Descargar la presentación
La descarga está en progreso. Por favor, espere

1
ADAPTACIÓN METABÓLICA AL AYUNO RESPUESTA METABÓLICA AL ESTRÉS
Paula Andújar Plata Servicio de Endocrinología y Nutrición

2
Involuntario/voluntario/terapéutico/diagnóstico/patológico.
Punto de vista fisiológico: Situación en la que la ingesta es insuficiente para cubrir las necesidades de macronutrientes– adaptación metabólica que permite obtener energía y cubrir los requerimientos tisulares. Breve / prolongado. Total / parcial. Involuntario/voluntario/terapéutico/diagnóstico/patológico. -Breve: días/Prolongado (semanas o meses). -Total (nula salvo ingesta de agua)/ Parcial (cuando se cubren parcialmente los requerimientos) -Involuntario: desastres, voluntario: huelga de hambre; terapéutico: obesidad morbida; diagnostico: hipoglucemia; patologico: AN
. -Total (nula salvo ingesta de agua)/ Parcial (cuando se cubren parcialmente los requerimientos) -Involuntario: desastres, voluntario: huelga de hambre; terapéutico: obesidad morbida; diagnostico: hipoglucemia; patologico: AN.")
3
La repercusión metabólica del ayuno prolongado ha sido estudiada sobre todo en 4 situaciones:
Ensayos clínicos controlados (en pacientes con sobrepeso). Personas que han sufrido inanición (campos de concentración,…). Ayuno prolongado voluntario (huelgas de hambre). Ayuno parcial prolongado (AN).
. Personas que han sufrido inanición (campos de concentración,…). Ayuno prolongado voluntario (huelgas de hambre). Ayuno parcial prolongado (AN).")
4
METABOLISMO DE LOS PRINCIPIOS INMEDIATOS EN CONDICIONES NORMALES
Tras la ingesta → HC, lípidos y proteínas → energía (ATP). Cerebro y células sanguíneas: sólo se nutren de glucosa, salvo en situaciones especiales como el ayuno prolongado. Tras ingesta → azúcares simples (glucosa, fructosa y galactosa) → se absorben. La glucosa es empleada como fuente de energía por los distintos tejidos (a través del ciclo de Krebs se obtiene ATP), o se almacena en hígado y músculo (GLUCÓGENO). HIDRATOS DE CARBONO - HC, lípidos y proteínas: cada uno es utilizado por sus vías para dar lugar a energía. Cada uno a través de sus vías dará lugar a ATP, que se almacenará para ser utilizado como fuente de energía. - HC:Una vez ingeridos se transforman en azúcares simples (glucosa, fructosa y galactosa)- se absorben y se transportan por la porta y van al hígado donde se acumulan.
. Cerebro y células sanguíneas: sólo se nutren de glucosa, salvo en situaciones especiales como el ayuno prolongado. Tras ingesta → azúcares simples (glucosa, fructosa y galactosa) → se absorben. La glucosa es empleada como fuente de energía por los distintos tejidos (a través del ciclo de Krebs se obtiene ATP), o se almacena en hígado y músculo (GLUCÓGENO). HIDRATOS DE CARBONO. - HC, lípidos y proteínas: cada uno es utilizado por sus vías para dar lugar a energía. Cada uno a través de sus vías dará lugar a ATP, que se almacenará para ser utilizado como fuente de energía. - HC:Una vez ingeridos se transforman en azúcares simples (glucosa, fructosa y galactosa)- se absorben y se transportan por la porta y van al hígado donde se acumulan.")
5
Son el menor depósito de energía. Glucosa circulante (20 g, 80 kcal).
Glucógeno hepático ( g) y muscular ( g). Cada gramo → 4kcal. El depósito hepático de glucosa supone unas kcal. De todo el glucógeno almacenado, en condiciones normales sólo está disponible el hepático. El glucógeno muscular no puede liberar glucosa al plasma (uso local o situaciones de emergencia). Las medidas de glucógeno: en un individuo de peso medio. kcal: una tercera parte de los requerimientos calóricos diarios
 y muscular ( g). Cada gramo → 4kcal. El depósito hepático de glucosa supone unas kcal. De todo el glucógeno almacenado, en condiciones normales sólo está disponible el hepático. El glucógeno muscular no puede liberar glucosa al plasma (uso local o situaciones de emergencia). Las medidas de glucógeno: en un individuo de peso medio kcal: una tercera parte de los requerimientos calóricos diarios.")
6
LÍPIDOS En el intestino se transforman en quilomicrones → sistema linfático → plasma donde son captados por el hígado o el tejido adiposo. La enzima lipoproteín lipasa (presente en los 2 órganos) los convierte en glicerol y ácidos grasos libres, que son transportados a los distintos tejidos. Los AGL pueden seguir 2 caminos: Producción de energía mediante la betaoxidación mitocondrial Almacenamiento en tejido adiposo en forma de triglicéridos, preparados para ser usados como fuente de energía cuando sea necesario. Producción de energía Almacenamiento en tejido adiposo (TG)
 los convierte en glicerol y ácidos grasos libres, que son transportados a los distintos tejidos. Los AGL pueden seguir 2 caminos: Producción de energía mediante la betaoxidación mitocondrial. Almacenamiento en tejido adiposo en forma de triglicéridos, preparados para ser usados como fuente de energía cuando sea necesario. Producción de energía. Almacenamiento. en tejido adiposo (TG)")
7
El valor energético de la grasa es elevado: 9 kcal por gramo.
GRASA constituye ± el 15-25% del peso corporal → unos 10.5 kg en una persona de 70 kg →94500 kcal almacenadas. El tejido adiposo es la principal reserva de energía del organismo. - Cuerpos cetónicos: desempeñan un papel fundamental en el ayuno prolongado.

