Descargar la presentación
La descarga está en progreso. Por favor, espere

1
BIOQUIMICA DE LAS PROTEINAS

2
CONCEPTOS INICIALES

3
Bioquímica Concepto: Es el estudio de las sustancias presentes en los organismos vivos y de las reacciones químicas en las que se basan los procesos vitales.

4
Objeto Su objetivo principal es el conocimiento de la estructura y comportamiento de las moléculas biológicas, que son compuestos de carbono que forman las diversas partes de la célula y llevan a cabo las reacciones químicas que le permiten crecer, alimentarse, reproducirse y usar y almacenar energía.

5
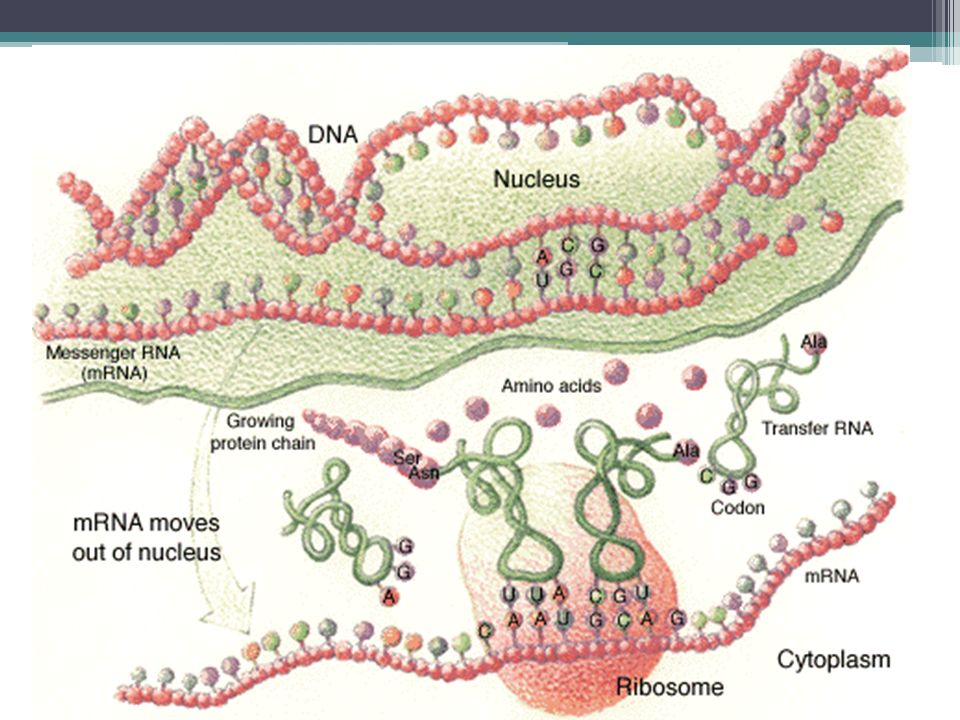
Acidos nucléicos Los ácidos nucleicos son responsables del almacen y transferencia de la información genética. Son moléculas grandes formadas por cadenas largas de unas subunidades llamadas bases, que se disponen según una secuencia exacta. Éstas, son "leídas" por otros componentes de las células y utilizadas como patrones para la fabricación de proteinas. Hay 2 tipos de ácidos nucleicos el ácido desoxirribonucleico (ADN) y el ácido ribonucleico (ARN), y están presentes en todas las células.
 y el ácido ribonucleico (ARN), y están presentes en todas las células.")
7
Síntesis de proteína El mensaje genético se encuentra en las cadenas de ADN. Para que la célula se divida este ADN dedbe duplicarse: REPLICACIÓN, repartiéndose entre las células hijas. Durante la interfase el funcionamiento de la célula está dirigido por las proteínas. A partir del ADN se forma una molécula de ARN (TRANSCRIPCIÓN) que sale del núcleo: ARN y es "leído" por el RNAr con la ayuda del RNAt que le provee los aminoácidos para la formación de las proteínas: TRADUCCIÓN
 que sale del núcleo: ARN y es leído por el RNAr con la ayuda del RNAt que le provee los aminoácidos para la formación de las proteínas: TRADUCCIÓN.")
9
El apareamiento normal en una doble cadena de DNA sería:
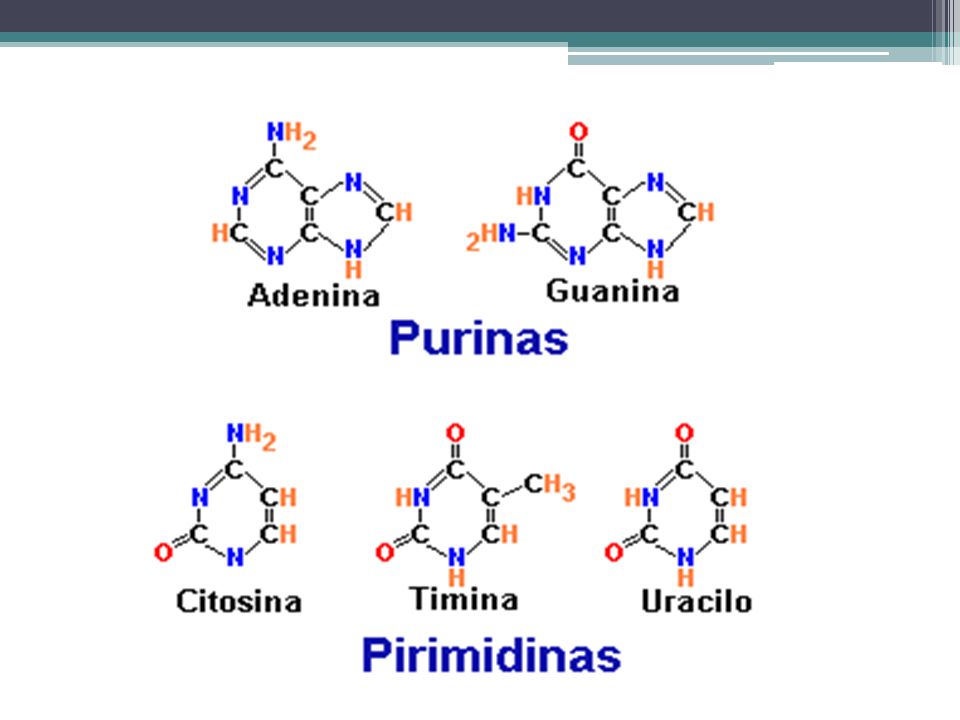
DNA – DNA A – T G – C T A C – G El apareamiento de bases entre DNA y RNA sería el siguiente: DNA – RNA A – U G – C T – A C – G

10
Proteínas Las proteínas son moléculas grandes formadas por pequeñas subunidades denominadas aminoácidos. Utilizando sólo 20 aminoácidos distintos, la célula elabora miles de proteínas diferentes, cada una de las cuales desempeña una función altamente especializada.

12
Proteínas Son biopolímeros grandes muchos de los cuales funcionan como enzimas. Otros sirven como componentes estructurales importantes de las células y de los organismos. Su estructura está determinada en gran parte, por el orden de los aminoácidos que se unen. Este orden, a su vez, está codificado por los genes.

13
Carbohidratos Los hidratos de carbono son las moléculas energéticas básicas de la célula. Contienen proporciones aproximadamente iguales de carbono e hidrógeno y oxígeno. Las plantas obtienen lo carbohidratos de la fotosíntesis Los animales, obtienen sus hidratos de carbono de los alimentos

14
Lípidos Los lípidos son sustancias grasas que desempeñan diversos papeles en la célula. Algunos se almacenan para ser utilizados como combustible de alto valor energético, mientras que otros se emplean como componentes esenciales de la membrana celular

15
Metabolismo Todas éstas moléculas y la misma célula, se hallan en constante variación. Una célula no puede mantenerse viva a menos que esté continuamente formando y rompiendo proteínas, hidratos de carbono y lípidos; reparando los ácidos nucleicos dañados y utilizando y almacenando energía. El conjunto de estos procesos activos y dependientes de la energía se denomina metabolismo

16
Proteínas La función primordial de la proteína es:
producir tejido corporal y sintetizar enzimas. Las proteínas animales y vegetales no se utilizan en la misma forma en que son ingeridas, sino que las enzimas digestivas (proteasas) deben descomponerlas en aminoácidos que contienen nitrógeno.
 deben descomponerlas en aminoácidos que contienen nitrógeno.")
17
Proteínas Rompiendo los enlaces de péptidos que ligan los aminoácidos ingeridos para que éstos puedan ser absorbidos por el intestino hasta la sangre y reconvertidos en tejido.

18
Aminoacidos De los 20 aminoácidos que componen las proteínas, ocho se consideran esenciales es decir: como el cuerpo no puede sintetizarlos, deben ser tomados ya listos a través de los alimentos. Si estos aminoácidos esenciales no están presentes al mismo tiempo y en proporciones específicas, los otros aa, no pueden utilizarse para construir las proteínas.

19
Aminoacidos Existen en la naturaleza aproximadamente unos 300 aminoácidos diferentes, pero solo 20 de ellos son de importancia al encontrarse en la formación de las moléculas de proteína de todas las formas de vida: vegetal, animal o microbiana. Estos son sintetizados a partir de precursores más sencillos o absorbidos como nutrientes.

20
Aminoacidos Los aminoácidos se subdividen en:
Cetogénicos: Dan origen a acetil-CoA y acetoacetil CoA Glucogénicos: Dan origen a glucosa por medio de oxalacetato y piruvato. Estrictamente acetogénicos: Que dan origen acetocetil CoA o acetil CoA (lisina y leusina).
.")
21
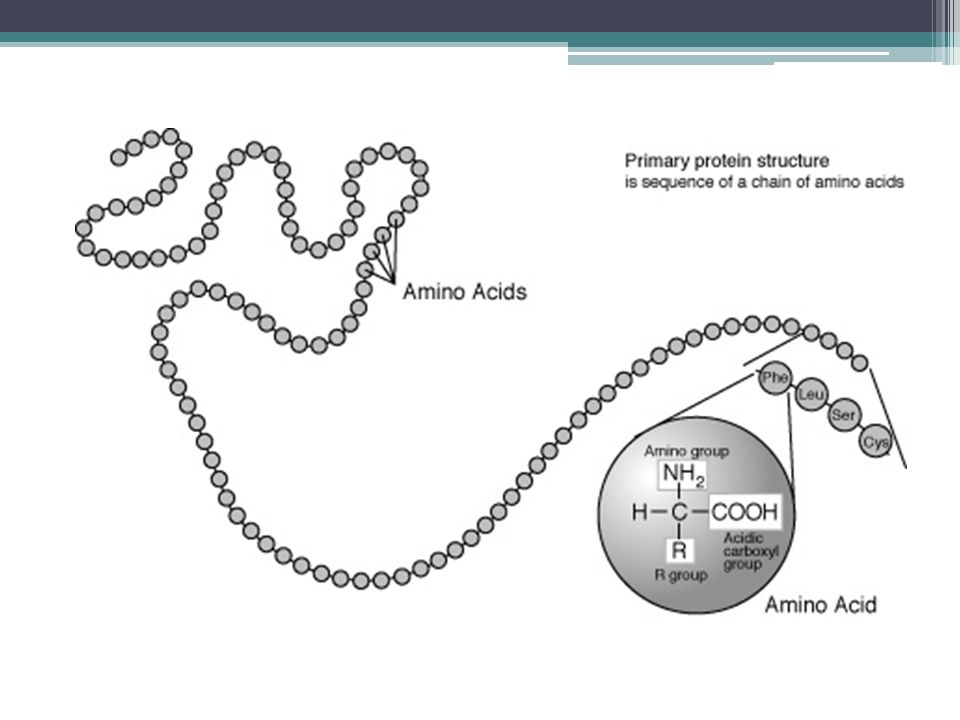
Aminoacidos Un aminoácido, como su nombre indica, es una molécula orgànica con un grupo amino (-NH2) y un grupo carboxilo (-COOH; ácido). Los aminoácidos más frecuentes y de mayor interés son aquellos que forman parte de las proteínas. Dos aminoácidos se combinan en una reacción de condensación que libera agua formando un enlace peptídico. Estos dos "residuos" aminoacídicos forman un dipéptido. Si se une un tercer aminoácido se forma un tripéptido y así, sucesivamente, para formar un polipéptido. Esta reacción ocurre de manera natural en los ribosomas, tanto los que están libres en el citosol como los asociados al retículo endoplasmático.
 y un grupo carboxilo (-COOH; ácido). Los aminoácidos más frecuentes y de mayor interés son aquellos que forman parte de las proteínas. Dos aminoácidos se combinan en una reacción de condensación que libera agua formando un enlace peptídico. Estos dos residuos aminoacídicos forman un dipéptido. Si se une un tercer aminoácido se forma un tripéptido y así, sucesivamente, para formar un polipéptido. Esta reacción ocurre de manera natural en los ribosomas, tanto los que están libres en el citosol como los asociados al retículo endoplasmático.")
22
Aminoacidos Todos los aminoácidos componentes de las proteínas son alfa-aminoácidos, lo que indica que el grupo amino está unido al carbono alfa, es decir, al carbono contiguo al grupo carboxilo. Por lo tanto, están formados por un carbono alfa unido a un grupo carboxilo, a un grupo amino, a un hidrógeno y a una cadena (habitualmente denominada R) de estructura variable, que determina la identidad y las propiedades de los diferentes aminoácidos; existen cientos de cadenas R por lo que se conocen cientos de aminoácidos diferentes, pero sólo 20 forman parte de las proteínas y tienen codones específicos
 de estructura variable, que determina la identidad y las propiedades de los diferentes aminoácidos; existen cientos de cadenas R por lo que se conocen cientos de aminoácidos diferentes, pero sólo 20 forman parte de las proteínas y tienen codones específicos.")
23
Aminoácidos "R" representa la cadena lateral, específica para cada aminoácido.

