Descargar la presentación
La descarga está en progreso. Por favor, espere

1
M.Orozco J.L.Gelpi M.Rueda J.R.Blas

2
No a la Guerra Curso Universitat de Barcelona –CESCA
Xarxa Catalana Bioinformàtica No a la Guerra Xarxa catalana de Bioinformàtica

3
Clase 1: Elementos de estructura

4
Estructura de proteínas
Estructura de ácidos nucleicos

5
Estructura de proteínas
Estructura de ácidos nucleicos

6
Secuencia DNA Gens RNA(m) Proteínas
 Proteínas")
9
NIVELES DE ESTRUCTURA Estructura primaria (secuencia)
Estructura secundaria (hélices) Estructura terciaria (superhélices,...) Estructura cuaternaria (complejos,...)
 Estructura terciaria (superhélices,...) Estructura cuaternaria (complejos,...)")
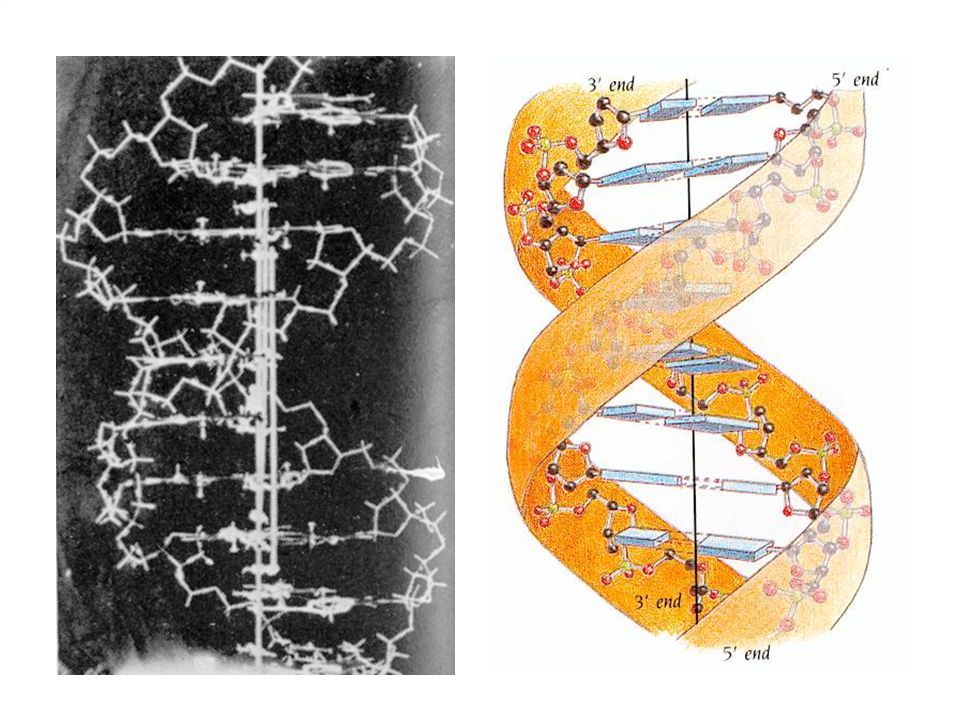
10
CONSTITUYENTES ESTRUCTURALES DE LOS ÁCIDOS NUCLEICOS