8
PROTEÍNAS Función energética pero también estructural!!
La ingesta mínima necesaria: 1 g/kg/día. Proteínas → aa → sangre → células. No existe un tejido que almacene específicamente proteínas así que toda proteína no utilizada es desaminada por el hígado. Las porciones hidrocarbonadas restantes son empleadas para la gluconeogénesis, la lipogénesis o con mayor frecuencia, para entrar en el ciclo de Krebs y ser utilizados como energía. 4 kcal/g. Se estima que las proteínas constituyen el 20% del peso, repartidas entre el tejido muscular (50%) y los tejidos viscerales. - Las proteínas son hidrolizadas a aa y pasan a la sangre y de ahí a las células para formar parte de las proteínas celulares.
 y los tejidos viscerales. - Las proteínas son hidrolizadas a aa y pasan a la sangre y de ahí a las células para formar parte de las proteínas celulares.")
9
FISIOLOGÍA DEL AYUNO PRECOZ
Desde el final de la absorción de una comida hasta la otra. Glucosa Glucagón Insulina Catecolaminas - Desde el final de una comida hasta la otra: unas horas. - Los niveles de glucosa y por tanto de insulina van disminuyendo y aumentan los de glucagón y catecolaminas– esto lleva al inicio de la glucógenolisis hepática, que en condiciones normales es frenado por una nueva comida. Esta situación es la que se produce a primera hora de la mañana. GLUCOGENÓLISIS HEPÁTICA

10
SE AGOTA EL PRIMER MECANISMO DE RESERVA DE ENERGÍA.
Cuando el período de ayuno es > horas, se estima que las reservas de glucógeno hepático están agotadas. SE AGOTA EL PRIMER MECANISMO DE RESERVA DE ENERGÍA. Algunos tejidos necesitan glucosa Esta vía se pone en marcha a las horas de ayuno como complementos de la glucogenólisis y la suplanta totalmente una vez pasado el primer día → mantiene el aporte de hidratos requerido por el cerebro (150 g de glucosa al día). GLUCONEOGÉNESIS - Algunos tejidos: (cerebro, hematíes y médula renal)necesitan glucosa– se inicia gluconeogénesis.
. GLUCONEOGÉNESIS. - Algunos tejidos: (cerebro, hematíes y médula renal)necesitan glucosa– se inicia gluconeogénesis.")
11
Cetogénesis Fase post-absortiva Gluconeogénesis Glucogenólisis 6 horas 16 horas 30 horas

12
Los SUSTRATOS PARA LA PRODUCCIÓN HEPÁTICA DE GLUCOSA SON:
GLICEROL. Generado de la hidrólisis de los TG junto con los AGL. El rendimiento del metabolismo del glicerol en glucosa es escaso. AA. Liberados por las proteínas tisulares. Son el sustrato más importante para la síntesis de glucosa. Contribuyen a la producción de ± 75 g al día. Con este fin se envían al hígado cerca de g de aa (alanina). La alanina y la glutamina constituyen el 50-60% de los aa que se liberan del músculo durante el ayuno. 3. LACTATO. Liberado del músculo a través de la glucólisis anaerobia. g de aa, fundamentalmente alanina. Gran parte de la alanina empleada en la gluconeogénesis se obtiene a partir del piruvato derivado del glucógeno muscular y otra parte se obtiene a partir de la glutamina originada como consecuencia del catabolismo de los aa ramificados. La glutamina generada a nivel muscular puede tener varios destinos: a) síntesis de nucleótidos por los tejidos periféricos; b) combustible para enterocitos, c) produccíon de amoniaco a nivel renal para neutralizar los cuerpos cetónicos.
. La alanina y la glutamina constituyen el 50-60% de los aa que se liberan del músculo durante el ayuno. 3. LACTATO. Liberado del músculo a través de la glucólisis anaerobia g de aa, fundamentalmente alanina. Gran parte de la alanina empleada en la gluconeogénesis se obtiene a partir del piruvato derivado del glucógeno muscular y otra parte se obtiene a partir de la glutamina originada como consecuencia del catabolismo de los aa ramificados. La glutamina generada a nivel muscular puede tener varios destinos: a) síntesis de nucleótidos por los tejidos periféricos; b) combustible para enterocitos, c) produccíon de amoniaco a nivel renal para neutralizar los cuerpos cetónicos.")
13
A pesar de la gluconeogénesis →
Prácticamente toda la glucosa por la gluconeogénesis está destinada a cerebro y hematíes. El resto: se nutren de AGL. A pesar de la gluconeogénesis → REDUCCIÓN DE RESERVAS PROTEICAS. Por eso durante el ayuno precoz se ponen en marcha otros procesos adaptativos. Se inhibe la oxidación periférica de glucosa Se utilizan como fuente primaria de energía las grasas. Se produce la hidrólisis de TG en el tejido adiposo y se genera glicerol y AGL.– las grasas se convierten en la principal fuente de energía del individuo en ayunas. Parte de esos AGL se transforman en cuerpos cetónicos, produciendo unas 4.5 kcal/g. ENERGÍA INSUFICIENTE