24
Aminoácidos Cuando los grupos amino y carboxilo de los aminoácidos se combinan para formar enlaces peptídicos, los aminoácidos constituyentes se denominan residuos de un aminoácido. Un péptido consiste en dos o más residuos de aminoácido unidos por enlaces peptídicos. Los péptidos con más de 10 residuos de aminoácidos se denominan polipéptidos.

25
Aminoácidos El orden de unión de los residuos diferentes tiene gran importancia ya que determina la proteína y consecuentemente su función. Los biopolímeros tienen propiedades distintivas muy diferentes a las unidades que las constituyen, así por ejemplo el almidón no es soluble en agua ni tiene sabor dulce, si bien es un polímero de la glucosa.

26
Aminoacidos esenciales
Valina (Val) Leucina (Leu) Treonina (Thr) Lisina (Lys) Triptófano(Trp) Histidina (His) Fenilalanina (Phe) Isoleucina (Ile) Arginina (Arg) (Requerida en niños y tal vez ancianos) Metionina (Met)
 Leucina (Leu) Treonina (Thr) Lisina (Lys) Triptófano(Trp) Histidina (His) Fenilalanina (Phe) Isoleucina (Ile) Arginina (Arg) (Requerida en niños y tal vez ancianos) Metionina (Met)")
27
Aminoacidos sintetizados por el cuerpo
Alanina (Ala) Prolina (Pro) Glicina (Gly) Serina (Ser) Cisteína (Cys) Asparagina (Asn) Glutamina (Gln) Tirosina (Tyr) Ácido aspártico (Asp) Ácido glutámico (Glu)
 Prolina (Pro) Glicina (Gly) Serina (Ser) Cisteína (Cys) Asparagina (Asn) Glutamina (Gln) Tirosina (Tyr) Ácido aspártico (Asp) Ácido glutámico (Glu)")
28
METABOLISMO DE LAS PROTEÍNAS

29
Metabolismo de Amino Acidos

30
USOS METABÓLICOS DE LOS AMINOÁCIDOS
Los aminoácidos absorbidos por la sangre, procedentes de los intestinos, se utilizan de muchas maneras en el cuerpo. Son nuestra principal fuente de nitrógeno, un elemento esencial para la vida. Las células utilizan también los aminoácidos para sintetizar las proteínas de los tejidos que se utilizan en la formación de células nuevas o para acondicionar las viejas. Se necesitan grandes cantidades de proteínas durante los períodos de crecimiento rápido o de larga convalecencia (quemaduras, hemorragias e infecciones). También los aminoácidos se usan en síntesis de otros aminoácidos, enzimas, hormonas, anticuerpos y compuestos no proteínicos que contienen nitrógeno como los ácidos nucleicos y los grupos hemo.
. También los aminoácidos se usan en síntesis de otros aminoácidos, enzimas, hormonas, anticuerpos y compuestos no proteínicos que contienen nitrógeno como los ácidos nucleicos y los grupos hemo.")
31
USOS METABÓLICOS DE LOS AMINOÁCIDOS
Los aminoácidos no se almacenan en el cuerpo, como los carbohidratos (en forma de glucógeno) y los lípidos (en forma de grasa). En cambio el cuerpo mantiene una reserva de aminoácidos, cuyo contenido cambia de manera constante (1- 2%), ya que las proteínas de los tejidos se degradan y sintetizan continuamente.
 y los lípidos (en forma de grasa). En cambio el cuerpo mantiene una reserva de aminoácidos, cuyo contenido cambia de manera constante (1- 2%), ya que las proteínas de los tejidos se degradan y sintetizan continuamente.")
32
BIOSÍNTESIS DE AMINOÁCIDOS
La Biosíntesis de aminoácidos comprende la síntesis proteínica, la degradación y conversión de los esqueletos del carbono de los aminoácidos a intermediarios anfibólicos, la síntesis de la urea, y la formación de una amplia variedad de compuestos funcionalmente activos, como la serotonina (neuro transmisor sintetizado en el tejido nervioso).
.")
33
BIOSÍNTESIS DE AMINOÁCIDOS
Para propósitos prácticos puede considerarse que existen 20 aminoácidos en las proteínas de los mamíferos. Si, durante la síntesis proteínica, falta uno solo de estos aminoácidos, la síntesis cesa.

34
BIOSÍNTESIS DE AMINOÁCIDOS
Los aminoácidos pueden ser sintetizados por el cuerpo, en un proceso llamado transaminación. La transaminación implica la transferencia de un grupo amino de un carbono a otro. El donador del grupo amino es un aminoácido y la molécula receptora es un - cetoácido (un ácido que contiene un grupo funcional cetona en el carbono , que es el carbono contiguo al grupo carboxilo)
")
35
BIOSÍNTESIS DE AMINOÁCIDOS
Los aminoácidos absorbidos por la sangre, procedentes de los intestinos, se utilizan de muchas maneras en el cuerpo. Son nuestra principal fuente de nitrógeno, un elemento esencial para la vida. Las células utilizan también los aminoácidos para sintetizar las proteínas de los tejidos que se utilizan en la formación de células nuevas o para acondicionar las viejas.

36
BIOSÍNTESIS DE AMINOÁCIDOS
Se necesitan grandes cantidades de proteínas durante los períodos de crecimiento rápido o de larga convalecencia (quemaduras, hemorragias e infecciones). También los aminoácidos se usan en síntesis de otros aminoácidos, enzimas, hormonas, anticuerpos y compuestos no proteínicos que contienen nitrógeno como los ácidos nucleicos y los grupos hemo.
. También los aminoácidos se usan en síntesis de otros aminoácidos, enzimas, hormonas, anticuerpos y compuestos no proteínicos que contienen nitrógeno como los ácidos nucleicos y los grupos hemo.")
37
BIOSÍNTESIS DE AMINOÁCIDOS
Los aminoácidos no se almacenan en el cuerpo, como los carbohidratos (en forma de glucógeno) y los lípidos (en forma de grasa). En cambio el cuerpo mantiene una reserva de aminoácidos, cuyo contenido cambia de manera constante (1- 2%), ya que las proteínas de los tejidos se degradan y sintetizan continuamente.
 y los lípidos (en forma de grasa). En cambio el cuerpo mantiene una reserva de aminoácidos, cuyo contenido cambia de manera constante (1- 2%), ya que las proteínas de los tejidos se degradan y sintetizan continuamente.")
38
BIOSÍNTESIS DE AMINOÁCIDOS
SÍNTESIS ESPECÍFICAS Los aminoácidos intervienen en la síntesis de muchos compuestos metabólicos. Los siguientes son solo algunos ejemplos. La tiroxina se utiliza para producir las hormonas adrenalina y noradrenalina y tiroxina, así como el pigmento de la piel llamado melanina. El triptófano se utiliza en la síntesis de la sustancia química llamada serotonina, que se relaciona con la transmisión nerviosa, y para sintetizar las coenzimas, NAD+ y NADP+. La serina se convierte en etanolamina que se encuentra en los lípidos, en la cisteína se utiliza en la síntesis de la sales biliares.

