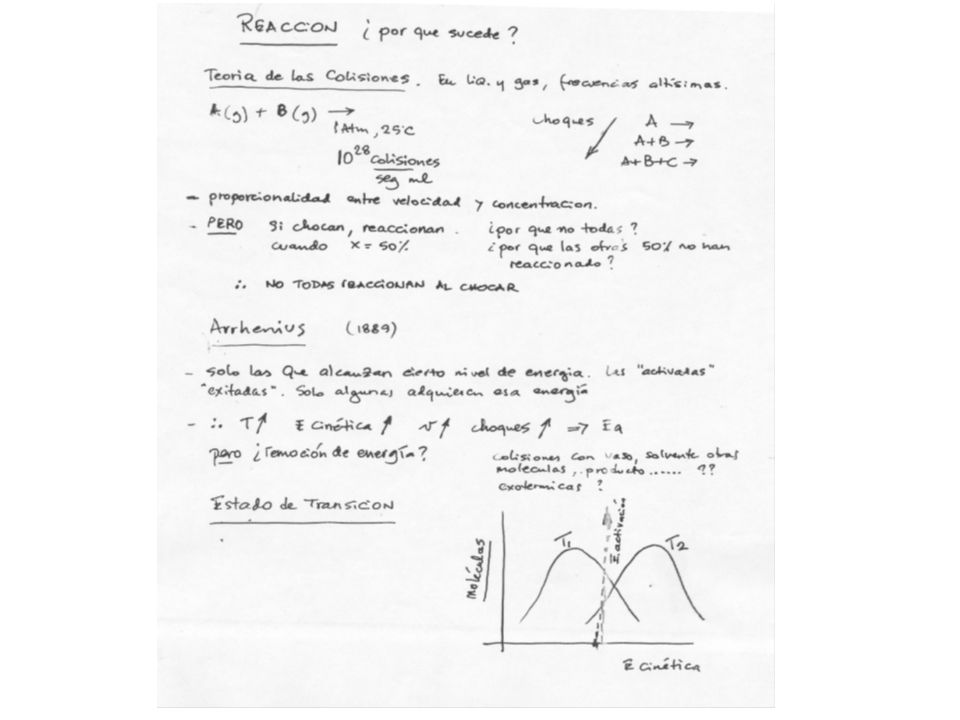
ENZIMAS

12
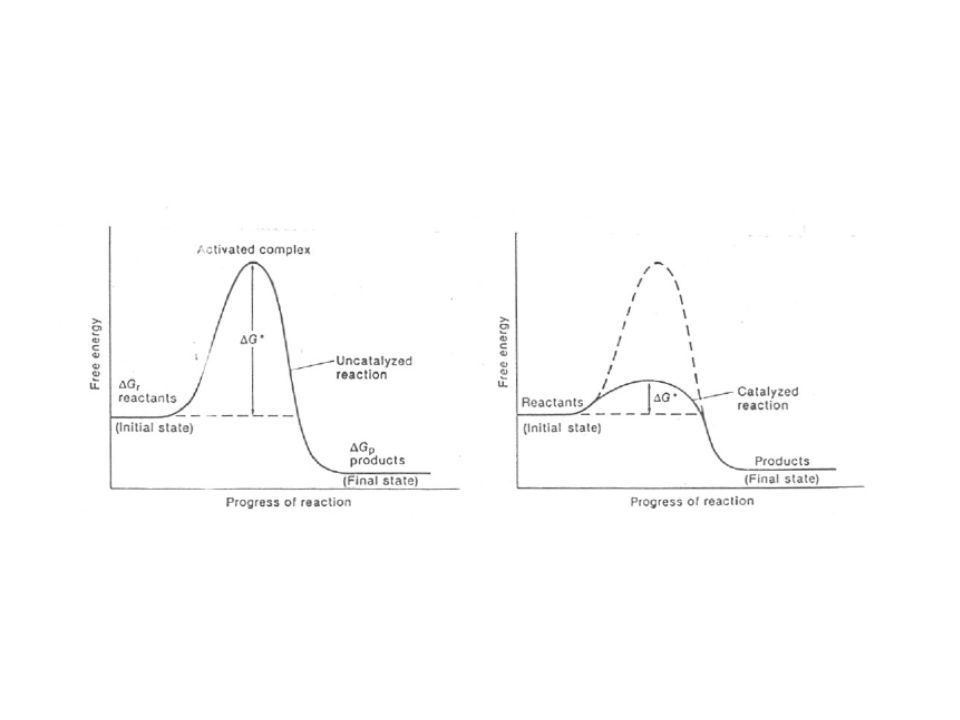
Features of a catalyst:
Makes an alternative reaction path in which less activation energy is needed. Equally increases the rate of the back and forth reaction reaction catalysis has no effect on equilibrium position!

13
Reaction equations and reaction rate
1 A reaction like A B may run via route 1; the rate is determined by the magnitude of Ea(1) Alternatively, the reaction runs via an intermediate, like 2. The rate is determined by the slowest step (= the one with the highest Ea). Alternatively, the reaction can run via more intermediates, like 3 Ea(1) 2 E 3 A B
 Alternatively, the reaction runs via an intermediate, like 2. The rate is determined by the slowest step (= the one with the highest Ea). Alternatively, the reaction can run via more intermediates, like 3. Ea(1) 2. E. 3. A. B.")
15
What kinds of catalysts are there around?
Organic Inorganic Biological (biocatalyst) Other way of subdivision: Homogeneous = freely dissolved in solution organic catalyst organometallic complex enzyme in water Heterogeneous = solid, in liquid or gaseous environment inorganic catalyst (e.g. zeolite) immobilised enzyme enzyme in an organic solvent
 Other way of subdivision: Homogeneous = freely dissolved in solution. organic catalyst. organometallic complex. enzyme in water. Heterogeneous = solid, in liquid or gaseous environment. inorganic catalyst (e.g. zeolite) immobilised enzyme. enzyme in an organic solvent.")
16
Determination of reaction rates
Determination of the concentration of a compound (substrate, product), as a function of time. The method used depends on the properties of the compound. Chromatography Example:
, as a function of time. The method used depends on the properties of the compound. Chromatography. Example:")
17
GC analysis of the esterification of racemic menthol with vinyl acetate, catalysed by Candida rugosa lipase, in diisopropyl ether as the solvent. I.S., internal standard (decane). An enantioselective GC column is used, so the enantiomers are separated. There is only one product peak because the reaction is stereoselective.
. An enantioselective GC column is used, so the enantiomers are separated. There is only one product peak because the reaction is stereoselective..")
18
Another example: the chlorination of barbituric acid (1) by chloroperoxidase from Caldariomyces fumago
 by chloroperoxidase from Caldariomyces fumago")
19
UV spectroscopy Example: chlorination of 1 by chloroperoxidase Can be done quantitatively through Lambert-Beer’s law: A = e.c.d

