Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Metabolismo de Proteínas y Prof. MV Enrique C. ALMIRON
Aminoácidos Prof. MV Enrique C. ALMIRON Bioquímica

3
Introducción Diferencias con glúcidos y lípidos
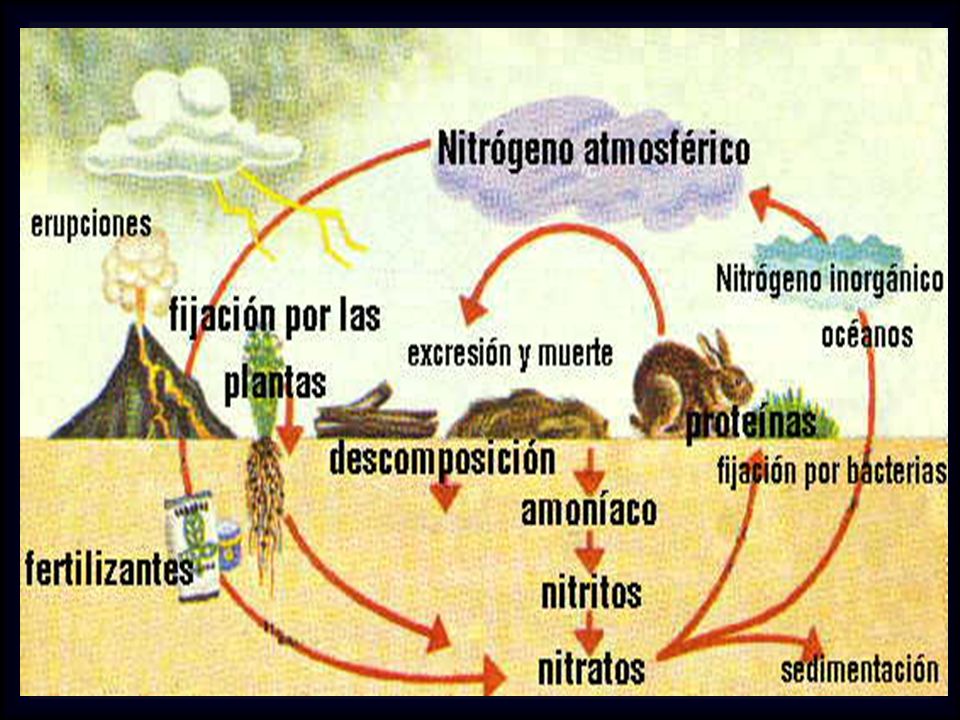
Fuentes de Sustancias Nitrogenadas - Balance Nitrogenado

4
Introducción Balance Nitrogenado
Balance Positivo: la ingesta supera a la pérdida (niños, preñez, etc) Balance Negativo: la pérdida supera la ingesta (cáncer, desnutrición, etc)
 Balance Negativo: la pérdida supera la ingesta (cáncer, desnutrición, etc)")
5
Introducción Diferencias con glúcidos y lípidos
Fuentes de Sustancias Nitrogenadas - Balance Nitrogenado Digestión

6
Introducción Digestión Proteínas AA Aminoácidos Sin modificación AA
Enzimas Hidrolíticas Absorción Intestinal AA (sangre) Proteínas (dieta) AA (tejidos) Sin modificación Transformación Degradación
 Proteínas. (dieta) AA. (tejidos) Sin modificación. Transformación. Degradación.")
7
Introducción Diferencias con glúcidos y lípidos
Fuentes de Sustancias Nitrogenadas - Balance Nitrogenado Digestión Clasificación de los Aminoácidos

8
Introducción Origen Destino de AA Absorción Intestinal AMINOÁCIDOS
Síntesis de aa (hígado) AMINOÁCIDOS Fondo metabólico Común o “pool de aa.” Degradación de Proteínas Tisulares
 AMINOÁCIDOS. Fondo metabólico. Común o pool de aa. Degradación. de Proteínas. Tisulares.")
9
Introducción Origen Destino de AA Síntesis de Proteínas Absorción
AMINOÁCIDOS Fondo metabólico Común o “pool de aa.” Origen Destino de AA Absorción Intestinal Degradación de Proteínas Tisulares Síntesis de aa (hígado) Síntesis de Proteínas Producción de Energía Síntesis de Compuestos Nitrogenados No proteicos
 Síntesis de. Proteínas. Producción. de Energía. Síntesis de. Compuestos. Nitrogenados. No proteicos.")
10
Catabolismo de los Aminoácidos
Separación del grupo amino: Transaminación Desaminación NH3 Tóxico -NH2 Urea Hígado ORINA

11
Catabolismo de los Aminoácidos
R. de Transaminación Transferencia del grupo -amino de un aa a un -cetoácido. aminoácido2 + cetoácido1 à cetoácido2 + aminoácido1 Es una reacción reversible. Las enzimas catalizadoras utilizan piridoxal fosfato como coenzima. Las transaminasas presentan isoenzimas con diferentes localizaciones.