14
Proteólisis Glucogenólisis AA GLUCOSA Gluconeogénesis GLICEROL Glucólisis anaerobia PIRUVATO LACTATO Lipólisis AGL

15
Tras 2-3 días de ayuno, la producción de cuerpos cetónicos alcanza su máximo nivel (130 g/día), pero los niveles en sangre no aumentan ya que son utilizados por los tejidos periféricos como fuente de energía. Las necesidades energéticas quedan cubiertas a nivel periférico por los CC y a nivel cerebral por el aporte de glucosa vía gluconeogénesis. Ambiente hormonal de hipoinsulinemia, con escaso aumento de las hormonas contrarreguladoras. De esta manera, las necesidades energéticas,… Todo esto se realiza en un ambiente hormonal de…

16
FISIOLOGÍA DEL AYUNO PROLONGADO
Durante los primeros 5 días de ayuno la pérdida de proteínas es de 75 g/día → supone el catabolismo diario de 300 g de músculo. Para evitar continuar con la pérdida proteica → mecanismos de adaptación. Cuando el ayuno supera las 3 semanas → descenso en la excreción de nitrógeno urinario, reflejo de la disminución de la gluconeogénesis y por lo tanto del catabolismo proteico. Esto implica un descenso de los niveles de glucosa plasmática, que en esta fase de ayuno prolongado es sintetizada también en el riñón. - Mecanismos de adaptación que permiten disminuir el consumo de proteínas

17
La reducción del metabolismo de los hidratos de carbono produce un déficit de oxalacetato, necesario para introducir los cuerpos cetónicos en el ciclo de Krebs. Por lo tanto, la utilización periférica de los CC disminuye y su nivel plasmático aumenta hasta el punto de que atraviesa la barrera hematoencefálica. En la fase de ayuno prolongado, el 50-75% de la energía requerida por el cerebro se obtiene de los cuerpos CC. - atraviesa la barrera hematoencefálica y son empleados por el cerebro.

18
Por otra parte, en el músculo esquelético, los CC son sustituidos por AGL como fuente de energía principal. El ambiente biológico de ayuno → cociente glucagón/insulina elevado, lo que facilita la movilización de AGL hacia el hígado (estímulo para formar CC). Tanto en la fase de ayuno precoz como en la de prolongado, el 30% de los AGL liberados del tejido adiposo son convertidos en el hígado en CC. En el ayuno prolongado se ponen en marcha mecanismos que disminuyen los requerimientos energéticos y reducen la proteólisis al mínimo para producir el amonio suficiente para eliminar los CC por la orina. Esta situación permite al individuo mantenerse con vida mientras tenga suficiente reserva de grasa (± 1 mes).
. Tanto en la fase de ayuno precoz como en la de prolongado, el 30% de los AGL liberados del tejido adiposo son convertidos en el hígado en CC. En el ayuno prolongado se ponen en marcha mecanismos que disminuyen los requerimientos energéticos y reducen la proteólisis al mínimo para producir el amonio suficiente para eliminar los CC por la orina. Esta situación permite al individuo mantenerse con vida mientras tenga suficiente reserva de grasa (± 1 mes).")
19
Cetogénesis CUERPOS CETÓNICOS

20
EVOLUCIÓN DE LOS NIVELES PLASMÁTICOS DE LOS PRINCIPALES METABOLITOS
GLUCEMIA. Descenso brusco al tercer día, manteniéndose posteriormente en valores de mg/dL. AGL. Se elevan de forma importante los tres primeros días, para mantenerse altos hasta que finaliza el ayuno. CC. No aumentan los primeros días porque aunque aumentan, aumenta también su consumo. El aumento se produce a partir del 4-5 dias, en relación con la disminución de su utilización por los tejidos, excepto el cerebro. Alcanzan el máximo a las 3 semanas de ayuno. - AGL: coincidiendo con los de glucosa…

21
AA glucogénicos (alanina principalmente)
AA glucogénicos (alanina principalmente). Parten de niveles altos y van disminuyendo por la gluconeogénesis. AA ramificados (valina, leucina, isoleucina) aumentan los 3-5 primeros días por aumento de proteólisis muscular y luego descenso progresivo.
. Parten de niveles altos y van disminuyendo por la gluconeogénesis. AA ramificados (valina, leucina, isoleucina) aumentan los 3-5 primeros días por aumento de proteólisis muscular y luego descenso progresivo.")
22
RESPUESTA HORMONAL AL AYUNO BREVE
INSULINA: papel importante en la regulación de la homeostasis de los tres principios inmediatos. Aumento glucosa en plasma en periodo postabsortivo Elevación niveles de insulina Aumento de uso periférico de glucosa y disminución de producción endógena / Síntesis proteínas y almacenamiento de TG en el tejido adiposo En condiciones normales: aumento de glucos… Producción endógena: glucogenólisis y gluconeogénesis El aumento de insulina estimula también la síntesis de proteínas y el almacenamiento de TG en el tejido adiposo.

