Descargar la presentación
La descarga está en progreso. Por favor, espere

1
La genética mendeliana La teoría cromosómica de la herencia
Tema 63 La genética mendeliana La teoría cromosómica de la herencia Las mutaciones

2
1. La genética mendeliana
La metodología mendeliana Gregor Mendel puso de manifiesto, por primera vez y con su Investigación sobre la hibridación en plantas, que la herencia de los caracteres respondía a ciertas leyes. Su gran mérito fue el uso de una metodología científica con base estadísti-ca: formuló hipótesis, hizo comprobaciones experimentales y utilizó el méto-do científico para probar la certeza de sus hipótesis. Para sus experimentos utilizó el guisante (Pisum sativum), que presenta características que la hacen idónea para los experimentos planeados: Resultaba fácil conseguir muchas variedades. Las plantas eran fáciles de cultivar, el tiempo de generación es relativa-mente corto, y el número de descendientes de cada siembra, alto. Los descendientes obtenidos al cruzar distintas variedades eran tan fér-tiles corno sus progenitores. No presentan polinización cruzada, pues la corola encierra al estigma y a las anteras, provocando la autopolinización. Esta característica ha desarro-llado razas puras en la naturaleza.
, que presenta características que la hacen idónea para los experimentos planeados: Resultaba fácil conseguir muchas variedades. Las plantas eran fáciles de cultivar, el tiempo de generación es relativa-mente corto, y el número de descendientes de cada siembra, alto. Los descendientes obtenidos al cruzar distintas variedades eran tan fér-tiles corno sus progenitores. No presentan polinización cruzada, pues la corola encierra al estigma y a las anteras, provocando la autopolinización. Esta característica ha desarro-llado razas puras en la naturaleza.")
3
En sus cruzamientos controlados abría las yemas florales antes de que madurase el polen y retiraba los estambres, evitando así la autopoliniza-ción. A continuación espolvoreaba el estigma con polen de otra planta seleccionada a la que se le había abierto la yema floral de la misma forma. De esta forma, tuvo un control total sobre la descendencia. De entre las 34 variedades que Mendel cultivó durante 2 años seleccionó 22 que mostraban constancia para los caracteres escogidos en las distin-tas generaciones (razas puras). Para ello centró su atención en 7 carac-teres que presentaban formas alternativas perfectamente diferenciables. Carácter Variedades Forma de la semilla Lisa y rugosa Color de la semilla Verde y amarilla Altura del tallo Alto y bajo Posición de las flores Apical y lateral Color de la flores Blanco y púrpura Forma de la vaina Color de la vaina
. Para ello centró su atención en 7 carac-teres que presentaban formas alternativas perfectamente diferenciables. Carácter. Variedades. Forma de la semilla. Lisa y rugosa. Color de la semilla. Verde y amarilla. Altura del tallo. Alto y bajo. Posición de las flores. Apical y lateral. Color de la flores. Blanco y púrpura. Forma de la vaina. Color de la vaina.")