12
Catabolismo de los Aminoácidos

13
Destino final del grupo amino de aminoácidos
Q H I L M S Y W V Cisteína Transaminación -CG + piruvato -cetoácido + alanina Transaminación -CG + oxaloacetato - cetoácido + aspartato + -ceto glutarato - cetoácido + glutamato Desaminación oxidativa - ceto glutarato + NH3 GLUTAMINA UREA

14
Catabolismo de los Aminoácidos
R. de Desaminación de Glutamato El - CG es el sustrato más frecuente en las reacciones de transaminación Glutamato Glutamato: desaminación y oxidación catalizado por la glutamato deshidrogenasa (enzima de la mitocondrial). La misma enzima cataliza la reacción inversa. Esta enzima utiliza NAD o NADP como coenzima Esta reacción provee la mayor parte del NH3 tisular.
. La misma enzima cataliza la reacción inversa. Esta enzima utiliza NAD o NADP como coenzima. Esta reacción provee la mayor parte del NH3 tisular.")
15
Catabolismo de los Aminoácidos

16
Catabolismo de los Aminoácidos

17
Vías Metabólicas del Amoníaco
Principal fuente de NH3: DO de glutamato El Hígado elimina la casi totalidad de NH3 NH4+ + -CG Glutamato + H2O Vías de eliminación de NH3 Sintesis de Glutamina Síntesis de Urea

18
Vías Metabólicas del Amoníaco
Síntesis de Glutamina El NH3 puede ser unido al ácido glutámico por acción de la glutamina sintetasa (enzima mitocondrial). La reacción es prácticamente irreversible. Este mecanismo es especialmente importante en cerebro. La reacción inversa ocurre por acción de la glutaminasa (hígado y riñón) y no es la inversión de la formación de glutamina
. La reacción es prácticamente irreversible. Este mecanismo es especialmente importante en cerebro. La reacción inversa ocurre por acción de la glutaminasa (hígado y riñón) y no es la inversión de la formación de glutamina.")
19
Vías Metabólicas del Amoníaco
Síntesis de Glutamina

20
Vías Metabólicas del Amoníaco
Síntesis de Urea

21
Vías Metabólicas del Amoníaco
Síntesis de Urea CO2 +NH3 +4 ~P (3 ATP) +Aspartato +H2O UREA +2ADP +2Pi +AMP +PPi + Fumarato Los dos nitrógenos de la urea proceden de cualquiera de los aa que participan en las transaminaciones. El NH3 que ingresa en la primera reacción proviene ppalmente de la desaminación oxidativa del glutamato, que a su vez lo adquiere por transferencia de otro aa. El segundo nitrógeno es introducido por el aspartato y puede derivar de los aa que transaminan con oxalacetato.
 +Aspartato +H2O UREA +2ADP +2Pi +AMP +PPi + Fumarato. Los dos nitrógenos de la urea proceden de cualquiera de los aa que participan en las transaminaciones. El NH3 que ingresa en la primera reacción proviene ppalmente de la desaminación oxidativa del glutamato, que a su vez lo adquiere por transferencia de otro aa. El segundo nitrógeno es introducido por el aspartato y puede derivar de los aa que transaminan con oxalacetato.")
22
CICLO DE LA UREA

