Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Replicación del DNA

2
Dogma Central proteina RNA DNA replicación transcripción traducción
inversa

3
Cromosomas Procariontes

4
Cromosoma lineal eucarionte

5
Semi-continua ó semi-discontinua Alta fidelidad
Características Generales de la Replicación Semi-conservativa Bidireccional Semi-continua ó semi-discontinua Alta fidelidad

6
Replicación semiconservativa

7
Replicación bidireccional

8
Replicación semidiscontinua
la polimerización se lleva a cabo en dirección 5´--3´

10
Replicación del DNA y ciclo celular
La replicación en bacterias toma 40 min a una velocidad casi invariable de 50,000 bases/min. Después de que termina, existe un periodo de aprox. 20 minutos para que la célula se divida, por lo que todo el proceso duraría mínimamente 60 min. Los tiempos de división bacteriana pueden ir desde 18 min, hasta 160 min.

11
Etapas de la Replicación
I. Inicio: es el punto en donde se regula el proceso II. Elongación III.Terminación

12
I. Inicio de la replicación
Ori C Secuencia rica en T-A

13
Minicromosomas para determinar el origen mínimo
E. coli DNA cromosomal (E. coli) + Enzima de restricción DNA plasmídico + Enzima de restricción origen R + + R mezclado Medio con antibiótico Transformar bacterias ligasa
 + Enzima de restricción. DNA plasmídico + Enzima de restricción. origen. R. + + R. mezclado. Medio con antibiótico. Transformar. bacterias. ligasa.")
14
R Ori-coli X DNA NO se replica Se replica Medio con antibiótico

15
Metodo de footprinting para determinar el sitio de origen de la replicación

16
Dna A reconoce el sitio de inicio de la replicación
Nucleasa + DNA Ori Dna A + DNA Ori DnaB + DNA Ori DnaC + DNA Ori 245 1

17
Secuencia del origen de Replicación bacteriano
5 cajas DNAA con secuencia muy parecida, tienen diferentes afinidades por la proteína. DnaA se une a 3 sitios con alta afinidad R1,R2 y R4 durante todo el tiempo. Las otras dos cajas con afinidad intermedia son reconocidas por el complejo DnaA-ATP. Además, este complejo reconoce los sitios I y t que son de baja afinidad. Organización espacial entre los sitios es importante para regular la iniciación. The I-sites bear 9-mer sequences that differ by 3–4 bases from the DnaA box 9-mer consensus sequence (Ryan et al., 2002). In addition, the s-sites bear the 6-mer consensus TGATCC sequence, which is shared by the I2 and I3 sites 5 Nonámeros o cajas DnaA Región de los treceámeros (tres repeticiones de 13 nucleótidos), 70% AT once sitios GATC en la secuencia de 245pb
. In addition, the s-sites bear the 6-mer consensus TGATCC sequence, which is shared by the I2 and I3 sites. 5 Nonámeros o cajas DnaA. Región de los treceámeros (tres repeticiones de 13 nucleótidos), 70% AT. once sitios GATC en la secuencia de 245pb.")
18
Organización del OriC R1=R4>R2>R3=R5 DUE: DNA unwinding element
Each DnaA box has different affinities for DnaA, the order in affinities of R boxes would be: R1 = R4 > R2 > R3 = R5. DnaA boxes R1 and R4, which are located at the edges of the DnaA assembly region, have the highest affinity (Kd of 3–9 nM) for DnaA regardless of its nucleotide forms. Specific binding sites for bacterial histone-like proteins IHF and Fis are also present in the DnaA assembly region. These proteins bend DNA at the binding sites . IHF can stimulate initiation at oriC in vitro and in vivo. IhF assists in the formation of DnaA multimers by binding to a specific site in oriC, leading to bending of the DNA and enhancing ATP–DnaA binding to the I1–I3 sites in particular, the T-rich upper-strand of ssDUE preferentially binds ATP-DnaA molecules (Ozaki et al.,2008). This ATP-DnaA ssDUE interaction is crucial for open complex formation DUE: DNA unwinding element IHF: Integration host factor FIS factor R1=R4>R2>R3=R5
 for DnaA regardless of. its nucleotide forms. Specific binding sites for bacterial histone-like proteins IHF and Fis are also present in the DnaA assembly region. These proteins bend DNA at the binding sites . IHF can stimulate initiation at oriC in vitro and in vivo. IhF assists in the formation of DnaA multimers by binding to a specific site in oriC, leading to bending of the DNA and enhancing ATP–DnaA binding to the I1–I3 sites in particular, the T-rich upper-strand of ssDUE. preferentially binds ATP-DnaA molecules (Ozaki et al.,2008). This ATP-DnaA ssDUE interaction is crucial for open complex formation. DUE: DNA unwinding element. IHF: Integration host factor. FIS factor. R1=R4>R2>R3=R5.")
20
DnaA A region involved in membrane binding is located between domains III and IV that is postulated to induce the dissociation of ADP from DnaA in vivo so that DnaA can bind ATP to support initiation