5
La primera ley de Mendel
En el primer grupo de experimentos, Mendel cruzó entre sí dos progenito-res de raza pura que diferían en un sólo carácter, repitiendo el experi-mento con las 7 razas puras seleccionadas y efectuando los cruces en las dos direcciones posibles. Fecundó, por ejemplo, flores de plantas de semilla lisa con polen de flores de plantas de semilla rugosa (generación parental o P), y a la inversa, y obtuvo en ambos casos una generación de plantas todas de semilla lisa, a la que llamó primera generación filial (F1). De los dos caracteres alternativos paternos, sólo uno, al que llamó dominante, aparecía en el híbrido de la primera generación filial, quedando oculto el otro carácter, al que llamó recesivo. Basándose en estos hechos, Mendel estableció lo que posteriormente Correns denominaría la 1ª ley de Mendel o Ley de la uniformidad: «cuando se cruzan dos razas puras, todos los descendientes son iguales entre sí»
, y a la inversa, y obtuvo en ambos casos una generación de plantas todas de semilla lisa, a la que llamó primera generación filial (F1). De los dos caracteres alternativos paternos, sólo uno, al que llamó dominante, aparecía en el híbrido de la primera generación filial, quedando oculto el otro carácter, al que llamó recesivo. Basándose en estos hechos, Mendel estableció lo que posteriormente Correns denominaría la 1ª ley de Mendel o Ley de la uniformidad: «cuando se cruzan dos razas puras, todos los descendientes son iguales entre sí»")
6
(P) ♀ Liso x ♂ Rugoso ♀ Rugoso x ♂ Liso (AA) (aa) (aa) (AA) aa¯ ¯
(F1) Liso (Aa) Liso (Aa) Según Mendel los rasgos alternativos para un mismo carácter están regidos por factores (que luego Johannsen llamó genes) que son transmitidos a la descendencia por los gametos de los progenitores. Cada factor puede existir en dos formas alternativas (hoy denominados alelos), cada una de las cuales es responsable de una de las formas alternativas (fenotipos) del carácter. A la forma alternativa dominante se representa por una letra mayúscula (A) y a la recesiva con la misma letra pero minúscula (a).
 Liso (Aa) Liso (Aa) Según Mendel los rasgos alternativos para un mismo carácter están regidos por factores (que luego Johannsen llamó genes) que son transmitidos a la descendencia por los gametos de los progenitores. Cada factor puede existir en dos formas alternativas (hoy denominados alelos), cada una de las cuales es responsable de una de las formas alternativas (fenotipos) del carácter. A la forma alternativa dominante se representa por una letra mayúscula (A) y a la recesiva con la misma letra pero minúscula (a).")
7
1/4 Liso (AA) 2/4 Liso (Aa) 1/4 Liso (aa)
La segunda ley de Mendel En un segundo grupo de experimentos, Mendel plantó y dejó que se auto- fecundaran de forma natural los híbridos obtenidos en los experimentos anteriores (F1). El resultado fue que el carácter recesivo reaparecía en la segunda generación filial (F2), en una relación a 1:3 (un individuo recesivo por cada tres dominantes). (F1) Liso x Liso aaaaaaaaa (Aa) (Aa) Aaaaaaaaaaaaaa ¯ (F2) /4 Liso : 1/4 Rugoso 1/4 Liso (AA) 2/4 Liso (Aa) 1/4 Liso (aa)
. El resultado fue que el carácter recesivo reaparecía en la segunda generación filial (F2), en una relación a 1:3 (un individuo recesivo por cada tres dominantes). (F1) Liso x Liso. aaaaaaaaa (Aa) (Aa) Aaaaaaaaaaaaaa ¯ (F2) 3/4 Liso : 1/4 Rugoso. 1/4 Liso (AA) 2/4 Liso (Aa) 1/4 Liso (aa)")
8
(F1) 1/2 Liso (Aa) : 1/2 Rugoso (aa)
Después, realizó un cruzamiento prueba o retrocruzamiento, que con- siste entre el cruce artificial entre los híbridos y la raza pura recesiva, y obtuvo una descendencia compuesta por la mitad de individuos con el carácter dominante, y la otra mitad con el recesivo. (P) Liso x Rugoso aaaaaaaaa (Aa) (aa) Aaaaaaaaaaaaaa ¯ (F1) /2 Liso (Aa) : 1/2 Rugoso (aa)
 Liso x Rugoso. aaaaaaaaa (Aa) (aa) Aaaaaaaaaaaaaa ¯ (F1) 1/2 Liso (Aa) : 1/2 Rugoso (aa)")
