Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Esteban Osorio Cadavid, PhD. Curso Biología Celular y Bioquímica I
ENZIMAS Esteban Osorio Cadavid, PhD. Curso Biología Celular y Bioquímica I

2
ENZIMAS Proteínas especificas que catalizan las reacciones químicas en los sistemas biológicos.

3
Las enzimas aceleran las reacciones multiplicando su velocidad por un millón de veces, o aún más.
CO2 + H2O H2CO3 Cada molécula enzimática puede hidratar 105 moléculas de CO2 en un segundo. Esta reacción catalizada es 107 veces más rápida que la misma reacción no catalizada. Anhidrasa carbónica

4
Las enzimas son altamente Específicas tanto en la reacción que catalizan como en la selección de las sustancias reaccionantes, denominadas SUSTRATOS. El grado de especificidad del Sustrato es normalmente elevado, a veces prácticamente absoluto.

5
Especificidad de la Tripsina: Rompe los enlaces peptídicos sólo por el lado carboxílico de los residuos Lisina y de Arginina.

6
2. Especificidad de la Trombina, un factor de coagulación
2. Especificidad de la Trombina, un factor de coagulación. Rompe el enlace peptídico, cuando el lado carboxilo es la arginina y el lado amino es la glicina

7
3. Actividad enzimática de la Subtilisina: rompe cualquier enlace peptídico, independiente de la secuencia de aminoácidos.

8
4. Enzimas proteolíticas: Catalizan la hidrólisis de un enlace peptídico.

9
La mayoría de las enzimas proteolíticas también catalizan una reacción diferente, pero relacionada: la hidrólisis de un enlace éster.

10
Regulación de la Actividad de Algunas Enzimas
Zimógenos: Son los precursores enzimáticamente inactivos de las enzimas proteolíticas. se sintetizan en forma de un precursor inactivo son activadas a un tiempo y en un lugar fisiológicamente apropiado. El tripsinógeno es activado por la rotura de un enlace peptídico en el intestino delgado para formar tripsina. Se sintetiza en el páncreas.

11
Secreción de Zimógenos en el páncreas

12
Activación de zimógenos por clivaje proteolítico.

13
Figure 6.21 Activation of some pancreatic zymogens. Initially, enteropeptidase catalyzes the activation of trypsinogen to trypsin. Trypsin then activates chymotrypsinogen, proelastase, and additional trypsinogen molecules.

14
Figure 6.22 Polypeptide chains of chymotrypsinogen (blue) [PDB 2CGA] and -chymotrypsin (green) [PDB 5CHA]. Ile-16 and Asp-194 in both zymogen and the active enzyme are shown in yellow. The catalytic-site residues (Asp-102, His-57, and Ser-195) are shown in red.
 [PDB 2CGA] and -chymotrypsin (green) [PDB 5CHA]. Ile-16 and Asp-194 in both zymogen and the active enzyme are shown in yellow. The catalytic-site residues (Asp-102, His-57, and Ser-195) are shown in red.")
15
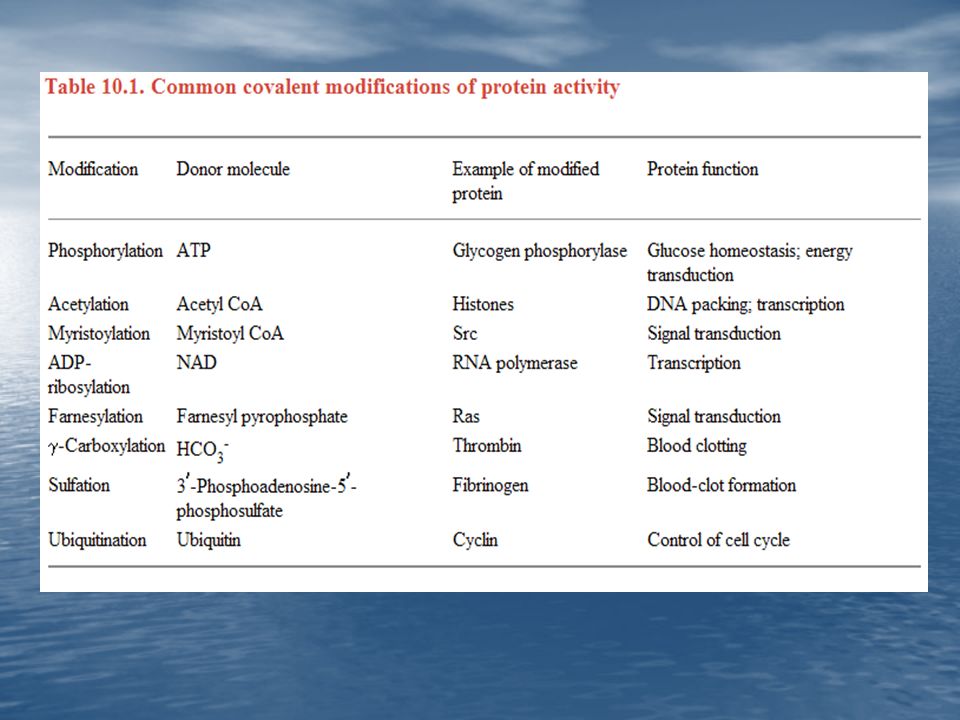
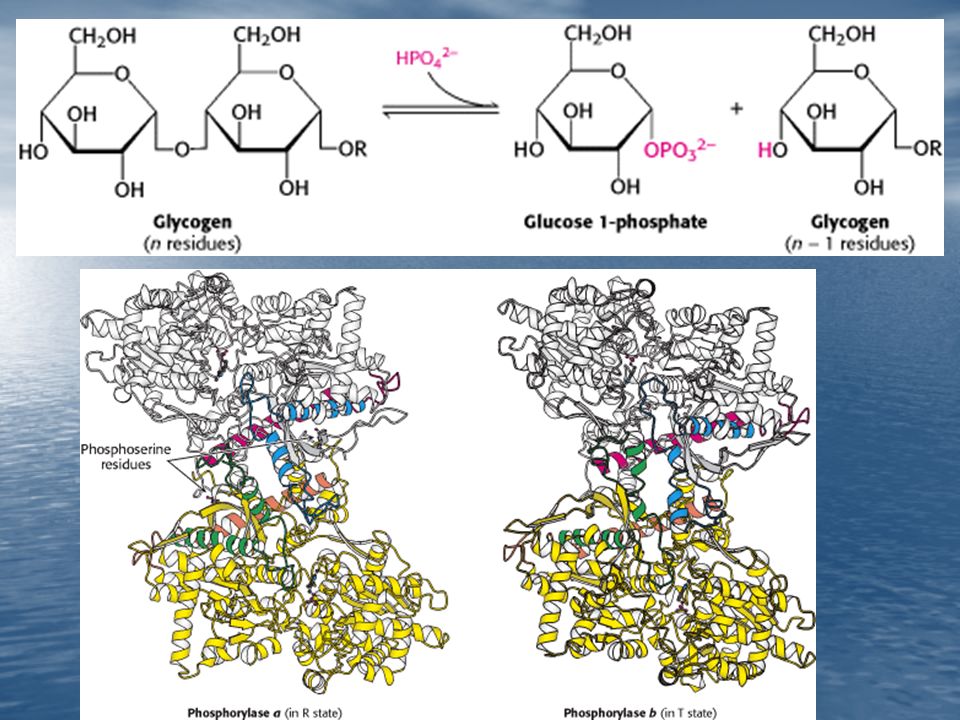
2. Modificación Covalente: Este mecanismo de control enzimático consiste en la inserción de un pequeño grupo sobre la enzima. Las enzimas que degradan el glucógeno están reguladas por la introducción de un grupo fosforilo a un residuo específico de serina de estas enzimas.

