Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Proteínas de transporte. Proteínas nutrientes y de reserva.
TEMA 1. INTRODUCCIÓN A LAS PROTEÍNAS, FUNCIONES: Enzimas. Proteínas de transporte. Proteínas nutrientes y de reserva. Proteínas contráctiles o motiles. Proteínas estructurales. Proteínas de defensa. Proteínas reguladoras. Otras proteínas.

2
TEMA 1. INTRODUCCIÓN A LAS PROTEÍNAS
Muchas proteínas contienen sólo residuos de aminoácidos y ningún otro grupo químico adicional; a este tipo de proteínas se las considera proteínas simples. Sin embargo, algunas proteínas contienen componentes químicos diferentes; estas proteínas se denominan proteínas conjugadas. La parte no aminoácido de una proteína conjugada se denomina habitualmente grupo prostético. TABLA Proteínas conjugadas. Clase Grupo prostético Ejemplo Lipoproteínas Lípidos Lipoproteína b1 de la sangre Glicoproteínas Glícidos Inmunoglobulina G Fosfoproteínas Grupos fosfatos Caseína de la leche Hemoproteínas Hemo Hemoglobina Flavoproteínas Nucleótidos de flavina Succinato deshidrogenasa Metaloproteínas Hierro Ferritina Zinc Alcohol deshidrogenasa Calcio Calmodulina Molibdeno Dinitrogenasa Cobre Plastocianina

3
Estructura general de un aminoácido.
Grupo carboxilo Grupo amino Grupo de cadena lateral Átomo de carbono a Esta estructura es común a todos los a-aminoácidos. El grupo R o cadena lateral (en rojo) unido al carbono a (azul) es diferente para cada aminoácido.
 unido al carbono a (azul) es diferente para cada aminoácido.")
4
A) B) Estereoisomería de los aminoácidos. A) El átomo de carbono a es asimétrico (excepto en la glicina) por lo que existen isómeros especulares (o estereoisó-meros) D y L. B) Convención de Fisher.
 El átomo de carbono a es asimétrico (excepto en la glicina) por lo que existen isómeros especulares (o estereoisó-meros) D y L. B) Convención de Fisher.")
5
Los aminoácidos en disolución, a pH neutro, son predominantemente iones dipolares (zwitteriones) en vez de moléculas no iónicas. En la forma dipolar de un aminoácido el grupo amino está protonizado (–) y el grupo carboxilo está disociado (–COO-). El estado de ionización de un aminoácido varía con el pH.
 y el grupo carboxilo está disociado (–COO-). El estado de ionización de un aminoácido varía con el pH..")
6
Aminoácidos con cadenas apolares alifáticas.

