Descargar la presentación
La descarga está en progreso. Por favor, espere

1
TRANSFERENCIA DE LA INFORMACIÓN GENÉTICA: TRANSCRIPCIÓN
Se denomina transcripción al proceso de trasvase de información, contenida en el ADN, a una molécula de ARN. Constituye el primer paso en la expresión de los genes y mediante esta ruta se sintetizan todos los tipos de ARN que existen en las células. A primera vista, las cadenas de ARN y ADN pueden parecer similares, con un grupo OH en la posición 2´de la pentosa y la sustitución de la T por U como únicas diferencias. Sin embargo, al contrario del ADN, la mayoría de los ARN son de cadena sencilla. Estas cadenas se pliegan sobre sí mismas, dando lugar a una diversidad estructural mucho más amplia que la observada en el ADN, gracias a la cual el ARN es capaz de asumir una amplia variedad de funciones celulares.

2
CARACTERÍSTICAS GENERALES DE LA TRANSCIPCIÓN
Desde el punto de vista del mecanismo, la transcripción es semejante a la replicación del ADN. Es una reacción de polimerización, se utilizan sustratos activados (nucleósidos trifosfato), se necesita un molde (de ahí el nombre de síntesis de ARN dependiente de ADN), la dirección de síntesis es también 5´→ 3´ y transcurre en 3 etapas: iniciación, elongación y terminación. Existen también una serie de características que diferencian ambos procesos: La transcripción es un proceso monocatenario. Con pocas excepciones, sólo se transcribe una de las cadenas del ADN. La situación de los genes a copiar puede localizarse en cualquiera de las dos cadenas del ADN. Por convenio, a la cadena utilizada como molde se la llama hebra molde, antisentido, no codificante o hebra (-) y a la cadena complementaria se la denomina hebra no molde, con sentido, codificadora o hebra (+). La doble cadena se escribe de izquierda a derecha en el sentido 5´→ 3´ de la hebra codificadora. La hebra codificadora del ADN tiene la misma secuencia que la cadena de ARN transcrito excepto que la timina sustituye al uracilo (Figura 1).
, se necesita un molde (de ahí el nombre de síntesis de ARN dependiente de ADN), la dirección de síntesis es también 5´→ 3´ y transcurre en 3 etapas: iniciación, elongación y terminación. Existen también una serie de características que diferencian ambos procesos: La transcripción es un proceso monocatenario. Con pocas excepciones, sólo se transcribe una de las cadenas del ADN. La situación de los genes a copiar puede localizarse en cualquiera de las dos cadenas del ADN. Por convenio, a la cadena utilizada como molde se la llama hebra molde, antisentido, no codificante o hebra (-) y a la cadena complementaria se la denomina hebra no molde, con sentido, codificadora o hebra (+). La doble cadena se escribe de izquierda a derecha en el sentido 5´→ 3´ de la hebra codificadora. La hebra codificadora del ADN tiene la misma secuencia que la cadena de ARN transcrito excepto que la timina sustituye al uracilo (Figura 1).")
3
Es un proceso selectivo
Es un proceso selectivo. La transcripción se limita a una porción del ADN. En una célula eucariota diferenciada, la parte del ADN total que se transcribe es muy pequeña. Incluso en los organismos unicelulares, en los que prácticamente todas las secuencias del ADN pueden transcribirse, en un momento dado se transcribe mucho menos de la mitad de los genes. En consecuencia, gran parte del interés por la transcripción se centra en los mecanismos utilizados para seleccionar determinados genes y cadenas molde, puesto que esta selección es la que gobierna en gran medida las capacidades metabólicas de una célula. La transcripción es un proceso reiterativo, puede repetirse infinidad de veces durante la vida celular. A este respecto, existe una gran variabilidad en cuanto a la frecuencia de transcripción de distintas regiones del ADN. Es un proceso conservador. No afecta a la estructura del ADN. No requiere cebador.