20
Example: oxime formation
IR spectroscopy Example: oxime formation Problem: not quantitative Polarimetry (only for chiral compounds) Example: hydrolysis of sucrose: C12H22O11 + H2O C6H12O C6H12O6 sucrose + water glucose fructose [a]D20 = +66,5° [a]D20 = +52° [a]D20 = –92° (together [a]D20 = –40°) Can be done quantitatively
 Example: hydrolysis of sucrose: C12H22O11 + H2O C6H12O6 + C6H12O6. sucrose + water glucose + fructose. [a]D20 = +66,5° [a]D20 = +52° [a]D20 = –92° (together [a]D20 = –40°) Can be done quantitatively.")
21
Sucrose Glucose Fructose + Invertase (IT) IT Reducing sugars
Non-reducing sugar Reducing Power IT Sucrose Glucose Fructose + HOCH2 O OH HOCH2 O OH O HOCH2 OH HOCH2 HO O 6 5 4 3 2 1 1 2 3 4 5 6 1 2 b b H2O 1 2 3 4 5 6 CHO H-C-OH HO-C-H H2-C-OH H2C-OH C=O HO-C-H H-C-OH H2-C-OH Michaelis 及 Menten 兩位以當時一個普遍研究的酵素 invertase (轉化脢) 作為研究對象,其催化反應如上圖所示,可以把非還原糖的蔗糖,轉化成為兩個具有還原力的單糖 (葡萄糖及果糖)。因此做實驗時,只要把蔗糖加入轉化脢後,測量所生成還原力的增高,即可得知此酵素的活性。 他們在進行實驗時,觀察到一件事實: 當酵素的量固定時,若增加基質的量,則反應速率會跟著上升;但此種上升並非沒有限制,當基質增加到某一濃度後,反應速率即不會再增加了,反應速率似乎是受限於其中的酵素分子數量。因此,確定酵素可能要先與基質結合,才能進行催化反應。 HOCH2 O O Juang RH (2004) BCbasics
 作為研究對象,其催化反應如上圖所示,可以把非還原糖的蔗糖,轉化成為兩個具有還原力的單糖 (葡萄糖及果糖)。因此做實驗時,只要把蔗糖加入轉化脢後,測量所生成還原力的增高,即可得知此酵素的活性。 他們在進行實驗時,觀察到一件事實: 當酵素的量固定時,若增加基質的量,則反應速率會跟著上升;但此種上升並非沒有限制,當基質增加到某一濃度後,反應速率即不會再增加了,反應速率似乎是受限於其中的酵素分子數量。因此,確定酵素可能要先與基質結合,才能進行催化反應。 HOCH2. O. O. Juang RH (2004) BCbasics.")
22
NMR spectrometry deuterated solvents are necessary can be done quantitatively Titration Example: ester hydrolysis Ester + water acid + alcohol Excellent for quantitative measurements; hooked up to computer. ml base added vs time; slope = reaction rate

23
Catálisis: Leyes Generales de la Catálisis
Catalizadores biológicos Velocidades más elevadas de reacción Condiciones de reacción mas suaves Mayor especificidad de reacción Capacidad para la regulación

25
Tipos de reacciones Acido-base Oxidación-reducción
Radicales libres: adición y abstracción Condensación Adición a dobles enlaces Sustitución nucleofílica

26
Reacciones de oxidación y reducción

28
Coenzymes and Prosthetic Groups
some enzymes employ coenzymes and prosthetic groups at their active sites used for reactions that amino acid R groups can’t perform coenzymes metals or small organic molecules not covalently bound to protein often function as co-substrates precursors are often vitamins prosthetic groups small organic molecules covalently linked to protein Enzymes and Enzyme Kinetics

30
Rompimiento homolítico de enlaces

31
Rompimiento heterolítico de enlaces

32
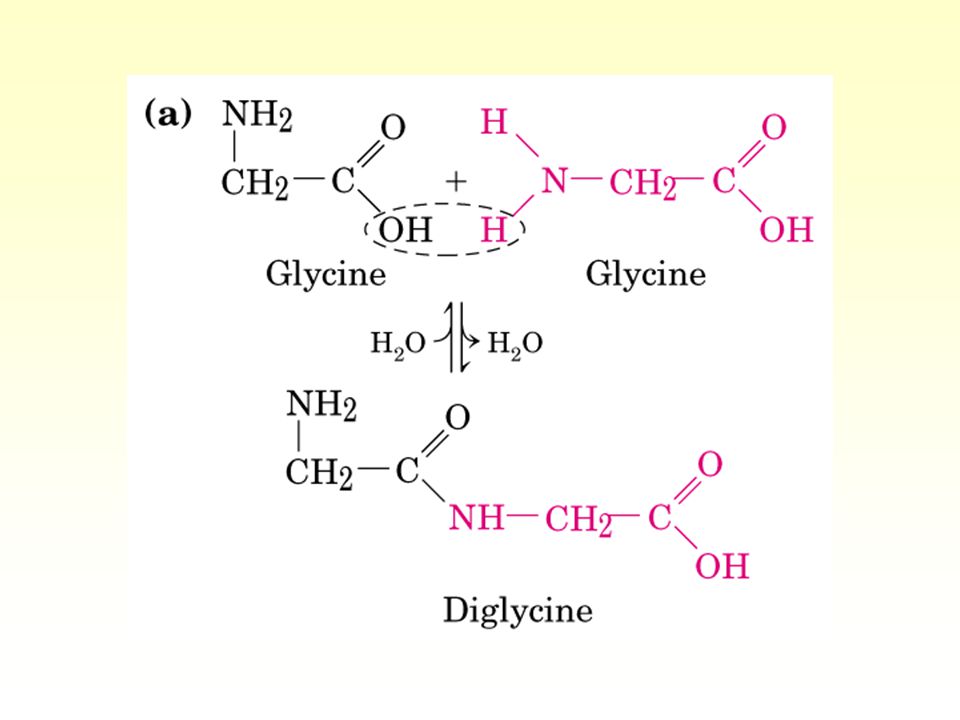
Reacciones de condensación
Condensación: formación de un enlace covalente concomitante con la pérdida de una molécula de agua Ester: ácido carboxílico + alcohol Eter: dos alcoholes (autocondensación) Amida: amina (1a o 2a) + alcohol
 Amida: amina (1a o 2a) + alcohol.")
34
La reacción opuesta a la condensación: la hidrólisis