7
Aminoácidos con cadena lateral aromática

8
Aminoácidos con cadenas laterales sin carga

9
Aminoácidos con cadena lateral cargada positivamente:
Aminoácidos con cadena lateral cargada negativamente:

11
Propiedades ácido-base de los aminoácidos.
Cuando se disuelve en agua un aminoácido tal como la alanina, se encuentra en solución en forma de ion dipolar, o zwitterion, el cual puede actuar como ácido (dador de protones): o como base (aceptor de protones): Las sustancias con esta naturaleza dual son anfóteras llamándoselas a menudo anfolitos, de "electrólitos anfóteros" Punto isoeléctrico: valor de pH al cual la carga neta es cero.
: o como base (aceptor de protones): Las sustancias con esta naturaleza dual son anfóteras llamándoselas a menudo anfolitos, de electrólitos anfóteros Punto isoeléctrico: valor de pH al cual la carga neta es cero.")
12
Ecuación de Henderson-Hasselbach
pK1 – carboxilo (-COOH); pKR – imidazol (-NH) (R); pK2 – amino (- NH2) pK1 – carboxilo (-COOH); pKR – carboxilo (-COOH) (R); pK2 – amino (- NH2) Ecuación de Henderson-Hasselbach
; pKR – imidazol (-NH) (R); pK2 – amino (- NH2) pK1 – carboxilo (-COOH); pKR – carboxilo (-COOH) (R); pK2 – amino (- NH2) Ecuación de Henderson-Hasselbach.")
13
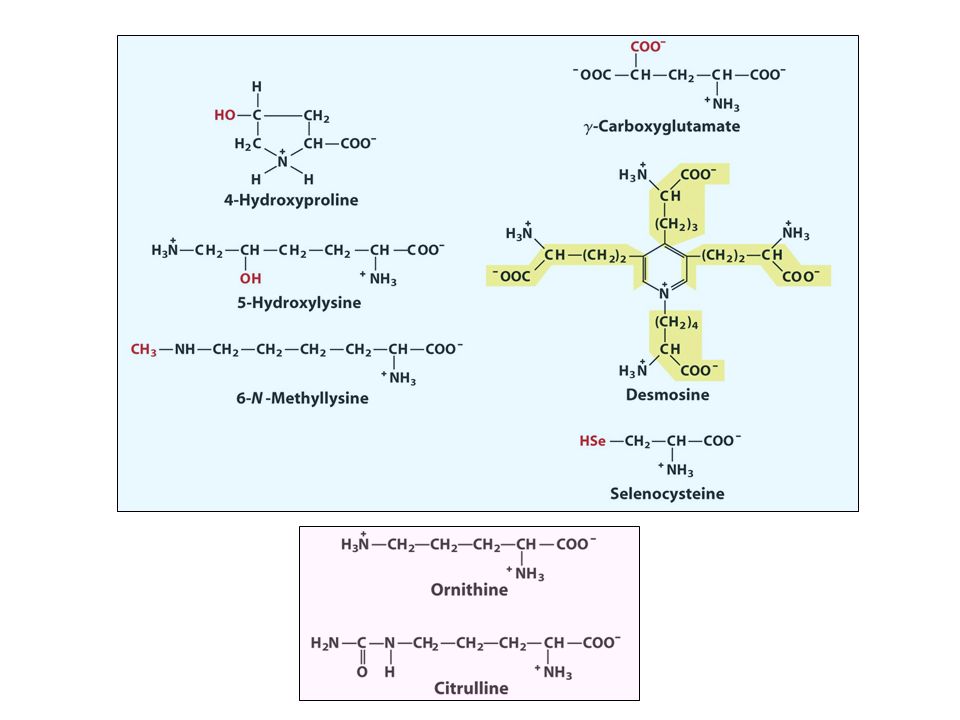
Propiedades y nomenclatura aminoácidos estándar

14
Niveles estructurales de las proteínas.
Las proteínas están constituidas por secuencia de aminoácidos, unidos mediante enlaces peptídicos. Cada aminoácido en la secuencia se denomina residuo. Estructura primaria: La secuencia lineal de aminoácidos que componen una proteína unidos entre sí por enlaces peptídicos. Estructura secundaria: disposición regular y repetitiva en el espacio de los residuos de aminoácido adyacentes en la cadena polipeptídica. Estructura terciaria: estructura tridimensional completa y biológicamente activa de un polipéptido. Incluye la relación espacial entre todos los aminoácidos de un polipéptido (adyacentes o alejados -interacciones de largo alcance). Estructura cuaternaria: relación espacial entre los polipéptidos o subunidades que constituyen una proteína oligomérica. Las proteínas con una sola cadena polipeptídica carecen de estructura cuaternaria.
. Estructura cuaternaria: relación espacial entre los polipéptidos o subunidades que constituyen una proteína oligomérica. Las proteínas con una sola cadena polipeptídica carecen de estructura cuaternaria.")
15
Formación de un enlace peptídico (sombreado en gris) en un dipeptido
Formación de un enlace peptídico (sombreado en gris) en un dipeptido. Es ésta una reacción de condensación. El grupo a-amino del aminoácido que actúa como nucleófilo desplazando el grupo hidroxilo del grupo carboxilo del aminoácido 1 (rojo). Los grupos amino son buenos nucleófilos, pero el grupo hidroxilo es un grupo saliente pobre por lo que no es desplazado fácilmente. A pH fisiológico la reacción, tal como se muestra, no tiene lugar de forma apreciable. La formación del enlace peptídico es endergónica con una variación de energía libre de alrededor de +21 kJ/mol.
 en un dipeptido. Es ésta una reacción de condensación. El grupo a-amino del aminoácido que actúa como nucleófilo desplazando el grupo hidroxilo del grupo carboxilo del aminoácido 1 (rojo). Los grupos amino son buenos nucleófilos, pero el grupo hidroxilo es un grupo saliente pobre por lo que no es desplazado fácilmente. A pH fisiológico la reacción, tal como se muestra, no tiene lugar de forma apreciable. La formación del enlace peptídico es endergónica con una variación de energía libre de alrededor de +21 kJ/mol.")
16
Punto isoeléctrico y carga de las proteínas
Punto isoeléctrico (pI): valor de pH al cual la proteína posee carga neta cero ( cargas positivas = cargas negativas). pH = pI carga neta cero pH < pI carga neta positiva pH > pI carga neta negativa
: valor de pH al cual la proteína posee carga neta cero ( cargas positivas = cargas negativas). pH = pI carga neta cero. pH < pI carga neta positiva. pH > pI carga neta negativa.")
17
Propiedades del enlace peptídico
1. El enlace peptídico tiene carácter de doble enlace debido a que presenta estructuras resonantes. El híbrido de resonancia resultante posee 40% de doble enlace. Orbitales p

