Descargar la presentación
La descarga está en progreso. Por favor, espere

1
METABOLISMO CELULAR: ENZIMAS

2
Sistema abierto Intercambio de materia y energía
Los seres vivos y las células que los forman son sistemas abiertos, en equilibrio y que realizan un trabajo. Sistema abierto Intercambio de materia y energía Equilibrio. Sus variables se mantienen dentro de unos niveles de tolerancia. Trabajo. Realiza trabajos dentro de su propia actividad de ser vivo (moverse, reproducirse, renovar tejidos…) Célula Energía Materia Energía Materia
 Célula. Energía. Materia. Energía. Materia.")
3
Concepto de metabolismo
El metabolismo es el conjunto de reacciones químicas que se producen en el interior de las células y que conducen a la transformación de unas biomoléculas en otras. Las distintas reacciones químicas del metabolismo se denominan vías metabólicas y las moléculas que intervienen se llaman metabolitos. Todas las reacciones del metabolismo están reguladas por enzimas, que son específicas para cada metabolito inicial o sustrato y para cada tipo de transformación. Las sustancias finales de una vía metabólica se denominan productos. Las conexiones existentes entre diferentes vías metabólicas reciben el nombre de metabolismo intermediario.

4
Se pueden considerar tres fases en el metabolismo:
Catabolismo: Transformación de moléculas orgánicas complejas en otras más sencillas, con liberación de energía que se almacena en ATP. Anabolismo: Síntesis de moléculas orgánicas complejas a partir de otras más sencillas. Se necesita suministrar energía, en forma de ATP Anfibolismo: (una fase intermedia). Procesos en los que se almacena gran cantidad de energía (para los posteriores procesos anabólicos)
. Procesos en los que se almacena gran cantidad de energía (para los posteriores procesos anabólicos)")
5
Moléculas que intervienen en el metabolismo Metabolitos
Glucosa, ácidos grasos… Nucleótidos NAD, FAD, NADP… Moléculas con enlaces ricos en energía ATP, coA Moléculas ambientales O2, H2O, CO2

6
Tipos de metabolismo Las células se encuentran siempre en un proceso constante de autodestrucción y autoregeneración. El metabolismo forma una unidad, aunque se estudia fragmentado en rutas o vías metabólicas. Las rutas metabólicas no son independientes entre si , poseen encrucijadas comunes. Un mismo metabolito común a dos rutas podrá seguir por una o por otra en función de las condiciones celulares.

7
Para crecer y desarrollarse, todos los seres vivos necesitan incorporar materia y energía y en función de estas clasificamos los distintos tipos de metabolismo de los seres vivos. MATERIA. Si la fuente de carbono es el dióxido de carbono (CO2 atmosférico) o carbono inorgánico, se habla de metabolismo autótrofo Si la fuente es la propia materia orgánica (formas más o menos reducidas del carbono como metano, glucosa, grasas, etc., es decir, el llamado carbono orgánico), se habla de metabolismo heterótrofo. ENERGIA Fotosintéticos si la fuente de energía es la luz. Quimiosíntéticos si es energía desprendida en reacciones químicas.
 o carbono inorgánico, se habla de metabolismo autótrofo. Si la fuente es la propia materia orgánica (formas más o menos reducidas del carbono como metano, glucosa, grasas, etc., es decir, el llamado carbono orgánico), se habla de metabolismo heterótrofo. ENERGIA. Fotosintéticos si la fuente de energía es la luz. Quimiosíntéticos si es energía desprendida en reacciones químicas.")
8
TIPO DE ORGANISMO FUENTE DE ENERGÍA FUENTE DE C ORGANISMOS
Fotolitótrofo Luz solar CO2 Vegetales. Bact. fotosintéticas Fotoorganótrofo Comp. orgánicos Bacterias purpúreas Quimiolitótrofo Reacciones redox Bacterias desnitrificantes Quimioorganótrofo Animales y Hongos

9
ATP + H2O ADP + Pi + energía (7,3 kcal/mol)
El ATP Puede actuar como molécula energética, al ser capaz de almacenar o ceder energía gracias a sus dos enlaces éster-fosfóricos que son capaces de almacenar cada uno de ellos, 7,3 kcal/mol. ATP + H2O ADP + Pi + energía (7,3 kcal/mol) ADP + H2O AMP + Pi + energía (7,3 kcal/mol) También se pueden dar las reacciones inversas (almacén de energía) Se dice que el ATP es la moneda energética de la célula, pues representa la manera de tener almacenado un tipo de energía de pronto uso. En ocasiones son utilizados para el mismo fin otros nucleótidos como el GTP el UTP o el CTP.
 ADP + H2O AMP + Pi + energía (7,3 kcal/mol) También se pueden dar las reacciones inversas (almacén de energía) Se dice que el ATP es la moneda energética de la célula, pues representa la manera de tener almacenado un tipo de energía de pronto uso. En ocasiones son utilizados para el mismo fin otros nucleótidos como el GTP el UTP o el CTP.")
10
La síntesis de ATP puede realizarse por dos vías:
Fosforilación a nivel de sustrato. Síntesis de ATP gracias a la energía que se libera de una biomolécula al romperse uno de sus enlaces ricos en energía, (ocurre en algunas reacciones de la glucólisis y del ciclo de Krebs). Las enzimas que regulan estos procesos se denominan quinasas.
. Las enzimas que regulan estos procesos se denominan quinasas.")
11
Fosforilación en el transporte de electrones.
Mediante enzimas del grupo de las ATP-sintetasas existentes en las crestas de las mitocondrias (fosforilación oxidativa) o en los tilacoides de los cloroplastos (fotofosforilación), cuando dichas enzimas son atravesadas por un flujo de protones (H+ ).
 o en los tilacoides de los cloroplastos (fotofosforilación), cuando dichas enzimas son atravesadas por un flujo de protones (H+ ).")
12
Muchas de las reacciones del catabolismo suponen la oxidación de un sustrato, lo cual libera electrones. Por el contrario, el anabolismo frecuentemente consiste en reacciones de reducción que requieren electrones. Los electrones son transportados desde las reacciones catabólicas de oxidación hasta las reacciones anabólicas de reducción. Intervienen coenzimas transportadores de electrones, como el NAD o el FAD, que llevan electrones de un punto a otro de la célula de un modo similar a como el ATP transporta la energía. Cuando uno de estos coenzimas se encuentra cargado de electrones, en estado oxidado, se dice que tiene poder reductor, puesto que al liberarse de los electrones podrá reducir a otro compuesto.