4
ARN POLIMERASAS El descubrimiento de la ADNp y su dependencia de un molde de ADN fue un estímulo para la búsqueda de una enzima que sintetizase ARN complementario de un molde de ADN. Hacia 1960, cuatro grupos de investigación independientes habían detectado una enzima en extractos celulares capaz de formar un polímero de ARN a partir de ribonucleótidos 5´-trifosfato. Investigaciones posteriores con la ARN polimerasa (ARNp) purificada de E. coli contribuyeron a definir las propiedades fundamentales de la transcripción. La ARNp realiza múltiples funciones en la transcripción: Busca en el ADN los puntos de iniciación, también llamados secuencias promotoras o, simplemente, promotores. Desenrolla una parte de la doble hélice del ADN para producir un molde de ADN de una sola hebra. Selecciona el ribonucleótido trifosfato correcto y cataliza la formación del enlace fosfodiéster. Este proceso se repite tantas veces como la enzima se desplace unidireccionalmente a lo largo del molde de ADN. A este respecto, la ARNp es totalmente procesiva: una única molécula de ARNp realiza la transcripción desde el inicio hasta el final. Detecta las señales de terminación que indican dónde finaliza la transcripción. Interacciona con las proteínas activadoras y represoras (factores de transcripción) que modulan en un amplio intervalo la velocidad de transcripción.
 purificada de E. coli contribuyeron a definir las propiedades fundamentales de la transcripción. La ARNp realiza múltiples funciones en la transcripción: Busca en el ADN los puntos de iniciación, también llamados secuencias promotoras o, simplemente, promotores. Desenrolla una parte de la doble hélice del ADN para producir un molde de ADN de una sola hebra. Selecciona el ribonucleótido trifosfato correcto y cataliza la formación del enlace fosfodiéster. Este proceso se repite tantas veces como la enzima se desplace unidireccionalmente a lo largo del molde de ADN. A este respecto, la ARNp es totalmente procesiva: una única molécula de ARNp realiza la transcripción desde el inicio hasta el final. Detecta las señales de terminación que indican dónde finaliza la transcripción. Interacciona con las proteínas activadoras y represoras (factores de transcripción) que modulan en un amplio intervalo la velocidad de transcripción.")
5
La química de la síntesis del ARN tiene mucho en común con la síntesis de ADN. La ARNp elonga una cadena de ARN por adición de ribonucleótidos al extremo 3´-OH de la cadena y sintetiza el ARN en dirección 5´→3´. El 3´-OH actúa como nucleófilo, atacando el fosfato del ribonucleótido trifosfato entrante y liberando pirofosfato (Figura 2). La reacción global es: (NMP)n + NTP → (NMP)n+1 + PPi ARN ARN alargado
n + NTP → (NMP)n+1 + PPi. ARN ARN alargado.")
6
La ARNp requiere ADN para su actividad, siendo más activa con un molde de ADN bicatenario. La hebra molde es copiada en la dirección 3´→5´ (antiparalela a la nueva cadena de ARN) al igual que en al replicación. Cada nucleótido en el ARN recién formado es seleccionado según las interacciones de apareamiento de bases de Watson y Crick; en este caso los residuos de uridilato se insertan en el ARN para aparearse con residuos adenilato del ADN molde. La geometría de los pares de bases también puede jugar un papel en la selección de las bases, tal y como hemos visto en la replicación. A diferencia de la ADNp, la ARNp no necesita un cebador para iniciar la síntesis. El grupo 5´-trifosfato del primer residuo de una cadena de nueva síntesis no es cortado para liberar PPi, sino que se mantiene intacto durante toda la transcripción. La ARNp dependiente de ADN de E. coli es una enzima compleja y de gran tamaño formada por 6 subunidades de 5 tipos distintos (2´). El núcleo de la enzima está constituido por 2´ y presenta la actividad catalítica. La subunidad se une transitoriamente al núcleo y dirige a la enzima hacia sitios específicos de unión en el ADN (promotores), participa en la iniciación de la síntesis y luego se disocia del resto de la enzima. Estas 6 subunidades contituyen la holoenzima ARNp. (Figura 3 y Tabla 1) Tabla 1. Composición de subunidades de la ARN polimerasa de E. coli
. El núcleo de la enzima está constituido por 2´ y presenta la actividad catalítica. La subunidad se une transitoriamente al núcleo y dirige a la enzima hacia sitios específicos de unión en el ADN (promotores), participa en la iniciación de la síntesis y luego se disocia del resto de la enzima. Estas 6 subunidades contituyen la holoenzima ARNp. (Figura 3 y Tabla 1) Tabla 1. Composición de subunidades de la ARN polimerasa de E. coli.")