11
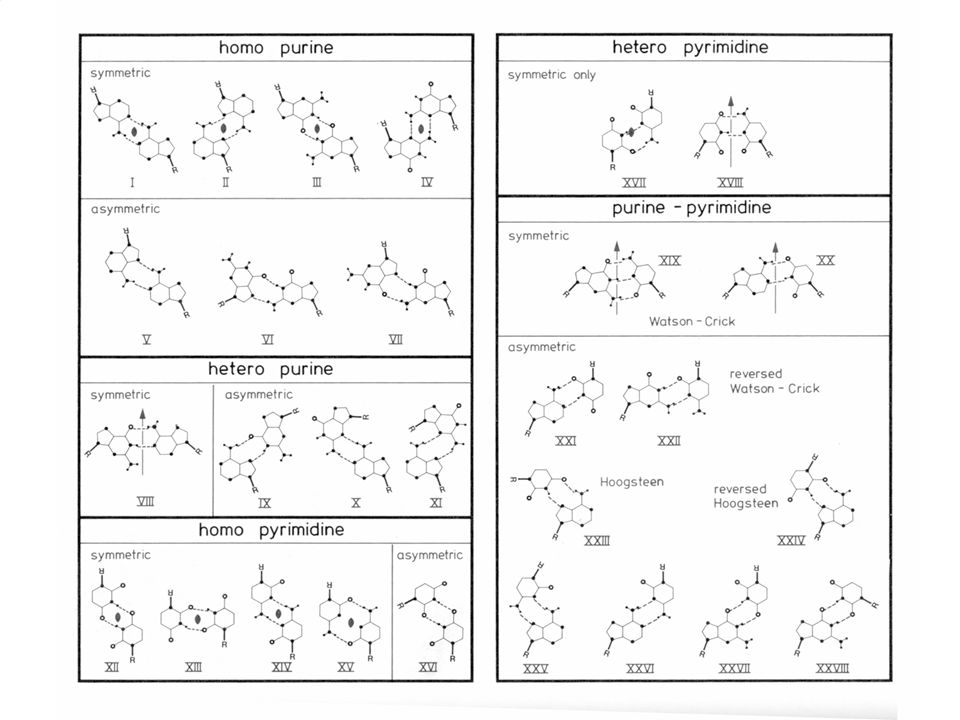
BASES NITROGENADAS Pirimidinas Purinas

12
LAS BASES POSEEN UNA HUELLA DACTILAR UNICA
BASADA EN SU PAUTA DE PUENTES DE HIDROGENO

13
EL AZUCAR: RIBO: RNA; DEOXY: DNA

14
Como regla general,... Las bases son responsables de delimitar las propiedades de reconocimiento específico del DNA. El esqueleto de fosforibosa (conformación del azucar, enlaces fosfodiester,...) marcan la flexibilidad conformacional.
 marcan la flexibilidad conformacional.")
15
El paso entre confórmeros del azucar se da siempre a través de otros especies no planas

16
CONFORMACION DE LA RIBOSA
(PSEUDOROTACION) La transición entre puckerings del azucar se da siguiendo el denominado Círculo Pseudorotacional 360>P>0 para tm constante
 La transición entre. puckerings del. azucar se da siguiendo. el denominado. Círculo. Pseudorotacional. 360>P>0. para tm constante.")
17
La transición C2’endo<->C3’ endo es siempre por O4’endo
nunca por O4’exo S<->N por E nunca por W

18
PUCKERING Las formas más abundantes en nucleósidos son la N (C3’endo) y la S (C2’endo) La forma E (O4’endo) se encuentra a veces, pero nunca la W (O4’exo) Las formas B del DNA presentan puckerings S y en algún caso E. Las formas A del DNA y el RNA presentan siempre puckerings N
 se encuentra a veces, pero nunca la W (O4’exo) Las formas B del DNA presentan puckerings S y en algún caso E. Las formas A del DNA y el RNA presentan siempre puckerings N.")
19
NUCLEOSIDO (base + azucar)
")
20
NUCLEOTIDO (base + azucar + fosfato)
")
21
ENLACE GLICOSIDICO syn anti

22
ENLACE GLICOSIDICO Normalmente los nucleósidos y nucleótidos están en la conformación anti La Guanosina es el único nucleósido (de los A.N.) con posibilidad de estar en conformación syn Grupos voluminosos en la posición 8 de la purina pueden forzar la conformación syn.
 con posibilidad de estar en conformación syn. Grupos voluminosos en la posición 8 de la purina pueden forzar la conformación syn.")
23
ENLACE C4’-C5’ gauche-gauche trans-gauche gauche-trans

24
ENLACE C4’-C5’ Las tres conformaciones se detectan
La gg parece un poco más estable, pero la diferencia es muy pequeña El puckering del azucar influye mucho en la conformación del enlace C4’-C5’

25
EL DNA ES EL FRUTO DE LAS INTERACCIONES DE LOS NUCLEOTIDOS
Interacciones de puente de hidrógeno. Interacciones de stacking (apilamiento). Screening (apantallamiento de la repulsión fosfato-fosfato) Efecto solvatación del agua y contraiones
. Screening (apantallamiento de la repulsión fosfato-fosfato) Efecto solvatación del agua y contraiones.")
26
Apareamientos canónicos (Watson-Crick)
")
27
Los apareamientos W.C no son los únicos posibles!