23
Reacciones: 1. El primer grupo amino que ingresa al ciclo proviene del amoníaco libre intramitocondrial. El amoníaco producido en las mitocondrias, se utiliza junto con el bicarbonato (producto de la respiración celular), para producir carbamoil-fosfato. Reacción dependiente de ATP y catalizada por la carbamoil-fosfato-sintetasa I. Enzima alostérica y modulada (+) por el N-acetilglutamato. 2. El carbamoil-fosfato cede su grupo carbamoilo a la ornitina, para formar citrulina y liberar Pi. Reacción catalizada por laornitina transcarbamoilasa. La citrulina se libera al citoplasma. 3. El segundo grupo amino procedente del aspartato (producido en la mitocondria por transaminación y posteriormente exportado al citosol) se condensa con la citrulina para formar argininosuccinato. Reacción catalizada por laargininosuccinato sintetasa citoplasmática. Enzima que necesita ATP y produce como intermediario de la reacción citrulil-AMP. 4. El argininosuccinato se hidroliza por la arginino succinato liasa, para formar arginina libre y fumarato. El fumarato liberado en una de las reacciones es intermediario del ciclo de Krebs 5. El fumarato ingresa en el ciclo de Krebs y la arginina libre se hidroliza en el citoplasma, por la arginasa citoplasmática para formar urea y ornitina. 6. La ornitina puede ser transportada a la mitocondria para iniciar otra vuelta del ciclo de la urea. En resumen, el ciclo de la urea consta de dos reacciones mitocondriales y cuatro citoplasmáticas .
, para producir carbamoil-fosfato. Reacción dependiente de ATP y catalizada por la carbamoil-fosfato-sintetasa I. Enzima alostérica y modulada (+) por el N-acetilglutamato. 2. El carbamoil-fosfato cede su grupo carbamoilo a la ornitina, para formar citrulina y liberar Pi. Reacción catalizada por laornitina transcarbamoilasa. La citrulina se libera al citoplasma. 3. El segundo grupo amino procedente del aspartato (producido en la mitocondria por transaminación y posteriormente exportado al citosol) se condensa con la citrulina para formar argininosuccinato. Reacción catalizada por laargininosuccinato sintetasa citoplasmática. Enzima que necesita ATP y produce como intermediario de la reacción citrulil-AMP. 4. El argininosuccinato se hidroliza por la arginino succinato liasa, para formar arginina libre y fumarato. El fumarato liberado en una de las reacciones es intermediario del ciclo de Krebs. 5. El fumarato ingresa en el ciclo de Krebs y la arginina libre se hidroliza en el citoplasma, por la arginasa citoplasmática para formar urea y ornitina. 6. La ornitina puede ser transportada a la mitocondria para iniciar otra vuelta del ciclo de la urea. En resumen, el ciclo de la urea consta de dos reacciones mitocondriales y cuatro citoplasmáticas. .")
24
ENERGÉTICA DEL CICLO El ciclo de la urea reúne dos grupos amino y un bicarbonato, para formar una molécula de urea: 1. La síntesis de la urea requiere 4 Pi de alta energía. 2 ATP para formar el carbamoil - P y un ATP para producir argininosuccinato. En la segunda reacción el ATP se hidroliza a AMP y PPi, que puede ser nuevamente hidrolizado para dar 2 Pi. 2. Se ha calculado que los animales ureotélicos pierden cerca del 15% de la energía procedente de los aminoácidos en la producción de urea. 3. Algunos animales compensan esta perdida (bovinos) por transferencia de la urea al rumen, donde los microorganismos la utilizan como fuente de amoníaco para la síntesis de aminoácidos. Este proceso incluso disminuye el consumo de agua. La conexión entre ambos ciclos, de la urea y de los ácidos tricarboxílicos, reducen el coste energético de la síntesis de urea. El ciclo de la urea conlleva la conversión de oxalacetato en fumarato y la posterior conversión del fumarato hasta oxalacetato producirá un NADH, que podrá generar 3 ATP en la respiración mitocondrial, lo que reduce el coste de la síntesis de urea.
 por transferencia de la urea al rumen, donde los microorganismos la utilizan como fuente de amoníaco para la síntesis de aminoácidos. Este proceso incluso disminuye el consumo de agua. La conexión entre ambos ciclos, de la urea y de los ácidos tricarboxílicos, reducen el coste energético de la síntesis de urea. El ciclo de la urea conlleva la conversión de oxalacetato en fumarato y la posterior conversión del fumarato hasta oxalacetato producirá un NADH, que podrá generar 3 ATP en la respiración mitocondrial, lo que reduce el coste de la síntesis de urea.")
25
Vías Metabólicas del Amoníaco
Síntesis de Urea - ENERGÉTICA DEL CICLO El balance energético del ciclo da como resultado el consumo de 4 enlaces de alta energía.