35
Sustitución nucleofílica

36
Reacción de 1er orden: La velocidad solamente depende de la concentración del sustrato orgánico y no de Z

37
Reacción de 2o orden: La velocidad depende tanto de la concentración del sustrato orgánico como de Z

38
Factores que influyen en la reacción de sustitución
1.-El sustrato -Los compuestos con impedimento estérico siguen el mecanismo SN1 -Los carbonos terciarios, alílicos o bencílicos siguen un mecanismo SN1 ya que pueden formar carbocationes estables R R-C-L CH2=CHCH2L C6H5CH2L -Los carbonos simples siguen un mecanismo SN2 CH3-L R-CH2-L R-CH-L

39
Factores que influyen en la reacción de sustitución
2.- El nucléofilo -Mientras más fuerte sea el nucleófilo, mayor será la velocidad de la reacción SN2 Un nucleófilo con carga negativa es más fuerte que su ácido conjugado HO- > H2O RO-> ROH RS-> RSH La fuerza del nucleófilo es paralela a la basicidad, en un grupo de nucleófilos donde el átomo nucleofílico es el mismo RO- > HO- > ArO- > RCOO- > ROH > H2O Cuando se examina la fuerza relativa de los compuestos que pertenecen al mismo grupo de la tabla periódica, se encuentra que el que tiene el átomo nucleofílico más grande es un nucleófilo más fuerte RSH > ROH RS- > RO- I- > Br- > Cl- > F- -El impedimento estérico que tenga el nucléofilo, importante para la reacción SN2