29
Los apareamientos WC no saturan todas las posibilidades de reconocimientos específicos

30
Apareamiento d(A:T)
")
31
INTERACCIONES DE STACKING
Son interacciones (básicamente de van der Waals) entre anillos aromáticos de las bases nitrogenadas Intensas, pero menos específicas que los ptes de hidrógeno Menos dependientes del solvente que los ptes de hidrógeno Las purina-purina son las más intensas
 entre anillos aromáticos de las bases nitrogenadas. Intensas, pero menos específicas que los ptes de hidrógeno. Menos dependientes del solvente que los ptes de hidrógeno. Las purina-purina son las más intensas.")
32
d(A:T)2 El stacking y el puente de hidrógeno mantienen la estructura de la doble hélice
2 El stacking y el puente de hidrógeno mantienen la estructura de la doble hélice")
33
d(A:T)2 El stacking y el puente de hidrógeno mantienen la estructura de la doble hélice
2 El stacking y el puente de hidrógeno mantienen la estructura de la doble hélice")
35
EL DNA PRESENTA ES UNA HELICE NO PLANA SINO CON PERFILES (SURCOS)
LOS SURCOS SON TOTALMENTE DIFERENTES EN EL A- Y EN EL B- DNA

36
EL DNA PRESENTA ES UNA HELICE NO PLANA SINO CON PERFILES (SURCOS)
EN CADA PAR DE BASES SE DEFINEN DOS SURCOS EL MAJOR Y EL MINOR GROOVES GRAN PARTE DE LA REACTIVIDAD ESTA LOCALIZADA EN LOS SURCOS

37
Diversos fármacos interaccionan específicamente con las bases por el MINOR groove. Muchas proteínas lo hacen por el MAJOR groove. Otros oligonucleótidos pueden hacerlo por el MAJOR groove

39
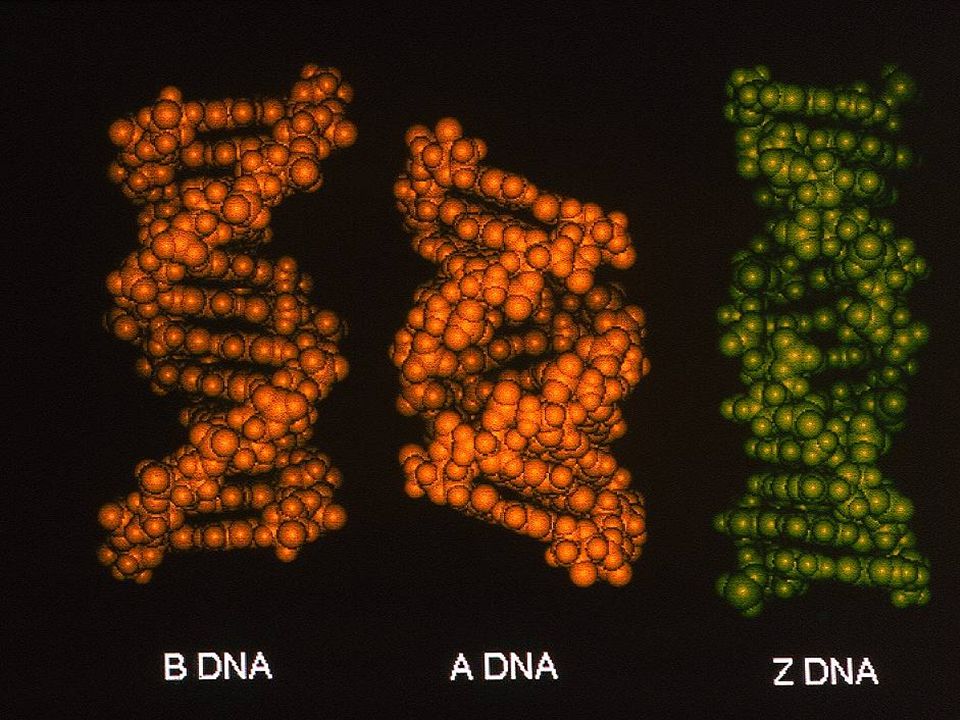
LA ESTRUCTURA SECUNDARIA DEL DNA ES POLIMORFICA
Depende de la secuencia Varía con la humedad del medio Cambia con la fuerza iónica Depende de la presencia de drogas Puede ser alterada por la acción de proteínas Ciertas sales pueden hacerla cambiar