7
Las ARNp carecen de actividad exonucleasa 3´→5´correctora de pruebas
Las ARNp carecen de actividad exonucleasa 3´→5´correctora de pruebas. En consecuencia, durante la transcripción se produce un error por cada 104 o 105 ribonucleótidos incorporados en el ARN. Dado que de un solo gen se producen muchas copias de ARN y que todos los ARN son finalmente degradados y reemplazados, un error en una molécula de ARN tiene menos consecuencias para la célula que un error en la información permanente almacenada en el ADN.

8
ETAPAS DE LA TRANSCRIPCIÓN EN PROCARIOTAS: INICIACIÓN, ELONGACIÓN Y TERMINACIÓN
La fase de iniciación comienza en los promotores del molde de ADN. Los promotores son secuencias de ADN que dirigen a la ARNp hacia secuencias específicas adyacentes para iniciar la transcripción. Cuando se comparan las secuencias de muchos promotores de procariotas se pone en evidencia un patrón llamativo (Figura 4). Existen dos tramos de secuencias consensos situados a 10 y 35 pb antes del inicio de la transcripción. Estas denominaciones hacen referencia a la hebra codificadora de ADN, aunque la hebra molde o no codificadora de ADN es la que se transcribe en ARN. Por convenio, se da el número +1 al par de bases en el que comienza la síntesis de ARN, y +n será el último par de bases donde acaba la síntesis. Este último par de bases estará hacia el extremo 3´ de la cadena codificadora, por lo que se dice que el movimiento de la ARNp en esta zona es “aguas abajo”. La secuencia promotora, situada hacia el extremo 5´ de la cadena codificadora, se dice que está “aguas arriba”, y se expresa -1 a –n. Las secuencias promotoras consenso están a situadas a -10 (caja TATA o de Pribnow) y -35, ya que se localizan hacia los nucleótidos 10 y 35 antes del punto de iniciación. Cada una de estas secuencias tiene una longitud de 6 pares de bases. A partir del análisis de muchos promotores se han deducido sus secuencias consenso (promedio), estas son: 5'-TTGACA TATAAT-----Punto de partida para la transcripción
. Existen dos tramos de secuencias consensos situados a 10 y 35 pb antes del inicio de la transcripción. Estas denominaciones hacen referencia a la hebra codificadora de ADN, aunque la hebra molde o no codificadora de ADN es la que se transcribe en ARN. Por convenio, se da el número +1 al par de bases en el que comienza la síntesis de ARN, y +n será el último par de bases donde acaba la síntesis. Este último par de bases estará hacia el extremo 3´ de la cadena codificadora, por lo que se dice que el movimiento de la ARNp en esta zona es aguas abajo . La secuencia promotora, situada hacia el extremo 5´ de la cadena codificadora, se dice que está aguas arriba , y se expresa -1 a –n. Las secuencias promotoras consenso están a situadas a -10 (caja TATA o de Pribnow) y -35, ya que se localizan hacia los nucleótidos 10 y 35 antes del punto de iniciación. Cada una de estas secuencias tiene una longitud de 6 pares de bases. A partir del análisis de muchos promotores se han deducido sus secuencias consenso (promedio), estas son: TTGACA TATAAT-----Punto de partida para la transcripción.")
9
Figura 4. Típicos promotores de E
Figura 4. Típicos promotores de E. coli reconocidos por el holoenzima de la ARN polimerasa que contiene 70. Se muestran las secuencias de la hebra no molde que son leídas en la dirección 3' 5', como se acostumbra en representaciones de este tipo. Las secuencias varían de un promotor a otro, pero la comparación de muchos promotores pone de manifiesto similitudes, particularmente en las regiones -10 y -35. La secuencia consenso de los promotores reconocidos por 70 es la segunda por arriba. Las regiones espaciadoras contienen un número variable de nucleótidos (N). Sólo se muestra el primer nucleótido que codifica el transcrito de ARN (en la posición +1).
. Sólo se muestra el primer nucleótido que codifica el transcrito de ARN (en la posición +1).")