40
Factores que influyen en la reacción de sustitución
3.- El grupo saliente -Los mejores grupos salientes (L) son aquellos que forman la molécula o ion más estable una vez que se separa. Mientras menos básico sea el grupo más fácilmente se separa. Por ejemplo, el agua es mejor grupo saliente que el ion hidróxido; los alcoholes son mejores grupos salientes que los alcóxidos; los iones haluro, sulfonato, sulfato y carboxilato son buenos grupos salientes. R-OH + X- = R-X + HO- R-OH2+ + X- = R-X + H2O X-: ion haluro En general, son buenos grupos salientes los que tienen átomos electronegativos y que pueden estabilizar electrones a su alrededor
 son aquellos que forman la molécula o. ion más estable una vez que se separa. Mientras menos básico sea el grupo. más fácilmente se separa. Por ejemplo, el agua es mejor grupo saliente que el ion hidróxido; los alcoholes. son mejores grupos salientes que los alcóxidos; los iones haluro, sulfonato, sulfato y carboxilato son buenos grupos salientes. R-OH + X- = R-X + HO- R-OH2+ + X- = R-X + H2O. X-: ion haluro. En general, son buenos grupos salientes los que tienen átomos electronegativos y. que pueden estabilizar electrones a su alrededor.")
42
Example: hydrolysis of alkyl bromides
R-Br + OH– R-OH + Br– (SN2 reaction) Reaction takes place by collision of the particles RBr and OH–, implying that the reaction rate is directly proportional to [RBr] and [OH–]. Alternative: These are the most common reaction situations: - bimolecular: 2 particles collide and react - monomolecular: 1 particle dissociates or reacts E.g., the reaction below runs through a large number of bi-and monomolecular steps: MnO4– + 5 Fe H+ Mn Fe H2O
 Reaction takes place by collision of the particles RBr and OH–, implying that the reaction rate is directly proportional to [RBr] and [OH–]. Alternative: These are the most common reaction situations: - bimolecular: 2 particles collide and react. - monomolecular: 1 particle dissociates or reacts. E.g., the reaction below runs through a large number of bi-and monomolecular steps: MnO4– + 5 Fe H+ Mn Fe H2O.")
43
Reacciones de sustitución nucleofílica Nu: + R-L = Nu-R + :L
Nucleófilo Sustrato Gpo. Saliente Producto Síntesis de HO- R-L (-L) R-OH Alcoholes H2O R-L (-HL) R-OH Alcoholes RO- R-L (-L) R-O-R Eteres R-OH R-L (-HL) R-O-R Eteres HS- R-L (-L) R-SH Tioles NH3 R-L (-HL) R-NH2 Amina 1a RNH2 R-L (-HL) R-NH-R Amina 2a R2NH R-L (-HL) R3N Amina 3a N3- R-L (-L) R-N3 Azida de alquilo I- R-L (-L) R-I Yoduro de alquilo RCOO- R-L (-L) RCOOR Ester RCOOH R-L (-HL) RCOOR Ester CN- R-L (-L) R-CN Nitrilo
 R-OH Alcoholes. H2O R-L (-HL) R-OH Alcoholes. RO- R-L (-L) R-O-R Eteres. R-OH R-L (-HL) R-O-R Eteres. HS- R-L (-L) R-SH Tioles. NH3 R-L (-HL) R-NH2 Amina 1a. RNH2 R-L (-HL) R-NH-R Amina 2a. R2NH R-L (-HL) R3N Amina 3a. N3- R-L (-L) R-N3 Azida de alquilo. I- R-L (-L) R-I Yoduro de alquilo. RCOO- R-L (-L) RCOOR Ester. RCOOH R-L (-HL) RCOOR Ester. CN- R-L (-L) R-CN Nitrilo.")
45
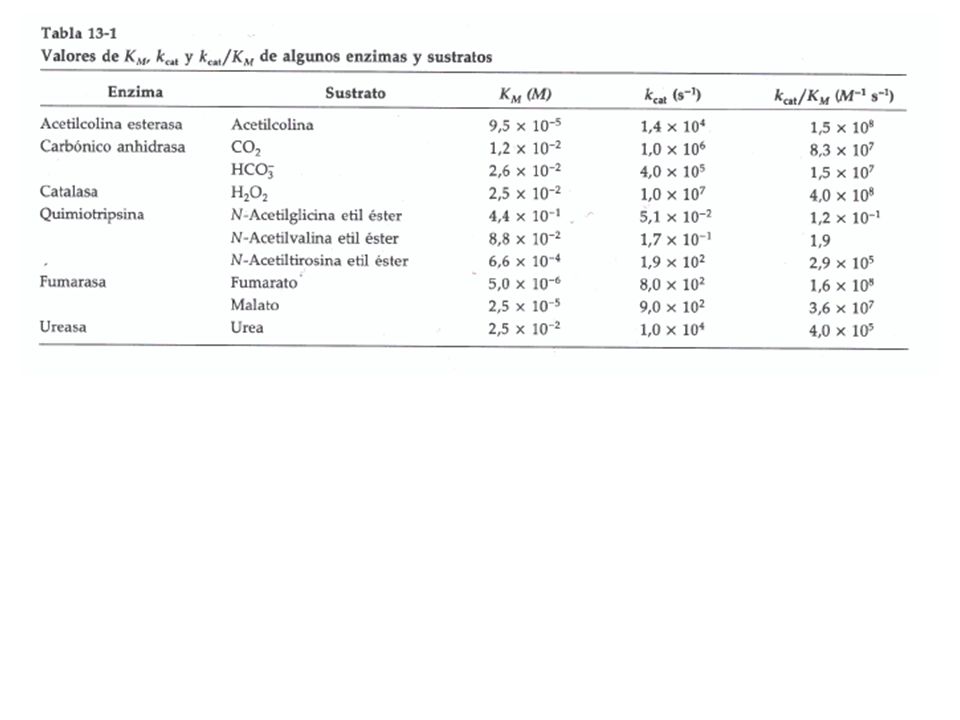
Turn Over Numbers of Enzymes