18
3. Inhibición por producto final: Mecanismo regulador que afecta a muchas secuencias de reacciones que dan por resultado la síntesis de moléculas pequeñas tales como los aminoácidos. La enzima que cataliza la primera etapa en tal vía biosintética es inhibida por el producto final.

19
Mecanismo de control denominado también “Inhibicion Feedback” o “retroalimentación”.
La treonina se convierte en isoleucina en 5 etapas, la primera es catalizada por treonina desaminasa. Cuando la isoleucina ha alcanzado un nivel alto, esta se une a la enzima en un lugar diferente al catalítico inactivándola.

20
4. Control Fisiológico: La síntesis de lactosa por la glándula mamaria.
La lactosa sintetasa, cataliza la síntesis de lactosa y consta de una subunidad catalítica y una subunidad modificadora. La subunidad modificadora altera la especificidad de la subunidad catalítica de tal manera que así une galactosa a glucosa para formar lactosa.

21
Durante el embarazo, la subunidad catalizadora se forma en la glándula mamaría, pero se forma poco subunidad modificadora. En el momento del nacimiento, se sintetiza la subunidad modificadora en grandes cantidades. La subunidad modificadora se une entonces a la subunidad catalitica para formar el complejo de Lactosa sintetasa activa.

23
La subunidad catalítica sola no puede sintetizar Lactosa, tiene un papel diferente: cataliza la unión de galactosa a las proteínas que contienen una cadena carbohidratada unida covalentemente.

24
Las Enzimas y su Papel en la Transformación de Energía
En muchas reacciones bioquímicas, la energía de las sustancias reactivas se convierten en una forma de energía diferente con una eficiencia muy elevada.

25
En la fotosíntesis, la energía lumínica
En la respiración, la energía libre contenida en los alimentos energía química de enlace. adenosina trifosfato (ATP).
.")
26
3. En la contracción muscular, la energía química del enlace ATP
energía mecánica Estas transformaciones de energía son llevadas a cabo por moléculas enzimáticas que son parte integrante de conjuntos altamente especializados.

27
ATPasa catalizando el transporte de Ca2+ a través de la membrana célular.

28
Las enzimas no alteran los equilibrios de reacción:
Una enzima es un catalizador y como consecuencia no puede alterar el equilibrio de una reacción química. Una enzima acelera la reacción en un sentido u otro precisamente con el mimo factor.

29
La concentración de equilibrio de B es 100 veces la de A, haya o no enzima presente.
Cuando la enzima está presente el equilibrio se obtendría en un segundo. Las enzimas aceleran la consecución del equilibrio, pero no varían su posición.

30
Las enzimas disminuyen la energía de activación de las reacciones catalizadas.

31
= GEstado de transición - G Sustrato
Una reacción química A B transcurre a través de un Estado de Transición que tiene un energía mayor que la de A o la de B. La velocidad de reacción hacia adelante depende de la temperatura y de la diferencia de energía libre de activación de Gibbs y se simboliza como = GEstado de transición - G Sustrato

32
La Formación de un complejo Enzima –Sustrato es el primer paso en la catálisis enzimática. La existencia de los complejos ES ha sido demostrada de varias maneras: Algunos complejos ES se han visualizado directamente por microscopía electrónica y por cristalografía de rayos X.

33
2. Las propiedades físicas de un enzima, como solubilidad o estabilidad al calor, cambian frecuentemente con la formación del complejo ES. 3. Las características espectroscópicas de muchos enzimas y sustratos cambian con la formación del complejo ES.