22
The relative increases and decreases in the concentrations of factors at oriC as a function of progression through the cell cycle, as derived from experimental observations10,98, are shown. a | Before the start of initiation, DnaA (green) is bound to several high-affinity DnaA boxes at oriC. Fis (factor for inversion stimulation, orange) is also associated with its primary binding site, whereas IHF (integration host factor, blue) is absent. b | At the start of initiation, the local concentration of ATP–DnaA at oriC increases, while at the same time Fis occupancy decreases. c | As initiation progresses, IHF binds oriC. The severe bending of DNA by IHF probably assists the interaction of DnaA with its weaker affinity binding sites during the formation of the final nucleoprotein complex and unwinding of the DNA-unwinding element (DUE).
 is bound to several high-affinity DnaA boxes at oriC. Fis (factor for inversion stimulation, orange) is also associated with its primary binding site, whereas IHF (integration host factor, blue) is absent. b | At the start of initiation, the local concentration of ATP–DnaA at oriC increases, while at the same time Fis occupancy decreases. c | As initiation progresses, IHF binds oriC. The severe bending of DNA by IHF probably assists the interaction of DnaA with its weaker affinity binding sites during the formation of the final nucleoprotein complex and unwinding of the DNA-unwinding element (DUE)..")
23
Activación 20-30 DnaA DiaA: DnaA initiator-associating protein
IHF: Integration Host factor Activación 20-30 DnaA DiaA enhances ATP–DnaA binding to all low-affinity sites16,17,24 (FIG. 2). DiaA forms homotetramers in solution that bind multiple DnaA molecules and stimulate DnaA assembly on oriC
. DiaA forms homotetramers in solution that bind multiple DnaA molecules and stimulate DnaA. assembly on oriC.")
24
Arginina 285 es importante en la oligomerización

25
Proteína DnaA Monómero 52kDa
Se une con alta afinidad y de forma cooperativa a las cajas dnaA Estequiometria de hasta subunidades de DnaA por oriC Une ATP y lo hidroliza a una velocidad muy baja Una vez abiertos los treceámeros, DnaA reconoce las cajas presentes en el DUE y se une a la cadena sencilla estabilizandola.

27
Regulación negativa del inicio de la replicación

28
1) Hda (RIDA: regulatory inactivation of DnaA)
Después del ensamblaje del replisoma y que se completó un fragmento de Okazaki, la subunidad b del replisoma permanece unida al DNA y forma un complejo con ADP-Hda. Reduce la presencia del complejo activo DnaA-ATP estimulando la hidrólisis de ATP en una forma dependiente de la replicación. Este es el mecanismo más importante que reprime la iniciación. Hda homólogo de DnaA con un sitio AAA+ que interactúa con DnaA , estimulando la hidrólisis. Mutantes de DnaA con poca actividad hidrolítica o de Hda, producen excesivas iniciaciones. Hda es activada por la unión de ADP. Dado que la cantidad de ATP es alta, cualquier síntesis de novo de DnaA se acompleja con ATP.

29
2) dat locus (DnaA titulación): sitio en el cromosoma que une
grandes cantidades de proteína DnaA Aproximadamente moléculas de DnaA pueden unirse al locus dat. El locus dat (1kb) contiene 5 cajas DnaA y un sitio para IHF DDAH: dat dependent DnaA-ATP hydrolisis Deleciones en dat, resultan en una extrainiciación, aunque no inhiben el crecimiento. Esta titulación se incrementa cuando el cromosoma se duplica.
 contiene 5 cajas DnaA. y un sitio para IHF. DDAH: dat dependent DnaA-ATP hydrolisis. Deleciones en dat, resultan en una extrainiciación, aunque no inhiben el crecimiento. Esta titulación se incrementa cuando el cromosoma se duplica.")
30
3) Interacción de OriC con SeqA y la membrana
Immediately following replication, the newly synthesized strand is unmethylated, and the resulting hemimethylated origin is sequestered at the lipid bilayer of membrane by SeqA. This is not accessible to replicatively active ATP–DnaA. After approximately one-third of the cell cycle, the sequestered origin is released and methylated by Dam methylase. At this point in the cell cycle, the levels of ATP–DnaA are not sufficient to catalyze a new round of replication. As such, sequestration serves as a mechanism to prevent secondary initiations