Substrate kcat (s-1) Catalase H2O2 40,000,000 Carbonic anhydrase HCO3- 400,000 Acetylcholinesterase Acetylcholine 140,000 b-Lactamase Benzylpenicillin 2,000 Fumarase Fumarate 800 常數 k3 控制的是生成物的產生速率,因此也可看作一個酵素最後轉換出生成物的速率,特稱之為 turn over number (t.o.n),也是一個重要指標。 注意其單位為 s-1,亦即每秒鐘能夠轉換得到生成物的分子數目;例如上面 RecA 的 t.o.n. 為 0.4,表示此酵素要 2.5 秒才能使用一分子 ATP 生成 ADP (DNA 重組時需要此酵素)。 RecA protein (ATPase) ATP 0.4 The number of product transformed from substrate by one enzyme molecule in one second Adapted from Nelson & Cox (2000) Lehninger Principles of Biochemistry (3e) p.263
 Catalase H2O2. 40,000,000. Carbonic anhydrase HCO3- 400,000. Acetylcholinesterase Acetylcholine. 140,000. b-Lactamase Benzylpenicillin. 2,000. Fumarase Fumarate 常數 k3 控制的是生成物的產生速率,因此也可看作一個酵素最後轉換出生成物的速率,特稱之為 turn over number (t.o.n),也是一個重要指標。 注意其單位為 s-1,亦即每秒鐘能夠轉換得到生成物的分子數目;例如上面 RecA 的 t.o.n. 為 0.4,表示此酵素要 2.5 秒才能使用一分子 ATP 生成 ADP (DNA 重組時需要此酵素)。 RecA protein (ATPase) ATP The number of product transformed from substrate. by one enzyme molecule in one second. Adapted from Nelson & Cox (2000) Lehninger Principles of Biochemistry (3e) p.263.")
46
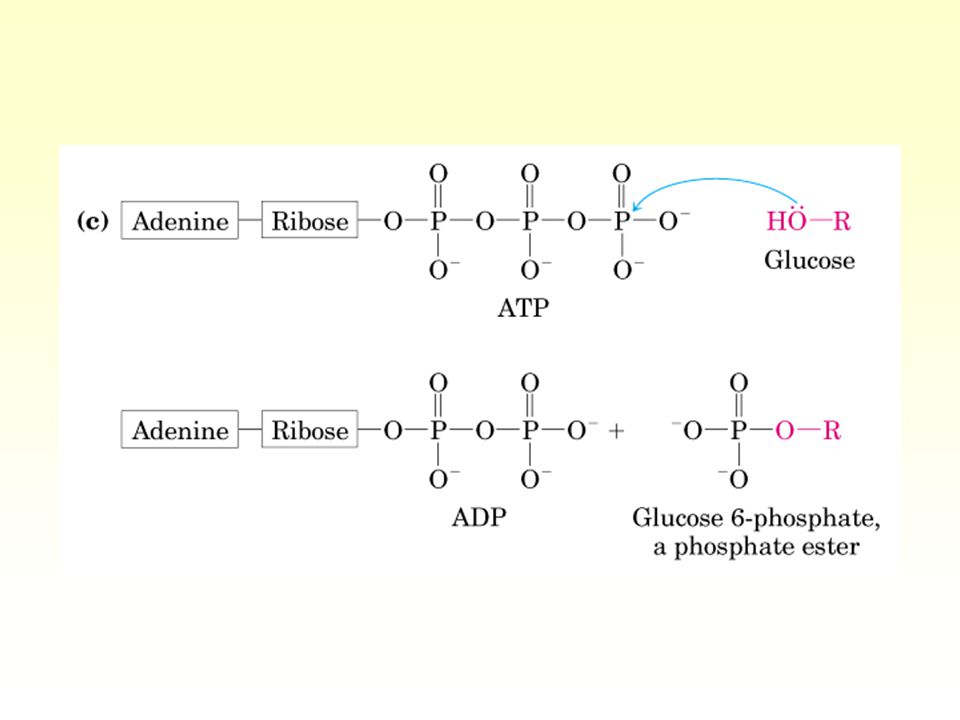
Km = 8 8,000 5 mM Glucose + ATP → Glc-6-P + ADP Km: Hexokinase Example
Allose Mannose Substrate number CHO H-C-OH HO-C-H H2-C-OH CHO H-C-OH H2-C-OH CHO HO-C-H H-C-OH H2-C-OH 1 2 3 4 5 6 Hexokinase 可以催化數種六碳糖 (如 glucose, allose, mannose 等),但三者的 Km 相差甚大。 Hexokinase 對 glucose 及 mannose 有較大的親和力 (因 Km 低),而對 allose 親和力很差 (Km 為 8,000 mM)。 檢查比對這三種單糖的分子構造,發現三號碳上面 -OH 基的立體構形有很大的影響;hexokinase 偏好類似 glucose 的構形。 由此結果反推回去想像 hexokinase 的構造,可以推測 hexokinase 與基質結合的活性區,一定有相當專一的空間排列,其空間排列形狀可以與葡萄糖的形狀互補;而且對葡萄糖分子上的三號碳特別挑剔,但對二號碳則較為寬容。 Km = , mM Juang RH (2004) BCbasics
,但三者的 Km 相差甚大。 Hexokinase 對 glucose 及 mannose 有較大的親和力 (因 Km 低),而對 allose 親和力很差 (Km 為 8,000 mM)。 檢查比對這三種單糖的分子構造,發現三號碳上面 -OH 基的立體構形有很大的影響;hexokinase 偏好類似 glucose 的構形。 由此結果反推回去想像 hexokinase 的構造,可以推測 hexokinase 與基質結合的活性區,一定有相當專一的空間排列,其空間排列形狀可以與葡萄糖的形狀互補;而且對葡萄糖分子上的三號碳特別挑剔,但對二號碳則較為寬容。 Km = 8 8,000 5 mM. Juang RH (2004) BCbasics.")
47
Chymotrypsin Has Distinct kcat / Km to Different Substrates
O R O H3C–C–N–C–C–O–CH3 H H = – kcat / Km R = Glycine –H 1.3 ╳ 10-1 –CH2–CH2–CH3 Norvaline 3.6 ╳ 102 –CH2–CH2–CH2–CH3 Norleucine 3.0 ╳ 103 用 kcat/Km 可以同時兼顧前半與後半反應,也就是說同時監視酵素對基質的親和力,以及該酵素的 Vmax (kcat),是一個比較理想的酵素行為指標; 此一比值越大者,有越好的催化力。由上例可以看出,酵素對不同基質會有不同的表現,chymotrypsin 顯然偏好較大的胺基酸基團,且最好有芳香基團。 酵素的成功催化首先需與基質碰撞,而兩者的碰撞率決定在細胞內的擴散率,但是細胞內的擴散率有其極限;有人由此算出若在最高的擴散速率下,且每次碰撞都完成催化,則一個酵素最高的催化極限,其 kcat/Km 將在 10 8~10 9 (M-1s-1) 之間;的確最有效的酵素催化也都趨近此一範圍,目前最高的記錄是 triose phosphate isomerase 的 2.4×10 8。 –CH2– Phenylalanine 1.0 ╳ 105 (M-1 s-1) Adapted from Mathews et al (2000) Biochemistry (3e) p.379
,是一個比較理想的酵素行為指標; 此一比值越大者,有越好的催化力。由上例可以看出,酵素對不同基質會有不同的表現,chymotrypsin 顯然偏好較大的胺基酸基團,且最好有芳香基團。 酵素的成功催化首先需與基質碰撞,而兩者的碰撞率決定在細胞內的擴散率,但是細胞內的擴散率有其極限;有人由此算出若在最高的擴散速率下,且每次碰撞都完成催化,則一個酵素最高的催化極限,其 kcat/Km 將在 10 8~10 9 (M-1s-1) 之間;的確最有效的酵素催化也都趨近此一範圍,目前最高的記錄是 triose phosphate isomerase 的 2.4×10 8。 –CH2– Phenylalanine. 1.0 ╳ 105. (M-1 s-1) Adapted from Mathews et al (2000) Biochemistry (3e) p.379.")
48
Lipases catalyse the hydrolysis of triglycerides:
The active site of lipases contain 3 essential amino acids: asp, his and ser. Lipases have ~250 other amino acid residues. Why? Tripeptide asp-his-ser enhances the hydrolysis rate of triglycerides by a factor 2-10; lipases by a factor How do they do that?