23
En AYUNO BREVE: Disminución concentración plasmática de glucosa Descenso de los niveles de insulina Reducción de la utilización periférica de glucosa y un aumento de sus síntesis mediante la glucogenólisis y gluconeogénesis. El descenso de insulina favorece también la proteólisis y la liberación de AGL procedentes de la lipólisis

24
Las hormonas contrarreguladoras (cortisol, GH, adrenalina, glucagón) tienen un efecto opuesto a la insulina. Estas hormonas aumentan la proteólisis lo que favorece la liberación de aa (sustrato para la gluconeogenesis). Cuando los HDC son insuficientes → liberación masiva de AGL y glicerol a la circulación, para ser convertidos en CC.
. Cuando los HDC son insuficientes → liberación masiva de AGL y glicerol a la circulación, para ser convertidos en CC.")
25
RESPUESTA HORMONAL AL AYUNO PROLONGADO
INSULINA. Los niveles plasmáticos disminuyen al principio del ayuno. T3, GH. También disminuyen al inicio del ayuno. GLUCAGÓN. Se eleva inicialmente, para bajar después y mantenerse en valores superiores a los de la fase postabsortiva

26
RESPUESTA NEUROENDOCRINA AL AYUNO
LEPTINA Polipéptido sintetizado en los adipocitos. Concentración proporcional a masa grasa. Ejerce su función a nivel del SNC. Cuando el organismo tiene los depósitos de energía llenos → aumentan los niveles de leptina en sangre y en el SNC se desencadena una respuesta por la que disminuye el apetito y aumenta el gasto metabólico. En ayuno, se produce un descenso de la leptina, favoreciendo la conservación de energía. A nivel hipotalámico inhibe funciones no esenciales como la capacidad reproductora.

27
La acción de la leptina está modulada por otros neuropéptidos y hormonas.
La insulina junto con la leptina constituyen una señal de feed back negativo para el cerebro → ante un balance energético negativo como en una situación de ayuno, el descenso de ambas desencadena una respuesta a nivel hipotalámico que se traduce en aumento de apetito.

28
Pertenece a la familia de los polipéptidos pancreáticos.
NEUROPÉPTIDO Y Pertenece a la familia de los polipéptidos pancreáticos. Se expresa en múltiples regiones cerebrales. Se sintetiza en el hipotálamo, en las neuronas del núcleo arqueado, desde donde envía señales a otra áreas hipotalámicas, incluido el núcleo paraventricular. Estimula la ingesta, disminuye el gasto energético mediante la reducción del flujo del sistema nervioso simpático y favorece la síntesis y almacenamiento de grasa. -A través del núcleo paraventricular, estimula la ingesta… En animales sometidos a ayuno, se ha observado un aumento de la expresión del gen del neuropéptido Y y que la insulina y la leptina inhiben su expresión, mientras que los GC la estimulan

29
El ayuno induce la activación de este eje a través de varias vías:
EJE HHSUPRARRENAL El ayuno induce la activación de este eje a través de varias vías: La síntesis de la hormona liberadora de corticotropina (CRH), principal regulador de eje, que se mantiene igual o disminuye tras reducción ponderal por ayuno. El aumento de la liberación de otros secretagogos de ACTH, como urocortina y hormona antidiuretica. El neuropéptido Y activa el eje suprarrenal a nivel hipotalámico. - El neuropéptido Y activa el eje suprarrenal a nivel hipotalámico.; mientras que su incremento en el ayuno depende , en parte, de la estimulación por los GC; por lo tanto, la activación suprarrenal por el neuropéptido Y durante el ayuno puede estimular más la síntesis de este neuropéptido
, principal regulador de eje, que se mantiene igual o disminuye tras reducción ponderal por ayuno. El aumento de la liberación de otros secretagogos de ACTH, como urocortina y hormona antidiuretica. El neuropéptido Y activa el eje suprarrenal a nivel hipotalámico. - El neuropéptido Y activa el eje suprarrenal a nivel hipotalámico.; mientras que su incremento en el ayuno depende , en parte, de la estimulación por los GC; por lo tanto, la activación suprarrenal por el neuropéptido Y durante el ayuno puede estimular más la síntesis de este neuropéptido.")
30
UROCORTINA, ARGININA, VASOPRESINA
AYUNO NEUROPÉPTIDO Y ↑ CRH N o ↓ - El papel de CRH en la estimulación del eje corticoideo durante la reducción ponderal puede ser suplantado en el tiempo por otros factores hipotalámicos. La capacidad de otros secretagogos de ACTH para activar el eje anula el feed back negativo hipotalámico de los esteroides, dando lugar a la supresión indirecta de CRh. UROCORTINA, ARGININA, VASOPRESINA ACTH ↑ LEPTINA ↓ INSULINA ↓ CORTISOL ↑