9
De estos dos últimos resultados, Mendel concluyó que los factores here-ditarios que informan para un mismo carácter son independientes, se separan y reparten entre los descendientes, emparejándose al azar. Esto dio origen, posteriormente, al enunciado de la 2ª ley de Mendel o Ley de la segregación: «los dos factores hereditarios que informan sobre un mismo carácter no se mezclan, sino que permanecen diferenciados durante toda la vida del individuo y se segregan (reparten) en el momento de la formación de los gametos»
 en el momento de la formación de los gametos»")
10
(P) Liso Amarillo x Rugoso Verde aaaaaaaaa (LLAA) (llaa)
La tercera ley de Mendel Siguiendo la misma sistemática que en los experimentos anteriores, Mendel realizó cruces entre dihíbridos, es decir, de progenitores que diferían entre sí en dos caracteres. Para ello cruzó las líneas puras (P) liso y amarillo con rugoso y verde, y obtuvo toda la descendencia (F1) de fenotipo uniforme: liso y amarillo. (P) Liso Amarillo x Rugoso Verde aaaaaaaaa (LLAA) (llaa) Aaaaaaaaaaaaaa ¯ (F1) Liso Amarillo (LlAa)
 liso y amarillo con rugoso y verde, y obtuvo toda la descendencia (F1) de fenotipo uniforme: liso y amarillo. (P) Liso Amarillo x Rugoso Verde. aaaaaaaaa (LLAA) (llaa) Aaaaaaaaaaaaaa ¯ (F1) Liso Amarillo. (LlAa)")
11
(F1) Liso Amarillo x Liso Amarillo aaaaaaaaa (LlAa) (LlAa)
Seguidamente, permitió la autofecundación de los individuos de la F1 (dihíbridos) y obtuvo, para los individuos de la F2, las proporciones: 9:3:3:1 (F1) Liso Amarillo x Liso Amarillo aaaaaaaaa (LlAa) (LlAa) Aaaaaaaaaaaaaa ¯ (F2) /16 Liso Amarillo (L_A_) (F2) /16 Liso Verde (L_aa) (F2) /16 Rugoso Amarillo (llA_) (F2) /16 Rugoso Verde (llaa)
 y obtuvo, para los individuos de la F2, las proporciones: 9:3:3:1. (F1) Liso Amarillo x Liso Amarillo. aaaaaaaaa (LlAa) (LlAa) Aaaaaaaaaaaaaa ¯ (F2) 9/16 Liso Amarillo (L_A_) (F2) 3/16 Liso Verde (L_aa) (F2) 3/16 Rugoso Amarillo (llA_) (F2) 1/16 Rugoso Verde (llaa)")
12
Estos resultados dieron lugar al enunciado de lo que se conoce como 3ª ley de Mendel o Ley de la independencia: «los factores hereditarios no antagónicos se heredan independientemente a través de las generaciones, agrupándose al azar en los descendientes»

13
Caracteres patológicos
Las leyes de Mendel se cumplen siempre, pero el concepto de herencia mendeliana se suele re-servar para casos sencillos. En la tabla siguiente se citan algu-nos ejemplos de este tipo de herencia en humanos. Caracteres normales Dominante Recesivo Ojos sesgados Ojos no sesgados Ventanas nasales anchas Ventanas nasales angostas Pestañas largas Pestañas cortas Cabello rizado Cabello lacio Caracteres patológicos Polidactilia Dedos normales Sindactilia Miopía Visión normal Hipermetropía Normal Sordomudez Albinismo Culture in Mendel’s Garden by Ellen K. Levy, 1999

14
Ampliaciones del mendelismo
Existen caracteres que presentan patrones de herencia algo más com-plejos que lo vistos hasta ahora. En estos casos, a pesar de que se siguen las leyes mendelianas, existen diferencias en las frecuencias fenotípicas y/o genotípicas observadas con respecto a las esperadas, por lo que su estudio se considera como una ampliación del mende-lismo. a) Herencia intermedia Entre los dos alelos de un gen que se encuentran en un individuo hetero-cigoto, además de la relación de dominancia-recesividad, se pueden establecer otro tipo de relaciones: Se habla de dominancia parcial cuando la manifestación fenotípica del heterocigoto se encuentra entre los fenotipos de los homocigotos, pareciéndose más a uno que a otro, o incluso siendo exactamente intermedio entre los dos. En este último caso se habla de herencia intermedia y las proporciones fenotípicas que aparecen en la F2 del cruce de dos razas puras es 1:2:1, que es diferente de las esperadas 3:1 (En general AA ≠ Aa ≠ aa).