18
Propiedades del enlace peptídico (cont.)
2. El enlace peptídico tiene carácter polar. El oxígeno es más electronegativo que el nitrógeno por lo que atrae a los electrones compartidos dando lugar a un reparto desigual de los electrones deslocalizados. El oxígeno tiene una carga parcial negativa y el nitrógeno tiene una carga parcial positiva 3. El enlace peptídico es plano. el carácter parcial de doble enlace del enlace peptídico evita la libre rotación en el enlace amida C-Na y coloca a los seis átomos del grupo peptídico en el mismo plano.

19
Las consideraciones anteriores son importantes porque indican que el esqueleto de una proteína es una secuencia ligada de grupos peptídicos planos rígidos La rigidez de los enlaces peptídicos limita el número de conformaciones que puede adoptar una cadena polipeptídica.

20
Ángulos de torsión del enlace peptídico
Como se ha mencionado anteriormente no existe rotación en el enlace peptídico pero es posible la rotación alrededor de los enlaces Ca-C y N-Ca. Los ángulos de rotación o de torsión de los enlaces se denominan por convenio y (psi) para el enlace Ca-C y f (phi) para el enlace N-Ca(A). Por convenio, los ángulos y y f valen 0º cuando los dos enlaces peptídicos conectados a un mismo Ca están en un mismo plano (B). En principio, ambos ángulos pueden tener valores entre –180º y +180º. Positivo si el ángulo resultante se forma por una rotación en el sentido de las agujas del reloj (en el caso de f cuando el giro en el sentido de las agujas del reloj lo efectúa el grupo peptídico que precede al enlace Ca-N, visto desde el Ca al N1; en el caso y el grupo peptídico siguiente alrededor del los Ca-C2, visto desde el Ca al C2); negativo si el giro se efectúa en contra de las agujas del reloj. En principio, f y y pueden adoptar cualquier valor entre –180º y +180º, pero muchos de estos valores no son accesibles a causa de las interferencias estéricas entre átomos del esqueleto polipeptídico y de las cadenas laterales de los aminoácidos. La conformación en la que f y valen 0º es una de las inaccesibles; este valor se usa meramente como referencia B
 para el enlace Ca-C y f (phi) para el enlace N-Ca(A). Por convenio, los ángulos y y f valen 0º cuando los dos enlaces peptídicos conectados a un mismo Ca están en un mismo plano (B). En principio, ambos ángulos pueden tener valores entre –180º y +180º. Positivo si el ángulo resultante se forma por una rotación en el sentido de las agujas del reloj (en el caso de f cuando el giro en el sentido de las agujas del reloj lo efectúa el grupo peptídico que precede al enlace Ca-N, visto desde el Ca al N1; en el caso y el grupo peptídico siguiente alrededor del los Ca-C2, visto desde el Ca al C2); negativo si el giro se efectúa en contra de las agujas del reloj. En principio, f y y pueden adoptar cualquier valor entre –180º y +180º, pero muchos de estos valores no son accesibles a causa de las interferencias estéricas entre átomos del esqueleto polipeptídico y de las cadenas laterales de los aminoácidos. La conformación en la que f y valen 0º es una de las inaccesibles; este valor se usa meramente como referencia. B.")
21
(a) Cada una de las estructuras secundarias posibles queda completamente descrita mediante los ángulos de enlace f y y, que se repiten en cada residuo. Los valores permitidos para f y y pueden visualizarse gráficamente representando f frente a y, en lo que se denomina la representación de Ramachandran. Esta representación visualiza los ángulos f y y estéricamente posibles en base a modelos moleculares que asumen que los átomos son esferas rígidas con radio de Van der Waals. (b) (a) Representación de Ramachandran para los residuos de L-Ala. (b) Isómeros de prolina.
 (a) Representación de Ramachandran para los residuos de L-Ala. (b) Isómeros de prolina.")
23
Determinación de la secuencia de aminoácidos de una proteína (estructura primaria).
1. Determinación del residuo amino terminal. Método de Sanger:

24
1. Determinación del residuo amino terminal (cont)
1. Determinación del residuo amino terminal (cont). Degradación de Edman: Aminoácido PTH
. Degradación de Edman: Aminoácido PTH.")
25
2. Identificación del residuo carboxilo-terminal:
Carboxipeptidasas Método de la hidracina

26
3. Rotura de los puentes disulfuro

27
4. Determinación de la composición de aminoácidos de la proteína.
5. Ruptura de la cadena polipéptidica TABLA Especificidad de algunos métodos usuales de fragmentación de cadenas polipeptídicas. Tratamiento Puntos de corte* Tripsina Lys, Arg (C) Proteasa de Submaxillarus Arg (C) Quimotripsina Phe, Trp, Tyr (C) Proteasa V8 de Staphylococcus aureus Asp, Glu (C) Proteasa Asp-N Asp, Glu (N) Pepsina Phe, Trp, Tyr (N) Endoproteinasa Lys C Lys (C) Bromuro de cianógeno Met (C) *Residuos que aportan el punto de reconocimiento primario de la proteasa o el reactivo; la ruptura del enlace peptídico tiene lugar en el lado carbonilo (C) o amino (N) del residuo aminoácido indicado
 Proteasa de Submaxillarus. Arg (C) Quimotripsina. Phe, Trp, Tyr (C) Proteasa V8 de Staphylococcus aureus. Asp, Glu (C) Proteasa Asp-N. Asp, Glu (N) Pepsina. Phe, Trp, Tyr (N) Endoproteinasa Lys C. Lys (C) Bromuro de cianógeno. Met (C) *Residuos que aportan el punto de reconocimiento primario de la proteasa o el reactivo; la ruptura del enlace peptídico tiene lugar en el lado carbonilo (C) o amino (N) del residuo aminoácido indicado.")
28
6. Secuenciación de los fragmentos peptídicos.
7. Ordenación de los fragmentos peptídicos. 8. Ruptura de la cadena polipéptidica.

29
La secuencia de aminoácidos de una proteína o estructura primaria tiene gran importancia en bioquímica porque aporta información sobre: 1. Aporta información sobre su localización celular y función biológica. 2. Ayuda a entender la estructura tridimensional de una proteína y las reglas que gobiernan el plegamiento de las proteínas. 3. Contribuye al diagnóstico y conocimientos de muchas enfermedades. 4. El estudio de la secuencia de aminoácidos de proteínas homólogas de distintas especies permite determinar las rela-ciones filogenéticos de dife-rentes organismos.

Presentaciones similares