13
Reacciones catabólicas (oxidación de moléculas)
Liberación de e- que van a los coenzimas NAD FAD NADH FADH2 Liberación de e- desde los coenzimas que van a reducir otras moléculas Reacciones anabólicas (reducción)
")
14
Balance del metabolismo
Número de moléculas con enlaces ricos en energía (ATP u otros), que se producen por cada metabolito oxidado. En general: Rutas catabólicas: Balance positivo Rutas anabólicas: Balance negativo En las reacciones metabólicas, la energía generada se transforma, parte en ATP que si puede ser utilizado por la célula, y otra parte, se transfiere al entorno en forma de calor: Por ejemplo: Un mol de glucosa por combustión genera 680 Kcal. Mediante reacciones metabólicas da 36 ATP (262,8 Kcal) y 417 Kcal se pierden en forma de calor
, que se producen por cada metabolito oxidado. En general: Rutas catabólicas: Balance positivo. Rutas anabólicas: Balance negativo. En las reacciones metabólicas, la energía generada se transforma, parte en ATP que si puede ser utilizado por la célula, y otra parte, se transfiere al entorno en forma de calor: Por ejemplo: Un mol de glucosa por combustión genera 680 Kcal. Mediante reacciones metabólicas da 36 ATP (262,8 Kcal) y 417 Kcal se pierden en forma de calor.")
15
ATP/metabolito oxidado
Balance energético Relación: ATP/metabolito oxidado Balance positivo Se obtiene ATP Ruta catabólica Glucolisis Balance negativo Se gasta ATP Ruta anabólica Síntesis de proteínas

16
Control del metabolismo
1.- El control bioquímico Las sustancias que intervienen en el metabolismo celular son muy estables a temperatura ambiente Sin “ayuda” no reaccionarían o lo harían tan lentamente que no sería posible la vida. Esta dependencia de ayuda es paradójicamente una gran ventaja, ya que permite al organismo regular qué reacciones se han de dar y en que momento, es decir, el control bioquímico del metabolismo 2.- Control hormonal o sistema endocrino. El elemento fundamental de este sistema de control son las hormonas, que actúan específicamente sobre determinadas células como mensajeros químicos, regulando el metabolismo interno

17
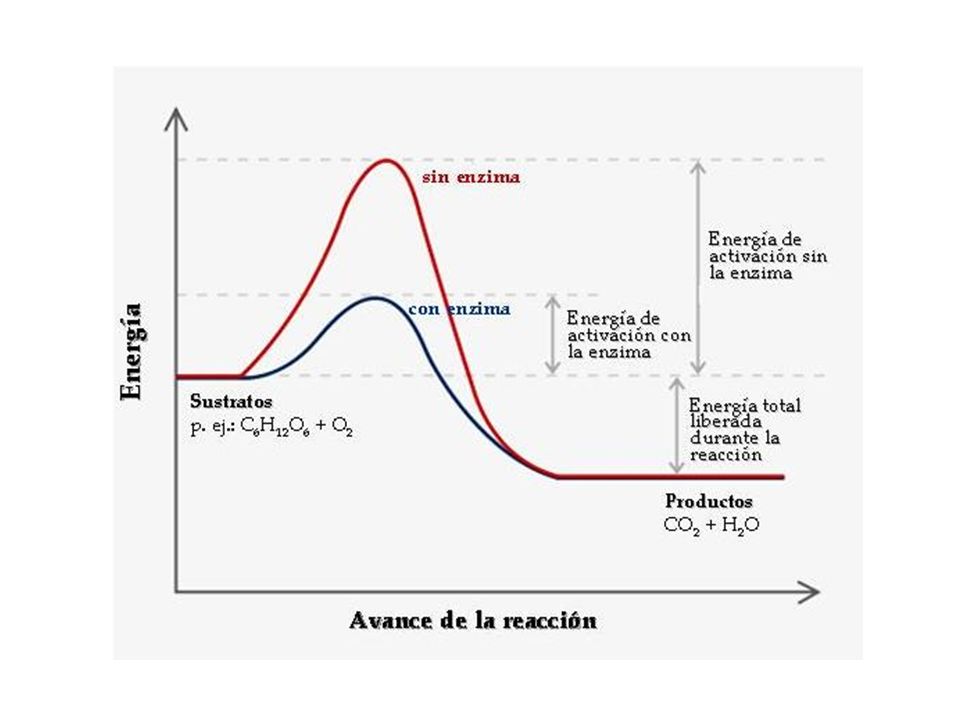
Muchas sustancias químicas, desprenden energía calorífica, son las reacciones exergónicas.
Este desprendimiento se debe a que la energía interna de los reactivos, (energía química de los enlaces), es mayor que la energía interna de las sustancias producidas (productos). Estas reacciones no se dan de forma espontánea porque para iniciar una reacción, primero es necesario suministrar la energía suficiente para debilitar los enlaces de los reactivos y posibilitar así su rotura. Reactivos Productos E. interna (reactivos) > E. interna (productos)
, es mayor que la energía interna de las sustancias producidas (productos). Estas reacciones no se dan de forma espontánea porque para iniciar una reacción, primero es necesario suministrar la energía suficiente para debilitar los enlaces de los reactivos y posibilitar así su rotura. Reactivos. Productos. E. interna (reactivos) > E. interna (productos)")
18
Tirar por la ventana un objeto que está sobre el suelo.
Este paso intermedio, que requiere un aporte de energía, recibe el nombre de estado de transición, y en él hay tantas posibilidades de que las moléculas acaben formando el producto como de que retrocedan. Energía de activación Estado de transición Productos Reactivos Ejemplos: Tirar por la ventana un objeto que está sobre el suelo. El papel no arde espontáneamente pese a la presencia de oxígeno en el aire, y sí lo hace cuando se calienta hasta una determinada temperatura.