 Herencia intermedia. Entre los dos alelos de un gen que se encuentran en un individuo hetero-cigoto, además de la relación de dominancia-recesividad, se pueden establecer otro tipo de relaciones: Se habla de dominancia parcial cuando la manifestación fenotípica del heterocigoto se encuentra entre los fenotipos de los homocigotos, pareciéndose más a uno que a otro, o incluso siendo exactamente intermedio entre los dos. En este último caso se habla de herencia intermedia y las proporciones fenotípicas que aparecen en la F2 del cruce de dos razas puras es 1:2:1, que es diferente de las esperadas 3:1 (En general AA ≠ Aa ≠ aa).")
15
Esto ocurre, por ejemplo, en el Don Diego de noche (Minabilis jalaba)
Esto ocurre, por ejemplo, en el Don Diego de noche (Minabilis jalaba). Cuando se cruzan líneas puras de plantas con flores rojas con otras de flores blancas se obtienen una F1 totalmente rosa. Otro caso es el de la superdominancia, en la que el fenotipo del heterocigoto es “superior” (más grande, más viable, etc.) a los dos fenotipos homocigotos (Aa > AA, aa).
. Cuando se cruzan líneas puras de plantas con flores rojas con otras de flores blancas se obtienen una F1 totalmente rosa. Otro caso es el de la superdominancia, en la que el fenotipo del heterocigoto es superior (más grande, más viable, etc.) a los dos fenotipos homocigotos (Aa > AA, aa).")
16
b) Codominancia c) Multialelismo
Es el caso especial en el que los dos alelos del heterocigoto se manifiestan en el fenotipo. En este caso, las frecuencias observadas en la F2 también son 1:2:1. Un ejemplo es el sistema sanguíneo MN en humanos: los individuos homocigotos para el alelo LM presentan el antígeno M en sus hematíes y los individuos homocigotos para el alelo LN presentan el antígeno N, mientras que los heterocigotos presentan ambos (fenotipo Aa = fenotipo AA + fenotipo aa). c) Multialelismo Se habla de multialelismo cuando un carácter está controlado por un sistema alélico múltiple, es decir, está determinado por más de dos alelos diferentes. A pesar de ser una ampliación de las leyes mendelia-nas, las proporciones de sus cruces son las típicas de un caso de mono-hibridismo, ya que cada uno de los individuos que participa en el cruce tendrá sólo dos alelos (iguales o distintos) de la serie alélica. Un ejemplo es el de la herencia del grupo sanguíneo AB0 que está determinado por tres alelos, dos de ellos codominantes entre si (IA e IB) y dominantes sobre el tercero (i).
. c) Multialelismo. Se habla de multialelismo cuando un carácter está controlado por un sistema alélico múltiple, es decir, está determinado por más de dos alelos diferentes. A pesar de ser una ampliación de las leyes mendelia-nas, las proporciones de sus cruces son las típicas de un caso de mono-hibridismo, ya que cada uno de los individuos que participa en el cruce tendrá sólo dos alelos (iguales o distintos) de la serie alélica. Un ejemplo es el de la herencia del grupo sanguíneo AB0 que está determinado por tres alelos, dos de ellos codominantes entre si (IA e IB) y dominantes sobre el tercero (i).")