34
Piridoxal Fosfato: grupo prostético de la enzima triptófano sintetasa.
La intensidad de fluorescencia del piridoxal fosfato en el centro activo de la triptófano sintetasa cambia por la adición de serina e indol, los sustratos. Piridoxal Fosfato: grupo prostético de la enzima triptófano sintetasa. L – serina + Indol L - triptófano

35
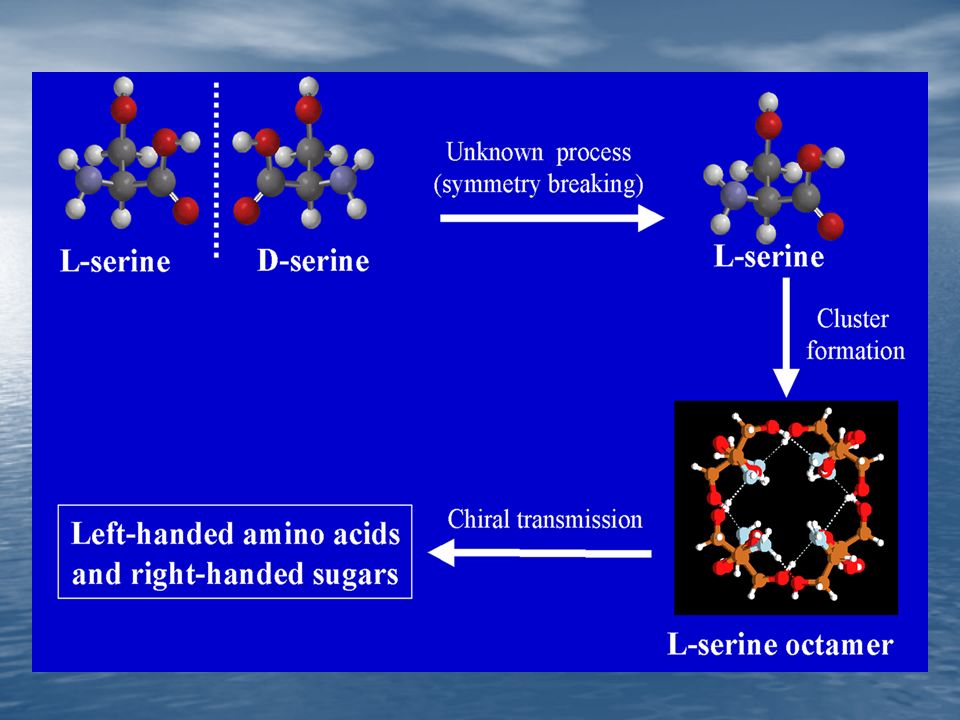
4. En la formación de complejos ES se muestra un alto grado de estero especificidad.
Ejemplo: La D-Serina no es sustrato de la Triptofano Sintetasa.

37
5. Los complejos ES algunas veces pueden se aislados en forma pura
5. Los complejos ES algunas veces pueden se aislados en forma pura. A+B = C Puede aislarse el complejo EA, si la enzima tiene una afinidad alta para A, y B está ausente de la mezcla.

38
A una concentración de enzima constante, la velocidad de reacción aumenta con la concentración de sustrato, hasta que alcanza una velocidad máxima. La primera etapa de la catálisis enzimática es la formación de un complejo enzima-sustrato.

40
La formación y rotura de un enlace químico por un enzima viene precedida por la formación de un complejo Enzima-Sustrato (ES). El sustrato queda ligado a una región específica del enzima llamado Centro activo.

41
A una concentración de enzima constante, la velocidad de reacción aumenta con la concentración del sustrato hasta una velocidad máxima. En 1913, Leonor Michaelis, interpretó la velocidad máxima de una reacción catalizada por un enzima en términos de la formación de un complejo Enzima-Sustrato (ES) directo, a concentraciones de sustrato suficientemente elevadas, los centros catalíticos están ocupados por él, y por lo tanto, la velocidad de reacción alcanza un máximo.
 directo, a concentraciones de sustrato suficientemente elevadas, los centros catalíticos están ocupados por él, y por lo tanto, la velocidad de reacción alcanza un máximo.")
42
CARACTERÍSTICAS DE LOS CENTROS ACTIVOS
El centro activo de una enzima es la región que une al sustrato y contribuye con los residuos que participan directamente en la producción y rotura de enlaces.

43
El centro activo supone una porción relativamente pequeña del volumen total de enzima. Casi todas las enzimas están constituidas por más de 100a.a., una masa mayor a 10kdal y un diámetro mayor de 25Å

44
2. El centro activo es una identidad tridimensional compleja, compuesta de grupos que proceden de diferentes partes de la secuencia lineal de a.a. 3. Los sustratos se unen a las enzimas por fuerzas relativamente débiles.

45
Los sitios activos pueden incluir residuos distantes

46
Puentes de Hidrógeno entre una enzima y un sustrato

47
4. Los centros activos son hoyos o hendiduras que contienen varios residuos polares que son esenciales para la unión o catálisis. 5. La especificidad del enlace depende de la disposición exactamente definida de los átomos del centro activo.