19
Enzimas Para acelerar una reacción química también hay dos soluciones:
Calentar los reactivos. Añadir un catalizador, En los seres vivos, un aumento de temperatura podría provocar la muerte, por lo que se sigue el segundo mecanismo, es decir, el concurso de catalizadores biológicos o biocatalizadores. Las moléculas que desempeñan esta función son las enzimas

20
Enzimas Las enzimas son los catalizadores de las reacciones biológicas. Actúan rebajando la energía de activación, y por tanto acelerando la velocidad de la reacción, la cual se puede medir por la cantidad de producto que se forma por unidad de tiempo. Exceptuando las ribozimas, son proteínas globulares, solubles en agua, que se difunden bien en los líquidos orgánicos, y que pueden actuar a nivel intracelular, es decir, en el interior de la célula donde se han formado, o a nivel extracelular, en la zona donde se segregan, como sucede con las enzimas digestivas. Las ribozimas son unos ARN capaces de catalizar a otros ARN, quitándoles o añadiéndoles nucleótidos, sin consumirse ellos mismos. Se considera que en la primera materia viva la función catalítica la realizaba el ARN, luego aparecieron las proteínas, en las que se delegó la función enzimática, y los ADN, en los que se delegó, por su mayor estabilidad, la función de almacenar la información.

22
Las enzimas cumplen las dos características de todos los catalizadores:
Incluso en cantidades muy pequeñas, aceleran la reacción. No se obtiene más producto, sino la misma cantidad en menos tiempo. No se consumen durante la reacción biológica. Además, a diferencia de los catalizadores no biológicos, las enzimas presentan estas características: Son muy específicas. Pueden actuar en una reacción determinada sin alterar otras. Actúan siempre a temperatura ambiente, la temperatura del ser vivo. Son muy activas. Algunas consiguen aumentar la velocidad de reacción mas de un millón de veces, muy superior a los catalizadores no biológicos. Presentan un peso molecular muy elevado. Dada su naturaleza proteica, su síntesis implica una codificación genética.

23
Diferencias entre catalizadores biológicos y químicos
Son específicos para una determinada reacción química o para un grupo de reacciones químicas a para un sustrato o grupo de sustratos. Aceleran cualquier reacción inespecíficamente. Son proteínas (aunque hay ARN –Ribozimas- con función enzimática). Son sustancias simples finamente divididas. Son saturables No son saturables. Son altamente eficaces (son eficaces en bajas concentraciones). Son medianamente eficaces. Puede ser regulada su actividad catalítica. No pueden ser regulados. Son termolábiles y su actividad puede variar también de acuerdo al pH del medio. No son termolábiles ni se alteran con cambios de pH.
. Son sustancias simples finamente divididas. Son saturables. No son saturables. Son altamente eficaces (son eficaces en bajas concentraciones). Son medianamente eficaces. Puede ser regulada su actividad catalítica. No pueden ser regulados. Son termolábiles y su actividad puede variar también de acuerdo al pH del medio. No son termolábiles ni se alteran con cambios de pH.")
24
Factores que afectan la actividad enzimática
Influencia de la temperatura. Si a una reacción enzimática se le suministra energía calorífica, las moléculas aumentan su movilidad y el número de encuentros moleculares, por lo que aumenta la velocidad en que se forma el producto. Existe una temperatura óptima para la cual la actividad enzimática es máxima. Si la temperatura aumenta, se dificulta la unión enzima-sustrato y a partir de cierta temperatura la enzima se desnaturaliza, pierde su estructura terciaria y cuaternaria si la tiene y, por tanto, pierde su actividad enzimática.