31
Regulacion de la expresión de DnaA
Promotor de DnaA tiene cajas DnaA y secuencias GATC. Regulación de DnaA Notably, several GATC sites in oriC coincide with weak DnaA-binding sites, allowing SeqA binding to directly block DnaA association and reset oriC to a pre-initiation state. DnaA titration by datA could effectively reduce the concentration of the initiator at the origin to levels below those required to stimulate further rounds of initiation. One of the most notable examples of DnaA-mediated repression occurs Figure 6 | Initiation regulator mechanism. a | The structure of the Escherichia coli SeqA C-terminal DNA-binding domain in complex with a hemimethylated GATC-containing oligonucleotide (RCSB Protein Data Bank (PDB) ID 1LRR)61. The top and bottom strands of the GATC sequence are shown in orange. The methylated adenine is indicated in green. The residues that are involved in recognizing the central AT base pair (N150 and N152) and in interacting with the DNA minor groove (R86 and R116) are labelled. b | The structure of the E. coli SeqA N-terminal multimerization domain (PDB ID 1XRX)63. The N-terminal domain is shown as a dimer in which monomer A and B are shown as a magenta and brown ribbon, respectively. Transparent subunits reflect higher-order contacts that are present between the dimers in the crystal. c | SeqA filamentation during oriC replication. SeqA initially recognizes hemimethylated GATC sequences as a dimer. Oligomerization between SeqA dimers forms extended filaments and leads to the rapid sequestration of newly replicated DNA. d | A model for the inactivation of DnaA by the Hda regulator. Interaction with the β-clamp (blue) promotes localization of Hda (grey) to oriC (top panel). Hda may interact with the exposed ATP end of the DnaA filament (green), promoting ATP hydrolysis in either a sequential (bottom-left panel) or catastrophic (bottom-right panel) manner, to inactivate DnaA. at the transcriptional unit for the dnaA gene itself69,70. A collection of two DnaA boxes, as well as four ATP– DnaA boxes, is located between the two promoters for dnaA. Although only the two DnaA boxes are occupied in the presence of ADP–DnaA, all possible binding sites are filled by ATP–DnaA, resulting in stronger repression of dnaA transcription. Regulatory inactivation of DnaA (RIDA) is another control event, and directly stimulates ATP hydrolysis by DnaA after the initiator melts the DUE
 ID 1LRR)61. The top and bottom strands of the GATC sequence are shown in orange. The methylated adenine is indicated in green. The residues that are involved in recognizing the central AT base pair (N150 and N152) and in interacting with the DNA. minor groove (R86 and R116) are labelled. b | The structure of the E. coli SeqA N-terminal multimerization domain (PDB ID. 1XRX)63. The N-terminal domain is shown as a dimer in which monomer A and B are shown as a magenta and brown ribbon, respectively. Transparent subunits reflect higher-order contacts that are present between the dimers in the crystal. c | SeqA filamentation during oriC replication. SeqA initially recognizes hemimethylated GATC sequences as a dimer. Oligomerization between SeqA dimers forms extended filaments and leads to the rapid sequestration of newly replicated DNA. d | A model for the inactivation of DnaA by the Hda regulator. Interaction with the β-clamp (blue) promotes localization of Hda (grey) to oriC (top panel). Hda may interact with the exposed ATP end of the DnaA filament (green), promoting ATP hydrolysis in either a sequential (bottom-left panel) or catastrophic (bottom-right panel) manner, to inactivate DnaA. at the transcriptional unit for the dnaA gene itself69,70. A collection of two DnaA boxes, as well as four ATP– DnaA boxes, is located between the two promoters for dnaA. Although only the two DnaA boxes are occupied. in the presence of ADP–DnaA, all possible binding sites are filled by ATP–DnaA, resulting in stronger repression of dnaA transcription. Regulatory inactivation of DnaA (RIDA) is another. control event, and directly stimulates ATP hydrolysis by DnaA after the initiator melts the DUE.")
32
Reactivación de DnaA-ADP a DnaA-ATP
Recently, a system for the regeneration of ATP–DnaA from ADP–DnaA was discovered in E. coli59. Two chromosomalDNA regions termed DnaA-reactivating sequence 1 (DARS1) and DARS2 bear threeflanking DnaA boxes. ADP–DnaA oligomers are formed on these sites, promoting dissociation of ADP from DnaA and subsequent ATP binding in vitro DARS: DnaA reactivation sequence: sitios en el cromosoma que contienen cajas DnaA en orientaciones diferentes
 and DARS2 bear threeflanking DnaA boxes. ADP–DnaA oligomers are formed on these sites, promoting dissociation of ADP from DnaA and subsequent. ATP binding in vitro. DARS: DnaA reactivation sequence: sitios en el cromosoma que contienen cajas DnaA. en orientaciones diferentes.")
34
DiaA: DnaA initiator-associating protein
IHF: Integration Host factor Activación 20-30 DnaA 2. Formación del pre-primosoma DiaA enhances ATP–DnaA binding to all low-affinity sites16,17,24 (FIG. 2). DiaA forms homotetramers in solution that bind multiple DnaA molecules and stimulate DnaA assembly on oriC
. DiaA forms homotetramers in solution that bind multiple DnaA molecules and stimulate DnaA. assembly on oriC.")
35
HELICASA DnaB C-terminal ATPasa Se mueve en dirección 5´---3´
Following unwinding of the DUE, another important function for DnaA is to recruit and properly position the DnaB helicase. Interestingly, the helicase loader that helps to mediate this reaction, DnaC, is both an AAA+ ATPase and a DnaA paralogue41,43. Although helicase loading depends on the association of the N-terminal domain of DnaA with DnaB, the phylogenetic relationship between DnaA and DnaC raises the possibility that a specific association between these two ATPases could further regulate the process After DnaB loads at oriC , DnaC must dissociate so that DnaB can unwind the parental duplex DNA. Recent evidence indicates that primer formation by primase, which interacts with the smaller domain of DnaB during primer synthesis [42,84,85 ], causes the dissociation of DnaC bound to the larger C-terminal domain of DnaB For DnaB moving either the rightward or leftward, the interaction of primase with DnaB leads to primer formation, which is followed by the binding to the primer end by one unit of a dimer of DNA polymerase III holoenzyme. This unit of the dimer synthesizes the leading strand as it interacts via its t subunit with DnaB helicase