48
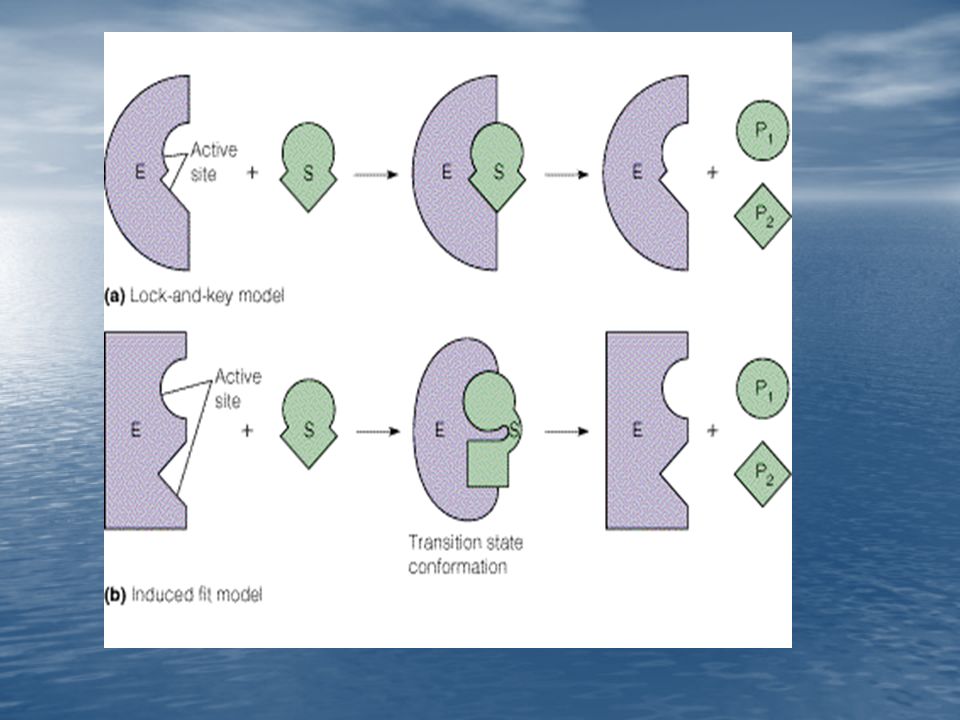
Modelo Llave-Cerradura

49
Modelo del Ajuste Inducido

51
Los centros activos de algunas enzimas no son rígidos
Los centros activos de algunas enzimas no son rígidos. La forma del centro activo viene modificada por su unión al sustrato. El centro activo tiene una forma complementaria a la del sustrato solamente después de que el sustrato queda ligado.

52
Con platino Coloidal (Catalizador inorgánico) 13
Valores de la reacción H2O H2O + ½ O2 a diferentes condiciones experimentales. Condición Kcal.mol-1 Sin catalizadores 18 Con platino Coloidal (Catalizador inorgánico) 13 Con la enzima Catalasa (peroxidasa) 7
 13. Con la enzima Catalasa (peroxidasa) 7.")
53
Clasificación de las enzimas de acuerdo a la comisión de enzimas de la IUB. (Internacional Union of Biochemistry) Grupo Nº Nombre Genérico Reacción-Ejemplo 1 Oxido – reductasas Oxidación y reducción AH2 + B A + BH2 2 Transferasas Transferencia de grupos A–P + B A + B–P 3 Hidrolasas Hidrólisis A – B + H2O A –OH +B-H

54
Formación de nuevos enlaces con la ruptura de ATP
Clasificación de las enzimas de acuerdo a la comisión de enzimas de la IUB. (Internacional Union of Biochemistry) Grupo Nº Nombre Genérico Reacción-Ejemplo 4 Liasas Adición a dobles enlaces 5 Isomerasas Isomerización enantiómero D enantiómero L 6 Ligasas Formación de nuevos enlaces con la ruptura de ATP A + B + ATP A – B + ADP + Pi C + X 2
 Grupo Nº. Nombre Genérico. Reacción-Ejemplo. 4. Liasas. Adición a dobles enlaces. 5. Isomerasas. Isomerización. enantiómero D enantiómero L. 6. Ligasas. Formación de nuevos enlaces con la ruptura de ATP. A + B + ATP. A – B + ADP + Pi. C. + X. 2.")
55
Oxido – reductasas (Reacciones de oxido-reducción)
Clasificación internacional de las enzimas (denominaciones de las clases, números de códigos, tipos de reacciones catalizadas.) Oxido – reductasas (Reacciones de oxido-reducción) 1.1 actúan sobre CH – OH C = O C = C H C H – N H2 C H – N H 1.6 actúan sobre NADH o NADPH
 Oxido – reductasas (Reacciones de oxido-reducción) 1.1 actúan sobre CH – OH. 1.2 C = O. 1.3 C = C H. 1.4 C H – N H C H – N H. 1.6 actúan sobre NADH o NADPH.")
56
Oxido-reductasas

57
2. Transferasas (Transferencia de grupos funcionales)
Clasificación internacional de las enzimas (denominaciones de las clases, números de códigos, tipos de reacciones catalizadas.) 2. Transferasas (Transferencia de grupos funcionales) 2.1 Grupos de un átomo de carbono 2.2 Grupos aldehídicos o cetónicos 2.3 Grupos acila 2.4 Grupos glucosilo 2.5 Grupos fosfato 2.6 Grupos que contienen azufre
 2. Transferasas (Transferencia de grupos funcionales) 2.1 Grupos de un átomo de carbono. 2.2 Grupos aldehídicos o cetónicos. 2.3 Grupos acila. 2.4 Grupos glucosilo. 2.5 Grupos fosfato. 2.6 Grupos que contienen azufre.")
58
Transferasa

59
3. Hidrolasas (Reacciones de hidrólisis) 3.1 ésteres
Clasificación internacional de las enzimas (denominaciones de las clases, números de códigos, tipos de reacciones catalizadas.) 3. Hidrolasas (Reacciones de hidrólisis) 3.1 ésteres 3.2 enlaces glucosídicos 3.3 enlaces peptídicos 3.4 otros enlaces C – N 3.5 anhídridos de ácido
 3. Hidrolasas (Reacciones de hidrólisis) 3.1 ésteres. 3.2 enlaces glucosídicos. 3.3 enlaces peptídicos. 3.4 otros enlaces C – N. 3.5 anhídridos de ácido.")
60
Hidrolasas

61
4. Liasas (adición a los dobles enlaces)
4.1 C = C 4.2 C = O 4.3 C = N 5. Isomerasas (reacciones de isomerización) 5.1 racemasas
 5.1 racemasas.")
62
6. Ligasas (Formación de enlaces con escisión de ATP)
6.2 C – S 6.3 C – N 6.4 C – C

63
KM de algunas enzimas Enzima y sustrato KM (mn) Catalasa H2O2 25
Hexoquinasa Glucosa Fructosa Quimotripsina N – Benzoiltirosinamida N – Formiltirosinamida N - Acetiltirosinamida Gliciltirosinamida Anhidrasa Carbónica Glutamato –deshidrogenasa Glutamato L. Cetoglurarato NH