25
Factores que afectan la actividad enzimática
Influencia del pH. Las enzimas presentan dos valores límite de pH entre los cuales son eficaces; traspasados estos valores, las enzimas se desnaturalizan y dejan de actuar. Entre los dos límites existe un pH óptimo en el que la enzima presenta su máxima eficacia. El pH óptimo está condicionado por el tipo de enzima y de sustrato, debido a que el pH influye en el grado de ionización de los radicales del centro activo de la enzima y también de los radicales del sustrato. Las variaciones de pH provocan cambios en las cargas eléctricas, alterando la estructura terciaria del enzima y por tanto, su actividad.

26
Factores que afectan la actividad enzimática
Inhibidores. Los inhibidores son sustancias que disminuyen la actividad de una enzima o bien impiden completamente la actuación de la misma. Pueden ser perjudiciales o beneficiosos como, por ejemplo, la penicilina, que es un inhibidor de las enzimas que regulan la síntesis de la pared bacteriana, por lo que es útil contra las infecciones bacterianas, y el AZT, que es un inhibidor de la transcriptasa inversa, por lo que retrasa el desarrollo del SIDA.

27
Factores que afectan la actividad enzimática
Concentración del sustrato A mayor concentración del sustrato, a una concentración fija de la enzima se obtiene la velocidad máxima. Después de que se alcanza esta velocidad, un aumento en la concentración del sustrato no tiene efecto en la velocidad de la reacción. Concentración de la enzima Siempre y cuando haya sustrato disponible, un aumento en la concentración de la enzima aumenta la velocidad enzimática hacia cierto límite.