17
(bw+bw+ cd+cd+) (bwbw cdcd)
d) Interacciones génicas y epistasia Se dice que existen interacciones génicas cuando dos o más genes intervienen en la determinación fenotípica de un mismo carácter. La interacción génica se pone de manifiesto cuando entre los individuos de la F2 aparecen fenotipos diferentes a los de los parentales, aunque sus proporciones se ajustan a la relación 9:3:3:1 (para el caso de dos loci). Un ejemplo de esto ocurre con el color de los ojos en Drosophila. Si se cruza una cepa normal de ojos rojos (fenotipo silvestre) con otra de color rojo claro (mutante) se obtienen los siguientes resultados: (P) Ojos rojos x Ojos rosa claro (bw+bw+ cd+cd+) (bwbw cdcd) (F1) Ojos rojos (bw+bw cd+cd) (F2) /16 Ojos rojos (bw+_ cd+_) 3/16 Ojos marrones (bwbw cd+_) 3/16 Ojos rojo cardenal (bw+_ cdcd) 1/16 Ojos rosa claro (bwbw cdcd)
 Interacciones génicas y epistasia. Se dice que existen interacciones génicas cuando dos o más genes intervienen en la determinación fenotípica de un mismo carácter. La interacción génica se pone de manifiesto cuando entre los individuos de la F2 aparecen fenotipos diferentes a los de los parentales, aunque sus proporciones se ajustan a la relación 9:3:3:1 (para el caso de dos loci). Un ejemplo de esto ocurre con el color de los ojos en Drosophila. Si se cruza una cepa normal de ojos rojos (fenotipo silvestre) con otra de color rojo claro (mutante) se obtienen los siguientes resultados: (P) Ojos rojos x Ojos rosa claro. (bw+bw+ cd+cd+) (bwbw cdcd) (F1) Ojos rojos (bw+bw cd+cd) (F2) 9/16 Ojos rojos (bw+_ cd+_) 3/16 Ojos marrones (bwbw cd+_) 3/16 Ojos rojo cardenal (bw+_ cdcd) 1/16 Ojos rosa claro (bwbw cdcd)")
18
Las moscas con ojos color rosa claro de la F2 (1/16) tienen un genotipo doble homocigoto recesivo, mientras que las normales (9/16) llevarán, como mínimo, un alelo dominante de cada gen. Las proporciones 3/16 corresponden a la homocigosis recesiva para un loci y la presencia de, al menos, un alelo dominante en el otro loci. En este caso los genes que intervienen son el brown (bw+/bw) y el cardinal (cd+/cd). Cuando la interacción génica tiene como consecuencia la alteración de las proporciones mendelianas 9:3:3:1, se habla de epistasis. En este caso, el gen de un locus (epistático) suprime o enmascara la acción de un gen en otro locus (hipostático). Cuando la epistasis opera entre dos loci, el número de fenotipos que aparecen en la descendencia de progenitores dihíbridos será menor de 4. Existen seis tipos de proporciones epistáticas reconocidas común-mente, tres de las cuales tienen tres fenotipos, mientras que las tres restantes sólo tienen dos.
 suprime o enmascara la acción de un gen en otro locus (hipostático). Cuando la epistasis opera entre dos loci, el número de fenotipos que aparecen en la descendencia de progenitores dihíbridos será menor de 4. Existen seis tipos de proporciones epistáticas reconocidas común-mente, tres de las cuales tienen tres fenotipos, mientras que las tres restantes sólo tienen dos.")
19
Epistasis dominante (12:3:1)
Cuando el alelo dominante de un locus, por ejemplo, el alelo A, produce un cierto fenotipo sin importar la condición alélica del otro locus, enton-ces se dice que el locus A es epistático para el locus B. Además, debido a que el alelo dominante A es capaz de expresarse aun en presencia de B o b, éste es un caso de epistasis dominante. Sólo cuando el genotipo de un individuo es homo-cigoto recesivo para el locus epistático (aa), los alelos del locus hipostáti-co (B o b) podrán expre-sarse. Así, los genotipos A_B_ y A_bb producen el mismo fenotipo y aaB_ y aabb producen dos feno-tipos adicionales. La pro-porción 9:3:3:1 se modifi-ca a una proporción 12:3:1
, los alelos del locus hipostáti-co (B o b) podrán expre-sarse. Así, los genotipos A_B_ y A_bb producen el mismo fenotipo y aaB_ y aabb producen dos feno-tipos adicionales. La pro-porción 9:3:3:1 se modifi-ca a una proporción 12:3:1.")