31
RESPUESTA METABÓLICA AL ESTRÉS
Proceso patológico con gravedad suficiente para desencadenar una respuesta funcional, endocrina y metabólica → con el objetivo de proporcionar los sustratos energéticos necesarios para el mantenimiento de las funciones vitales y la recuperación de los órganos lesionados. La respuesta fisiológica al estrés agudo incluye cambios en el metabolismo de los principios inmediatos dirigidos a aumentar la disponibilidad de glucosa, aa y AGL. Este proceso permite la síntesis de proteínas de fase aguda y la activación del sistema inmune, lo que conlleva la infiltración leucocitaria de los tejidos lesionados y la liberación sistémica de citoquinas. Esta respuesta se produce de forma independiente de la ingesta de nutrientes. - Por ejemplo el paciente crítico: politraumatismo, sepsis, cirugía mayor y quemados extensos. Glucosa, aa y AGL: que serán utilizados principalmente por órganos vitales. CK: (interfereon gamma, TNF,..)
")
32
HIPODINÁMICA CATABÓLICA
Desde el punto de vista metabólico se distinguen 2 fases en la evolución de la respuesta fisiológica al estrés: HIPODINÁMICA CATABÓLICA La respuesta a la situación de estrés se produce por la liberación de citoquinas y tras una serie de cambios neuroendocrinos, que incluyen la activación del eje hipotálamo-hipofisario suprarrenal, la hipersecreción de PRL y GH en presencia de bajos niveles circulantes de IGF-1, así como un descenso de la actividad de los ejes tiroideo y gonadal.

33
FASE HIPODINÁMICA Comienza tras la agresión y dura 24-72 horas.
Es una situación de HIPERCATABOLISMO y por tanto de gran disponibilidad de sustratos, pero éstos no pueden ser empleados adecuadamente por la disminución de la perfusión y de la oxigenación de los tejidos secundaria a la reducción del volumen sanguíneo y del gasto cardíaco.

34
HIDRATOS DE CARBONO La CK proinflamatorias liberadas a nivel sistémico en respuesta a la agresión potencian la actividad de otras CK así como de hormonas catabólicas (cortisol, glucagón y catecolaminas) Estimulan la glucogenólisis y la movilización de glucosa. Se reducen rápidamente los depósitos de glucógeno, momento en que empieza a ponerse en marcha la gluconeogénesis y los lípidos se convierten en la principal fuente de energía, ya al final de esta fase. La glucosa obtenida es metabolizada principalmente mediante la glucólisis anaerobia, generando ácido láctico. Los niveles de insulina suelen estar bajos en esta fase.
 Estimulan la glucogenólisis y la movilización de glucosa. Se reducen rápidamente los depósitos de glucógeno, momento en que empieza a ponerse en marcha la gluconeogénesis y los lípidos se convierten en la principal fuente de energía, ya al final de esta fase. La glucosa obtenida es metabolizada principalmente mediante la glucólisis anaerobia, generando ácido láctico. Los niveles de insulina suelen estar bajos en esta fase.")
35
PROTEÍNAS Aumento del catabolismo proteico.
El flujo principal de aa procede de la porción móvil de la musculatura esquelética, el tejido conectivo y el intestino en reposo. Dichos aa son captados por el hígado, donde se utilizarán en la síntesis de proteínas, principalmente reactantes de fase aguda (disminuyendo así otras como la albúmina). Esta proteólisis acelerada se mantiene, provocando una importante pérdida proteica a nivel muscular y visceral, que acaba afectando a enzimas y otros péptidos con funciones específicas. En paciente que reciben un soporte nutricional adecuado con aporte de aa la probabilidad de supervivencia es algo mayor, si bien no suprime por completo la respuesta catabólica. En un paciente estresado, no alimentado, se pierden hasta 250 g de masa muscular diaria. En esta fase se produce un aumento del catabolismo proteico. Esta proteólisis acelerada no se atenúa en el tiempo (como en el ayuno)
. Esta proteólisis acelerada se mantiene, provocando una importante pérdida proteica a nivel muscular y visceral, que acaba afectando a enzimas y otros péptidos con funciones específicas. En paciente que reciben un soporte nutricional adecuado con aporte de aa la probabilidad de supervivencia es algo mayor, si bien no suprime por completo la respuesta catabólica. En un paciente estresado, no alimentado, se pierden hasta 250 g de masa muscular diaria. En esta fase se produce un aumento del catabolismo proteico. Esta proteólisis acelerada no se atenúa en el tiempo (como en el ayuno)")
36
LÍPIDOS Las hormonas catabólicas son las principales estimulantes de la hidrólisis de TG que genera glicerol y AGL. El glicerol se emplea como sustrato en la gluconeogénesis. Los AGL, al no poder ser oxidados directamente por los tejidos periféricos (por la hipoperfusión y la hipoxia tisular) participan en la cetogénesis hepática. Los CC obtenidos sirven de sustrato a los tejidos periféricos y contribuyen a atenuar la proteólisis.
 participan en la cetogénesis hepática. Los CC obtenidos sirven de sustrato a los tejidos periféricos y contribuyen a atenuar la proteólisis.")
37
FASE CATABÓLICA Transición progresiva hacia la fase catabólica, a medida que aumentan la gluconeogénesis y la captación tisular de O2. Se produce gracias a la recuperación del volumen sanguíneo y del gasto cardiaco. A su vez, esta fase tiene una parte aguda que se caracteriza por un aumento del catabolismo de todos los principios inmediatos, unido a una utilización aumentada de los sustratos disponibles, que permite la reparación tisular y la supervivencia sin aporte exógeno de nutrientes; en la segunda parte que sería adaptativa, el organismo va recuperando progresivamente la normalidad, preparando el anabolismo. La mejor oxigenación de los tejidos → mayor actividad metabólica → mayor gasto metabólico basal; además favorece la termogénesis → contribuye a aumentar más el gasto energético basal. Aquí se hace referencia principalmente a la primera fase. Estos efectos son contrarios a los que se produce en ayuno (se reduce el gasto metabólico basal y la Tª corporal).
.")
38
HIDRATOS DE CARBONO Aumento de la producción de glucosa → hiperglucemia mantenida, a pesar de una mayor captación de glucosa por los tejidos. Los niveles de insulina plasmáticos están elevados en respuesta a la hiperglucemia y a la resistencia a la insulina, favorecida por las hormonas contrarreguladoras y las CK proinflamatorias.