10
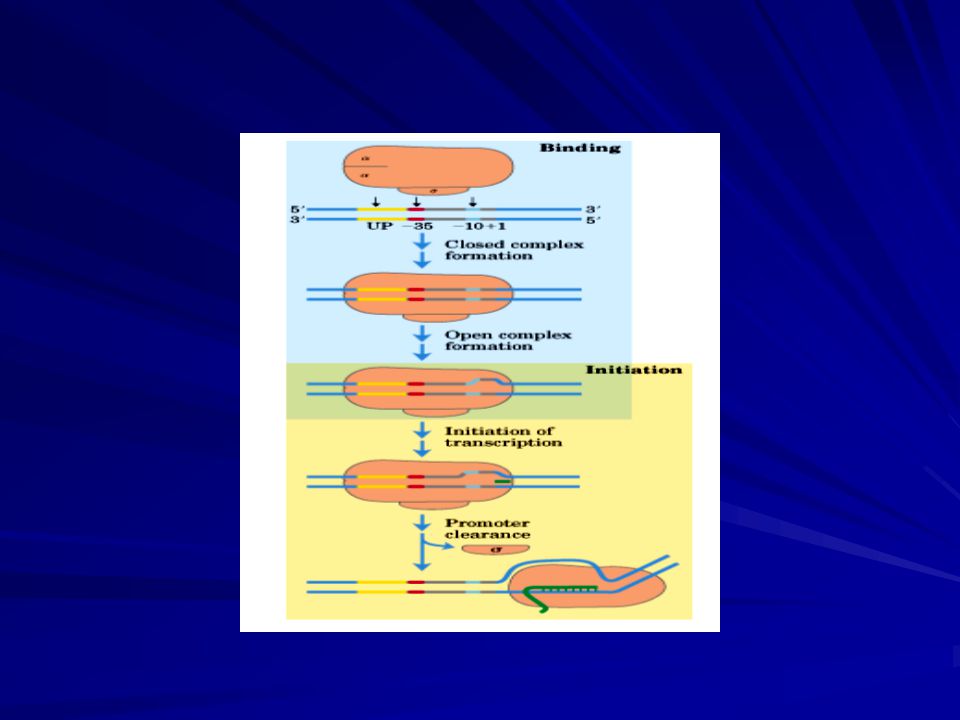
INICIACIÓN Los promotores difieren ampliamente en su eficacia. Los genes con promotores muy activos se transcriben con mucha frecuencia, tan a menudo como cada 2 segundos en E. coli, en tanto que los genes con promotores poco activos pueden transcribirse una vez cada 10 minutos o más. Las secuencias -10 y -35 de los promotores activos tienen secuencias muy similares a las secuencias consenso, en tanto que los promotores poco activos suelen presentar múltiples sustituciones en estas secuencias. En efecto, la mutación de sólo una base tanto en la secuencia -10 como en la secuencia -35 puede variar la actividad del promotor. La separación entre estas secuencias consenso también es importante: la distancia óptima es de 17 nucleótidos. Por tanto, la eficiencia o actividad de una secuencia promotora sirve para regular la transcripción. Las proteínas reguladoras que se unen a secuencias específicas próximas a los promotores y que interaccionan con la ARN polimerasa ejercen también una marcada influencia sobre la frecuencia de transcripción de muchos genes. El núcleo 2´ de la ARNp es incapaz de iniciar la transcripción en las secuencias promotoras. Más bien, el holoenzima completo 2´ es indispensable para la iniciación en el punto de partida correcto. La subunidad contribuye concretamente a la iniciación de dos maneras. 1º) Disminuye la afinidad de la ARN polimerasa por las regiones generales del ADN. En su ausencia, el núcleo del enzima se une al ADN fuerte e indiscriminadamente. 2º) La subunidad permite a la ARN polimerasa el reconocimiento de las secuencias promotoras. Se ha encontrado que un amplio fragmento de la subunidad presenta en su superficie una hélice ; ésta hélice se ha asociado con el reconocimiento de la secuencia de la región -10 (Figura 6). El holoenzima se une a la doble hélice del ADN y se desplaza a lo largo de ella en busca del promotor, haciendo una búsqueda unidimensional utilizando el ADN como si fuesen los raíles de un tren. La búsqueda es rápida ya que la ARN polimerasa se desliza a lo largo del ADN en vez de asociarse y disociarse. La subunidad se libera cuando la cadena naciente de ARN tiene una longitud de 9 ó 10 nucleótidos. Tras esta liberación, la subunidad puede colaborar en la iniciación con otro núcleo enzimático. (Figura 7)