39
Síntesis de proteínas Las proteínas, por su tamaño, no pueden atravesar la membrana plasmática de la célula, por eso es que existe en su interior un mecanismo que las construye (síntesis) según las necesidades que tenga en ese momento la célula.
 según las necesidades que tenga en ese momento la célula.")
40
SÍNTESIS DE PROTEINAS Etapas de la biosíntesis de proteínas :
a) Transcripción b) Traducción Iniciación de la síntesis. Elongación de la cadena polipeptídica. Terminación de la síntesis.
 Transcripción. b) Traducción. Iniciación de la síntesis. Elongación de la cadena polipeptídica. Terminación de la síntesis.")
41
Síntesis de proteínas A) Transcripción:
Ocurre dentro del núcleo de las células eucariotas, aquí la secuencia de nucleótidos que denominamos gen (segmento de ADN que determina una proteína) se transcribe en una molécula de ARN.
 se transcribe en una molécula de ARN.")
42
Síntesis de proteínas Para formar la hebra de ARN a partir del ADN se debe tener en cuenta que cada nucleótido del ADN se ensambla con un determinado nucleótido del ARN. La molécula helicoidal de ADN se desenrolla y deja accesible la hebra paralela, a partir de la cual se inicia la síntesis (armado) del ARN. La enzima (polimesara del ARN) que controla la reacción detecta una región de la secuencia del ADN, llamada promotor, que marca el punto de inicio de la síntesis. La enzima se une al ADN en el sitio preciso para iniciar la síntesis de ARN y selecciona el primer nucleótido, que se convertirá en el extremo 5' de la cadena
 del ARN. La enzima (polimesara del ARN) que controla la reacción detecta una región de la secuencia del ADN, llamada promotor, que marca el punto de inicio de la síntesis. La enzima se une al ADN en el sitio preciso para iniciar la síntesis de ARN y selecciona el primer nucleótido, que se convertirá en el extremo 5 de la cadena.")
43
Síntesis de proteínas A continuación, se desplaza rápidamente por la cadena de ADN, añadiendo los nucleótidos correspondientes a la cadena de ARN. Según se forma, el ARN se va separando del ADN, comenzando por el extremo 5'; no obstante, hasta que no se llega al extremo 3' no se separa la molécula de ARN. Los nucleótidos se añaden uno por uno en orden complementario, de esta manera la adenina del ADN se combina con el uracilo del ARN (A – U), en el mismo orden, la timina se ensambla con la adenina (T – A), y la citosina se combina con la guanina y viceversa (C – G, G – C). Hay por lo tanto complementariedad entre el ARN y el ADN de donde se copia. Al conservar la información impresa en esta parte del genoma (dotación genética), el ARN se constituye en portador de las instrucciones que determinan la secuencia de aminoácidos de una proteína
, en el mismo orden, la timina se ensambla con la adenina (T – A), y la citosina se combina con la guanina y viceversa (C – G, G – C). Hay por lo tanto complementariedad entre el ARN y el ADN de donde se copia. Al conservar la información impresa en esta parte del genoma (dotación genética), el ARN se constituye en portador de las instrucciones que determinan la secuencia de aminoácidos de una proteína.")
44
Síntesis de proteínas Dichas instrucciones , se descifran leyendo los nucleótidos de tres en tres ("tripletes de nucleótidos“ o codon), y cada triplete de nucleótido determina uno de los 20 aminoácidos existentes- Durante la traducción, a medida que se "leen" los codones, se van añadiendo los aminoácidos correspondientes a la proteína que se está formando.
, y cada triplete de nucleótido determina uno de los 20 aminoácidos existentes- Durante la traducción, a medida que se leen los codones, se van añadiendo los aminoácidos correspondientes a la proteína que se está formando.")
45
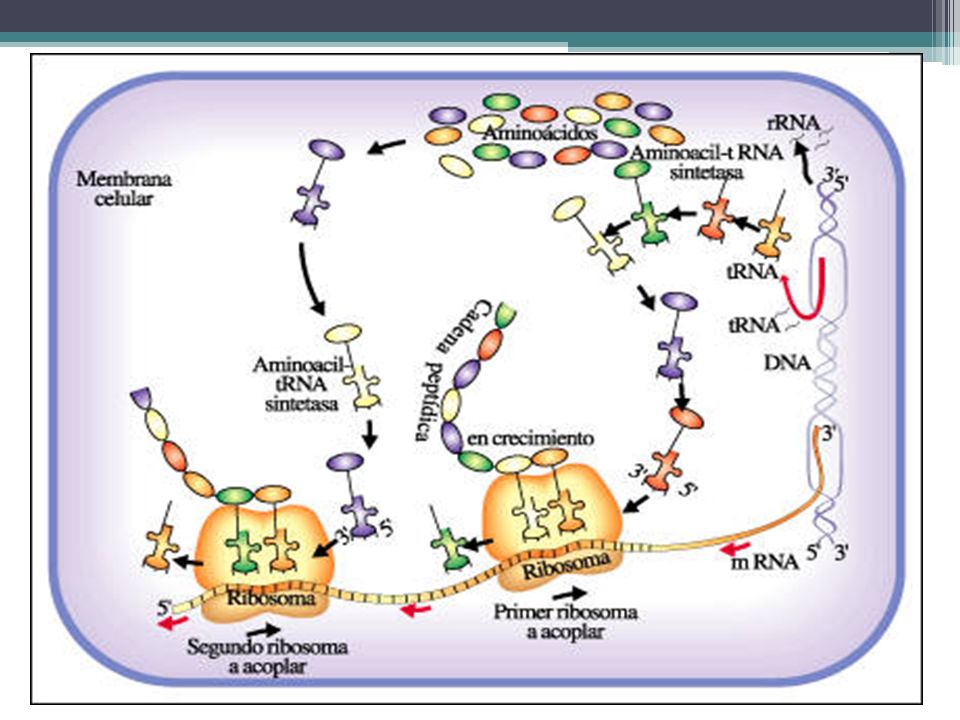
SÍNTESIS DE PROTEINAS La síntesis de proteínas o traducción tiene lugar en los ribosomas simples del citoplasma o los adheridos al retículo endoplasmático. En las Eucariotas la trasncripción ocurre en el nucleo Los aminoácidos son transportados por el ARN de transferencia (ARNt), específico para cada uno de ellos, hasta el citoplasma donde se aparea con el ARN mensajero (ARNm), donde se aparean el codón de éste y el anticodón del ARN de transferencia, por complementariedad de bases. Así, siguiendo la secuencia dictada originalmente por el DNA, las unidades de aminoácidos son alineadas una tras otra y, a medida que se forman los enlaces peptídicos entre ellas, se unen en una cadena polipeptídica.
, específico para cada uno de ellos, hasta el citoplasma donde se aparea con el ARN mensajero (ARNm), donde se aparean el codón de éste y el anticodón del ARN de transferencia, por complementariedad de bases. Así, siguiendo la secuencia dictada originalmente por el DNA, las unidades de aminoácidos son alineadas una tras otra y, a medida que se forman los enlaces peptídicos entre ellas, se unen en una cadena polipeptídica.")
46
SÍNTESIS DE PROTEINAS La molécula de tRNA es el adaptador (anticodon) que aparea el aminoácido correcto con cada codón de mRNA durante la síntesis de proteínas. Las enzimas conocidas como aminoacil-tRNA sintetasas catalizan la unión de cada aminoácido a su molécula de tRNA específica.
 que aparea el aminoácido correcto con cada codón de mRNA durante la síntesis de proteínas. Las enzimas conocidas como aminoacil-tRNA sintetasas catalizan la unión de cada aminoácido a su molécula de tRNA específica.")
47
Síntesis de proteínas El trabajo de los ARNt consiste en tomar del citosol a los aminoácidos y conducirlos al ribosoma en el orden marcado por los nucleótidos del ARNm, que son los moldes del sistema. La síntesis de las proteínas comienza con la unión entre sí de dos aminoácidos y continúa por el agregado de nuevos aminoácidos -de a uno por vez- en uno extremos de la cadena.

48
Síntesis de proteínas Cada tipo de ARNt lleva antepuesto el nombre del aminoácido que transporta: lisinil-ARNt para el de la lisina, fenilalanil-ARNt para el de la fenilalanina, metionil-ARNt para el de la metionina, etc. Por su lado el ARNt unido al aminoácido compatible con él se designa aminoacil-ARNtaá, en el que "aá" corresponde a la sigla del aminoácido. Por ejemplo, leucinil- ARNtLeu, lisinil-ARNtlys, fenilalanil-ARNtPhe, metionil-ARNtMet, etc

49
Síntesis de proteínas 2) Traducción :
Es la síntesis de proteína propiamente dicha) el ARN pasa del núcleo al citoplasma donde es traducida por los ribosomas que arman una proteína
 el ARN pasa del núcleo al citoplasma donde es traducida por los ribosomas que arman una proteína.")
50
SÍNTESIS DE PROTEINAS La síntesis de proteínas o traducción del ADN es el proceso anabólico mediante el cual se forman las proteína partir de los aa. Es el paso siguiente a la transcripción del ADN a ARN. Como existen veinte aminoácidos diferentes y sólo hay cuatro nucleótidos en el ARN (Adenina, Uracilo, Citosina y Guanina), es evidente que la relación no puede ser un aminoácido por cada nucleótido, ni tampoco por cada dos nucleótidos, ya que los cuatro tomados de dos en dos, sólo dan dieciséis posibilidades. La colinearidad debe establecerse como mínimo entre cada aminoácido y tripletes de nucleótidos. Los tripletes que codifican aminoácidos se denominan codones.
, es evidente que la relación no puede ser un aminoácido por cada nucleótido, ni tampoco por cada dos nucleótidos, ya que los cuatro tomados de dos en dos, sólo dan dieciséis posibilidades. La colinearidad debe establecerse como mínimo entre cada aminoácido y tripletes de nucleótidos. Los tripletes que codifican aminoácidos se denominan codones.")
51
Síntesis de proteínas Como se ha explicado, la clave de la traducción reside en el código genético, compuesto por combinaciones de tres nucleótidos consecutivos -o tripletes- en el ARNm. Los distintos tripletes se relacionan específicamente con tipos de aminoácidos usados en la síntesis de las proteínas. Cada triplete constituye un codón, existen en total 64 codones (cuatro nucleótidos se combinan de a tres, así que: 43 = 64), 61 de los cuales sirven para cifrar aminoácidos y 3 para marcar el cese de la traducción.
, 61 de los cuales sirven para cifrar aminoácidos y 3 para marcar el cese de la traducción.")
52
Síntesis de proteínas Dado que existen más codones que tipos de aminoácidos, casi todos pueden ser reconocidos por más de un codón, por lo que algunos tripletes a como "sinónimos". Solamente el triptófano y la metionina -dos de los aminoácidos menos frecuentes en las proteínas - son codificados, cada uno, por un solo codón. Generalmente los codones que decodifican a un mismo aminoácido se parecen entre sí y es frecuente que difieran sólo en el tercer nucleótido. Es importante destacar que el número de codones en el ARNm determina la longitud de la proteína.