28
Velocidad de la reacción
Concentración de sustrato (concentración de enzima fija) Velocidad de la reacción Después de que se alcanza esta velocidad, un aumento en la concentración del sustrato no tiene efecto en la velocidad de la reacción. (todos los enzimas están ocupados) A medida que aumenta la concentración de sustrato, aumenta la velocidad de reacción (mientras queden enzimas libres).
 Velocidad de la reacción. Después de que se alcanza esta velocidad, un aumento en la concentración del sustrato no tiene efecto en la velocidad de la reacción. (todos los enzimas están ocupados) A medida que aumenta la concentración de sustrato, aumenta la velocidad de reacción (mientras queden enzimas libres).")
29
Velocidad de la reacción
Después de que se alcanza esta velocidad, un aumento en la concentración del enzima no tiene efecto en la velocidad de la reacción. (no hay más sustrato que procesar) Velocidad de la reacción A medida que aumenta la concentración de enzima, aumenta la velocidad de reacción (mientras queden sustrato sin reaccionar). Concentración de enzima (concentración de sustrato fija)
 Velocidad de la reacción. A medida que aumenta la concentración de enzima, aumenta la velocidad de reacción (mientras queden sustrato sin reaccionar). Concentración de enzima (concentración de sustrato fija)")
30
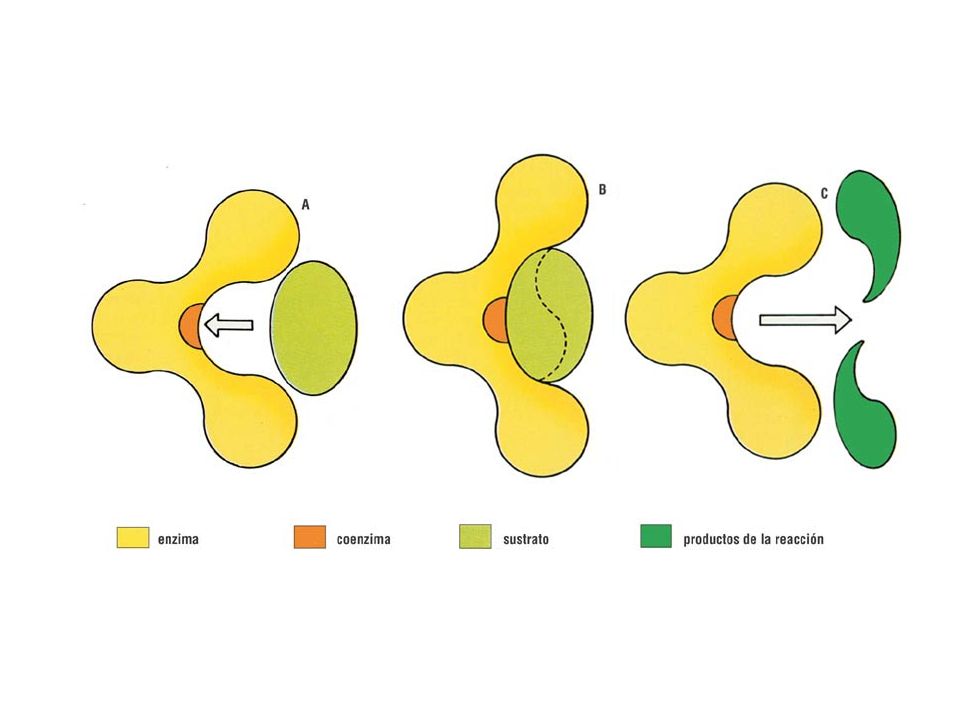
Cofactores enzimáticos
Enzimas Sólo proteínas Holoenzimas Cofactor Cationes metálicos (Ca2+ Fe2+..) Moléculas orgánicas NAD, FAD Coenzimas Grupo prostético (Grupo hemo) Apoenzima (parte proteica)
 Moléculas orgánicas. NAD, FAD. Coenzimas. Grupo prostético. (Grupo hemo) Apoenzima (parte proteica)")
31
Nomenclatura de los Enzimas
1. NOMENCLATURA ANTIGUA: SUFIJO -asa Nombre de la fuente u origen del enzima: Pancreasa Nombre del sustrato: Proteasa Tipo de reacción catalizada: Hidrolasa 2. NOMENCLATURA ACTUAL: Enzyme Commission [E.C.] de la IUBMB Clasificación de enzimas (6 clases) Asignación de código E.C.: Nombre sistemático: Sustrato:Cosustrato Tipo de Reacción -asa Etanol:NAD+ Oxidorreductasa
 Asignación de código E.C.: Nombre sistemático: Sustrato:Cosustrato Tipo de Reacción -asa. Etanol:NAD+ Oxidorreductasa.")
32
Clasificación de los Enzimas
CLASE TIPO DE REACCION CATALIZADA 1. OXIDO-REDUCTASAS Transferencia de electrones 20 subclases Sred + S’ox Sox + S’red 2. TRANSFERASAS Transferencia de grupos 9 subclases S-grupo + S’ S’-grupo + S 3. HIDROLASAS Rotura hidrolítica de enlaces 11 subclases A-B + H2O A-H + B-OH 4. LIASAS Rotura de enlaces A-B A+B 7 subclases Salida de grupos CX-CY C=C + X-Y Adición a dobles enlaces C=C + XY CX-CY 5. ISOMERASAS Cambios internos 6 subclases Transferencias internas de grupos 6. LIGASAS Formación de enlaces mediante reacciones de 5 subclases condensación con gasto de energía (ATP) International Union of Biochemistry and Molecular Biology [IUBMB]
 International Union of Biochemistry and Molecular Biology [IUBMB]")
33
La reacción enzimática
La enzima (E) actúa fijando al sustrato en su superficie (adsorción) mediante enlaces débiles Se forma el complejo enzíma-sustrato (ES). Se generan tensiones que debilitan los enlaces del sustrato, por lo que para llegar al estado de transición del complejo enzima-sustrato, (complejo activado) se requiere mucha menos energía que para llegar al estado de transición del sustrato solo. Se liberan la enzima intacta (E) y el producto (P)
 actúa fijando al sustrato en su superficie (adsorción) mediante enlaces débiles. Se forma el complejo enzíma-sustrato (ES). Se generan tensiones que debilitan los enlaces del sustrato, por lo que para llegar al estado de transición del complejo enzima-sustrato, (complejo activado) se requiere mucha menos energía que para llegar al estado de transición del sustrato solo. Se liberan la enzima intacta (E) y el producto (P)")
35
El centro activo de los enzimas
La actividad enzimática se inicia con la formación del complejo ES. Esta unión se realiza gracias a los radicales de algunos pocos aminoácidos que establecen enlaces con el sustrato (y con el grupo prostético si lo hay), fijándolo y luego rompiendo alguno de sus enlaces. La región de la enzima que se une al sustrato recibe el nombre de centro activo.
, fijándolo y luego rompiendo alguno de sus enlaces. La región de la enzima que se une al sustrato recibe el nombre de centro activo.")
36
Características del centro activo
Es una parte muy pequeña del volumen total de la enzima. Tienen una estructura tridimensional en forma de hueco que facilita encajar al sustrato. Están formados por aminoácidos lejanos en la secuencia polipeptídica, que debido a los repliegues de ésta, quedan próximos. Los radicales de estos aminoácidos presentan afinidad por el sustrato, lo atraen y establecen enlaces débiles con él. Esto facilita que, una vez roto alguno de sus enlaces, los productos resultantes se puedan separar con facilidad del centro activo.

37
En una enzima se pueden distinguir tres tipos de aminoácidos
Aminoácidos estructurales. Son los que no establecen enlaces químicos con el sustrato Son los más abundantes y los responsables de la forma de la enzima. Por ejemplo, en la lisozima de 129 aminoácidos, 124 son estructurales y sólo 5 no lo son. Aminoácidos de fijación. Son los que establecen enlaces débiles con el sustrato y lo fijan. Se encuentran en el centro activo de la enzima Aminoácidos catalizadores. Son los que al establecer enlaces, débiles o fuertes (covalentes), con el sustrato, provocan la rotura de alguno de sus enlaces. Son los responsables de su transformación. También están en el centro activo Centro activo
, con el sustrato, provocan la rotura de alguno de sus enlaces. Son los responsables de su transformación. También están en el centro activo. Centro activo.")
38
La especificidad de los enzimas
Fischer (1890) Modelo «llave (sustrato) - cerradura (enzima)»
 Modelo «llave (sustrato) - cerradura (enzima)»")
39
La especificidad de los enzimas
En la actualidad se ha visto que algunas enzimas, al establecer los enlaces con el sustrato, modifican la forma de sus centros activos para adaptarse mejor al sustrato, es decir, solamente son complementarias después de haberse unido a él, es el llamado acoplamiento inducido (como el guante (enzima) se adapta a la mano (sustrato)).
 se adapta a la mano (sustrato)).")
40
La especificidad puede darse en varios grados.
Especificidad absoluta. Se da cuando la enzima sólo actúa sobre un sustrato, por ejemplo, la ureasa sólo actúa sobre la urea. Especificidad de grupo. Se da cuando la enzima reconoce un determinado grupo de moléculas, por ejemplo, la β-glucosidasa que actúa sobre todos los β-glucósidos Especificidad de clase. Es la menos específica, dado que la actuación de la enzima no depende del tipo de molécula, sino del tipo de enlace Por ejemplo, las fosfatasas separan los grupos fosfato de cualquier tipo de molécula.