40
La estructura secundaria nativa del DNA es “siempre” helicoidal
Dobles hélices (A, B, Z) Triples hélices (d(Pu:Py:Pu); d(Pu:Py:Py)) Cuadruples hélices (motivo G-DNA)
 Triples hélices (d(Pu:Py:Pu); d(Pu:Py:Py)) Cuadruples hélices (motivo G-DNA)")
43
COMO REGLA GENERAL El DNA fisiológico adopta la forma dextrógira B
El RNA fisiológico adopta la forma dextrógira A Los híbridos B:A adoptan la forma dextrógira A

44
TRIPLES HELICES Se forman por una tercera cadena de oligonucleótidos reconoce a un duplex. Los motivos pirimidina (3a cadena poly-Y) son los más conocidos Se pueden formar con DNA, RNA, PNA,... Son estructuras que existen in vivo y pueden tener gran impacto biotecnológico y terapéutico
 son los más conocidos. Se pueden formar con DNA, RNA, PNA,... Son estructuras que existen in vivo y pueden tener gran impacto biotecnológico y terapéutico.")
45
Triplexes motivos pirimidina

47
LOS TRIPLEX SE FORMAN AL RECONOCER EL SURCO ANCHO UNA CADENA SIMPLE A UN DNA DUPLEX

48
ES POSIBLE INFLUIR EN LA FUNCIONALIDAD DEL DNA
MEDIANTE LA FORMACION DE ESTRUCTURAS HIBRIDAS DNA(T): ANTI-GENE & DNA-RNA: ANTISENSE
: ANTI-GENE & DNA-RNA: ANTISENSE.")
49
TETRAPLEXES: G-DNA Se forman fundamentalmente en secuencias ricas en poly(G) Claves para la estabilización de los telómeros Una de las dianas más prometedoras en tratamiento antineoplásico.

50
El G-DNA SE FORMA POR LA INTERACCION EN EL PLANO
DE 4 GUANINAS. LA REPETICIÓN DE ESTE MOTIVO DA LUGAR A UNA HELICE TETRACATENARIA DEXTROGIRA EL G-DNA NECESITA PARA SU ESTABILIDAD ELEVADAS CONCENTRACIONES DE NA+ O K+

51
ESTRUCTURAS ORDEN SUPERIOR
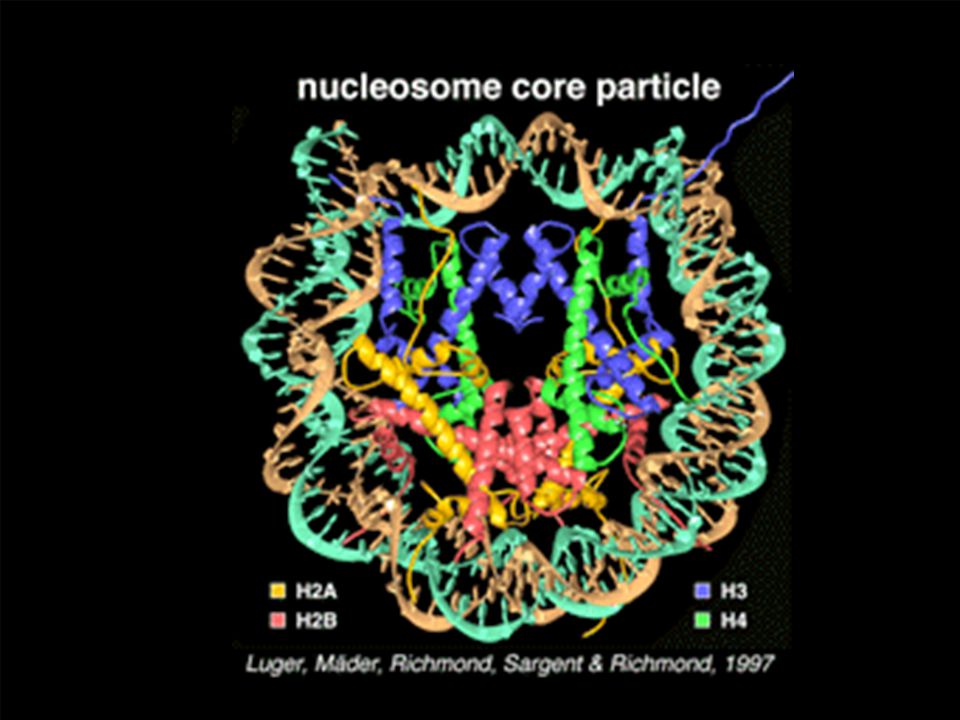
El DNA DEBE COMPACTARSE MUCHO EN LA CELULA ESTO DA LUGAR A ESTRUCTURAS DE ORDEN SUPERIOR LA PRIMERA ES EL NUCLEOSOMA, LA ULTIMA EL CROMOSOMA