53
Síntesis de proteínas Las moléculas intermediarias entre los codones del ARNm y los aminoácidos son los ARNt, los cuales tienen un dominio que se liga específicamente a uno de los 20 aminoácidos (en el extremo 3') y otro que lo hace, específicamente también, con el codón apropiado. El segundo dominio consta de una combinación de tres nucleótidos - llamado anticodón - que es complementaria de la del codón.
 y otro que lo hace, específicamente también, con el codón apropiado. El segundo dominio consta de una combinación de tres nucleótidos - llamado anticodón - que es complementaria de la del codón.")
54
Síntesis de proteínas La síntesis de proteínas o traducción del ARN es el proceso anabólico mediante el cual se forman las proteínas a partir de los aminoácidos. La Traducción se produce en el citoplasma donde se encuentran los ribosomas. Los Ribosomas están constituidos por una subunidad grande y pequeña que rodea el ARNm. En la subunidad menor algunas proteínas forman dos áreas: una al lado de la otra denominadas sitio P (por peptidil) y sitio A (por aminoacil).
 y sitio A (por aminoacil).")
55
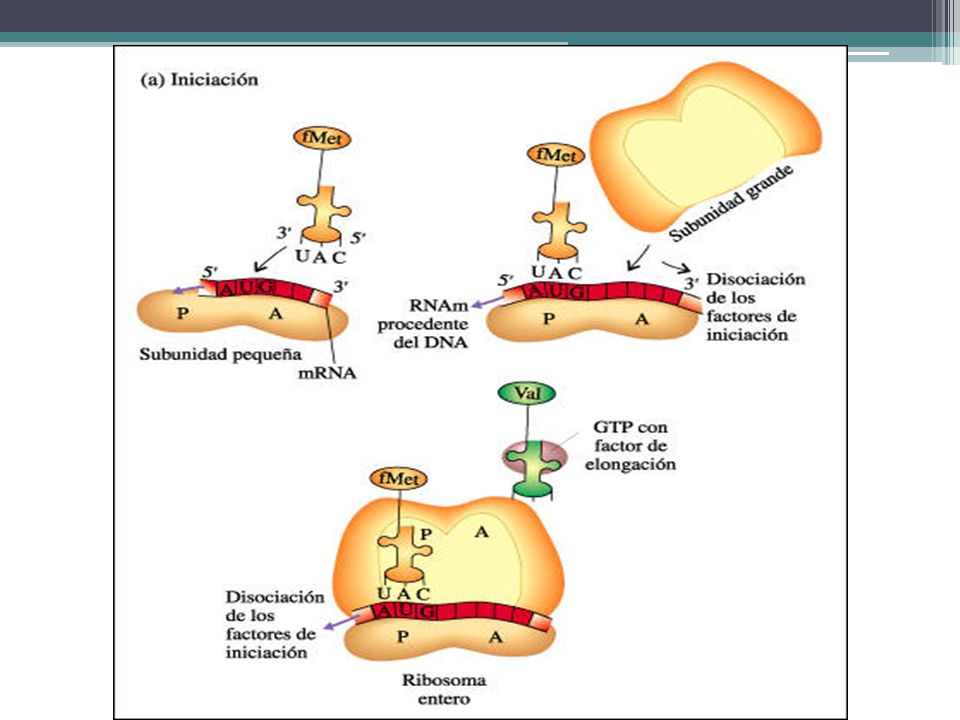
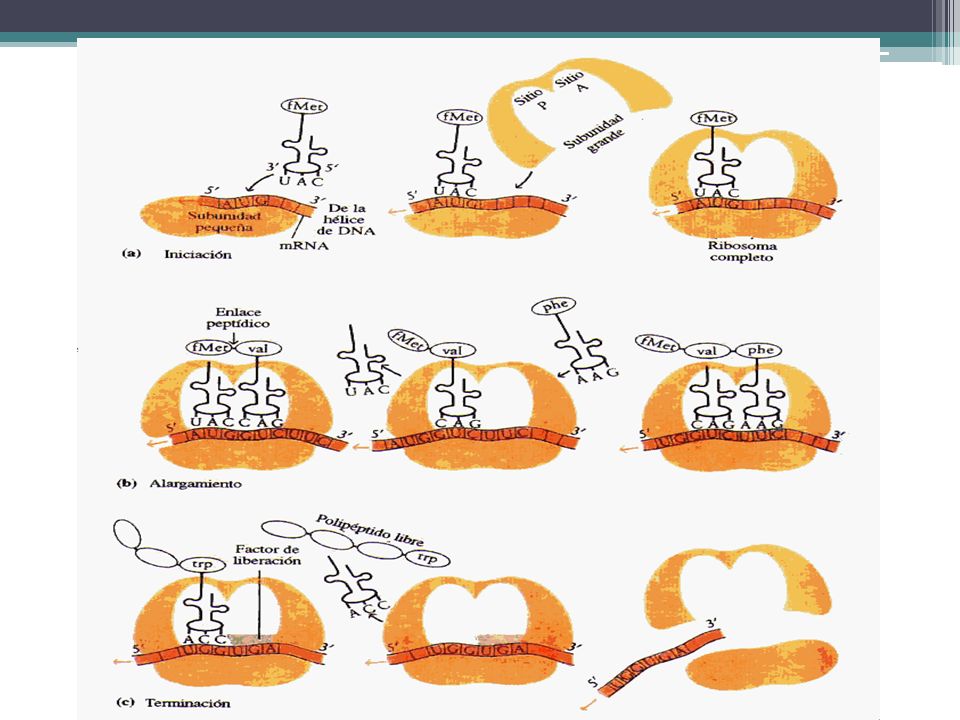
SÍNTESIS DE PROTEINAS Iniciación. La subunidad ribosómica más pequeña se une al extremo 5' de una molécula de mRNA. La primera molécula de tRNA, que lleva el aminoácido modificado fMet, se acopla según la complementaridad de las bases con el codón iniciador AUG de la molécula de mRNA. La subunidad ribosómica más grande se ubica en su lugar, el complejo tRNA-fMet ocupa el sitio P (peptídico). El sitio A (aminoacil) está vacante. El complejo de iniciación está completo ahora
. El sitio A (aminoacil) está vacante. El complejo de iniciación está completo ahora.")
57
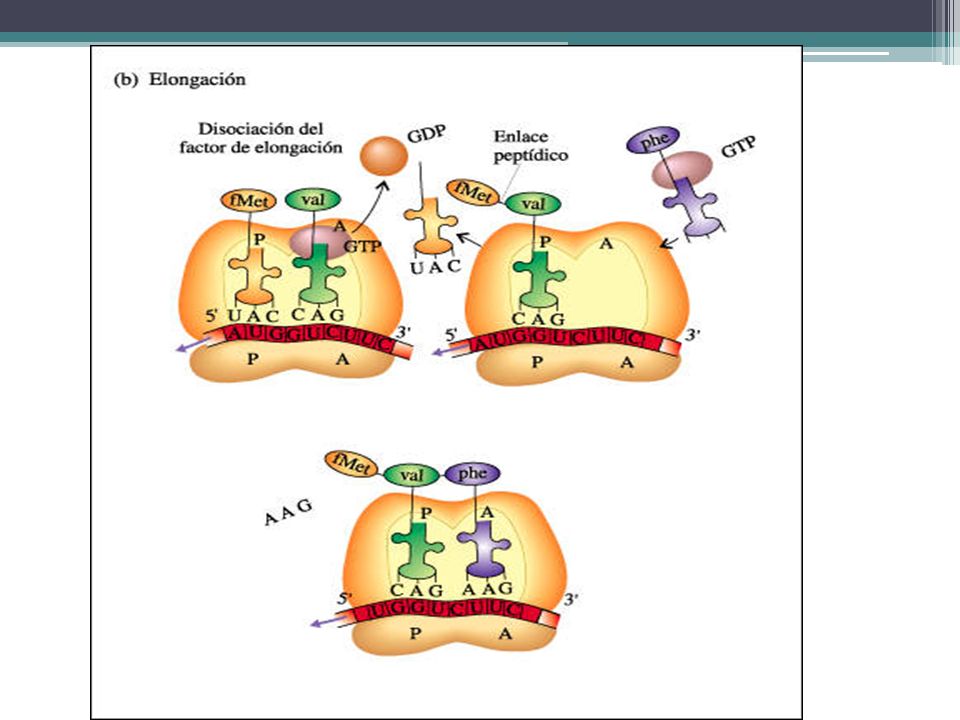
SÍNTESIS DE PROTEINAS Elongación de la cadena polipeptidica:
Un segundo tRNA, con su aminoácido unido, se coloca en el sitio A y su anticodón se acopla con el mRNA. Se forma un enlace peptídico entre los dos aminoácidos reunidos en el ribosoma. Al mismo tiempo, se rompe el enlace entre el primer aminoácido y su tRNA. El ribosoma se mueve a lo largo de la cadena de mRNA en una dirección 5' a 3', y el segundo tRNA, con el dipéptido unido, se mueve desde el sitio A al sitio P, a medida que el primer tRNA se desprende del ribosoma.

58
SÍNTESIS DE PROTEINAS El radical carboxilo (-COOH) del aminoácido iniciado se une con el radical amino (-NH2) del aminoácido siguiente mediante enlace peptídico. Esta unión es catalizada por la enzima peptidil-transferasa. El centro P queda pues ocupado por un ARNt sin aminoácido. El ARNt sin aminoácido sale del ribosoma. Se produce la translocación ribosomal.
 del aminoácido iniciado se une con el radical amino (-NH2) del aminoácido siguiente mediante enlace peptídico. Esta unión es catalizada por la enzima peptidil-transferasa. El centro P queda pues ocupado por un ARNt sin aminoácido. El ARNt sin aminoácido sale del ribosoma. Se produce la translocación ribosomal.")
59
SÍNTESIS DE PROTEINAS El dipeptil-ARNt queda ahora en el centro P. Todo ello es catalizado por los factores de elongación (FE) y precisa GTP. Según la terminación del tercer codón, aparece el tercer aminoacil-ARNt y ocupa el centro A. Luego se forma el tripéptido en A y posteriormente el ribosoma realiza su segunda translocación. Estos pasos se pueden repetir múltiples veces, hasta cientos de veces, según el número de aminoácidos que contenga el polipéptido. La traslocación del ribosama implica el desplazamiento del ribosama a lo largo de ARNm en sentido 5'-> 3'.
 y precisa GTP. Según la terminación del tercer codón, aparece el tercer aminoacil-ARNt y ocupa el centro A. Luego se forma el tripéptido en A y posteriormente el ribosoma realiza su segunda translocación. Estos pasos se pueden repetir múltiples veces, hasta cientos de veces, según el número de aminoácidos que contenga el polipéptido. La traslocación del ribosama implica el desplazamiento del ribosama a lo largo de ARNm en sentido 5 -> 3 .")
61
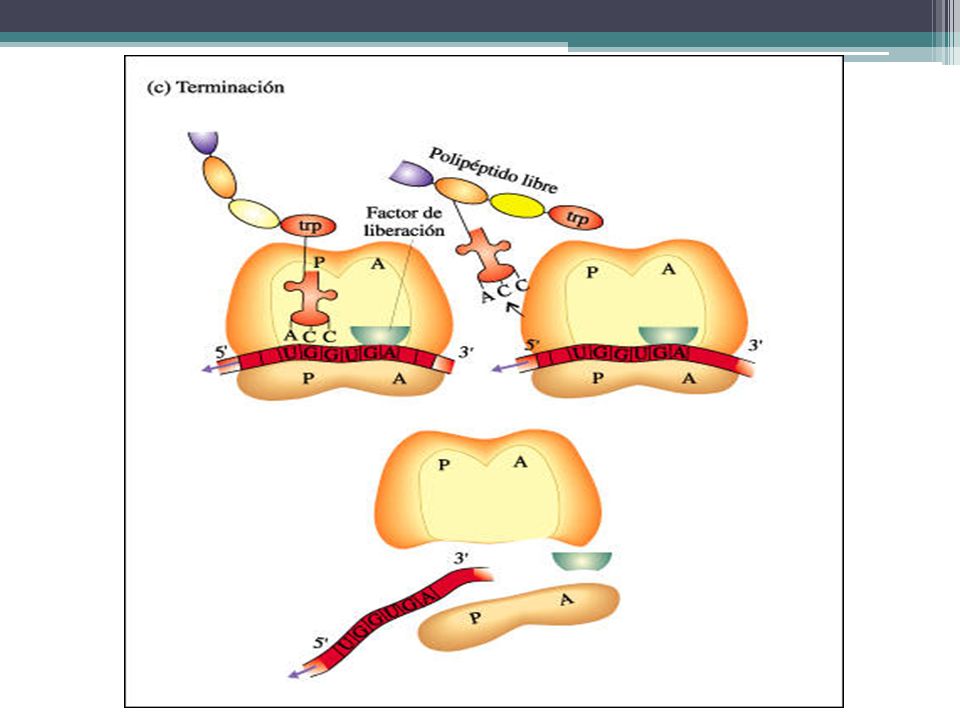
SÍNTESIS DE PROTEINAS Terminación de la síntesis de la cadena polipeptídica El final de la síntesis se presenta por los llamados tripletes sin sentido, también denominados codones stop. Son tres: UAA, UAG y UGA. No existe ningún ARNt cuyo anticodón sea complementario de ellos y, por lo tanto, la biosíntesis del polipéptido se interrumpe. Indican que la cadena polipeptídica ya ha terminado. Este proceso viene regulado por los factores de liberación, de naturaleza proteica, que se sitúan en el sitio A y hacen que la peptidil-transferasa separe, por hidrólisis, la cadena polipeptídica del ARNt.

65
Síntesis de proteínas Una vez finalizada la síntesis de una proteína, el ARN mensajero queda libre y puede ser leído de nuevo. De hecho, es muy frecuente que antes de que finalice una proteína ya está comenzando otra, con lo cual, una misma molécula de ARN mensajero, está siendo utilizada por varios ribosomas simultáneamente, esta estructura se conoce con el nombre de polirribosoma (polisoma).
.")
67
Síntesis de Proteínas

68
CATABOLISMO DE AMINOÁCIDOS:
Los aminoácidos que exceden las necesidades para la síntesis proteínica no pueden ser almacenados, ni excretados como tales. Los radicales aminos de los aminoácidos excedentes son eliminados son eliminados por transaminación o Desaminación oxidativa y los esqueletos de carbono son convertidos a intermediarios anfibólicos.

69
CATABOLISMO DE AMINOÁCIDOS
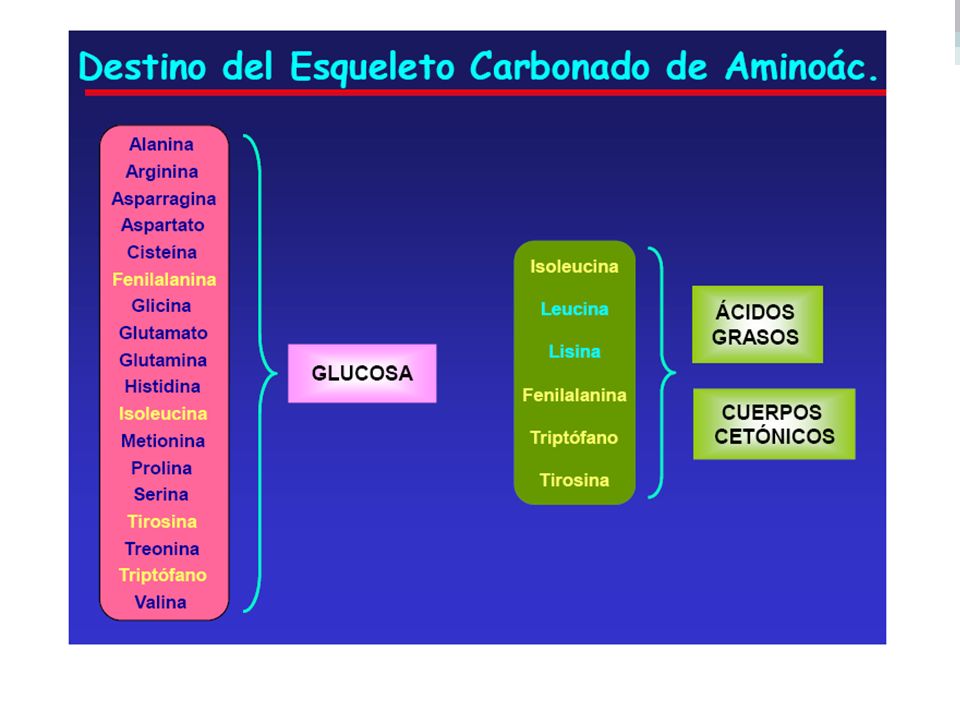
GLUCOGENICOS: precursores de carbohidratos. CETOGENICOS: precursores de lípidos. (Leu, Lys) AMINO ACIDOS QUE SON GLUCOGENICOS Y CETOGENICOS: ( Ile, Thr, Phe, Tyr, Trp)
 AMINO ACIDOS QUE SON GLUCOGENICOS Y CETOGENICOS: ( Ile, Thr, Phe, Tyr, Trp)")
70
CATABOLISMO DE AMINOÁCIDOS
Los aminoácidos están clasificados en esenciales y no esenciales. Los mamíferos pueden sintetizar los no esenciales, los esenciales deben adquirirlos de la dieta. El exceso de aminoácidos consumidos en la dieta, no puede ser almacenado para uso futuro, por el contrario, son transformados en intermediarios metabólicos comunes como el piruvato, oxaloacetato y alfa-cetoglutarato. Consecuentemente, los aminoácidos son precursores de glucosa, ácidos grasos y cuerpos cetónicos y por tanto son combustibles metabólicos