 Disminuye la afinidad de la ARN polimerasa por las regiones generales del ADN. En su ausencia, el núcleo del enzima se une al ADN fuerte e indiscriminadamente. 2º) La subunidad permite a la ARN polimerasa el reconocimiento de las secuencias promotoras. Se ha encontrado que un amplio fragmento de la subunidad presenta en su superficie una hélice ; ésta hélice se ha asociado con el reconocimiento de la secuencia de la región -10 (Figura 6). El holoenzima se une a la doble hélice del ADN y se desplaza a lo largo de ella en busca del promotor, haciendo una búsqueda unidimensional utilizando el ADN como si fuesen los raíles de un tren. La búsqueda es rápida ya que la ARN polimerasa se desliza a lo largo del ADN en vez de asociarse y disociarse. La subunidad se libera cuando la cadena naciente de ARN tiene una longitud de 9 ó 10 nucleótidos. Tras esta liberación, la subunidad puede colaborar en la iniciación con otro núcleo enzimático. (Figura 7)")
12
INICIACIÓN Aunque la ARN polimerasa puede localizar las secuencias promotoras cuando está unida a la doble hélice de ADN, antes de comenzar la síntesis se debe desenrollar un segmento de la hélice. Se debe desaparear una región de la doble cadena de ADN de tal modo que los nucleótidos de una de sus cadenas sean accesibles para emparejarse con los ribonucleósidos trifosfato entrantes. La cadena de del ADN molde selecciona al ribonucleósido trifosfato correcto mediante la formación de pares de bases Watson-Crick, al igual que en la síntesis del ADN. ¿Qué longitud de la cadena de ADN molde desenrolla la ARNp? Cada molécula de ARNp unida desenrolla un segmento de ADN formado por 17 pares de bases. La transición desde el complejo promotor cerrado (en el cual el ADN está como doble hélice) al complejo promotor abierto (en el cual un segmento de ADN está desenrollado) es un suceso esencial de la transcripción. En este momento todo está preparado para la formación del primer enlace fosfodiéster de la nueva cadena de ARN. A diferencia con la síntesis del ADN, la síntesis del ARN puede comenzar de novo, sin ser necesario un cebador. La mayoría de las cadenas nuevamente sintetizadas portan en un extremo 5´una etiqueta muy diferenciadora: la primera base en ese extremo es pppG o pppA. La presencia de un residuo trifosfato sugiere que la síntesis del ARN comienza por el extremo 5´. Al igual que en la síntesis del ADN, la cadena de ARN crece en sentido 5´ 3´.
 al complejo promotor abierto (en el cual un segmento de ADN está desenrollado) es un suceso esencial de la transcripción. En este momento todo está preparado para la formación del primer enlace fosfodiéster de la nueva cadena de ARN. A diferencia con la síntesis del ADN, la síntesis del ARN puede comenzar de novo, sin ser necesario un cebador. La mayoría de las cadenas nuevamente sintetizadas portan en un extremo 5´una etiqueta muy diferenciadora: la primera base en ese extremo es pppG o pppA. La presencia de un residuo trifosfato sugiere que la síntesis del ARN comienza por el extremo 5´. Al igual que en la síntesis del ADN, la cadena de ARN crece en sentido 5´ 3´.")