64
Ejemplos de algunas enzimas que contienen o requieren iones metálicos como cofactores.
alcohol deshidrogenasa arginasa anhidrasa carbónica Fosfotransferasas carboxipeptidasa o Tirosinasa piruvato quinasa Citocromo oxidasa K+

65
Ejemplos de algunas enzimas que contienen o requieren iones metálicos como cofactores.
Fosfohidrolasas citocromos Fosfotransferasas peroxidasas ferredoxina

66
niacina o nicotinamida átomo de hidrógeno (e-)
Ejemplos de algunas coenzimas con indicación de su procedencia vitamínica y los grupos involucrados en la reacción. Coenzima Vitamina precursora Grupos involucrados NAD+ o NADP+ niacina o nicotinamida átomo de hidrógeno (e-) FAD o FMN riboflavina (B2) Pirofosfato de Tiamina Tiamina (B1) aldehídos R C H O
 FAD o FMN. riboflavina (B2) Pirofosfato de Tiamina. Tiamina (B1) aldehídos. R. C. H. O.")
67
Cobalamina o cobalamida
Ejemplos de algunas coenzimas con indicación de su procedencia vitamínica y los grupos involucrados en la reacción. Coenzima Vitamina precursora Grupos involucrados Fosfato de piridoxal piridoxina (B6) amino –NH2 o –NH3 Coenzima A ácido pantoténico acilo Cobalamina o cobalamida cobalamina (B12) alquilo (grupo que contiene átomos de C y H) R C H O
 amino –NH2 o –NH3. Coenzima A. ácido pantoténico. acilo. Cobalamina o cobalamida. cobalamina (B12) alquilo (grupo que contiene átomos de C y H) R. C. H. O.")
68
meril, mutilen, formil, Formimino, etc.
Ejemplos de algunas coenzimas con indicación de su procedencia vitamínica y los grupos involucrados en la reacción. Coenzima Vitamina precursora Grupos involucrados Biotina o biocitina biotina carboxilo Tetrahidrofolato ácido fólico meril, mutilen, formil, Formimino, etc. C O H R = Grupo alquilo que contiene átomos de Carbono e Hidrógeno X = grupo diferente a un alquilo ó Hidrogeno

69
CINÉTICA DE LA CATÁLISIS ENZIMÁTICA

70
Las enzimas aceleran las reacciones por la disminución de la energía libre de activación.

71
Figure 6.3 Enzymatic catalysis of the reaction A + B --> A - B(a) Energy diagram for an uncatalyzed reaction. (b) Effect of reactant binding. Collection of the two reactants in the EAB complex properly positions them for reaction, makes formation of the transition state more frequent, and hence lowers the activation energy. (c) Effect of transition-state stabilization. An enzyme binds the transition state more tightly than it binds substrates, further lowering the activation energy. Thus, an enzymatic reaction has a much lower activation energy than an uncatalyzed reaction. (The breaks in the reaction curves indicate that the enzymes provide multistep pathways.)
 Energy diagram for an uncatalyzed reaction. (b) Effect of reactant binding. Collection of the two reactants in the EAB complex properly positions them for reaction, makes formation of the transition state more frequent, and hence lowers the activation energy. (c) Effect of transition-state stabilization. An enzyme binds the transition state more tightly than it binds substrates, further lowering the activation energy. Thus, an enzymatic reaction has a much lower activation energy than an uncatalyzed reaction. (The breaks in the reaction curves indicate that the enzymes provide multistep pathways.)")
72
Figure 6.3 Enzymatic catalysis of the reaction A + B --> A - B(a) Energy diagram for an uncatalyzed reaction. (b) Effect of reactant binding. Collection of the two reactants in the EAB complex properly positions them for reaction, makes formation of the transition state more frequent, and hence lowers the activation energy. (c) Effect of transition-state stabilization. An enzyme binds the transition state more tightly than it binds substrates, further lowering the activation energy. Thus, an enzymatic reaction has a much lower activation energy than an uncatalyzed reaction. (The breaks in the reaction curves indicate that the enzymes provide multistep pathways.)
 Energy diagram for an uncatalyzed reaction. (b) Effect of reactant binding. Collection of the two reactants in the EAB complex properly positions them for reaction, makes formation of the transition state more frequent, and hence lowers the activation energy. (c) Effect of transition-state stabilization. An enzyme binds the transition state more tightly than it binds substrates, further lowering the activation energy. Thus, an enzymatic reaction has a much lower activation energy than an uncatalyzed reaction. (The breaks in the reaction curves indicate that the enzymes provide multistep pathways.)")
73
Figure 6.3 Enzymatic catalysis of the reaction A + B --> A - B(a) Energy diagram for an uncatalyzed reaction. (b) Effect of reactant binding. Collection of the two reactants in the EAB complex properly positions them for reaction, makes formation of the transition state more frequent, and hence lowers the activation energy. (c) Effect of transition-state stabilization. An enzyme binds the transition state more tightly than it binds substrates, further lowering the activation energy. Thus, an enzymatic reaction has a much lower activation energy than an uncatalyzed reaction. (The breaks in the reaction curves indicate that the enzymes provide multistep pathways.)
 Energy diagram for an uncatalyzed reaction. (b) Effect of reactant binding. Collection of the two reactants in the EAB complex properly positions them for reaction, makes formation of the transition state more frequent, and hence lowers the activation energy. (c) Effect of transition-state stabilization. An enzyme binds the transition state more tightly than it binds substrates, further lowering the activation energy. Thus, an enzymatic reaction has a much lower activation energy than an uncatalyzed reaction. (The breaks in the reaction curves indicate that the enzymes provide multistep pathways.)")
74
Transcurso de la formación de un complejo enzima-sustrato en función del tiempo e iniciación del estado estacionario, tal como se deduce de la resolución por computador de los datos obtenidos en un experimento real con una enzima típica.