20
Epistasis recesiva (9:3:4)
Si el genotipo recesivo en un locus (por ejemplo aa) suprime la expre-sión de los alelos en el locus B, se dice que el locus A muestra epistasis recesiva sobre el locus B. Sólo si está presente el alelo dominante en el locus A, los alelos del locus hipostático B pueden expresarse. Los geno-tipos A_B_ y A_bb producen dos fenotipos adicionales. La proporción 9:3 :3:1 se convierte en una proporción 9:3:4.
 suprime la expre-sión de los alelos en el locus B, se dice que el locus A muestra epistasis recesiva sobre el locus B. Sólo si está presente el alelo dominante en el locus A, los alelos del locus hipostático B pueden expresarse. Los geno-tipos A_B_ y A_bb producen dos fenotipos adicionales. La proporción 9:3 :3:1 se convierte en una proporción 9:3:4.")
21
Genes duplicados con efecto acumulativo (9:6:1)
Si la condición dominante (homocigoto o heterocigoto) en uno o en otro locus (pero no en ambos) produce el mismo fenotipo, la proporción de la F2 se convierte en 9:6:1. Por ejemplo, cuando los genes epistáticos están involucrados en la producción de varias cantidades de una sustan-cia como los pigmentos, puede considerarse que los genotipos domi-nantes de cada locus producen independientemente una unidad de pigmento. Así, los genotipos A_bb y aaB_ producen cada uno una unidad de pigmento y tienen, por lo tanto el mismo fenotipo. El genotipo aabb no produce pigmento, pero en el genotipo A_B_ el efecto es acumulativo y se producen dos unidades de pigmento.
 en uno o en otro locus (pero no en ambos) produce el mismo fenotipo, la proporción de la F2 se convierte en 9:6:1. Por ejemplo, cuando los genes epistáticos están involucrados en la producción de varias cantidades de una sustan-cia como los pigmentos, puede considerarse que los genotipos domi-nantes de cada locus producen independientemente una unidad de pigmento. Así, los genotipos A_bb y aaB_ producen cada uno una unidad de pigmento y tienen, por lo tanto el mismo fenotipo. El genotipo aabb no produce pigmento, pero en el genotipo A_B_ el efecto es acumulativo y se producen dos unidades de pigmento.")
22
Genes dominantes duplicados (15:1)
La proporción 9:3:3:1 se modifica a 15:1 si los alelos dominantes de ambos loci producen cada uno el mismo fenotipo sin efecto acumulativo.

23
Genes recesivos duplicados o complementarios (9:7)
En el caso en donde ambos genotipos homocigotos recesivos producen fenotipos idénticos, la proporción en la F2 se modifica a 9:7. Los genotipos aaB_, A_bb y aabb producen un solo fenotipo. Cuando se presentan juntos ambos alelos dominantes, se complementan uno con otro y producen un fenotipo diferente.

24
Interacción de dominantes y recesivos (13:3)
Sólo resultan dos fenotipos en la F2 cuando un genotipo dominante en un locus (por ejemplo A_) y el genotipo recesivo en el otro (bb) producen el mismo efecto fenotípico. Así, A_B_, A_bb y aabb producen un mismo fenotipo y aaB_ producen otro en la proporción 13:3.
 y el genotipo recesivo en el otro (bb) producen el mismo efecto fenotípico. Así, A_B_, A_bb y aabb producen un mismo fenotipo y aaB_ producen otro en la proporción 13:3.")
25
Resumen de las relaciones epistáticas
Genotipos A_B_ A_bb aaB_ aabb Proporción clásica 9 3 1 Epistasis dominante 12 Epistasis recesiva 4 Genes duplicados con efecto acumulativo 6 Genes dominantes duplicados 15 Genes recesivos duplicados 7 Interacción de genes dominantes y recesivos 13

26
e) Herencia multifactorial
En este tipo de herencia están implicados varios genes que contribuyen a la expre-sión del fenotipo de un modo acumula-tivo o cuantitativo. Los individuos de la F1 presentan un carácter intermedio entre los dos progenitores y en la F2, la mayor parte de los individuos son intermedios (como los de la F1), mientras que sólo unos pocos presentan los caracteres ex-tremos de los parentales. Este patrón fenotípico se corresponde con una distri-bución de tipo normal (campana Gauss), es decir, los caracteres presentan una variabilidad continua. Un ejemplo lo constituye la herencia de la coloración de las semillas de trigo.