36
DnaB Dna C Monómeros de 60kDa que forman un homohexámero
Actividad de helicasa Dominios para : Unión a DNA de cadena doble Unión a DNA cadena sencilla Interacción con DnaC y DnaA Hidrólisis de NTPs (ATP) Monómeros de 28 kDa Homohexámero que permite la llegada de DnaB ATPasa: al hidrolizar ATP pierde afinidad por DnaB As DnaB unwinds DNA, it moves in the 50 -to-30 direction on this parental DNAStrand DnaB must be complexed with DnaC for the helicase to be able to bind to the respective replication origin (reviewed in Ref. [49 ]). However bound to DnaB, DnaC inhibits its helicase and ATPase activity, so DnaC must dissociate for DnaB to be active. DnaC has been described as the ‘helicase loader’ [54–57 ]. This term is a misnomer because it implies an active role for DnaC; the evidence thus far indicates that DnaA is the helicase loader, and DnaC has a passive but essential role in the delivery of DnaB at oriC . Bound to the larger domain of DnaB to form the DnaB– DnaC complex [58,59 ], DnaC apparently interacts via an N-terminal region to inhibit the ATPase and helicase activity of DnaB
 Monómeros de 28 kDa. Homohexámero que permite la llegada de DnaB. ATPasa: al hidrolizar ATP pierde afinidad por DnaB. As DnaB unwinds DNA, it moves in the 50 -to-30 direction on this parental DNAStrand DnaB must be complexed with DnaC for the helicase to be able to bind to the respective replication origin (reviewed in Ref. [49 ]). However bound to DnaB, DnaC inhibits its helicase and ATPase activity, so DnaC must dissociate for DnaB to be active. DnaC has been described as the ‘helicase loader’ [54–57 ]. This term is a misnomer because it implies an active role for DnaC; the evidence thus far indicates that DnaA is the helicase loader, and DnaC has a passive but essential role in the delivery of DnaB at oriC . Bound to the larger domain of DnaB to form the DnaB– DnaC complex [58,59 ], DnaC apparently interacts via an N-terminal region to inhibit the ATPase and helicase activity of DnaB.")
37
Pre-primosoma y primosoma

38
II. Elongación

39
La replicación del DNA requiere un cebador catalizado por la DNA Primasa
Dna G, Primasa, ≈ 60 kDa RNA polimerasa Sintetiza cebador de ≈ 12nt DNA Primasa reconoce a DnaB

40
Proteínas de unión a cadena sencilla: SSB
Las proteínas SSB se unen con alta afinidad al DNA de cadena sencilla y lo protegen de nucleasas y de asociaciones intracatenarias

41
DNA polimerasas de bacterias

42
Reacción de polimerización

43
DNA polimerasas en E. coli

44
Características de la DNA polimerasa I

45
Actividades de la DNA polimerasa I

46
Mutantes de E.coli en el gen de la Pol I son “viables”, por lo tanto la DNA polI no es la principal enzima replicativa (pero no soportan deleciones del gen). Mutantes de DNA pol II son viables, aún con el gen deletado. La DNApolIII es la principal enzima en la replicación del DNA. Las mutaciones son letales.