49
Why are lipases such active enzymes?
Exact alignment of the asp, his and ser side chains Stabilisation of the intermediate C-O Ensure the correct asp, his and ser Stabilise against proteolysis and denaturation

50
E5-10 Charge Relay in Active Site - O–CH2 O C–O - = Ser H–N N H–O–CH2
195 H–O–CH2 Asp 102 His 57 Ser 195 His 57 Asp 102 - O–CH2 O C–O–H = N N–H C H CH2 Active Ser Adapted from Alberts et al (2002) Molecular Biology of the Cell (4e) p.158 Chymotrypsin 活性區上面有三個極為重要的胺基酸,雖然原來在蛋白質長鏈上並沒連在一起 (57, 102, 195),但經蛋白質摺疊後,卻湊在一起成為一直線。然後發生一件非常重要的事,就是 Asp 102 搶走中間 His 57 的氫,然後 His 57 去搶 Ser 195 的氫,最後 Ser 195 留下一個光禿禿的氧原子,而此氧原子因失去氫而變得極為活躍 (因為有多餘的負電荷);這就是著名的催化鐵三角 (catalytic triad) 的形成。 E5-10
 Molecular Biology of the Cell (4e) p.158. Chymotrypsin 活性區上面有三個極為重要的胺基酸,雖然原來在蛋白質長鏈上並沒連在一起 (57, 102, 195),但經蛋白質摺疊後,卻湊在一起成為一直線。然後發生一件非常重要的事,就是 Asp 102 搶走中間 His 57 的氫,然後 His 57 去搶 Ser 195 的氫,最後 Ser 195 留下一個光禿禿的氧原子,而此氧原子因失去氫而變得極為活躍 (因為有多餘的負電荷);這就是著名的催化鐵三角 (catalytic triad) 的形成。 E5-10.")
51
Imidazole on Histidine Is Affected by pH
H–N N–H C H H–N N C H + H+ Adapted from Dressler & Potter (2000) Discovering Enzymes, p.163 + Ser 195 His 57 Asp 102 H–O–CH2 O C–O - = H–N N–H C C-H CH2 H Inactive 上述 酸鹼度的實驗中,當 pH 由 7 降到 6 時 chymotrypsin 的活性急遽下降,是否有什麼胺基酸在這個改變的 pH 範圍中,產生很大的改變? Chymotrypsin 是由胺基酸所組成,而其性質當然是由所組成胺基酸來決定。 回顧一下各種胺基酸的 pKa,在中性 pH 附近的只有 His 的 imidazole (6.0),因此是否因為 pH 的改變影響 His 的帶電性質,因而造成活性的下降? 的確,若把 pH 調到 6 以下,則催化鐵三角中的 His 將會多帶一個質子,而此質子將會加在吸引 Ser 195 質子的那個氮原子上,如此 His 將無法吸走 Ser 195 的質子,後者也就無法成為活性基團,只是個普通的醇基。 Adapted from Alberts et al (2002) Molecular Biology of the Cell (4e) p.158 E5-13
 Discovering Enzymes, p Ser His 57. Asp 102. H–O–CH2. O. C–O - = H–N. N–H. C. C-H. CH2. H. Inactive. 上述 酸鹼度的實驗中,當 pH 由 7 降到 6 時 chymotrypsin 的活性急遽下降,是否有什麼胺基酸在這個改變的 pH 範圍中,產生很大的改變? Chymotrypsin 是由胺基酸所組成,而其性質當然是由所組成胺基酸來決定。 回顧一下各種胺基酸的 pKa,在中性 pH 附近的只有 His 的 imidazole (6.0),因此是否因為 pH 的改變影響 His 的帶電性質,因而造成活性的下降? 的確,若把 pH 調到 6 以下,則催化鐵三角中的 His 將會多帶一個質子,而此質子將會加在吸引 Ser 195 質子的那個氮原子上,如此 His 將無法吸走 Ser 195 的質子,後者也就無法成為活性基團,只是個普通的醇基。 Adapted from Alberts et al (2002) Molecular Biology of the Cell (4e) p.158. E5-13.")
52
pH Influences Chymotrypsin Activity
Relative Activity 早期 對 chymotrypsin 的觀察發現,其酵素活性受到酸鹼度的影響很大,酵素活性在大約中性時最大,往酸鹼兩方向都會降低活性,而且降幅極大。 為何 chymotrypsin 活性會受到 pH 如此大的影響? 可以分成酸鹼兩個方向來探討,因為其機制不一樣。 pH Adapted from Dressler & Potter (1991) Discovering Enzymes, p.162 E5-11
 Discovering Enzymes, p.162. E5-11.")
53
+ - + E5-12 pH Influences Net Charge of Protein 10 9 8 7 6 5 4 3
Buffer pH 3 4 5 6 7 8 9 10 Isoelectric point, pI 通常 一個蛋白質分子上都帶有電荷,有正電荷、也有副電荷,這些正、負電荷的淨值,即為此蛋白質所帶的 淨電荷;蛋白質的淨電荷可能為正、也可能為負,在某 pH 下蛋白質的淨電荷可能為零,則此 pH 稱為此蛋白質的『等電點』(isoelectric point, pI),一個蛋白質的 pI 通常不會改變。 當環境的 pH 大於某蛋白質的的 pI (如上圖某蛋白質的 pI = 6,環境 pH = 9),則此蛋白質的淨電荷為負;反之則為正值。另外,環境的 pH 離其 pI 越遠,則其所帶的淨電荷數目將會越大;越接近 pI 時,所帶淨電荷變小,最後在其 pI 處淨電荷為零。因此,蛋白質溶液的 pH 要很小心選擇,以便使該蛋白質帶有我們所需要的淨電荷,或者不帶有淨電荷。 + - Juang RH (2004) BCbasics + Net Charge of a Protein E5-12
,一個蛋白質的 pI 通常不會改變。 當環境的 pH 大於某蛋白質的的 pI (如上圖某蛋白質的 pI = 6,環境 pH = 9),則此蛋白質的淨電荷為負;反之則為正值。另外,環境的 pH 離其 pI 越遠,則其所帶的淨電荷數目將會越大;越接近 pI 時,所帶淨電荷變小,最後在其 pI 處淨電荷為零。因此,蛋白質溶液的 pH 要很小心選擇,以便使該蛋白質帶有我們所需要的淨電荷,或者不帶有淨電荷。 + - Juang RH (2004) BCbasics. + Net Charge of a Protein. E5-12.")
54
Ejemplo de Catálisis, una Serina proteasa