75
Números máximos de recambio de algunas enzimas
Número de recambio (por segundo) Anhidrasa Carbónica Acetilcolinesterasa 25.000 Penicilinasa 2.000 Quimotripsina 100 DNA polimerasa I 15 Lisozima 0.5
 Anhidrasa Carbónica Acetilcolinesterasa Penicilinasa Quimotripsina DNA polimerasa I. 15. Lisozima")
76
Diagrama de energía para una reacción química

78
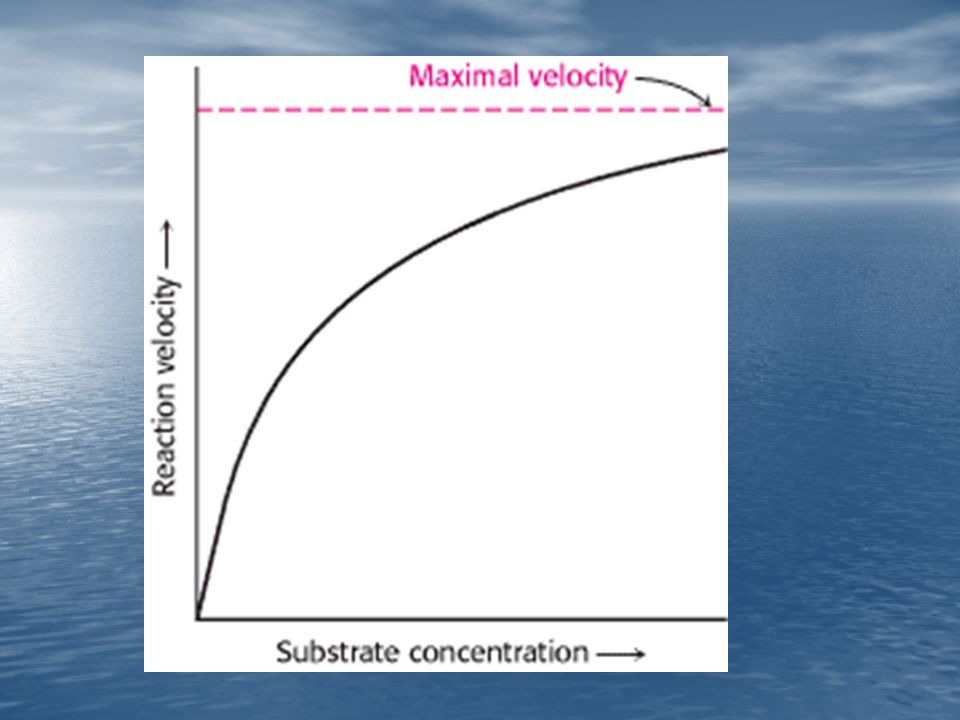
Para muchas enzimas, la velocidad de catálisis (V), varía con la concentración de sustrato [S], en la forma como muestra la figura. A una determinada concentración de enzima, la V es casi proporcional a [S] cuando la [S] es pequeña. Cuando la [S] es elevada, la V es prácticamente independiente de [S].
![Para muchas enzimas, la velocidad de catálisis (V), varía con la concentración de sustrato [S], en la forma como muestra la figura.](http://slideplayer.es/slide/1061018/3/images/78/Para+muchas+enzimas%2C+la+velocidad+de+cat%C3%A1lisis+%28V%29%2C+var%C3%ADa+con+la+concentraci%C3%B3n+de+sustrato+%5BS%5D%2C+en+la+forma+como+muestra+la+figura..jpg "A una determinada concentración de enzima, la V es casi proporcional a [S] cuando la [S] es pequeña. Cuando la [S] es elevada, la V es prácticamente independiente de [S].")
79
En 1913, Leonor Michaelis y Maud Menten hicieron la propuesta de un modelo sencillo.
E + S ES E + P K1 K3 K2

81
El complejo ES tiene dos destinos posibles:
Una enzima se combina con S para formar un complejo ES con una constante de velocidad K1. El complejo ES tiene dos destinos posibles: Puede disociarse hasta E y S, con una constante de velocidad K2; o puede continuar hasta formar un producto P, con una constante de velocidad K3.

82
La velocidad catalítica es igual al producto de la concentración del complejo ES por K3.

83
Las velocidades de formación y de destrucción de ES vienen dadas por:
Velocidad de formación de ES = (3) Velocidad de destrucción de ES = (4)
 Velocidad de destrucción de ES = (4)")
84
En el estado estacionario las concentraciones de los intermediarios permanecen invariables y las concentraciones de los materiales de partida y de los productos van cambiando. El Estado estacionario ocurre cuando la velocidad de formación y de destrucción del complejo ES son iguales.