47
Fidelidad se refiere al seguimiento exacto de la secuencia de DNA que sirve como molde
La fidelidad de la replicación en bacterias: ≈10-8 a 10-10 Procesividad # de nucleótidos que se sintetizan en un solo evento de unión. Una enzima procesiva agrega miles.

48
Por qué no existe una DNA polimerasa 3´--5´?

49
DNA polimerasa III holoenzima
an asymmetric dimer

50
Subunidades de la DNA polimerasa III de E.coli
# por holoenzima Mr Función dnaE α 2 132,000 Subunidad catalítica dnaQ ε 27,000 Proofreading holE θ 10,000 Acoplador dnaX 71,000 Dimerización 1 48,000 holA δ 35,000 holB δ´ 33,000 holC χ 15,000 holD ψ 12,000 dnaN β 4 37,000 Procesividad Pol III Núcleo nuc/seg Pol III’ Abrazadera que carga las subunidades β al DNA Complejo ó complejo Pol III* Pol III holo 1,000 bases/seg

51
Procesividad de la polimerasa III
La procesividad de la DNApol lII* aumenta de 50 nts a más de 50,000 nts gracias a la subunidad β

52
Modelo del dímero

53
Cuál es la participación de las DNA polimerasas
en la replicación?

54
Maduración de los fragmentos de Okazaki
DNA polimerasa I ayuda a remover el cebador con su actividad de exonucleasa 5´-3´ Actividad de polimerasa reemplaza los nucleótidos eliminados ≈17 DNA ligasa cataliza la formación de enlace fosfodiéster

55
DNA ligasa

56
DNA topoisomerasas

57
Topoisomerasas en E. coli
Topoisomerasa I Topoisomerasa III Topoisomerasa II (DNA Girasa) Topoisomerasa IV IA. 5´ IB 3´ IIA. IIB TOPOISOMERASAS I Gyrase catalyzes the formation of negative supercoils and also can relax positive supercoils (a topologically equivalent reaction), but is inefficient at unknotting and decatenation. o meet their unique needs, most bacteria possess two specialized type IIA topos (Figure 1(a)): DNA gyrase, encoded by the gyrA and gyrB genes; and topo IV, encoded by parC and parE. The four DNA topoisomerases found in E. coli consist of the two type IA enzymes, DNA topoisomerases I and III, and the two type IIA enzymes, DNA gyrase and DNA topoisomerase IV. Although some overlap of function has been shown genetically, each of the DNA topoisomerases appears optimized to carry out its own particular set of topological manipulations. DNA gyrase is the only known topoisomerase able to generate negative supercoiling at the expense of ATP hydrolysis and is responsible for global generation of negative supercoils in the bacterial chromosome. Thus, topoisomerases III and IV appear to have overlapping functions. the bacterial topoisomerases I will relax native plasmid DNAs isolated directly from bacteria, whereas the E. coli and eukaryotic topoisomerases III (® and ¯) characterized to date require a hypernegatively supercoiled plasmid DNA substrate TOPOISOMERASAS II
 Topoisomerasa IV. IA. 5´ IB 3´ IIA. IIB. TOPOISOMERASAS I. Gyrase catalyzes the formation of negative supercoils and also can relax positive supercoils (a topologically equivalent reaction), but is inefficient at unknotting and decatenation. o meet their unique needs, most bacteria possess two specialized type IIA topos (Figure 1(a)): DNA gyrase, encoded by the gyrA and gyrB genes; and topo IV, encoded by parC and parE. The four DNA topoisomerases found in E. coli consist of the two type IA enzymes, DNA topoisomerases I and III, and the two type IIA enzymes, DNA gyrase and DNA topoisomerase IV. Although some overlap of function has been shown genetically, each of the DNA topoisomerases appears optimized to carry out its own particular set of topological manipulations. DNA gyrase is the only known topoisomerase able to generate negative supercoiling at the expense of ATP hydrolysis and is responsible for global generation of negative supercoils in the bacterial chromosome. Thus, topoisomerases III and IV appear to have overlapping functions. the bacterial topoisomerases I will relax native plasmid DNAs isolated directly from bacteria, whereas the E. coli and eukaryotic topoisomerases III (® and ¯) characterized to date require a hypernegatively supercoiled plasmid DNA substrate. TOPOISOMERASAS II.")
58
III. Termino de la replicación
when DnaB unwinds the duplex DNA ahead of the non-permissive face of the Tus–Ter complex, a GC base pair located at position 6 in the Ter core sequence is broken and the C(6) moves 14 A ˚ from its normal position to bind tightly in a specific cytosine-specific binding pocket at the surface of the non-permissive face of Tus. This base flipping results in a tighter interaction between Tus and Ter, called the Tus–Ter-lock (TT-lock), which stalls the replication fork until the second replisome arrives
 moves 14 A ˚ from its normal position to bind tightly in a specific cytosine-specific binding pocket at the surface of the non-permissive face of Tus. This base flipping results in a tighter interaction between Tus and Ter, called the Tus–Ter-lock (TT-lock), which stalls the replication fork until the second replisome arrives.")
59
ORIGEN DE REPLICACION EUCARIONTE
Solo uno o múltiples? Tiempo de replicación/tamaño de genoma Múltiples orígenes?/Orígenes diferenciales? Encendido durante el ciclo Existe una secuencia única? Experimentos con plásmidos Levaduras vs eucariotes superiores Replicación una vez por ciclo? Depende del ciclo celular?