41
Cinética de la actividad enzimática
En una reacción enzimática con una concentración de enzima constante, al incrementar la concentración del sustrato se produce un aumento de la velocidad de reacción. Este incremento en la velocidad de reacción se debe a que, al haber más moléculas de sustrato por unidad de volumen, se aumenta la probabilidad de encuentro entre sustrato y enzima. Llega un momento en que la velocidad de reacción deja de crecer, es decir, se llega a una velocidad máxima (Vmax). Esto se debe a que todas las moléculas de la enzima ya están ocupadas por moléculas de sustrato, formando el complejo enzima-sustrato, lo que se denomina saturación de la enzima.
. Esto se debe a que todas las moléculas de la enzima ya están ocupadas por moléculas de sustrato, formando el complejo enzima-sustrato, lo que se denomina saturación de la enzima.")
42
Constante de Michaelis-Menten (KM).
Es la concentración del sustrato a la cual la velocidad de reacción es la mitad de la velocidad máxima. KM depende de la afinidad que hay entre la enzima y su sustrato. Un valor de KM pequeño indica mucha afinidad (la mitad de la velocidad máxima se alcanza a concentraciones de sustrato pequeñas) Se denomina número de recambio (turnover) o constante catalítica al número de moléculas de sustrato transformadas por unidad de tiempo.
 Se denomina número de recambio (turnover) o constante catalítica al número de moléculas de sustrato transformadas por unidad de tiempo.")
43
Inhibición enzimática
La inhibición puede ser de dos tipos: irreversible y reversible. La inhibición irreversible. Los inhibidores irreversibles son los que se combinan o destruyen un sitio esencial para la actividad de la enzima. La inhibición reversible tiene lugar cuando no se inutiliza el centro activo, sino que sólo se impide temporalmente su normal funcionamiento. Existen tres modalidades: Competitiva No competitiva Acompetitiva

44
Inhibición enzimática
Inhibidores enzimáticos Reversibles Competitivos Conformación similar al sustrato No competitivos Se unen a un sitio distinto al centro activo (alosterismo) Acompetitivos Se unen al complejo ES Irreversibles Venenos
 Acompetitivos. Se unen al complejo ES. Irreversibles. Venenos.")
45
La inhibición reversible competitiva se debe a la presencia de un inhibidor cuya molécula es similar al sustrato por lo que compite con este en la fijación al centro activo del enzima. Si se fija el inhibidor, la enzima queda bloqueada. La velocidad de la reacción disminuye en función de la concentración del inhibidor.

46
La inhibición reversible no competitiva es cuando un determinado producto se une a un lugar del enzima distinto del centro activo, cambiando la forma del enzima e impidiendo la unión del sustrato

47
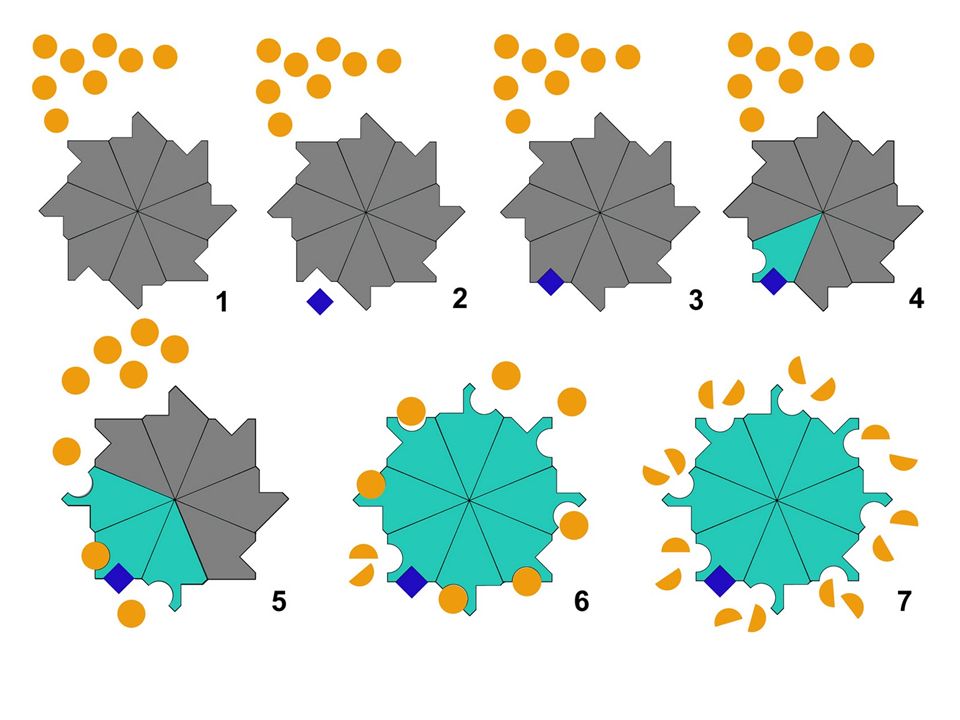
Enzimas alostéricas Las enzimas alostéricas son aquellas que pueden adoptar dos formas estables diferentes (activa e inactiva). Estas enzimas, además del centro activo, tienen al menos otro lugar, denominado centro regulador, al que se puede unir una determinada sustancia, denominada ligando. Los ligandos pueden ser activadores o inhibidores.