71
CATABOLISMO DE AMINOÁCIDOS
Un grupo pequeño de aminoácidos comprendidos por isoleucina, fenilalanina, treonina, triptófano, y tirosina dan lugar a precursores de la glucosa y de ácidos grasos y así son caracterizados como glucogénicos y cetogénicos. Finalmente, debe ser reconocido que los aminoácidos tienen un tercer posible destino. Durante etapas de hambruna los esqueletos de carbono reducidos se utilizan para la producción energética, con el resultado que se oxida a CO2 y H2O.

73
Catabolismo de amino ácidos

74
CATABOLISMO DE AMINOÁCIDOS:
El amoníaco es tóxico para el sistema nervioso central. El ácido úrico y sus sales son insolubles y precipitan en los tejidos y líquidos corporales cuando sus concentraciones exceden de varios mg/100 ml. Por lo tanto ninguno de estos productos finales de la degradación del nitrógeno es bien tolerado por los organismos superiores. Por ello han desarrollado la capacidad de convertir su excedente de nitrógeno en urea, compuesto sumamente soluble y no tóxico.

75
Biosíntesis de Aminoácidos no Esenciales Glutamato
El glutamato es sintetizado a partir de su α-ceto ácido precursor por una simple transaminacion reacción catalizada por el glutamato deshidrogenasa.

76
Biosíntesis de Aminoácidos no Esenciales. Aspartato
Como el glutamato, el aspartato es sintetizado por una simple 1-paso transaminacion reacción catalizado por aspartato aminotransferasa, AST.

77
Biosíntesis de Aminoácidos no Esenciales. Aspartato
El Aspartato también puede derivarse de asparragina a través de la acción de asparaginasa. La importancia de aspartato como precursor de ornitina para el ciclo de la urea es se describe en el metabolismo de nitrógeno.

78
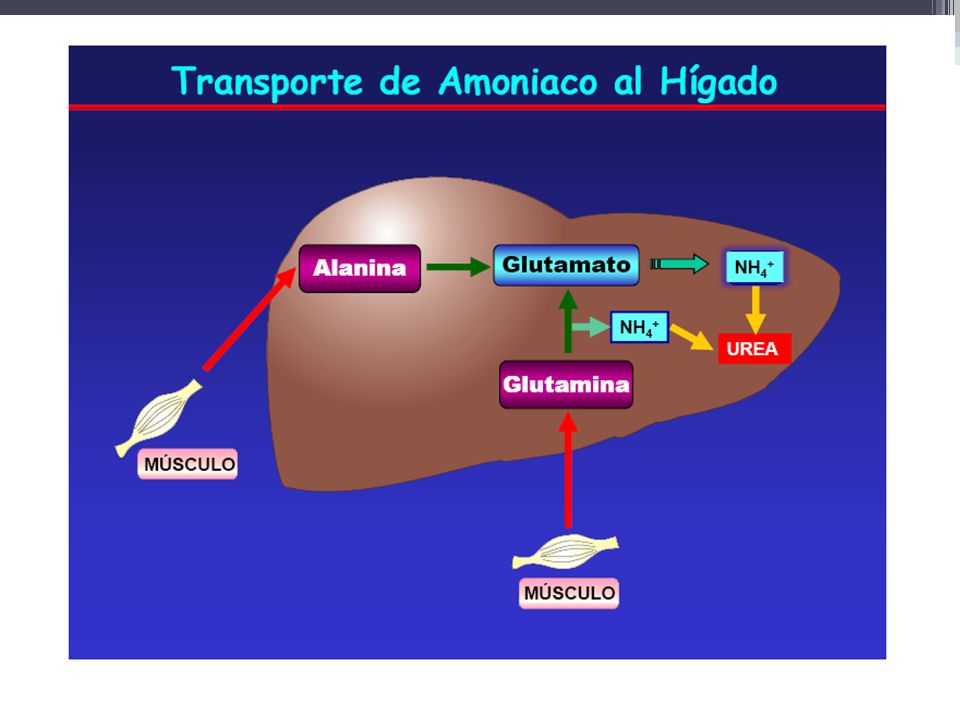
Alanina Hay 2 vías principales de producción de alanina muscular: directamente de la degradación de proteínas, y vía transaminación de piruvato por la alanina transaminasa, ALT (también designada como glutamato piruvato transaminasa sérica, SGPT).
.")
79
El ciclo de la glucosa-alanina se utiliza sobre todo como mecanismo del músculo esquelético para eliminar el nitrógeno al mismo tiempo que reabastece su suministro de energía

80
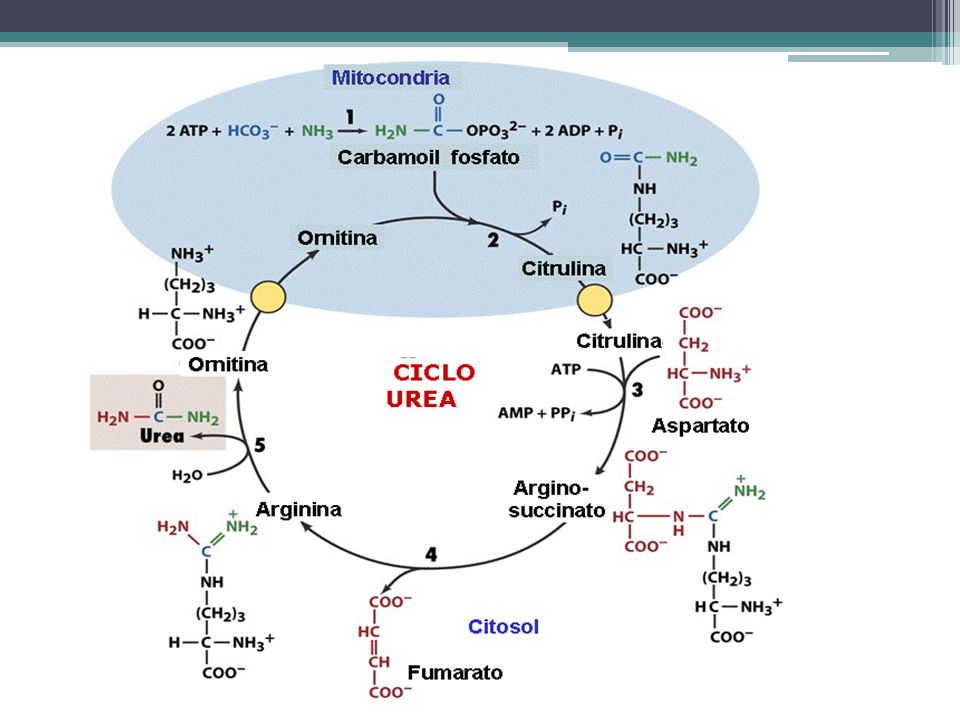
CICLO DE UREA Su importancia radica en que es el mecanismo más eficaz que dispone el organismo para la eliminación del amoníaco. La Urea es un compuesto de baja toxicidad. En este proceso 2 moléculas de amoníaco y una de CO2 son convertidas en urea. Es un proceso cíclico que consta de varias etapas que al final se obtiene arginina que al hidrolizarse libera Urea.

81
CICLO DE UREA Ocurre en el hígado
La síntesis de la Urea se lleva a cabo en el hígado y de este órgano alcanza al riñón, donde se elimina por la orina Cualquier situación que impida eliminar la urea por los riñones puede ser fatal (uremia).
.")
84
CICLO DE LA UREA El amoniaco que se forma en la Desaminación oxidativa, es toxico para célula; una concentración de 5mg/100ml de sangre es toxica para los seres humanos. El hígado transforma este amoniaco a urea, por medio de una serie de reacciones cíclicas llamadas ciclo de la urea o ciclo de Krebs para la ornitina. El amoníaco entra al ciclo como fosfato de carbamilo, y se une al aminoácido ornitina. Al completarse el ciclo, se forma una molécula de urea y se genera una molécula de ornitina para dar inicio al ciclo siguiente.

85
2 NH3 + CO2 + 3 ATP 3 H2O + Urea + 2 ADP + AMP + Pi
La ecuación de la reacción general es la siguiente. Reacción global de la síntesis de urea: 2 NH3 + CO2 + 3 ATP 3 H2O + Urea + 2 ADP + AMP + Pi Cinco reacciones enzimáticas: tres ocurren en el citosol y dos en la mitocondria. Mecanismos de transporte para citrulina y ornitina.

86
ENZIMAS DEL CICLO DE LA UREA:
CPS I (carbamilfosfato cintasa) TRANSCARBAMILASA DE ORNITINA SINTASA DE ARGININOSUCCINATO LIASA DE ARGININOSUCCINATO arginasa
 TRANSCARBAMILASA DE ORNITINA. SINTASA DE ARGININOSUCCINATO. LIASA DE ARGININOSUCCINATO. arginasa.")
87
CICLO DE LA UREA En resumen el amoniaco producido constantemente por los tejidos es transformado en glutamato, glutamina y urea. En gran medida la toxicidad del amoniaco se debe a la formación de glutamato y glutamina, lo que produce un déficit de a-cetoglutarato del ciclo de Krebs e inhibición de la respiración celular.

88
CICLO DE LA UREA El transporte de amoniaco desde otros tejidos al hígado o riñón se realiza como glutamina, gracias a la glutamina sintetasa que transforma el glutamato en glutamina, reacción muy importante en la detoxificación del cerebro. En el hombre entre 80 y 90% del N es eliminado como urea, cuya formación es dependiente de ATP, el cuál es también activador de la glutamato deshidrogenasa. En la síntesis de urea (H2N-CO-NH2), el C proviene del CO2 y los grupos NH2 son aportados por glutamato y aspartato.
, el C proviene del CO2 y los grupos NH2 son aportados por glutamato y aspartato.")
90
SÍNTESIS DE UREA La biosíntesis de la urea puede convencionalmente dividirse para su estudio en las siguientes etapas. A) Transaminación B) Desaminación Oxidativa C) Transporte de amonio D) Reacciones del ciclo de la urea
 Transaminación. B) Desaminación Oxidativa. C) Transporte de amonio. D) Reacciones del ciclo de la urea.")
91
Transaminación Es este un proceso, realizado en el citosol y en las mitocondrias, por el que un aminoácido se convierte en otro. Se realiza por medio de transaminasas que catalizan la transferencia del grupo alfa-amino (NH3+) de un aminoácido a un alfa-cetoácido, tal como piruvato, oxalacetato o más frecuentemente alfa-cetoglutarato. Consecuentemente se forma un nuevo aminoácido y un nuevo cetoácido.
 de un aminoácido a un alfa-cetoácido, tal como piruvato, oxalacetato o más frecuentemente alfa-cetoglutarato. Consecuentemente se forma un nuevo aminoácido y un nuevo cetoácido.")
92
Fig. 2 Transaminación (citosol)
")
93
aminoácido + enzima -ceto ácido + E-NH2
La reacción de aminotransferasa ocurre en dos pasos: 1.- el grupo amino de un aminoácido es transferido a la enzima, produciendo el correspondiente -ceto ácido y la enzima aminada aminoácido + enzima -ceto ácido + E-NH2

94
2. - el grupo amino es transferido al -ceto ácido aceptor (ej
2.- el grupo amino es transferido al -ceto ácido aceptor (ej. Alfa-cetoglutarato) formando el producto aminoácido (ej. Glutamato) y regenerando la enzima -cetoglutarato + enzima-NH2 enzima + glutamato
 formando el producto aminoácido (ej. Glutamato) y regenerando la enzima. -cetoglutarato + enzima-NH2 enzima + glutamato.")
95
A) Transaminación La transaminación, es catalizada por las enzimas llamadas: transaminasas o aminotransferasas, implica la ínter conversión de un par de aminoácidos y un par de cetoácidos. Estos generalmente son -amino y -cetoácidos. Los -cetoácidos son estados transitorios de los aminoácidos

96
Transaminación El fosfato piridoxal forma un parte esencial del ciclo activo de transaminasas y de muchas otras enzimas como aminoácido como sustratos. En todas las reacciones de los aminoácidos, dependiente del fosfato piridoxal, en fase inicial es la formación de una base intermediaria unida a la enzima. Este intermediario, estabilizado por la acción reciproca por una región catiónica del ciclo activo, se puede reestructurar de manera que incluya la liberación de un cetoácido con formación de fosfato de piridoxamina unido a la enzima..