60
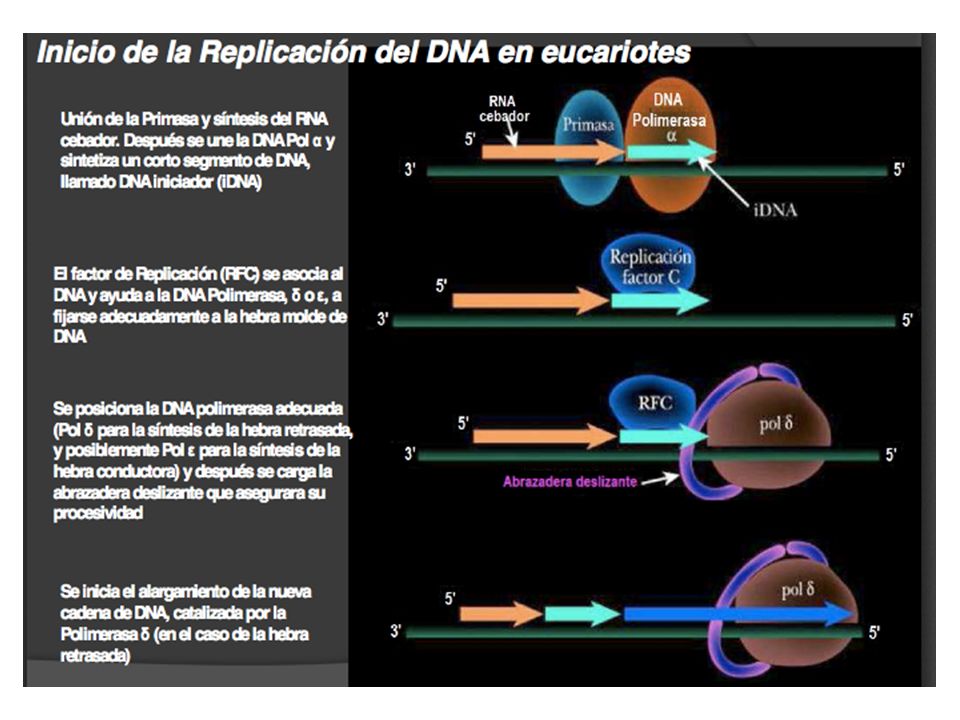
Inicio de la replicación en eucariotes
Mas de 10 kpb En S. cerevisiae en promedio cada 30,000. En mamíferos se encuentran entre 100,000 a 250,000. La tasa de replicación es aprox. 2000bp/min.

61
No todos los orígenes se activan al mismo tiempo

62
Eucromatina se duplica primero, heterocromatina al final

63
Ensayos para identificar secuencias que actúen como orígenes

64
Inicio de la replicación en eucariontes :Saccharomyces cerevisiae
ARS: Autonomously replicating sequence ATTTAATATTTTGGA % of origin function Mutaciones en A abaten la función. En B sólo se reduce