, mientras que sólo unos pocos presentan los caracteres ex-tremos de los parentales. Este patrón fenotípico se corresponde con una distri-bución de tipo normal (campana Gauss), es decir, los caracteres presentan una variabilidad continua. Un ejemplo lo constituye la herencia de la coloración de las semillas de trigo.")
27
f) Letalidad Las variantes alélicas de un gen no sólo pueden establecer distintos tipos de relación entre sí, sino también reducir (o aumentar) la super-vivencia de los individuos, incluso provocando su muerte, en fases tempranas o tardías del desarrollo (letalidad). Este efecto sobre la viabilidad complica el análisis mendeliano, ya que modifica, en muchos casos, las proporciones esperadas. Los genes letales dominantes producen efecto letal tanto en homoci-gosis como en heterocigosis, mientras que los genes letales recesivos sólo producen el efecto letal en homocigosis. Un ejemplo de esto ocurre en el caso del color del pelaje de los ratones. El alelo que proporciona el pelaje amarillo (Y) nunca puede dar origen a una raza pura, porque en homocigosis (YY), los embriones mueren tempranamente. Sólo sobreviven los genotipos Y+ (amarillo) y ++ (agutí). El alelo Y es dominante en cuanto al color del pelo, pero es recesivo respecto a la letalidad, ya que el gen sólo es letal en homocigosis.
 la super-vivencia de los individuos, incluso provocando su muerte, en fases tempranas o tardías del desarrollo (letalidad). Este efecto sobre la viabilidad complica el análisis mendeliano, ya que modifica, en muchos casos, las proporciones esperadas. Los genes letales dominantes producen efecto letal tanto en homoci-gosis como en heterocigosis, mientras que los genes letales recesivos sólo producen el efecto letal en homocigosis. Un ejemplo de esto ocurre en el caso del color del pelaje de los ratones. El alelo que proporciona el pelaje amarillo (Y) nunca puede dar origen a una raza pura, porque en homocigosis (YY), los embriones mueren tempranamente. Sólo sobreviven los genotipos Y+ (amarillo) y ++ (agutí). El alelo Y es dominante en cuanto al color del pelo, pero es recesivo respecto a la letalidad, ya que el gen sólo es letal en homocigosis.")
28
g) Herencia y sexo Se habla de este tipo de herencia cuando en ella intervienen genes cuyo locus se encuentra en los cromosomas sexuales. En función de la localización de los genes en las distintas regiones de estos cromosomas, se pueden establecer tres modos de herencia: herencia ligada al X (o al Z), herencia ligada al Y (o al W) y herencia parcialmente ligada al sexo. Además hay un cuarto caso que es la herencia influenciada por el sexo. Herencia ligada al cromosoma X (hologínica) Se da en genes que se encuentran en la región exclusiva del cromo-soma X. Por ejemplo, el gen para el daltonismo en humanos (d) recesivo respecto al alelo normal (+), se comporta de la siguiente manera en los cruces recíprocos:
, herencia ligada al Y (o al W) y herencia parcialmente ligada al sexo. Además hay un cuarto caso que es la herencia influenciada por el sexo. Herencia ligada al cromosoma X (hologínica) Se da en genes que se encuentran en la región exclusiva del cromo-soma X. Por ejemplo, el gen para el daltonismo en humanos (d) recesivo respecto al alelo normal (+), se comporta de la siguiente manera en los cruces recíprocos:")
29
Cruce A Padres ♀ XdXd x ♂ X+Y Gametos P Xd X+ Y Adultos F1 X+Xd XdY Fenotipo + Daltónico Gametos F1 Adultos F2 X+ Xd Xd Xd X+Y Dalt. Sexo Mujeres Hombres Cruce B Padres ♀ X+X+ x ♂ XdY Gametos P X+ Xd Y Adultos F1 X+Xd X+Y Fenotipo + Gametos F1 Adultos F2 X+X+ XdY Dalt. Sexo Mujeres Hombres En el cruce A, el resultado de la F1 (hembras normales, como el padre, y machos daltónicos, como la madre) nos da la pista de una herencia ligada al cromosoma X. En el cruce B, La F1 es normal, pero en la F2, aunque aparecen 3/4 normales + 1/4 daltónicos, como en el monohibridis-mo, los daltónicos son sólo hombres, lo que nos avisa de su ligamiento al cromo-soma X.