97
Transaminación La forma amino unida de la coenzima puede formar entonces una base análoga intermediaria con un acetoácido. Durante la transaminación la coenzima unida sirve como un transportador de grupo amino, la transaminación es un proceso libremente reversible lo cual le permite a las transaminasas funcionar tanto en el catabolismo de los aa como en la síntesis

98
Transaminación Cada transaminasa es especifica para el par especifico del aminoácido y cetoácido como un par de sustrato, pero inespecífica para el otro par, puesto que la alanina también un sustrato para la glutamato transaminasa todo el nitrógeno amino proveniente de los aminoácido que pueden experimentar la transaminación se pueden concentrar en el glutamato, siendo el glutamato el único aminoácido de los tejido del mamífero que puede experimentar desaminación oxidativa en una tasa apreciable. La mayor parte de los aminoácidos son sustrato para la transaminación. La transaminación forma glutamatos que transportan los grupos aminos

99
Transaminación La importancia de la transaminación es que forma glutamato que es el producto que se utiliza para la siguiente etapa. Este proceso se utiliza en citoplasma, debiendo el glutamato pasar a Desaminación oxidativa en el interior de la mitocondria.

100
Figura 3. Desaminación oxidativa
Aminoácido -cetoácido

101
Muchos de los aminoácidos son desaminados por transaminación
Muchos de los aminoácidos son desaminados por transaminación. En este proceso, se transfiere el grupo amino a un -ceto ácido para dar el -ceto ácido del aminoácido original y un nuevo aminoácido; estas reacciones son catalizadas por aminotransferasas o transaminasas, El aceptor principal de grupos amino es el -cetoglutarato que produce glutamato como nuevo aminoácido

102
El grupo amino del glutamato es transferido al oxaloacetato en una segunda reacción de transaminación dando aspartato:

103
Desaminación oxidativa
La transaminación no resulta en ninguna desaminación neta. La desaminación ocurre a través de la desaminación oxidativa del glutamato por la glutamato deshidrogensa que produce amonio. La reacción requiere de NAD+ o NADP+ y regenera -cetoglutarato para transaminaciones adicionales:

104
Desaminación oxidativa
La primera reacción en la ruptura de los aminoácidos es casi siempre la remoción de su grupo -amino con el objeto de excretar el exceso de nitrógeno y degradar el esqueleto de carbono restante. La urea, es el producto principal de la excreción de nitrógeno en los mamíferos terrestres se sintetiza a partir de amonio y aspartato. Ambas substancias son derivadas principalmente del glutamato, el producto principal de las reacciones de desaminación.

105
Desaminación oxidativa
Proceso, realizado en las mitocondrias, y en el que la enzima ácido glutámico-deshidrogenasa elimina el grupo amino del ácido glutámico. Se forma amoníaco que entra en el ciclo de la urea y los esqueletos carbonados vienen a ser productos intermedios glucolíticos y del ciclo de Krebs.

106
B) Desaminación oxidativa
Los aminoácidos que no se utilizan en procesos sintéticos pueden catabolizarse o degradarse para obtener amoníaco, CO2, agua y energía. Cuando lo anterior tiene lugar el grupo amino de los aminoácidos es removido, mediante un proceso llamado desaminación oxidativa, y entra al ciclo de la urea, lo que queda de la molécula llamado esqueleto hidrocarbonado del aminoácido, puede entrar al ciclo del ácido cítrico para suministrar la energía celular necesaria a convertirse en grasa corporal. Esto quiere decir que es posible engordar comiendo demasiada proteína, como lo es comiendo demasiados lípidos y carbohidratos.

107
Desaminación oxidativa
Los -acetoácido que se forma en la Desaminación oxidativa pueden ser utilizados de varias maneras. Se pueden oxidar en el ciclo del ácido cítrico para producir energía o convertirse en otros aminoácidos mediante las transaminación. Estos -cetoácidos pueden utilizarse así mismo en la síntesis de carbohidratos y grasas. La conversión oxidativa de muchos aminoácidos en sus correspondientes -cetoácidos ocurre en el hígado y riñón de los mamíferos. En ella participan fundamentalmente las enzimas: transaminasas y L- glutamato deshidrogenasa.

108
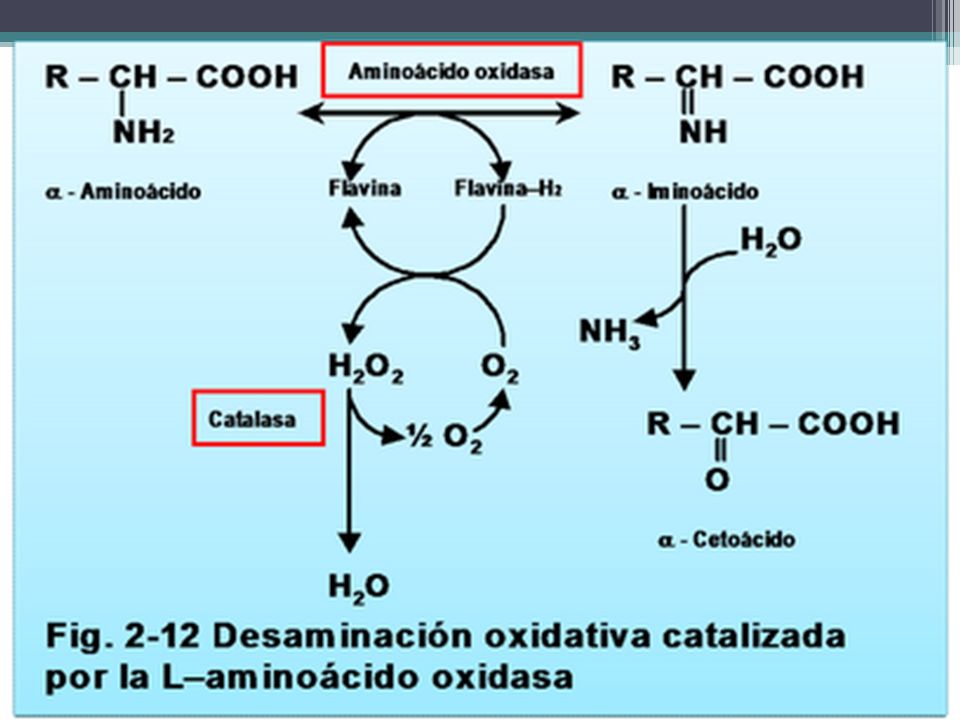
Desaminación oxidativa catalizada por la aminoácido oxidasas
Es una desaminación oxidativa que ocurre en un solo paso. Ocurre en el hígado y el riñón.

109
Desaminación oxidativa
Desaminación oxidativa catalizada por la aminoácido oxidasas Las aminoácido oxidasas son flavo-proteínas auto- oxidables, es decir, el FAD reducido es reoxidado directamente por el nitrógeno molécular, formando peróxido de hidrógeno el cuál es un producto tóxico que debe ser desdoblado en O2 y H2 por la enzima catalasa. En las reacciones de la aminoácido oxidasa, el aminoácido es deshidrogenado primero por la flavo- proteína de la oxidasa, formando un -aminoácido. Este adiciona agua espontáneamente y luego se descompone en el correspondiente -cetoácido con pérdida de nitrógeno -imino como ión amonio

111
Hay dos aminoácido oxidasas no específicas la L-aminoácido oxidasa y la D-aminoácido oxidasa, estas enzimas catalizan la oxidación de D y L aminoácidos respectivamente, utilizando FAD como coenzima redox (en vez de NAD(P)+). El FADH2 resultante es reoxidado por el oxígeno molecular. Aminoácido + FAD + H20 -ceto ácido + NH3 + FADH2 FADH2 + O2 FAD + H202

112
Desaminación oxidativa
Desaminación oxidativa catalizada por L- glutamato deshidrogenasa. Los grupos amino de la mayor parte de los aminoácidos son transferidos en último termino, al -cetoglutarato por .transaminación formando L-glutamato. La liberación de este nitrógeno como amoníaco es catalizada por la L-glutamato. La liberación de este nitrógeno como amoniaco es catalizada por la enzima L- glutamato deshidrogenasa, cuya actividad es afectada por modificadores alostéricos como el ATP, GTP, NADHA, que inhibe a la enzima , y el ADP que la activa.