65
Orc 6 es estructuralmente relacionada a TFIIB
ARS Proteínas ORC Origin recognition complex (ORC) complejo que se une a A y B1 ORC 1-6 Se encuentra unido durante todo el ciclo celular Cdc6 Cdt1 ORC 1-6 ORC comprende un complejo de 6 subunidades que se unen formando un especie de anillo. Existe una éptima proteína que sella la unión, cdc6. Este complejo identifica y marca los orígenes promoviendo la unión al sitio de la helicasa. Orc1, Orc 4 -5 y cdc6 tienen un dominio de ATPasa similar al de la DnaA. Existen además otros módulos como el de unión al DNA que es una variante del HTH. La actividad de ATPasa es importante para la actividad de la proteína, aunque esto no se conserva en los ortólogos. La unión de ORC al DNA no se ha entendido completamente porque no hay datos que indiquen que esa proteína puede abrir las cadenas Orc 6 es estructuralmente relacionada a TFIIB
 complejo que se une a A y B1. ORC Se encuentra unido durante todo. el ciclo celular. Cdc6. Cdt1. ORC ORC comprende un complejo de 6 subunidades que se unen formando un especie de anillo. Existe una éptima proteína que sella la unión, cdc6. Este complejo identifica y marca los orígenes promoviendo la unión al sitio de la helicasa. Orc1, Orc 4 -5 y cdc6 tienen un dominio de ATPasa similar al de la DnaA. Existen además otros módulos como el de unión al DNA que es una variante del HTH. La actividad de ATPasa es importante para la actividad de la proteína, aunque esto no se conserva en los ortólogos. La unión de ORC al DNA no se ha entendido completamente porque no hay datos que indiquen que esa proteína puede abrir las cadenas. Orc 6 es estructuralmente relacionada a TFIIB.")
66
Estructura y formación de la helicasa Mcm2-7
Subunidades distintas de una familia relacionada. El hexámero es similar superficialmente a la estructura en anillo que forma DnaB con un N-terminal de oligomerización y un C terminal que es el motor que rodea y se trasloca a lo largo del DNA. Este complejo se trasloca en dirección 3`--5`contrario a DNAB. Existe una unión entre las proteínas en el orden 5,3,7,4,6,2, siendo un punto natural de corte 2-5. En algunos casos la unión a ATP permite cerrar o abrir el anillo, pero no ocurre en todos los organismos. Traslocación 3´--5´

67
Complejo CMG: Cdc45-Mcm-GINS a través de Sld2 y Sld3
Pre-RC “licencia para replicar” Activación: eventos de fosforilación La unión de Mcm2-7 al DNA, junto con cdc6 y cdt1 marca el licenciamiento del inicio de la replicación. La unión de Mcm al DNA es mediante la accion conjunta de ORC, cdc6 y cdt1, formando el complejo de Pre-replicación. ATP se requiere para el posicionamiento, con la hidrólisis de ATP por Cdc6 y un recambio de nucleótido por Orc 1 promoviendo una asociación estable de la helicasa con el DNA.Cdc6 proteína muy inestable con una vida media de menos de 5 minutos. Se sintetiza durante G1. En mamíferos se controla diferente, es fosforilada durante la fase S y como resultado sale del núcleo. Su actividad de ATPasa sí se requiere. Las proteinas MCM son los factores de licenciamiento. Mutantes que fallan en el recambio de MCM afectan el sistema de ubiquitinación.MCM2,3,5 entran al núcleo solo durante mitosis. En levaduras, la presencia de Cdc6 en el origen permite a las proteínas MCM unirse. Cuando ocurre la iniciación Cdc6 y MCM son desplazadas. Origin firing is brought about by multiple phosphorylation events carried out by cyclin E-CDK2 at the onset of S phase and by Cdc7- Dbf4 (DDK kinase or Dbf4-dependent kinase) prior to individual origin firing (Figure 3B) [40–44]. At this point, origin melting occurs and DNA unwinding by the helicase generates ssDNA, exposing a template for replication (Figure 3C). Complejo CMG: Cdc45-Mcm-GINS a través de Sld2 y Sld3 Iniciación
 prior to individual origin firing (Figure 3B) [40–44]. At this point, origin melting occurs and DNA unwinding by the helicase generates ssDNA, exposing a template for replication (Figure 3C). Complejo CMG: Cdc45-Mcm-GINS. a través de Sld2 y Sld3. Iniciación.")
68
Complejo CMG Psf1 GINS Psf2 Psf3 CMG + Cdc45+ MCM2-7 Sld5
Incrementan la actividad de la helicasa al inicio para abrir las cadenas y permanecen a lo largo de la elongación