48
Inhibición alostérica.
sustrato Enzima activa Sin inhibidor Los inhibidores alostéricos se unen a una zona de la enzima y cambian la configuración del centro activo de tal manera que impiden que el sustrato se pueda unir a él. Enzima inactiva con inhibidor inhibidor

49
El alosterismo permite la autorregulación de la actividad enzimática
El alosterismo permite la autorregulación de la actividad enzimática. Hay dos casos: Regulación por retroinhibición o inhibición feed-back. Se da en enzimas cuya conformación inicial es la activa. Se produce cuando el producto final es el que al fijarse al centro regulador actúa como inhibidor, provocando la transición alostérica a la forma inactiva de la enzima. Regulación por inducción enzimática. Se da en enzimas cuya conformación inicial es la inactiva. Se produce cuando alguna sustancia inicial es la que al fijarse sobre el centro regulador provoca la transición alostérica a la forma activa de la enzima, por lo que ésta empieza a actuar sobre el sustrato

50
Cooperativismo Las enzimas alostéricas suelen estar formadas por varias subunidades moleculares denominadas protómeros. Cada protómero posee un centro activo y al menos un centro regulador. En muchas enzimas, cuando al centro regulador se une una molécula denominada activador o ligando, la conformación del protómero varía, haciendo funcional al centro activo. La variación en la conformación de este protómero se transmite instantáneamente a los otros protómeros asociados haciéndolos a su vez activos, efecto que se denomina transmisión alostérica. De esta forma la enzima pasa de estado inhibido a un estado activo o catalítico.

51
Como para que una enzima alostérica actué es precisa la unión de uno o más ligandos y luego la del sustrato, la velocidad de reacción en función de la concentración de sustrato no aumenta tan rápidamente como en las enzimas no alostéricas. En cambio, si esta enzima es una subunidad y su transformación al estado activo se transmite alostéricamente a todas las demás subunidades, son muchas las que súbitamente empiezan a actuar, lo que se denomina cooperativismo, y se produce un cambio de velocidad de reacción muy grande, la llamada «ley del todo o nada». La representación gráfica de la relación entre la velocidad de reacción y la concentración del sustrato no es una hipérbola, sino una curva sigmoidal o curva en «S»

52
Cooperatividad Positiva Muy sensible a cambios en la [sustrato] Negativa Insensible a cambios en la [sustrato] Representación de los distintos cambios conformacionales que padece la hemoglobina al unirse con una molécula de oxígeno Un molécula de oxígeno se puede unir con el hierro (II) del grupo hemo en cada una de las cuatro cadenas de la molécula de hemoglobina. La desoxihemoglobina tiene una relativa baja afinidad para el oxígeno, pero cuando una molécula se une a un único hemo, crece la afinidad por el oxígeno, permitiendo que la segunda molécula se una más fácilmente, y la tercera y cuarta aun más fácilmente.
![Cooperatividad Positiva. Muy sensible a cambios en la [sustrato] Negativa. Insensible a cambios en la [sustrato]](http://slideplayer.es/slide/101373/1/images/52/Cooperatividad+Positiva.+Muy+sensible+a+cambios+en+la+%5Bsustrato%5D+Negativa.+Insensible+a+cambios+en+la+%5Bsustrato%5D.jpg "Representación de los distintos cambios conformacionales que padece la hemoglobina al unirse con una molécula de oxígeno. Un molécula de oxígeno se puede unir con el hierro (II) del grupo hemo en cada una de las cuatro cadenas de la molécula de hemoglobina. La desoxihemoglobina tiene una relativa baja afinidad para el oxígeno, pero cuando una molécula se une a un único hemo, crece la afinidad por el oxígeno, permitiendo que la segunda molécula se una más fácilmente, y la tercera y cuarta aun más fácilmente.")
53
La cooperatividad es un fenómeno común y puede llegar a ser crucial en la regulación de la respuesta enzimática a cambios en la concentración de sustrato. La cooperatividad positiva hace que la enzima sea mucho más sensible a la concentración de sustrato, con lo que su actividad puede llegar a variar en gran medida aunque se mueva en rangos muy estrechos de concentración de sustrato. La cooperatividad negativa hace que la enzima sea insensible a pequeños cambios en la concentración de sustrato. Representación de los distintos cambios conformacionales que padece la hemoglobina al unirse con una molécula de oxígeno

54
Animaciones: Mecanismo de acción enzimática e inhibición Regulación alostérica de enzimas Retroinhibición de vías bioquímicas

56
Sustratos Si el ligando no se une al centro regulador el enzima no puede unirse al sustrato Ligandos Si el ligando se une al centro regulador , el centro activo se modifica y el enzima se une al sustrato