85
Igualando las ecuaciones (3) y (4) tenemos,
(5)
")
86
reagrupando la ecuación (5)
(6)
")
87
KM denominada la constante de Michaelis se define como :
(7)
")
88
Sustituyendo en la ecuación (6), resulta
, resulta")
89
La concentración de sustrato no combinado [S] es casi igual a la concentración total de sustrato, bajo el supuesto que la concentración de enzima es mucho más baja que la concentración de sustrato.
![La concentración de sustrato no combinado [S] es casi igual a la concentración total de sustrato, bajo el supuesto que la concentración de enzima es mucho más baja que la concentración de sustrato.](http://slideplayer.es/slide/1061018/3/images/89/La+concentraci%C3%B3n+de+sustrato+no+combinado+%5BS%5D+es+casi+igual+a+la+concentraci%C3%B3n+total+de+sustrato%2C+bajo+el+supuesto+que+la+concentraci%C3%B3n+de+enzima+es+mucho+m%C3%A1s+baja+que+la+concentraci%C3%B3n+de+sustrato..jpg "La concentración de sustrato no combinado [S] es casi igual a la concentración total de sustrato, bajo el supuesto que la concentración de enzima es mucho más baja que la concentración de sustrato.")
90
La concentración de enzima no combinada [E], es igual a la concentración total del enzima ET ,menos la concentración de la enzima formando parte del complejo ES. (9)
![La concentración de enzima no combinada [E], es igual a la concentración total del enzima ET ,menos la concentración de la enzima formando parte del complejo ES.](http://slideplayer.es/slide/1061018/3/images/90/La+concentraci%C3%B3n+de+enzima+no+combinada+%5BE%5D%2C+es+igual+a+la+concentraci%C3%B3n+total+del+enzima+ET+%2Cmenos+la+concentraci%C3%B3n+de+la+enzima+formando+parte+del+complejo+ES..jpg "(9)")
91
sustituyendo [E] en la ecuación (8) por esta expresión:
(10)
![sustituyendo [E] en la ecuación (8) por esta expresión:](http://slideplayer.es/slide/1061018/3/images/91/sustituyendo+%5BE%5D+en+la+ecuaci%C3%B3n+%288%29+por+esta+expresi%C3%B3n%3A.jpg "(10)")
92
Reagrupando la ecuación (10)
(11)
")
93
O bien, (12)
")
94
Sustituyendo en la ecuación (2) por esta expresión, obtenemos:
 por esta expresión, obtenemos:")
95
La velocidad máxima (Vmax) se obtiene cuando los centros del enzima están saturados con sustrato. (cuando [S] es mucho mayor que KM, se aproxima a 1).
..")
96
Entonces: (14)
")
97
Sustituyendo la ecuación (14) en la ecuación (13) se obtiene la ecuación de Michaelis–Menten.
(15)
")
98
A concentraciones bajas de sustrato, cuando [S] es mucho menor que KM,
la velocidad de la reacción es directamente proporcional a la concentración del sustrato.
![A concentraciones bajas de sustrato, cuando [S] es mucho menor que KM,](http://slideplayer.es/slide/1061018/3/images/98/A+concentraciones+bajas+de+sustrato%2C+cuando+%5BS%5D+es+mucho+menor+que+KM%2C.jpg "la velocidad de la reacción es directamente proporcional a la concentración del sustrato.")
99
A concentraciones elevadas de sustrato, cuando la [S] es mucho mayor que KM, V=Vmax. La velocidad es independiente de la concentración del sustrato.
![A concentraciones elevadas de sustrato, cuando la [S] es mucho mayor que KM, V=Vmax.](http://slideplayer.es/slide/1061018/3/images/99/A+concentraciones+elevadas+de+sustrato%2C+cuando+la+%5BS%5D+es+mucho+mayor+que+KM%2C+V%3DVmax..jpg "La velocidad es independiente de la concentración del sustrato..")
100
Dependencia de Velocidad vs. [S]
![Dependencia de Velocidad vs. [S]](http://slideplayer.es/slide/1061018/3/images/100/Dependencia+de+Velocidad+vs.+%5BS%5D.jpg "Dependencia de Velocidad vs. [S]")
101
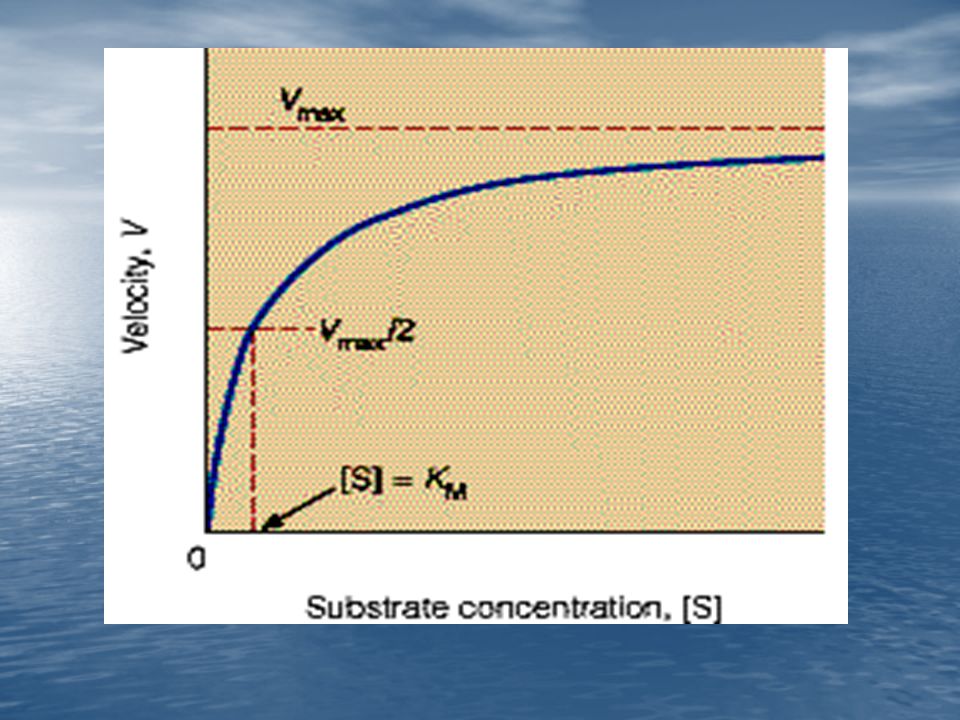
El significado de KM, es evidente en la ecuación (15) cuando [S]= KM, entonces V=Vmax/2.
KM es aquella concentración de sustrato a la cual la velocidad de reacción se hace la mitad de su valor máximo.
![El significado de KM, es evidente en la ecuación (15) cuando [S]= KM, entonces V=Vmax/2.](http://slideplayer.es/slide/1061018/3/images/101/El+significado+de+KM%2C+es+evidente+en+la+ecuaci%C3%B3n+%2815%29+cuando+%5BS%5D%3D+KM%2C+entonces+V%3DVmax%2F2..jpg "KM es aquella concentración de sustrato a la cual la velocidad de reacción se hace la mitad de su valor máximo.")
102
KM de algunas enzimas Enzima y sustrato KM (mn) Catalasa H2O2 25
Hexoquinasa Glucosa Fructosa Quimotripsina N – Benzoiltirosinamida N – Formiltirosinamida N - Acetiltirosinamida Gliciltirosinamida Anhidrasa Carbónica Glutamato –deshidrogenasa Glutamato L. Cetoglurarato NH