26
Destino del esqueleto carbonado de los aa.
Los aa pueden ser glucogénicos o cetogénicos, ya sea que participen en la síntesis de glucosa o de cuerpos cetónicos. Aminoácido cetogénicos: acetil-CoA, acetoacetato. - Aminoácido glucogénicos: piruvato e intermediarios del C.A.C.. dan lugar a glucosa. Casi todos los aa no esenciales son glucogénicos, por el contrario, casi todos los aa esenciales son cetogénicos

27
Ciclo de Krebs

28
CONEXIÓN ENTRE LOS CICLOS DE LA UREA Y DE KREBS

29
REGULACIÓN DEL CICLO La enzima carbamoil-fosfato-sintetasa I es activada alostéricamente por el N - acetilglutamato que se sintetiza a partir del acetil-CoA y el glutamato, por la N-acetilglutamato sintetasa; enzima que, a su vez, es activada por la arginina, aminoácido que se acumula cuando la producción de urea es lenta. En individuos con deficiencias congénitas de enzimas del ciclo, distintas a la arginasa, el sustrato correspondiente se acumula, lo que provoca un aumento de la velocidad de la reacción deficiente, por lo que la velocidad del ciclo se mantiene baja. No obstante se producen acumulaciones de los sustratos precedentes, hasta el amoniaco, lo que causa finalmente una hiperamonemia. El cerebro es particularmente sensible a las [ ] elevadas de amonio.

30
REGULACIÓN DEL CICLO

31
TRANSPORTE DEL GRUPO AMINO DESDE LOS TEJIDOS HACIA EL HÍGADO

32
Transporte desde el músculo:CICLO ALANINA-GLUCOSA

33
El hábitat natural determina la ruta de excreción del nitrógeno
La síntesis de urea no es la única ruta, ni siquiera la más común, para excretar el amoníaco. La base para las diferentes formas moleculares en que se excretan los grupos amino se encuentra en la anatomía y en la fisiología de los organismos en relación con su hábitat natural. En los peces óseos (animales amonotélicos), el hígado es el lugar principal del catabolismo de los aminoácidos. El amonio producido por transdesaminación se libera simplemente del hígado a la sangre para su transporte a las branquias, y es rápidamente eliminado de la sangre a medida que el agua pasa a través de las branquias. Así pues, los peces óseos no requieren un complejo sistema urinario. Los que convierten el nitrógeno amínico en purinas, las cuales se catabolizan a ácido úrico, un compuesto bastante insoluble que se excreta en las heces en forma de masa semisólida de cristales de ácido úrico. Para tener la ventaja de excretar el nitrógeno amínico en esta forma, las aves y reptiles llevan a cabo un trabajo metabólico considerable; la ruta desde los grupos amino de los aminoácidos a las purinas y al ácido úrico es un proceso complejo y que requiere energía.
, el hígado es el lugar principal del catabolismo de los aminoácidos. El amonio producido por transdesaminación se libera simplemente del hígado a la sangre para su transporte a las branquias, y es rápidamente eliminado de la sangre a medida que el agua pasa a través de las branquias. Así pues, los peces óseos no requieren un complejo sistema urinario. Los que convierten el nitrógeno amínico en purinas, las cuales se catabolizan a ácido úrico, un compuesto bastante insoluble que se excreta en las heces en forma de masa semisólida de cristales de ácido úrico. Para tener la ventaja de excretar el nitrógeno amínico en esta forma, las aves y reptiles llevan a cabo un trabajo metabólico considerable; la ruta desde los grupos amino de los aminoácidos a las purinas y al ácido úrico es un proceso complejo y que requiere energía.")
34
Otros mecanismos generales del metabolismo de aa
Descarboxilación: utiliza enzimas que se denominan descarboxilasas cuya coenzima es el fosfato de piridoxal. Genera aminas biogénas y también a los productos de los procesos de putrefacción de proteínas. Transferencia de restos monocarbonados: el donante de metilos es la metionina, a través de la SAM (S-adenosil metionina). Otros transportadores de grupos monocarbonados son el ácido THF y la Biotina Transpeptidación: :utiliza transferasas, las cuales transfieren aminoácidos entre oligopéptidos.
. Otros transportadores de grupos monocarbonados son el ácido THF y la Biotina. Transpeptidación: :utiliza transferasas, las cuales transfieren aminoácidos entre oligopéptidos.")
35
Clasificación de Seres Vivos
en base al producto final del metabolismo Proteico Ureotélicos Amoniotélicos Uricotélicos

Presentaciones similares