39
Persiste un aumento de la síntesis de reactantes de fase aguda y del catabolismo proteico.
Los aa obtenidos son fundamentalmente alanina y glutamina, y son destinados en su mayoría a la gluconeogénesis (sobre todo la alanina). La glutamina es además un sustrato metabólico para la mucosa intestinal y las células del sistema inmune. Se acaba produciendo un catabolismo neto de la masa magra, favorecido por otros factores como acidosis metabólica, la desnutrición previa o el reposo prolongado, que desde el punto de vista clínico se traduce en una pérdida de masa muscular y en un balance nitrogenado negativo. Por el contrario, en el ayuno prolongado el catabolismo proteico está atenuado. PROTEÍNAS
. La glutamina es además un sustrato metabólico para la mucosa intestinal y las células del sistema inmune. Se acaba produciendo un catabolismo neto de la masa magra, favorecido por otros factores como acidosis metabólica, la desnutrición previa o el reposo prolongado, que desde el punto de vista clínico se traduce en una pérdida de masa muscular y en un balance nitrogenado negativo. Por el contrario, en el ayuno prolongado el catabolismo proteico está atenuado. PROTEÍNAS.")
40
LÍPIDOS El catabolismo de los TG está acelerado, generando AGL que constituyen el principal sustrato energético. Parte de esos AGL son empleados en la cetogénesis, que no se atenúa a pesar de la administración de glucosa.

41
FASE HIPODINÁMICA FASE CATABÓLICA Duración 48-72 horas Varios días-semanas Perfusión tisular ↓ ↑ Consumo de O2 Actividad metabólica ↑↑ Catabolismo Producción de sustratos Utilización de sustratos Hidratos de carbono ↑ glucogenólisis ↑ gluconeogénesis Lípidos ↑ cetogénesis ↑ oxidación de ácidos grasos Proteínas ↑ proteólisis

42
ESTRÉS EN GRANDES QUEMADOS
Es un modelo típico de situación de estrés, aparte de tener algunas peculiaridades propias. A través de la quemadura se producen importantes pérdidas de calor, agua, nitrógeno, proteínas y micronutrientes. Además la solución de continuidad a nivel cutáneo deteriora la barrera defensiva lo que favorece infección que pueden complicar la situación del paciente. El soporte nutricional de estos pacientes es muy importante. Hay múltiples factores que limitan la ingesta y además se debe evitar la sobrealimentación. Factores que limitan la ingesta: (intervenciones, íleo paralñitico,..) (si la infusión de glucosa es excesiva aumenta la producción de dióxido de carbono y esto dificulta el manejo ventilatorio).
 (si la infusión de glucosa es excesiva aumenta la producción de dióxido de carbono y esto dificulta el manejo ventilatorio).")
43
AGL. En los quemados la lipólisis aumenta hasta 3-4 veces con respecto a la situación basal, para alcanzar una disponibilidad máxima de AGL, cuya oxidación están francamente aumentada, llegando al 60-70% de los sustratos oxidados. GLUCOSA. A pesar de la menor contribución de la glucosa y los aa a la elevación del gasto energético, en los quemados su tasa de oxidación está también aumentada en un 33%. En respuesta a una mayor captación tisular y oxidación de la glucosa, se acelera la gluconeogénesis. AA. El recambio proteico está elevado. También aumentan su tasa de oxidación y la producción de urea. La síntesis proteica está prácticamente duplicada. Situación de hipercatabolismo grave que implica un importante aumento de los requerimientos energéticos.