103
Números máximos de recambio de algunas enzimas
Número de recambio (por segundo) Anhidrasa Carbónica Acetilcolinesterasa 25.000 Penicilinasa 2.000 Quimotripsina 100 DNA polimerasa I 15 Lisozima 0.5
 Anhidrasa Carbónica Acetilcolinesterasa Penicilinasa Quimotripsina DNA polimerasa I. 15. Lisozima")
104
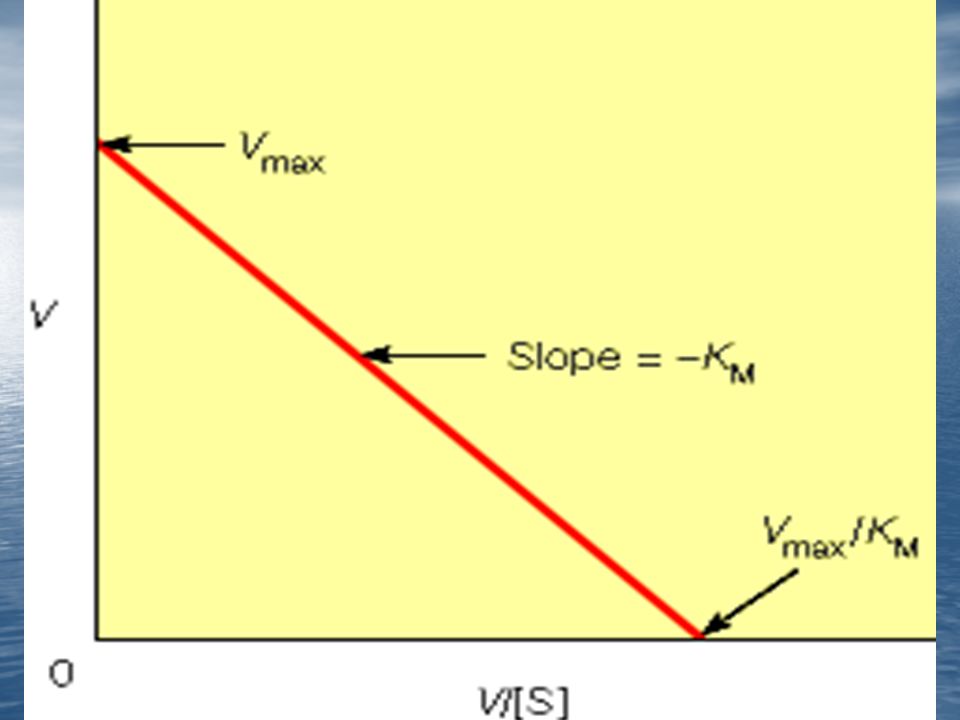
Es conveniente transformar la ecuación de Michaelis –Menten en otra que, representada gráficamente, resulte una línea recta.

106
Representación de Lineweaver-Burk: 1/v contra 1/(s)
")
108
Representación de Eadie-Hofstee: v contra v/(s)
")
109
Significado de los valores de KM y Vmax.
La constante de Michaelis, KM, tiene dos significados: La KM es la concentración de sustrato a la cual la mitad de los centros activos están ocupados. Una vez conocida KM , la fracción de centros llenos (fES), a cualquier concentración de sustrato puede calcularse a partir de
, a cualquier concentración de sustrato puede calcularse a partir de.")
110
(17)
")
111
2. KM se relacionan con la constante de velocidad de las etapas individuales, en el esquema catalítico de la ecuación (1). KM se define como: KM = (K2 + K3)K1 . Considerando K2 > K3 .
K1 . Considerando K2 > K3 .")
112
(18)
")
113
La constante de disociación del complejo ES viene dada por:
(19)
")
114
KM es igual a la constante de disociación del complejo ES, si K3 es mucho menor que K2.
KM es la medida de la estabilidad del complejo ES. Una KM alta indica unión débil, una KM baja indica unión fuerte.

115
La velocidad máxima, Vmax, revela el número de recambio de una enzima “Turnover number”, cuando se conoce la concentración de centros activos , porque (20)
")
116
La constante cinética K3 se denomina número de recambio.
El número de recambio de una enzima es el número de moléculas de sustrato convertidas en producto por unidad de tiempo, cuando la enzima está completamente saturada de sustrato.

117
Valores de la KM de algunas enzimas
Enzima Sustrato KM Quimotripsina Acetil –L-Triptofanamida 5 x 10-3M Lisozima Hexa –N- Acetilylucosamina 6 X 10-6M -Galactosidasa Lactosa 4 x 10-3M Treonina desaminasa Treonina 5 X 10-3M Anhidrasa Carbónica CO X 10-3M Penicilinasa Bencilpenicinina 5 X 10-5M Piruvato Carboxilasa Piruvato 4 X 10-4M

118
Valores de la KM de algunas enzimas
Enzima Sustrato KM Piruvato Carboxilasa HCO x10-3M ATP 6x10-5M Arginina –ERNA- Arginina 3x10-6M Sintetasa ERNA 4x10-7M ATP 3x10-4M

119
Números de recambio máximo de algunas enzimas.
Enzimas Número de recambio (por segundo) Anhidrasa Carbónica 3 – Cestosteroide isomerasa Acetilcolinesterasa Penicilinasa Lactosa deshidrogenasa Quimotripsina DNA polimerasa I Triptófano sintetasa Lisozima
 Anhidrasa Carbónica – Cestosteroide isomerasa Acetilcolinesterasa Penicilinasa Lactosa deshidrogenasa Quimotripsina 100. DNA polimerasa I 15. Triptófano sintetasa 2. Lisozima 0.5.")
Presentaciones similares