69
Control del inicio de la replicación
a lo largo del ciclo celular

70
Cdt1 y geminina limitan la replicación del DNA
To prevent multiple rounds of DNA replication during a single cell cycle, mechanisms have evolved to regulate limiting initiation factors: ORC, Cdt1, and Cdc6 (reviewed in [49]). After origin firing, Orc1, the largest ORC subunit, is ubiquitinated and degraded [50]. Phosphorylation of human Orc2 leads to the dissociation of Orc2–6 from chromatin to Thus, duplicating the genome once and only once per cell cycle is controlled at the level of licensing. These multiple and redundant mechanisms prevent origin assembly to restrict replication licensing once per cell cycle.These controls emphasize the importance of preventing overreplication of chromosomal DNA, in order to preserve genomic integrity. 3.2. The
. After origin firing, Orc1, the largest ORC subunit, is ubiquitinated and degraded [50]. Phosphorylation of human Orc2 leads to the dissociation of Orc2–6 from chromatin to Thus, duplicating the genome once and only once per cell cycle is controlled at the level of licensing. These multiple and redundant mechanisms prevent origin assembly to restrict replication licensing once per cell cycle.These controls emphasize the importance of preventing overreplication of chromosomal DNA, in order to preserve genomic integrity The.")
71
Pol -cadena lagging Pol -cadena líder PCNA (Helicasa) (PCNA-like)
MCM8 (primasa) Pol -cadena lagging Pol -cadena líder
 Pol -cadena lagging. Pol -cadena líder.")
73
Proliferating Cell Nuclear Antigen PCNA
Proteína 29kDa Incrementa procesividad de la DNA polimerasa Forma un homotrímero alrededor del DNA PCNA loading is accomplished by the replication factor C (RFC) complex, which is comprised of five AAA+ ATPases [73]. RFC recognizes primer-template junctions and loads PCNA at these sites [74,75]. The PCNA homotrimer is opened by RFC using energy from ATP hydrolysis and is then loaded onto DNA in the proper orientation to facilitate its association with the polymerase [76,77]. Clamp loaders can also unload PCNA from DNA, a mechanism that becomes important when replication must be terminated.].. The PCNA homotrimer is opened by RFC using energy from ATPhydrolysis and is then loaded onto DNA in the proper orientation to facilitate its association with the polymerase [76,77]. Clamp loaders can also unload PCNA from DNA, a mechanism that becomes important when replication must be terminated
 complex, which is comprised of five AAA+ ATPases [73]. RFC recognizes primer-template junctions and loads PCNA at these sites [74,75]. The PCNA homotrimer is opened by RFC using energy from ATP hydrolysis and is then loaded onto DNA in the proper orientation to facilitate its association with the polymerase [76,77]. Clamp loaders can also unload PCNA from DNA, a mechanism that becomes important when replication must be terminated.].. The PCNA homotrimer is opened by RFC using energy from ATPhydrolysis and is then loaded onto DNA in the proper orientation to facilitate its association. with the polymerase [76,77]. Clamp loaders can also unload PCNA from DNA, a mechanism that becomes important when replication must be terminated.")
74
RFC: “ clamp loader” de PCNA
Mechanism of RFC. (a) The clamp loader is in the closed form with amino-terminal domains closed upon the RFC-5 stator. (b) ATP binding to the RFC 234 motor releases the amino-terminal domains from RFC-5. (c) PCNA docks onto the amino-terminal domains and the RFC-1 wrench cracks one of the interfaces, allowing the PCNA ring to spring open like β. (d) The RFC complex binds DNA and places it through the open PCNA ring. (e) DNA binding results in ATP hydrolysis which closes the amino-terminal domains upon the RFC-5 stator, releasing the PCNA ring onto DNA. Protein subunits in the diagram are also labeled with the analogous proteins of the E. coli system. The analogous human subunits are given in
 The clamp loader is in the closed form with amino-terminal domains closed upon the RFC-5 stator. (b) ATP binding to the RFC 234 motor releases the amino-terminal domains from RFC-5. (c) PCNA docks onto the amino-terminal domains and the RFC-1 wrench cracks one of the interfaces, allowing the PCNA ring to spring open like β. (d) The RFC complex binds DNA and places it through the open PCNA ring. (e) DNA binding results in ATP hydrolysis which closes the amino-terminal domains upon the RFC-5 stator, releasing the PCNA ring onto DNA. Protein subunits in the diagram are also labeled with the analogous proteins of the E. coli system. The analogous human subunits are given in.")
75
Polimerasas de eucariotes

76
Horquilla de la Replicación Eucarionte

77
Procesamiento de los fragmentos de Okazaki

78
Terminación en eucariontes: El dilema de los cromosomas lineales

79
Telomerasa GGTTAG Telomerasa

80
T-loop

Presentaciones similares

















 de ARN ENZIMAS Y PROTEÍNAS IMPLICADAS EN LA REPLICACIÓN a) ARN polimerasa: sintetiza un fragmento de ARN constituido.>")