113
Desaminación oxidativa
Glutamato deshidrogenasa usa NAD+ o NADP + como substrato. La reacción es irreversible y funciona tanto en el catabolismo de los aminoácidos como en su biosíntesis. Por consiguiente, funciona no solo catalizando el nitrógeno del glutamato hacia urea (catabolismo), sino también catalizando la aminación del cetoglutarato por el amoniaco libre.
, sino también catalizando la aminación del cetoglutarato por el amoniaco libre.")
114
El glutamato es desaminado oxidativamente en la mitocondria por la glutamato deshidrogenasa, la única enzima conocida que al menos en algunos organismos, puede trabajar con NAD+ o NADP+ como coenzima redox. Se piensa que la oxidación ocurre con la transferencia de un ion hidruro del carbono del glutamato al NAD(P)+ formando -iminoglutarato el cual es hidrolizado a -cetoglutarato y amonio. La enzima es inhibida por GTP y activada por ADP in vitro lo que sugiere la regulan también in vivo.
+ formando -iminoglutarato el cual es hidrolizado a -cetoglutarato y amonio. La enzima es inhibida por GTP y activada por ADP in vitro lo que sugiere la regulan también in vivo..")
115
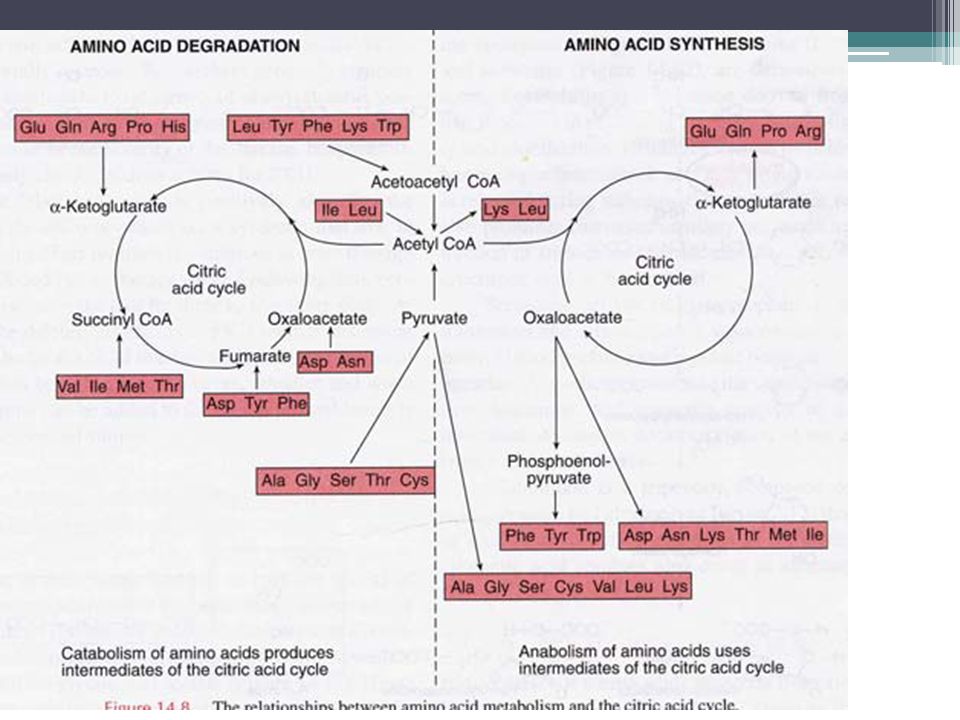
Productos de la desaminación de aa
Aminoácido(s) Producto Ile, Leu, Lys Acetil-CoA Tyr, Phe Acetoacetato Gln, Pro, Arg Glu y alfa-cetoglutarato His Glu y alfa-cetoglutarato Thr, Met , Val Succinil-CoA Tyr, Phe, Asp Fumarato Asp, Asn Oxaloacetato Ser, Gly, Cys Piruvato Trp Alanina y piruvato
 Producto Ile, Leu, Lys Acetil-CoA Tyr, Phe Acetoacetato Gln, Pro, Arg Glu y alfa-cetoglutarato His Glu y alfa-cetoglutarato Thr, Met , Val Succinil-CoA Tyr, Phe, Asp Fumarato Asp, Asn Oxaloacetato Ser, Gly, Cys Piruvato Trp Alanina y piruvato")
116
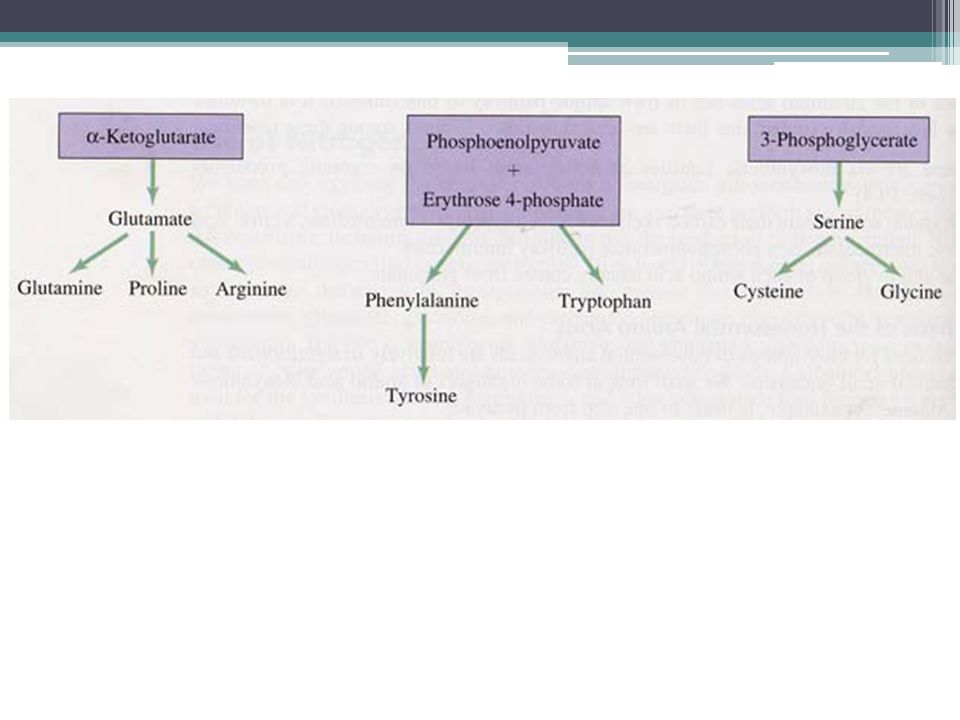
Sintesis de amino ácidos
Está unida al ciclo del ácido tricarboxílico (TCA), bien por transaminación o bien por fijación de amonio. El grupo alfa-amino es central a toda síntesis de aminoácidos y deriva del amonio de los grupos aminos del L-glutamato. De éstos se sintetizan glutamina, prolina y arginina. El ácido glutámico es la principal fuente de los grupos amino para la transaminación La cisteína se forma, en el citosol celular, a partir de serina y del aminoácido esencial metionina.
, bien por transaminación o bien por fijación de amonio. El grupo alfa-amino es central a toda síntesis de aminoácidos y deriva del amonio de los grupos aminos del L-glutamato. De éstos se sintetizan glutamina, prolina y arginina. El ácido glutámico es la principal fuente de los grupos amino para la transaminación La cisteína se forma, en el citosol celular, a partir de serina y del aminoácido esencial metionina.")
117
C) Formación y transporte de amoniaco
La producción de amoniaco por los riñones está marcadamente aumentada en la acidosis metabólica y deprimida en la alcalosis. Este amoniaco no solo se deriva de la urea, sino también de los aminoácidos intracelulares, particularmente de la glutamina. La liberación de amoniaco es catalizada por la glutaminasa renal.

118
Formación y transporte de amoniaco
Aunque el amoniaco puede ser excretado como sales de amonio, la mayor parte es excretada por urea. El principal componente nitrogenado de la orina. El amoniaco es producido constantemente en los tejidos pero en la sangre solo se encuentran concentraciones de 10 a 20 gr/100mL puesto que es rápidamente eliminado de la circulación por el hígado y convertido ya sea en glutamato, glutamina, o urea.

119
Formación y transporte de amoniaco
La formación de glutamina es catalizada por la glutaminsintasa. La síntesis de en enlace amídico de la glutamina se lleva a acabo de expensa de la hidrólisis de un equivalente de ATP en ADP y Pi.

120
Formación y transporte de amoniaco
La liberación del nitrógeno amídico de la glutamina como amoniaco sucede no por reversión dela reacción de la glutaminsintasa, sino por formación hidrolítica de amoniaco o catalizada por la glutaminasa. La reacción de la glutaminasa a diferencia de la reacción de la glutaminsintasa, no incluye la participación de nucleótido de adinina, favorece fuertemente la formación del glutamato y no funciona en la síntesis del glutamina. La glutaminsintasa y la glutaminasa sirven para catalizar interconversión del ion amonio libre y la glutamina.

121
Formación y transporte de amoniaco
Mientras que en encéfalo el mecanismo principal para la liberación del amoniaco es la formación de glutamina, en el hígado la vía más importante es la formación de urea. En la figura siguiente se muestra la formación hodrólitica del amoniaco catalizada por la enzima glutaminasa. La reacción de la glutaminasa procede esencialmente de manera irreversible en dirección de la formación de glutamato y radical NH4+.

122
Figura 5. Formación hodrólitica del amoniaco catalizada por la enzima glutaminasa

123
Figura 6. Reacción de la glutaminsintasa.

124
D) Reacciones del ciclo de la urea
Las reacciones de los intermediarios el la biosíntesis de un mol de urea a partir de un mol de amoniaco y otra a partir de CO2 (activados con Mg2+, y ATP), así como el nitrógeno alfa amino se muestran en la figura del ciclo de la urea. El proceso global requiere 3 moles de ATP (dos de los cuales son convertidos en ADP+Pi, y una en AMP+PPi) y la participación sucesiva de 5 enzimas que catalizan las reacciones del ciclo
, así como el nitrógeno alfa amino se muestran en la figura del ciclo de la urea. El proceso global requiere 3 moles de ATP (dos de los cuales son convertidos en ADP+Pi, y una en AMP+PPi) y la participación sucesiva de 5 enzimas que catalizan las reacciones del ciclo.")
125
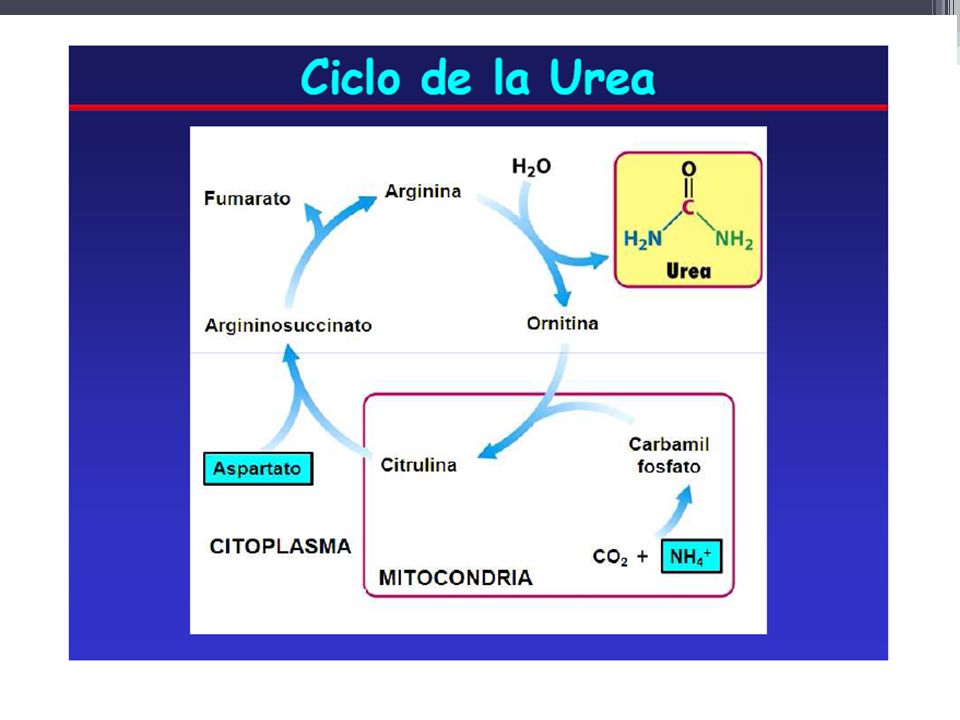
Reacciones del ciclo de la urea
De los 6 aminoácidos que intervienen en la síntesis de la urea, uno el N-acetil-glutamato, funciona como un activador enzimático y no como un intermediario. Los cinco restantes (aspartato, arginina, ornitina, citrulina y argininsuccinato) funcionan todos como transportadores de átomos que en último término se vuelven urea. El aspartato y arginina existen en las proteínas, mientras que la ornitina, citrulina y argininsuccinato, no
 funcionan todos como transportadores de átomos que en último término se vuelven urea. El aspartato y arginina existen en las proteínas, mientras que la ornitina, citrulina y argininsuccinato, no.")
126
Reacciones del ciclo de la urea
El principal papel metabólico de estos tres últimos aminoácidos es la síntesis de la urea (en mamíferos). Nótese que la formación de urea es, un proceso cíclico. La ornitina usada en la reacción número dos es regenerada en la reacción número cinco. Así, no hay pérdida ni ganancia neta de ornitina, citrulina, argininsuccinato, o de arginina durante la síntesis de la urea; sin embargo el amoniaco el CO2, ATP, y el aspartato si son consumidas.
. Nótese que la formación de urea es, un proceso cíclico. La ornitina usada en la reacción número dos es regenerada en la reacción número cinco. Así, no hay pérdida ni ganancia neta de ornitina, citrulina, argininsuccinato, o de arginina durante la síntesis de la urea; sin embargo el amoniaco el CO2, ATP, y el aspartato si son consumidas.")
127
Reacciones del ciclo de la urea
La ornitina puede atravesar la membrana intercelular donde se forma la citrulina, luego sale de mitocondrias a citoplasma (los aminoácidos se hallan siempre en citosol), para formar ácido arginosuccínico, luego ácido fumárico, y finalmente arginina que libera la ornitina y una molécula de urea.
, para formar ácido arginosuccínico, luego ácido fumárico, y finalmente arginina que libera la ornitina y una molécula de urea.")
128
Ciclo de la urea

129
Reacciones del ciclo de la urea
Reacción 1: Síntesis de carbamilfosfato La condensación de un mol de amoniaco de otro de CO2 y de una de fosfato (derivado del ATP) para formar carbamilfosfato es catalizada por la carbamilfosfato cintasa. Los dos moles de ATP hidrolizado durante esta reacción a portan la energía necesaria durante la síntesis de dos enlaces covalente de carbamilfosfato
 para formar carbamilfosfato es catalizada por la carbamilfosfato cintasa. Los dos moles de ATP hidrolizado durante esta reacción a portan la energía necesaria durante la síntesis de dos enlaces covalente de carbamilfosfato.")
130
Reacción 1: Síntesis de carbamilfosfato

131
Reacciones del ciclo de la urea
Reacción 2: Síntesis de citrulina La transferencia de la fracción carbamilo del carbamilfosfato a la ornitina, formando citrulina + PI, es cataliza por la L-ornitina transcarbamilasa de las mitocondrias del hígado. La reacción es altamente especifica para la ornitina y el equilibrio favorece grandemente la síntesis de la citrulina.