52
NUCLEOSOMA

54
Los nucleosomas se ordenan en superfibras
Los nucleosomas forman un rosario que se enrrolla sobre si mismo para dar una superfibra e 30 nm El modelo propuesto es el del solenoide

55
MODELO SOLENOIDE

56
Las fibras de nucleosomas definen el CROMOSOMA
Niveles de estructura-ción en lazos permiten la formación del cromosoma El cromosoma es el nivel máximo de empaquetamiento del DNA.

57
PARA REALIZAR SU FUNCIÓN BIOLÓGICA EL DNA DEBE SER CAPAZ DE DISTORSIONARSE: DOBLARSE, DESENROLLARSE, ALARGARSE,...

58
EL DNA ES DÚCTIL

59
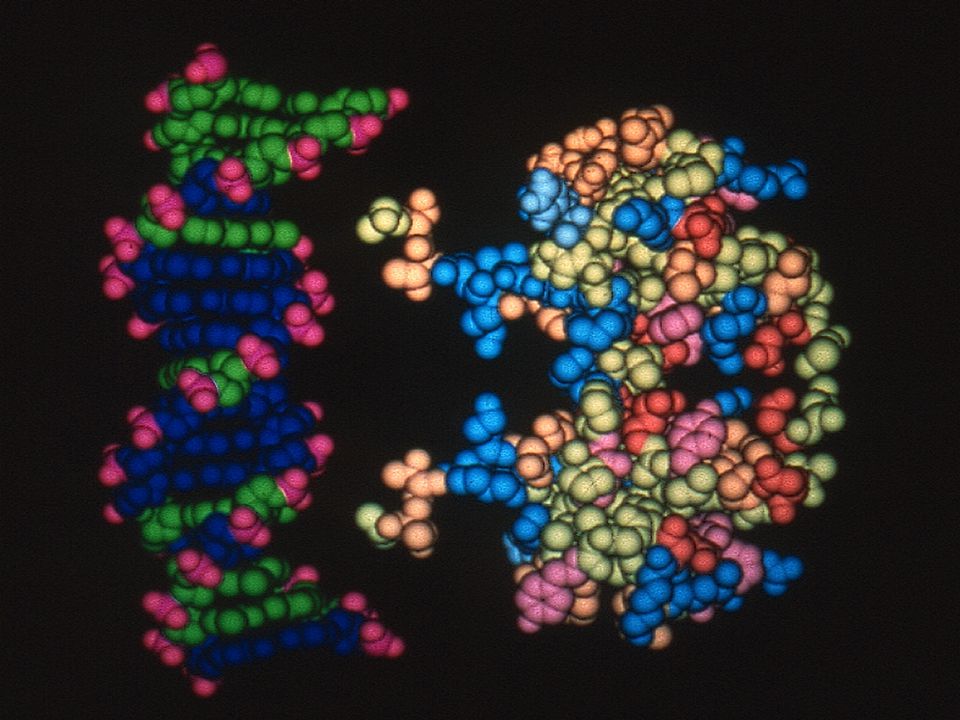
Complejo DNA-UvrB

60
EL DNA ES MOVIL

61
DNA bending Ejemplo de movimiento clave para la función del DNA

62
Estructura de proteínas
Estructura de ácidos nucleicos

63
Secuencia DNA Secuencia Proteína Reconocimiento Molecular Estructura 3D

64
Comunicación entre tejidos
Catálisis enzimática Comunicación entre tejidos Generación y transmisión del impulso nervioso Transporte y almacenaje Permebilidad de membranas Control crecimiento y diferenciación Movimiento Defensa

65
Síntesis “fine chemicals” Aditivos de alimentación
Transformación alimentos Diagnóstico clínico Tratamiento tejidos Terapia Reactivos laboratorio Aditivos limpieza