57
Inhibición acompetitiva
inhibidor Enzima sustrato El centro activo está ocupado. El sustrato no puede entrar Enzima sustrato Inhibición competitiva regulador Enzima sustrato Inhibición alostérica El centro activo se modifica por la acción del regulador. El sustrato no puede entrar sustrato El sustrato esta modificado, no puede entrar en el centro activo Inhibición acompetitiva

58
Eficacia de las vías metabólicas
En las vías metabólicas el producto generado por una enzima es el sustrato de la siguiente enzima, por ello, para aumentar la eficiencia del sistema hay distintos mecanismos: La compartimentación. Consiste en separar mediante membranas los lugares donde se realizan aquellas vías metabólicas que no se desea que se relacionen Complejo multienzimático. Es la asociación de varias enzimas que actúan sucesivamente en una vía. El complejo supramolecular resultante es más eficaz que si las enzimas estuvieran dispersas en el medio. Inclusión en membranas. Algunas enzimas y algunos complejos multienzimáticos se encuentran englobados de forma ordenada en las membranas, de forma que esto facilita la unión entre los sucesivos productos y las sucesivas enzimas.

59
Vitaminas Son moléculas muy variadas que pueden pertenecer a distintos grupos de principios inmediatos. Algunas son indispensables en la dieta, ya que no pueden ser sintetizadas por el organismo (excepto la B5). Otras vitaminas son necesarias para la actuación de determinados enzimas, ya que funcionan como coenzimas que intervienen en distintas rutas metabólicas y , por ello, una deficiencia en una vitamina puede originar importantes defectos metabólicos. Las cantidades necesarias son mínimas (una dieta variada garantiza las necesidades del organismo) Vitaminas Defecto Avitaminosis Hipovitaminosis Exceso Hipervitaminosis
. Otras vitaminas son necesarias para la actuación de determinados enzimas, ya que funcionan como coenzimas que intervienen en distintas rutas metabólicas y , por ello, una deficiencia en una vitamina puede originar importantes defectos metabólicos. Las cantidades necesarias son mínimas (una dieta variada garantiza las necesidades del organismo) Vitaminas. Defecto. Avitaminosis. Hipovitaminosis. Exceso. Hipervitaminosis.")
60
Las vitaminas se clasifican según su solubilidad en agua:
Vitaminas hidrosolubles. Actúan como coenzimas o precursores de coenzimas. Son las del complejo B o la vitamina C Vitaminas liposolubles. No son solubles en agua y si en disolventes no polares. Son lípidos insaponificables. No suelen ser cofactores o precursores. Son las vitaminas A,D,E y K

61
Enfermedades carenciales
VITAMINAS FUNCIONES Enfermedades carenciales C (ácido ascórbico) Coenzima de algunas peptidasas. Interviene en la síntesis de colágeno Escorbuto B1 (tiamina) Coenzima de las descarboxilasas y de las enzima que transfieren grupos aldehidos Beriberi B2 (riboflavina) Constituyente de los coenzimas FAD y FMN Dermatitis y lesiones en las mucosas B3 (ácido pantoténico) Constituyente de la CoA Fatiga y trastornos del sueño B5 (niacina) Constituyente de las coenzimas NAD y NADP Pelagra B6 ( piridoxina) Interviene en las reacciones de transferencia de grupos aminos. Depresión, anemia B12 (cobalamina) Coenzima en la transferencia de grupos metilo. Anemia perniciosa Biotina Coenzima de las enzimas que transfieren grupos carboxilo, en metabolismo de aminoácidos. Fatiga, dermatitis... A (retinol) Ciclo visual, crecimiento, protección y mantenimiento del tejido epitelial Ceguera nocturna, xeroftalmia, desecación epitelial D Metabolismo del Ca2+, esencial en el crecimiento y mantenimiento de los huesos Raquitismo, deformidades oseas
 Coenzima de algunas peptidasas. Interviene en la síntesis de colágeno. Escorbuto. B1 (tiamina) Coenzima de las descarboxilasas y de las enzima que transfieren grupos aldehidos. Beriberi. B2 (riboflavina) Constituyente de los coenzimas FAD y FMN. Dermatitis y lesiones en las mucosas. B3 (ácido pantoténico) Constituyente de la CoA. Fatiga y trastornos del sueño. B5 (niacina) Constituyente de las coenzimas NAD y NADP. Pelagra. B6 ( piridoxina) Interviene en las reacciones de transferencia de grupos aminos. Depresión, anemia. B12 (cobalamina) Coenzima en la transferencia de grupos metilo. Anemia perniciosa. Biotina. Coenzima de las enzimas que transfieren grupos carboxilo, en metabolismo de aminoácidos. Fatiga, dermatitis... A (retinol) Ciclo visual, crecimiento, protección y mantenimiento del tejido epitelial. Ceguera nocturna, xeroftalmia, desecación epitelial. D. Metabolismo del Ca2+, esencial en el crecimiento y mantenimiento de los huesos. Raquitismo, deformidades oseas.")
Presentaciones similares












![1 KM 1 1 Vo= Vmax [S] + Vmax Y = m x + b = m = b.](/2/161459/big_thumb.jpg "1 KM 1 1 Vo= Vmax [S] + Vmax Y = m x + b = m = b.>")