44
RESPUESTA NEUROENDOCRINA AL ESTRÉS
La respuesta neuroendocrina aguda al estrés se desencadena a nivel hipotalámico, donde estímulos como la hipovolemia, la hipoxemia o el dolor → activación del eje HHS, la hipersecreción de PRL y GH en presencia de bajos niveles circulantes de IGF1, así como el descenso de la actividad de los ejes tiroideo y gonadal. Desde el hipotálamo también se estimula el sistema nervioso simpático y se aumenta la liberación de glucagón. La fase aguda se caracteriza por una secreción activa aumentada de las hormonas de la hipófisis anterior, con una inactivación o descenso de las hormonas anabólicas (TSH, LH, FSH). En caso de que el cuadro no se resuelva, se instaura la fase crónica, en la que disminuye la liberación de hormonas hipofisarias anteriores por falta de estímulo hipotalámico.
. En caso de que el cuadro no se resuelva, se instaura la fase crónica, en la que disminuye la liberación de hormonas hipofisarias anteriores por falta de estímulo hipotalámico.")
45
INSULINA Inmediatamente después de la agresión, los niveles de insulina suelen ser bajos, lo que se atribuye a la descarga de catecolaminas. Posteriormente, la insulina se eleva en respuesta a la hiperglucemia y debido a la resistencia insulínica inducida por las hormonas contrarreguladoras y algunas CK que estimulan su secreción. La hiperinsulinemia favorece la captación de glucosa en los tejidos, mientras que en el músculo esquelético debido a la resistencia a la insulina, disminuye el consumo de glucosa y se permite la degradación de proteínas (para la gluconeogénesis)
")
46
EJE HHS Situación de hipercortisolismo asociado a un aumento de la liberación de ACTH mediado por CRH, a la presencia de CK y a la activación del sistema noradrenérgico. Se activa también el sistema renina-angiotensina-aldosterona. En esta fase, los niveles de la proteína transportadora del cortisol, así como de la albúmina, están disminuidos, lo que determina un aumento de la fracción libre de cortisol. Movilización rápida de grasas, hidratos de carbono y proteínas. Ejerce un papel especialmente importante en el músculo esquelético, donde propicia la proteólisis y la liberación de aa, e inhibe el flujo de entrada de éstos al músculo, favoreciendo la gluconeogénesis.

47
Esta situación puede persistir varias semanas y genera una pérdida importante de nitrógeno, que altera la capacidad de reparación tisular, la cicatrización y la competencia inmunitaria. La acción catabólica de los GC se ve potenciada por la disminución o la resistencia a la acción de hormonas anabolizantes como la GH, la insulina o la testosterona. El hipercortisolismo también favorece la retención de líquido intravascular e incrementa la respuesta ionotrópica y vasopresora a las catecolaminas y la angiotensina II respectivamente. En la fase crónica, se mantiene los niveles altos de cortisol aunque la ACTH es baja, lo que indica que el estímulo suprarrenal no depende de ésta. La concentración de la proteínas transportadora de cortisol aumenta, por lo que la fracción libre disminuye, alcanzando niveles casi normales en la fase de recuperación. No está claro por qué la ACTH disminuye.

48
EJE SOMATOTROPO En ausencia de fármacos supresores (dopamina, GC), en las primeras horas tras la agresión, los niveles de GH aumentan en la amplitud y la frecuencia de los pulsos, así como la concentración interpulsos. Sin embargo, los niveles de IGF-1 e IGFBP3 están disminuidos en sangre, lo que sugiere una resistencia a GH. Los efectos indirectos de la GH mediados por IGF1 y el feed back negativo de IGF1 están atenuados, con la consecuente elevación de GH y el predominio de su acción lipolítica, antagonista de la insulina y estimuladora del sistema inmune. - La disminución de la expresión de receptores de GH es un fenómeno temprano y se asocia con niveles deIGF1 bajos, lo que se supone que estimula el aumento de secreción de GH. Probablemente en esta resistencia también influyan CK inflamatorias.
, en las primeras horas tras la agresión, los niveles de GH aumentan en la amplitud y la frecuencia de los pulsos, así como la concentración interpulsos. Sin embargo, los niveles de IGF-1 e IGFBP3 están disminuidos en sangre, lo que sugiere una resistencia a GH. Los efectos indirectos de la GH mediados por IGF1 y el feed back negativo de IGF1 están atenuados, con la consecuente elevación de GH y el predominio de su acción lipolítica, antagonista de la insulina y estimuladora del sistema inmune. - La disminución de la expresión de receptores de GH es un fenómeno temprano y se asocia con niveles deIGF1 bajos, lo que se supone que estimula el aumento de secreción de GH. Probablemente en esta resistencia también influyan CK inflamatorias.")
49
El patrón de secreción de GH en la fase crónica es distinto:
disminuye la fracción pulsátil de GH, con una fracción no pulsátil todavía aumentada y una frecuencia de pulso aumentada. Esto supone una disminución de los niveles de IGF-1 e IGFBP3. Esto parece ocurrir cuando el paciente ha estado en situación crítica durante un tiempo prolongado. Se instaura entonces, un déficit relativo de GH que podría contribuir a la situación catabólica del paciente crítico crónico.