66
NIVELES DE ESTRUCTURA Estructura primaria (secuencia)
Estructura secundaria (hélices a, cadena b, giros b, lazos,...) Estructura supersecundaria Dominio estructural Estructura terciaria Estructura cuaternaria
 Estructura supersecundaria. Dominio estructural. Estructura terciaria. Estructura cuaternaria.")
67
Aminoácidos hidrofóbicos
Ala (A) Val (V) Leu (L) Ile (I) Met (M) Trp (W) Phe (F) Pro (P)
 Val (V) Leu (L) Ile (I) Met (M) Trp (W) Phe (F) Pro (P)")
68
Aminoácidos polares neutros
Gly (G) Asn (N) Gln (Q) Ser (S) Thr (T) Tyr (Y) Cys (C)
 Asn (N) Gln (Q) Ser (S) Thr (T) Tyr (Y) Cys (C)")
69
Aminoácidos cargados Arg (R) Lys (K) His (H) Glu (E) Asp (D)
 Lys (K) His (H) Glu (E) Asp (D)")
70
El enlace amida define la formación del polímero
+

71
La estructura secundaria es debida a la
ordenación local de residuos contiguos ... ... ... aa1 - aa aa3 - aa4 ... Esto se da por rotación de los enlaces sencillos Y (C-Ca) y F (N-Ca)
 y F (N-Ca)")
72
¿son posibles todas las conformaciones?
Diagrama de Ramachandran

73
¿son posibles todas las conformaciones?
El gráfico de Ramachandran define las estructuras secundarias: repetitivas (hélices o cadenas), y no repetitivas (giros, lazos)
, y no repetitivas (giros, lazos)")
74
Hélice a

75
Estructuras b (paralelas)
")
76
Estructuras b (antiparalelas)
")
77
Estructuras no periódicas giros, lazos

78
Estructura supersecundaria
Agrupaciones sencillas de elementos de estructura secundaria Es un nivel de estructuración local A menudo coincide con el concepto de dominio estructural

79
Dominio estructural Primer nivel de estructura tridimensional con entidad propia Es la unidad básica de plegamiento Puede contener una o varias estructuras supersecundarias

80
alfa-loop-alfa Unión de metales Unión a DNA

81
alfa-loop-alfa

82
alfa-loop-alfa

83
beta - hairpin

84
beta - hairpin

85
Greek key

86
Greek key Este motivo tiene un plegamiento especialmente favorable

87
alfa - beta La hélice se sitúa siempre en el mismo lado de la hoja

88
Agrupaciones todo a

89
Ortogonal b - sandwich

90
b – jelly roll

91
b - barrel Porina

92
a/b barrel

93
a/b doubly wound Rossman fold

94
Estructura terciaria Nivel de estructuración tridimensional global de las proteínas monoméricas Puede contener uno o varios dominios En unión ligera En unión íntima En disposición abrazada

96
Estructura cuaternaria
Nivel de estructuración tridimensional global de las proteínas oligoméricas Es el resultado de agrupar diversas estructuras terciarias Pueden ser temporales o permanentes

97
Fuerzas que estabilizan la estructura de las proteínas
Términos de enlace deben ser favorables (Mínimos en el mapa de Ramachandran) Puentes de hidrógeno (claves a nivel de estructura secundaria) Puentes disulfuro Puentes salinos Interacciones de van der Waals Efecto hidrofóbico
 Puentes de hidrógeno (claves a nivel de estructura secundaria) Puentes disulfuro. Puentes salinos. Interacciones de van der Waals. Efecto hidrofóbico.")
98
El número de estructuras terciarias es enorme
Pero es posible agruparlas estructuralmente

99
Proportion of "new folds" (light blue) and "old folds" (orange) for a given year as a number of protein chain.
 and old folds (orange) for a given year as a number of protein chain.")
100
Jerarquía CATH C: Clase (contenido en estructura secundaria)
A: Arquitectura (disposición de los elementos de estructura secundaria) T: Topología (disposición de las conexiones entre elementos) H: Homología (homología estructural) S: Secuencia (homología de secuencia)
 T: Topología (disposición de las conexiones entre elementos) H: Homología (homología estructural) S: Secuencia (homología de secuencia)")
102
SCOP. Estadísticas Class Number of folds Number of superfamilies
Number of families All alpha proteins 138 224 337 All beta proteins 93 171 276 Alpha and beta proteins (a/b) 97 167 374 Alpha and beta proteins (a+b) 184 263 391 Multi-domain proteins 28 35 Membrane and cell surface proteins 11 17 Small proteins 54 77 116 Total 605 947 1557
 Alpha and beta proteins (a+b) Multi-domain proteins Membrane and cell surface proteins Small proteins Total")
Presentaciones similares




















>")