55
Plegamiento: Proporciona un “andamio” que permite a los aminoácidos catalíticos estar en la posición espacial adecuada para llevar acabo su función, además, genera un medio ambiente adecuado para la captura de los sustratos.

56
La triada catalítica SER-HIS-ASP
Serin-proteasa de Bacillus lentus

57
X E5-16 Chymotrypsin Ser195 Inhibited by DIFP Ser 195 Ser 195
Diisopropyl-fluorophosphate (DIFP) X O (CH3)2CH–O– P –O–CH(CH3)2 F = O (CH3)2CH–O– P –O–CH(CH3)2 = CH2 Ser 195 O-…H CH2 Ser 195 DIFP 與 Ser 型的蛋白脢有相當專一的作用,會與其上的 Ser 195 反應,然後形成固定的鍵結,使得該酵素失去活性。 因此,DIFP 是 chymotrypsin 的專一性抑制劑,也是其假基質 (pseudosubstrate)。 Adapted from Dressler & Potter (1991) Discovering Enzymes, p.167 E5-16
 X. O. (CH3)2CH–O– P –O–CH(CH3)2. F. = O. (CH3)2CH–O– P –O–CH(CH3)2. = CH2. Ser 195. O-…H. CH2. Ser 195. DIFP 與 Ser 型的蛋白脢有相當專一的作用,會與其上的 Ser 195 反應,然後形成固定的鍵結,使得該酵素失去活性。 因此,DIFP 是 chymotrypsin 的專一性抑制劑,也是其假基質 (pseudosubstrate)。 Adapted from Dressler & Potter (1991) Discovering Enzymes, p.167. E5-16.")
58
Chymotrypsin Also Catalyzes Acetate
Hartley & Kilby Nitrophenol acetate O CH3–C–O– –NO2 O -C N- H + H2O Chymotrypsin Peptide bond HO– –NO2 O -C O- Acetate 酵素 雖然有專一性,但是還是對分子構造相似的基質,有時仍然無法辨認;例如水解胜肽鍵的蛋白脢有時也會水解類似的酯鍵,因此 chymotrypsin 可以使用 nitrophenol acetate 為基質,切出黃色的 nitrophenol,直接觀察黃色的深淺即可得知酵素活性。 但是到底 nitrophenol acetate 不是正常的基質,因此這種反應比較慢,慢到可以觀察到每一步反應過程,反而有利於探討酵素的催化機制。 同時,有一個奇怪的現象發生,理論上 nitrophenol acetate 水解後應該得到 nitrophenol 及 acetate,但是在反應的初期,都無法偵測得到 acetate,過一陣子後,醋酸才會跑出來。 那些醋酸根會跑到哪裡去了? 有一個可能就是,醋酸會不會在反應過程中,留在酵素上面,後來才被切出來? 科學家 (Hartley & Kilby) 的確是利用這樣的方法,證明 chymotrypsin 有兩階段的催化過程。 O CH3–C–OH Nitrophenol Ester bond No acetate was detected at early stage Adapted from Dressler & Potter (1991) Discovering Enzymes, p.168 E5-20
 的確是利用這樣的方法,證明 chymotrypsin 有兩階段的催化過程。 O. CH3–C–OH. Nitrophenol. Ester bond. No acetate was detected at early stage. Adapted from Dressler & Potter (1991) Discovering Enzymes, p.168. E5-20.")
59
Two-Stage Catalysis of Chymotrypsin
Acylation O CH3–C–O– –NO2 Nitrophenol acetate O C CH3–C HO– –NO2 O - C Kinetics of reaction Deacylation (slow step) CH3COOH + H2O Time (sec) Nitrophenol O-H C 現在 已經知道 chymotrypsin 的催化機制分成 acylation 及 deacylation 兩個階段,其研究是利用 nitrophenol acetate 為基質所進行的;在早期的觀察中,發現催化生成 nitrophenol 的速率有兩個 phases,起先相當快放出 nitrophenol,然後變慢並一直維持此一速度。 利用兩階段式催化機制,可說明為何開始速率會比較快。 這是因為第一階段的 acylation 反應很快,而剛開始酵素都空閒著,一抓到基質就水解並放出 nitrophenol,因此黃色產物迅速上升。但是因為醋酸根還留在酵素上,準備進行第二步的 deacylation,而此一步驟相當慢速,即為 速率決定步驟 rate-limiting step;酵素要在加入水分子,釋出醋酸後,才能繼續吸入 nitrophenol acetate 繼續下一輪反應。因此出現了兩相的反應速率。 Two-phase reaction Adapted from Dressler & Potter (1991) Discovering Enzymes, p.169 E5-21
 CH3COOH. + H2O. Time (sec) Nitrophenol. O-H. C. 現在 已經知道 chymotrypsin 的催化機制分成 acylation 及 deacylation 兩個階段,其研究是利用 nitrophenol acetate 為基質所進行的;在早期的觀察中,發現催化生成 nitrophenol 的速率有兩個 phases,起先相當快放出 nitrophenol,然後變慢並一直維持此一速度。 利用兩階段式催化機制,可說明為何開始速率會比較快。 這是因為第一階段的 acylation 反應很快,而剛開始酵素都空閒著,一抓到基質就水解並放出 nitrophenol,因此黃色產物迅速上升。但是因為醋酸根還留在酵素上,準備進行第二步的 deacylation,而此一步驟相當慢速,即為 速率決定步驟 rate-limiting step;酵素要在加入水分子,釋出醋酸後,才能繼續吸入 nitrophenol acetate 繼續下一輪反應。因此出現了兩相的反應速率。 Two-phase. reaction. Adapted from Dressler & Potter (1991) Discovering Enzymes, p.169. E5-21.")
60
Specificity of Ser-Protease Family
Trypsin Chymotrypsin Elastase cut at Lys, Arg cut at Trp, Phe, Tyr cut at Ala, Gly O O –C–N–C–C–N– C NH3 + O O –C–N–C–C–N– C O O –C–N–C–C–N– CH3 Shallow and non-polar pocket Deep and negatively charged pocket COO- C Asp Non-polar pocket Juang RH (2004) BCbasics Active Site
 BCbasics. Active Site.")
61
Modification of Subtilisin and Its Activity Change
Active Site Relative Modification Triad: Ser His Asp activity No enzyme Subtilisin ● ● ● ,000,000,000 Ser, His & Asp → Ala ○ ○ ○ 4,000 His & Asp → Ala ● ○ ○ 37,000 Ser → Ala ○ ● ● 5,000 Asp → Ala ● ● ○ ,000 為了 研究 Ser protease 的催化區,有人把 subtilisin 催化鐵三角上面的胺基酸修改,再看影響修飾後的蛋白脢活性大小。結果發現 Ser195 是絕對不可以改變的重要胺基酸,His57 及 Asp102 次之。同時,若把可以穩定過渡狀態分子的 Asn155 改成不具電荷的 Leu,活性也會下降一千倍。因此,這三個重要胺基酸是不能任意改變,所有 Ser protease 家族成員,都有這三個胺基酸在固定的位置。 Asn155 → Leu ● ● ● 10,000,000 (Asn155 stabilizes transition state) Adapted from Dressler & Potter (1991) Discovering Enzymes, p.245
 Adapted from Dressler & Potter (1991) Discovering Enzymes, p.245.")
62
Serine Protease and AchE
Chymotrypsin – Gly – Asp – Ser – Gly – Gly – Pro – Leu – Trypsin – Gly – Asp – Ser – Gly – Gly – Pro – Val – Elastase – Gly – Asp – Ser – Gly – Gly – Pro – Leu – Thrombin – Gly – Asp – Ser – Gly – Gly – Pro – Phe – Plasmin – Gly – Asp – Ser – Gly – Gly – Pro – Leu – Acetylcholinesterase – Gly – Glu – Ser – Ala – Gly – Gly – Ala – Ser 195 Chymotrypsin – Val – Thr – Ala – Ala – His – Cys – Gly – Trypsin – Val – Ser – Ala – Gly – His – Cys – Tyr – Elastase – Leu – Thr – Ala – Ala – His – Cys – Ile – Thrombin – Leu – Thr – Ala – Ala – His – Cys – Leu – Plasmin – Leu – Thr – Ala – Ala – His – Cys – Leu – Acetylcholinesterase – – – – – – – – – – – – – His – – – – – – – – His 57 Chymotrypsin – Thr – Ile – Asn – Asn – Asp – Ile – Thr – Trypsin – Tyr – Leu – Asn – Asn – Asp – Ile – Met – Elastase – Ser – Lys – Gly – Asn – Asp – Ile – Ala – Thrombin – Asn – Leu – Asp – Arg – Asp – Ile – Ala – Plasmin – Phe – Thr – Arg – Lys – Asp – Ile – Ala – Acetylcholinesterase – – – – – – – – – – – – – – Asp – – – – – – – 把 Ser 家族幾個成員拿出來比較,看催化鐵三角附近胺基酸序列的差異性如何。最重要的 Ser 附近的胺基酸保守性非常高,其它地方也都是半保守性地取代 (即極性取代極性、非極性取代非極性)。 成員中有一個 acetylcholinesterase (AchE) 似乎是認養來的,AchE 也有催化鐵三角,但是整體的胺基酸序列並不太像 (因此上圖無法列出比較的序列),好像沒有『血緣』關係。的確如此,AchE 原來並非 Ser 家族的嫡系成員,只是趨同演化出具有與鐵三角類似的催化機制。 Asp 102 Adapted from Dressler & Potter (1991) Discovering Enzymes, p.244
。 成員中有一個 acetylcholinesterase (AchE) 似乎是認養來的,AchE 也有催化鐵三角,但是整體的胺基酸序列並不太像 (因此上圖無法列出比較的序列),好像沒有『血緣』關係。的確如此,AchE 原來並非 Ser 家族的嫡系成員,只是趨同演化出具有與鐵三角類似的催化機制。 Asp 102. Adapted from Dressler & Potter (1991) Discovering Enzymes, p.244.")
63
AchE Has Similar Catalytic Mechanism
O CH3 CH3–C–O–CH2–CH2–N–CH3 CH3 + AchE O - C H O CH3–C CH3 O–CH2–CH2–N–CH3 H CH3 + O - C H H AchE H2O Acylation↑ CH3 HO–CH2–CH2–N–CH3 + O CH3–C–OH AchE O C H CH3–C ↓Deacylation H-O-H Acetylcholinesterase 的催化機制與胜肽鍵水解極相似,活性區有高反應性的 Ser 以及 His 和 Asp 的質子接力,也分成 acylation 及 deacylation 兩個階段。 AchE O - C H Adapted from Dressler & Potter (1991) Discovering Enzymes, p.243
 Discovering Enzymes, p.243.")
64
La posición espacial relativa de los elementos catalíticos se mantiene aunque la secuencia de aminoácidos cambie.