132
Reacción 2: Síntesis de citrulina

133
Reacciones del ciclo de la urea
Reacción 3: Síntesis del argininsuccinato En la reacción del argininsuccinato cintasa, el aspartato y la citrulina se unen por medio del grupo amino del aspartato la reacción requiere del ATP y el equilibrio favorece fuertemente la síntesis del argininsuccinato.

134
Reacción 3: Síntesis del argininsuccinato

135
Reacciones del ciclo de la urea
Reacción 4: Desdoblamiento del argininsuccinato en arginina y fumarato El desdoblamiento reversible del argininsuccinato en arginina y fumarato es cataliza por la argininsuccinasa. La reacción se lleva a cabo por un mecanismo de transeliminación. El fumarato formado puede ser convertido en oxalacetato mediante la reacciones de la fumarasa y de la malato deshidrogenasa y luego transaminado éste para regenerar el aspartato.

136
Reacción 4: Desdoblamiento del argininsuccinato en arginina y fumarato

137
Reacciones del ciclo de la urea
Reacción 5: Desdoblamiento de la arginina en ornitina y urea Esta reacción completa el ciclo de la urea y regenera la ornitina, sustrato de la reacción 2. el desdoblamiento hidrolítico del grupo guanidínico de la arginina es catalizado por la arginasa. La arginasa altamente purificada preparada el hígado de mamíferos es activada por el Co2+ o el Mn2+. La ornitina la lisina son potentes inhibidores que compiten con la arginina.

138
Reacción 5: Desdoblamiento de la arginina en ornitina y urea

140
Conexiones entre los ciclo de Krebbs y de la urea

141
Conexiones entre los ciclo de Krebbs y de la urea
1. El fumarato producido en la reacción de la argininosuccinato liasa, ingresa a la mitocondria, donde es blanco de la fumarasa y malato deshidrogenasa para formar oxalacetato. 2. El aspartato que actúa como dador de N en el ciclo de la urea se forma a partir del oxalacetato por transaminación desde el glutamato. 3. Dado que las reacciones de los dos ciclos están interconectados se les ha denominado como doble ciclo de Krebs

142
REGULACIÓN DEL CICLO El flujo del N a través del ciclo de la urea dependerá de la composición de la dieta. Una dieta rica en proteínas aumentará la oxidación de los aminoácidos, produciendo urea por el exceso de grupos aminos, al igual que en una inanición severa

143
Ciclo de la urea

144
DIGESTIÓN DE LAS PROTEÍNAS

145
DIGESTIÓN DE PROTEÍNAS
La digestión de las proteínas inicia en el estómago. El pH ácido de las secreciones gástricas activa a la enzima pepsina, la cual comienza la desintegración de proteínas en pequeños fragmentos llamados polipéptidos. Como la pepsina digiere las proteínas en las células que las elaboran, debe ser sintetizada en una forma inactiva denominada pepsinógeno, el cual se convierte en pepsina por la acción del ácido clorhídrico antes de ser segregada al estómago. La mucosidad estomacal protege a este órgano de la acción del ácido y la pepsina.

146
DIGESTIÓN DE PROTEÍNAS
Pero la mayor parte de la digestión de las proteínas se realiza en los intestinos y se debe a dos enzimas pancreáticas: tripsina y quimo tripsina. A semejanza de la pepsina, han de ser sintetizadas en forma s inactivas, tripsinógeno y quimotripsinógeno, ya que de lo contrario digerirían las proteínas de las células que las elaboran. La enterocinasa, una enzima sintetizada en el intestino, activa la tripsina, que a su vez hace lo mismo con la quimo tripsina.

147
DIGESTIÓN DE PROTEÍNAS
Después ambas prosiguen la desintegración de las proteínas y de grandes polipéptidos convirtiéndolos así en dipéptidos ( unión de dos aminoácidos) y tripéptidos (unión de tres aminoácidos). Unos y otros pasan por la digestión ulterior realizada por las enzimas de las micro vellosidades, y los aminoácidos libres así producidos son llevados a la corriente sanguínea.
 y tripéptidos (unión de tres aminoácidos). Unos y otros pasan por la digestión ulterior realizada por las enzimas de las micro vellosidades, y los aminoácidos libres así producidos son llevados a la corriente sanguínea.")
148
Enfermedades asociadas a deficiencias de proteínas (a.a)
HIDROXILASA DE FENILALANINA: FENILCETUNORIA. (Degradación de fenilalanina y tirosina) OXIDASA DE HOMOGENTISATO: ALCAPTONURIA. TIROSINASA: ALBINISMO
 OXIDASA DE HOMOGENTISATO: ALCAPTONURIA. TIROSINASA: ALBINISMO.")
149
Fenilcetonuria (PKU) Cambio de fenilalanina a tirosina.
Enzima: hidroxilasa de fenilalanina. Es un defecto genético. Acumulación de fenilalanina o sus metabolitos producen retardación mental severa. La condición debe ser identificada inmediatamente luego del nacimiento. (Todos los bebes en Estados Unidos se les hace la prueba de PKU cuando nacen). Tratamiento consiste de una dieta baja en Phe y el monitoreo hasta los 10 años.
. Tratamiento consiste de una dieta baja en Phe y el monitoreo hasta los 10 años.")
150
Alcaptonuria Cambio de homogentisato a maleilacetoacetato.
Enzima: dioxigenasa de homogentisato. Acumulación de homogentisato, grandes cantidades son excretadas. El producto es oxidado y torna la orina negra. Las personas son susceptibles a desarrollar alguna forma de artritis. (primera enfermedad genética identificada asociada a la deficiencia de una enzima).
.")
151
Albinismo Fenilalanina se convierte en tirosina.
La Tirosina es precursor de la síntesis de melanina, el pigmento oscuro de la piel y el pelo presente en los melanocitos. Una deficiencia en la monooxigenasa de tirosina (tyrosinase) produce inhibición de la síntesis de melanina.
 produce inhibición de la síntesis de melanina.")
152
Recambio proteico Casi todas las proteínas del organismo están en una constante dinámica de síntesis (1-2% del total de proteínas), a partir de aminoácidos, y de degradación a nuevos aminoácidos. Esta actividad ocasiona una pérdida diaria neta de nitrógeno, en forma de urea, que corresponde a unos gramos de proteína. Cuando la ingesta dietética compensa a las pérdidas se dice que el organismo está en equilibrio nitrogenado. El balance nitrogenado puede ser positivo o negativo. Es positivo cuando la ingesta nitrogenada supera a las pérdidas, como sucede en crecimiento, embarazo, convalecencia de enfermedades. Es negativo si la ingesta de nitrógeno es inferior a las pérdidas, tal como ocurre en: desnutrición, anorexia prolongada, postraumatismos, quemaduras, deficiencia de algún aminoácido esencial.
, a partir de aminoácidos, y de degradación a nuevos aminoácidos. Esta actividad ocasiona una pérdida diaria neta de nitrógeno, en forma de urea, que corresponde a unos gramos de proteína. Cuando la ingesta dietética compensa a las pérdidas se dice que el organismo está en equilibrio nitrogenado. El balance nitrogenado puede ser positivo o negativo. Es positivo cuando la ingesta nitrogenada supera a las pérdidas, como sucede en crecimiento, embarazo, convalecencia de enfermedades. Es negativo si la ingesta de nitrógeno es inferior a las pérdidas, tal como ocurre en: desnutrición, anorexia prolongada, postraumatismos, quemaduras, deficiencia de algún aminoácido esencial.")
154
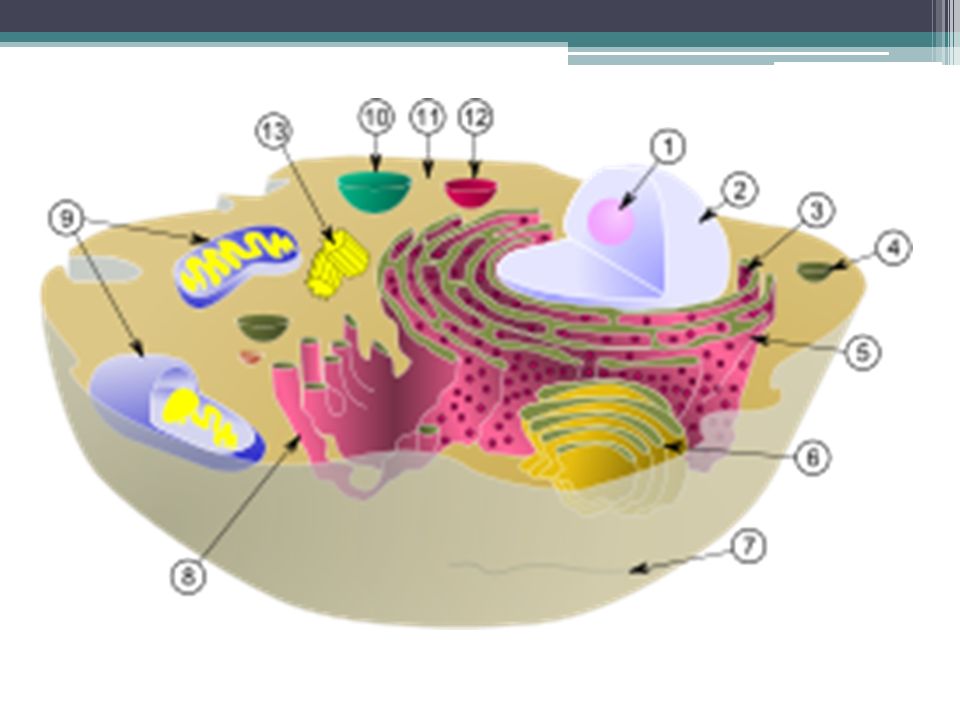
Estructura de la célula
Nucléolo Núcleo celular Ribosoma Vesículas de secreción Retículo endoplasmático rugoso Aparato de Golgi Citoesqueleto Retículo endoplasmático liso Mitocondria Vacuola Citosol Lisosoma Centríolo

155
GRACIAS POR SU ATENCION

156
Vías de degradación de las proteínas
Dos son las vías por la que son degradadas las proteínas mediante proteasas (catepsinas). 1. Vía de la ubiquitina (pequeña proteína básica). Fracciona proteínas anormales y citosólicas de vida corta. Es ATP dependiente y se localiza en el citosol celular. 2. Vía lisosómica. Fracciona proteínas de vida larga, de membrana, extracelulares y organelas tales como mitrocondrias. Es ATP independiente y se localiza en los
. 1. Vía de la ubiquitina (pequeña proteína básica). Fracciona proteínas anormales y citosólicas de vida corta. Es ATP dependiente y se localiza en el citosol celular. 2. Vía lisosómica. Fracciona proteínas de vida larga, de membrana, extracelulares y organelas tales como mitrocondrias. Es ATP independiente y se localiza en los.")
157
PRECURSORES DE AMINO ACIDOS
ALFA-CETOGLUTARATO OXALOACETATO PIRUVATO 3-FOSFOGLICERATO FOSFOENOLPIRUVATO + ERITROSA-4-FOSFATADA RIBOSA-5-FOSFATADA

Presentaciones similares