13
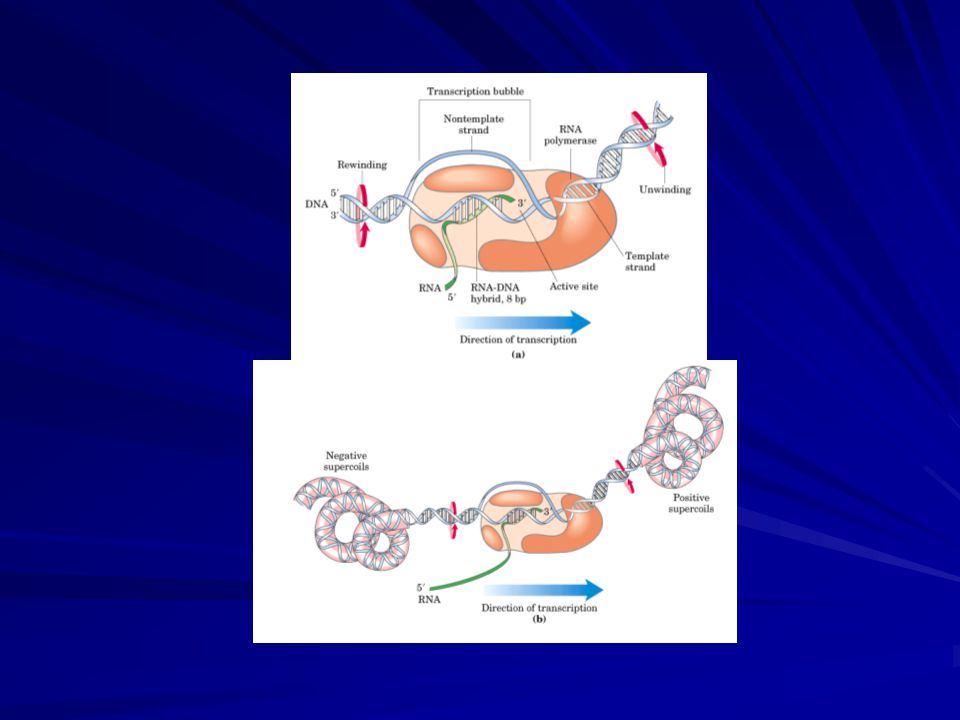
ELONGACIÓN La fase de elongación en la síntesis del ARN comienza nada más formarse el primer enlace fosfodiéster. Un cambio importante es la pérdida de ; recuérdese que el núcleo enzimático sin se une con mayor fuerza al molde de ADN. En efecto, la polimerasa permanece unida a su molde hasta que se alcanza una señal de terminación. La región que contiene la ARN polimerasa, ADN y la cadena creciente de ARN se denomina burbuja de transcripción. La cadena de ARN en crecimiento forma una hélice híbrida con la hebra molde de ADN. Esta hélice ARN-ADN tiene una longitud aproximada de 8 pares de bases, lo cual se corresponde, aproximadamente, con una vuelta de la doble hélice. En esta hélice híbrida el grupo 3'-OH del ARN está posicionado de tal modo que pueda atacar al átomo de fósforo de un ribonucleósido trifosfato entrante. Al igual que en la fase de iniciación, durante la fase de elongación se desenrollan aproximadamente 17 pares de bases del ADN. La burbuja de transcripción recorre unos 170 Å (17 nm) en un segundo, lo que equivale a una velocidad de elongación de aproximadamente 50 nucleótidos por segundo. Aunque rápida, es mucho menor que la velocidad de síntesis de ADN (800 nucleótidos por segundo). (Figura 8)
 en un segundo, lo que equivale a una velocidad de elongación de aproximadamente 50 nucleótidos por segundo. Aunque rápida, es mucho menor que la velocidad de síntesis de ADN (800 nucleótidos por segundo). (Figura 8)")
15
ELONGACIÓN Las longitudes del híbrido ARN-ADN y de la región desenrollada del ADN permanecen bastante constantes mientras la ARN polimerasa se desplaza a lo largo del molde de ADN. Este hecho indica que la velocidad de enrollado del ADN en la parte posterior de la ARNp es aproximadamente igual a la velocidad de desenrollado en su parte frontal. El híbrido ARN-ADN debe rotar también cada vez que se añade un nucleótido de tal modo que el extremo 3'-OH del ARN permanezca en el centro catalítico. La longitud del híbrido ARN-ADN está determinada por una estructura de la propia enzima que fuerza la separación del híbrido ARN-ADN, lo que permite a la cadena de ARN separarse de la enzima y a la cadena de ADN reunirse con su complementaria. La continua apertura por delante y cierre por detrás, de la burbuja de replicación, genera superenrollamientos positivos por delante y negativos por detrás que tienen que ser disipados por la acción de topoisomerasas.