65
TIM barrels Cholinesterase Haloalkane dehalogenase
Dienelactone hidrolase Lipase/Cutinase Thioestearase Serine Carboxypeptidase Prolyl Imino/Oligo peptidase Bromoperoxidase Haloalkane dehalogenase Fluoroacetate dehalogenase Epoxide hydrolase Hydroxynitrile lyase C-C Hydrolase 2,4-Dioxygenase

66
grupo guanidino de Arg grupos carboxilo de Glu y Asp grupo sulfhidrilo de Cys grupo imidazol de His grupo amino de Lys grupo tioéter de Met grupo indoílo de Trp grupo hidroxilo fenólico de Tyr pKa 12-13 6.5-7 9.6-10

67
Características funcionales de barriles TIM
Sitio activo en la terminal carboxilo de las hebras b Hélices orientadas paralelamente forman un dipolo Sitio activo: Atractor natural de cargas negativas Sustratos naturales fosforilados en la mayoría de los casos

68
Flexibilidad: La “tapa” del sitio activo de algunas enzimas presenta mobilidad.
Sitios Alostéricos: Activan a la proteína desde regiones alejados del sitio activo. Cambios conformacionales: Suceden al unir un sustrato. Glucocinasa Glucosamina 6-Fosfato-desaminasa

69
- Aminoácidos catalíticos: Son los involucrados en la función.
Están directamente involucrados en la catálisis. Ejercen un efecto en otro residuo o molécula de agua que está directamente relacionado con la catálisis. Estabilizan el estado de transición. Ejercen algún efecto en el sustrato o cofactor que ayuda a la catálisis. - Aminoácidos del sitio activo no catalíticos: Participan en el reconocimiento del sustrato y lo colocan en la orientación correcta.

70
Sitio Activo de la Tiamina Fosfato Sintasa
Aminoácidos del sitio activo no catalíticos que estabilizan al Pirofosfato. Aminoácidos catalíticos Arg59, Ser130, Lys 159 Mg++ Aminoácidos del sitio activo no catalíticos que estabilizan a los sustratos.

71
Analysis of Catalytic Residues in Enzyme Active Sites
JMB (2002) 324,
 324,")
72
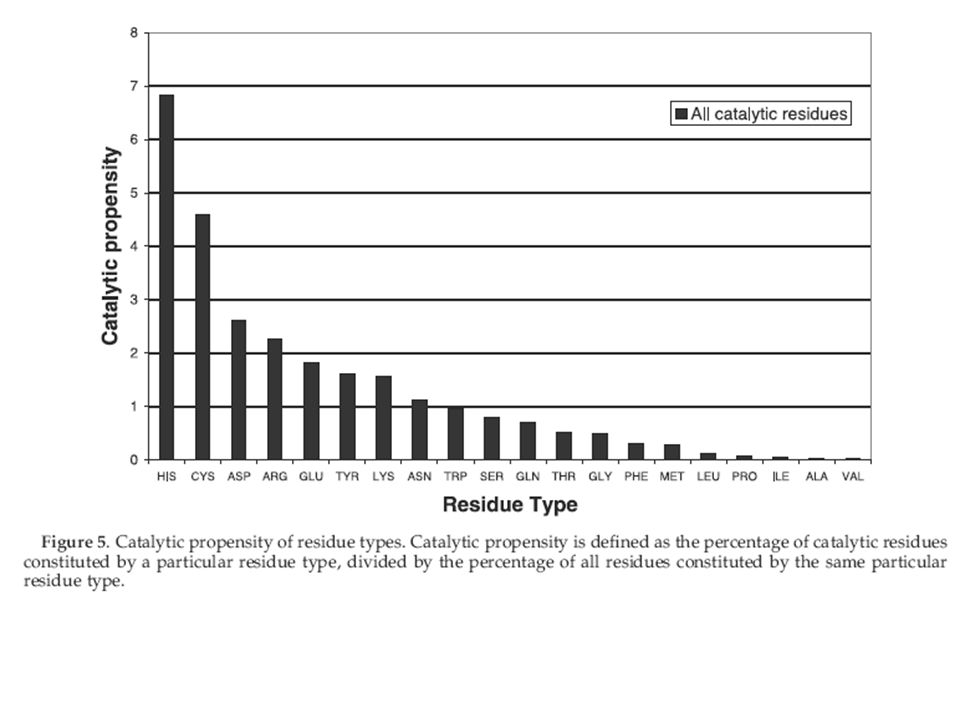
His, Asp y Glu casi siempre son catalíticos

75
- Están muy poco expuestos al solvente, a pesar de su polaridad.
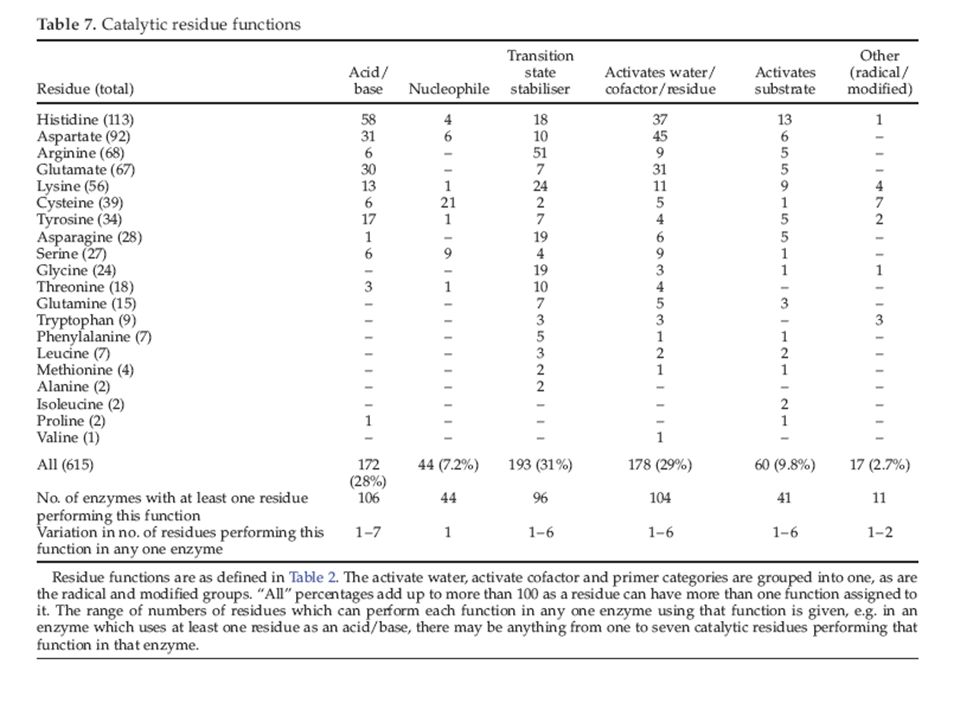
Conclusiones: - Los aminoácidos mayoritariamente catalíticos son histidina, cisteina, ácido glutámico, ácido aspártico, arginina, lisina. - Están muy poco expuestos al solvente, a pesar de su polaridad. - Casi todos están formando puentes de hidrógeno. - Tienen una movilidad muy baja en comparación con los demás aminoácidos. - La función de un aminoácido catalítico es estabilizar un intermediario del estado de transición, actuar como nucleófilo o ser donador o aceptor de protones.

Presentaciones similares

















. It features two verb changes that we will see very soon.>")