50
EJE TIROTROPO Sólo 2 horas tras la agresión, disminuyen los niveles plasmáticos de T3 por una reducción de la conversión periférica de T4 a T3 por la desyodinasa I. La gravedad de la enfermedad queda reflejada en la magnitud del descenso de T3 en las primeras 24 h. Este descenso de T3 conlleva un aumento leve de T4 y TSH, así como elevación de la T3 reversa. En pacientes muy graves la T4 también puede descender. En la fase crónica, a pesar del marcado descenso de T3, la TSH puede permanecer normal o incluso baja, pues disminuye la amplitud y la frecuencia de la pulsatilidad nocturna. Esta ausencia de elevación de TSH podría ser por un aumento del umbral de respuesta del eje. Estos cambios reflejan un intento de disminuir el gasto energético (no precisa tratamiento). - El TNF alfa, IL-6 IL1 podrían ser los responsables de esta respuesta.
. - El TNF alfa, IL-6 IL1 podrían ser los responsables de esta respuesta.")
51
EJE LACTOTROPO La PRL aumenta en las primeras 24 h. Los mediadores de esta elevación no están claros. En la fase crónica, la PRL permanece elevada. Su papel en la respuesta a la agresión parece estar en relación con la función inmunitaria. EJE GONADOTROPO La testosterona es el principal anabolizante endógeno. En situación de estrés grave se produce un hipogonadismo transitorio y por lo tanto la pérdida de su acción anabolizante.

53
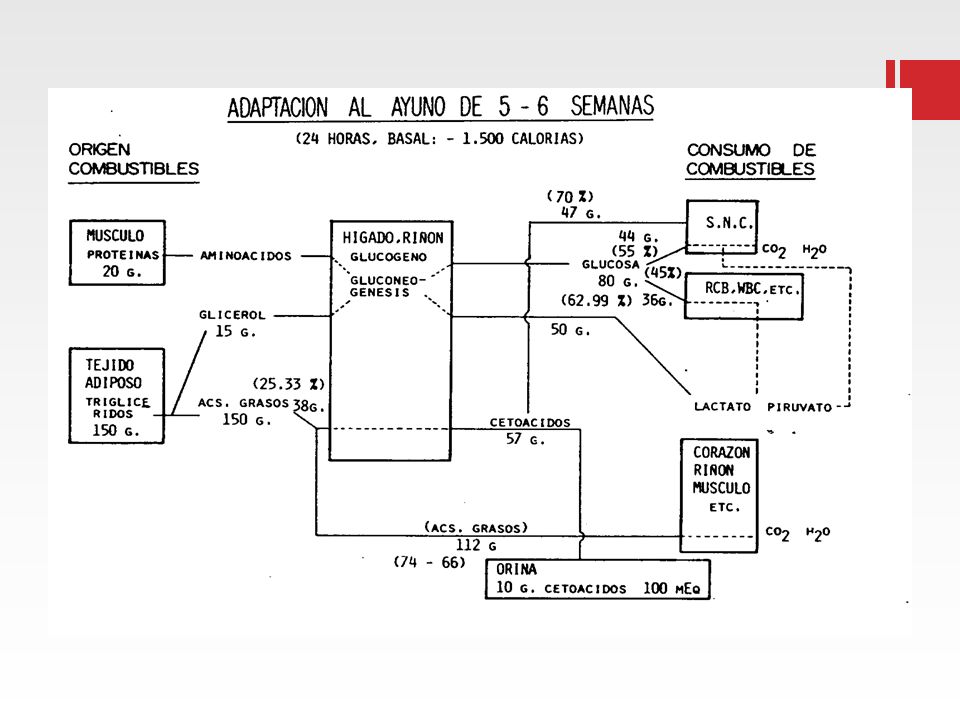
ADAPTACIÓN METABÓLICA EN AYUNO
(Adapt. Cahill)
")
54
Solo en ayuno prolongado
GRASA CORPORAL TEJ. ADIPOSO LIPOLISIS GLUCEMIA MUSCULO CATABOLISMO MASA MUSCULAR GLICEROL A. A. (ALANINA) NEOGLUCOGENESIS HEPATICA Ac. GRASOS CETOACIDOS OXIDACION (MÚSCULO, CORAZÓN, HÍGADO, RIÑÓN...) GLUCOSA OXIDACIÓN (C. SANGUÍNEAS- M. RENAL) (S. N. C.) Solo en ayuno prolongado
 NEOGLUCOGENESIS. HEPATICA. Ac. GRASOS. CETOACIDOS. OXIDACION. (MÚSCULO, CORAZÓN, HÍGADO, RIÑÓN...) GLUCOSA. OXIDACIÓN. (C. SANGUÍNEAS- M. RENAL) (S. N. C.) Solo en ayuno prolongado.")
56
COMPOSICIÓN CORPORAL UTILIZACIÓN DE MACRONUTRIENTES EN AYUNO
Total (Kg) Pérdida en ayuno (Kg) Duración (Días) 70 26.6 HC 0.3 0.3 (100%) < 1 Proteína 12.2 4.6 (37.7%) 60-65 Grasa 9 8 (89%) 48-61 (Tomado de Elia M 2000) Peso
 Pérdida en. ayuno (Kg) Duración. (Días) HC (100%) < 1. Proteína (37.7%) Grasa (89%) (Tomado de Elia M 2000) Peso.")
Presentaciones similares