16
TERMINACIÓN Al igual que su iniciación, la finalización de la transcripción viene controlada de un modo preciso. En la fase de terminación de la transcripción, cesa la formación del enlace fosfodiéster, el híbrido ARN-ADN se disocia, la región de ADN fundido se enrolla y la ARNp se desprende del ADN. ¿Qué es lo que determina dónde debe finalizar la transcripción? Las secuencias transcritas de los moldes de ADN contienen señales de parada. La más sencilla es una secuencia palindrómica rica en GC, seguida de una secuencia rica en AT. El ARN transcrito de esta secuencia palindrómica es autocomplementario. Por tanto, sus pares de bases pueden formar una estructura en horquilla con un vástago y un bucle, estructura favorecida por su alto contenido en residuos G y C. Los pares de bases G-C son más estables que los pares de bases A-T ya que tienen un puente de hidrógeno extra en cada par de bases. Esta horquilla estable es seguida por una secuencia de cuatro o más residuos de uracilo, los cuales son también cruciales para la terminación. La transcripción del ARN finaliza en ellos o justo detrás (Figura 9) ¿Cómo finaliza la transcripción en esta estructura combinada horquilla-oligo(U)? Primero, parece que la ARNp se detiene inmediatamente después de sintetizar la secuencia de ARN que se pliega en horquilla. Aún más, la hélice híbrida ARN-ADN que se forma tras la horquilla es inestable ya que sus pares de bases rU-dA son las más débiles de los cuatro tipos existentes. Por tanto, la parada en la transcripción causada por la horquilla permite al ARN en crecimiento débilmente enlazado disociarse del ADN y luego de la enzima. La hebra sencilla de ADN se reasocia a su complementaria para regenerar la doble hélice de ADN y se cierra la burbuja de transcripción. La ARNp no siempre precisa ayuda para finalizar la transcripción en una horquilla seguida de varios residuos U. En otros casos necesita la colaboración de un factor adicional para terminar, la proteína ro (). Esta proteína es un hexámero que se une específicamente a una hebra sencilla de ARN en un tramo de 72 nucleótidos, de tal forma que el ARN pasa a través del centro de la estructura. La proteína se activa mediante secuencias ricas en citosina y pobres en guanina situadas en el ARN en crecimiento. La actividad ATPasa de le permite tirar del ARN naciente mientras que sigue a la ARNp. Cuando alcanza a la ARNp en la burbuja de transcripción, corta la hélice híbrida ARN-ADN actuando como una ARN-ADN helicasa. (Figura 10)W3
 ¿Cómo finaliza la transcripción en esta estructura combinada horquilla-oligo(U) Primero, parece que la ARNp se detiene inmediatamente después de sintetizar la secuencia de ARN que se pliega en horquilla. Aún más, la hélice híbrida ARN-ADN que se forma tras la horquilla es inestable ya que sus pares de bases rU-dA son las más débiles de los cuatro tipos existentes. Por tanto, la parada en la transcripción causada por la horquilla permite al ARN en crecimiento débilmente enlazado disociarse del ADN y luego de la enzima. La hebra sencilla de ADN se reasocia a su complementaria para regenerar la doble hélice de ADN y se cierra la burbuja de transcripción. La ARNp no siempre precisa ayuda para finalizar la transcripción en una horquilla seguida de varios residuos U. En otros casos necesita la colaboración de un factor adicional para terminar, la proteína ro (). Esta proteína es un hexámero que se une específicamente a una hebra sencilla de ARN en un tramo de 72 nucleótidos, de tal forma que el ARN pasa a través del centro de la estructura. La proteína se activa mediante secuencias ricas en citosina y pobres en guanina situadas en el ARN en crecimiento. La actividad ATPasa de le permite tirar del ARN naciente mientras que sigue a la ARNp. Cuando alcanza a la ARNp en la burbuja de transcripción, corta la hélice híbrida ARN-ADN actuando como una ARN-ADN helicasa. (Figura 10)W3.")
17
Figura 10. Terminación de la transcripción dependiente del factor .

Presentaciones similares
















 de ARN ENZIMAS Y PROTEÍNAS IMPLICADAS EN LA REPLICACIÓN a) ARN polimerasa: sintetiza un fragmento de ARN constituido.>")