 nos da la pista de una herencia ligada al cromosoma X. En el cruce B, La F1 es normal, pero en la F2, aunque aparecen 3/4 normales + 1/4 daltónicos, como en el monohibridis-mo, los daltónicos son sólo hombres, lo que nos avisa de su ligamiento al cromo-soma X.")
30
Herencia ligada al cromosoma Y (holándrica)
Se da en genes situados en la región exclusiva del cromosoma Y, y sólo se transmite por vía masculina. Herencia parcialmente ligada al sexo Se da en genes situados en la región homóloga entre el cromosoma X y el Y (o entre el Z y el W). Por el hecho de que tanto las hembras como los machos pueden ser homo o heterocigotos, a esta región se le llama frecuentemente pseudoautosómica. Veamos el caso de la herencia del gen bobbed (bb), situado en la región homóloga entre el cromosoma X y el Y de Drosophila, que da lugar a quetas más cortas de lo normal.
. Por el hecho de que tanto las hembras como los machos pueden ser homo o heterocigotos, a esta región se le llama frecuentemente pseudoautosómica. Veamos el caso de la herencia del gen bobbed (bb), situado en la región homóloga entre el cromosoma X y el Y de Drosophila, que da lugar a quetas más cortas de lo normal.")
31
Cruce A Padres ♀ XbbXbb x ♂ X+Y+ Gametos P Xbb X+ Y Adultos F1 X+Xbb XbbY+ Fenotipo + Gametos F1 Y+ Adultos F2 XbbXbb X+Y+ Mutante Sexo Hembras Machos En el cruce A, la F2 da una proporción de 3/4 normales + 1/4 mutantes, pero éstos son exclusiva-mente hembras. Cruce B Padres ♀ X+X+ x ♂ XbbYbb Gametos P X+ Xbb Ybb Adultos F1 X+Xbb X+Ybb Fenotipo + Gametos F1 Adultos F2 X+X+ XbbYbb Mutante Sexo Hembras Machos En el cruce B, ocurre lo contrario, 1/4 de los in-dividuos de la F2 son mutantes y machos. Hay que tener cuidado con los resultados de este último cruce, ya que se pueden confundir con herencia ligada al cromosoma X.

32
Herencia influenciada por el sexo
Se da en genes autosómicos cuyos alelos invierten su relación de dominancia/recesividad según el sexo (debido, normalmente a la influen-cia de las hormonas sexuales). Por ejemplo, la presencia de cuernos en ciertas razas de ovinos depen-de de un gen con dos alelos, C y c, de tal manera que el heterocigoto tendrá cuernos si es macho y no los tendrá si es hembra. En este caso, el alelo C se comporta como dominante en machos y como recesivo en mujeres. Genotipo Machos Hembras CC Cuernos Cc Mocha cc Mocho
. Por ejemplo, la presencia de cuernos en ciertas razas de ovinos depen-de de un gen con dos alelos, C y c, de tal manera que el heterocigoto tendrá cuernos si es macho y no los tendrá si es hembra. En este caso, el alelo C se comporta como dominante en machos y como recesivo en mujeres. Genotipo. Machos. Hembras. CC. Cuernos. Cc. Mocha. cc. Mocho.")
33
2. La teoría cromosómica de la herencia
Walter Sutton Walter Flemming Carl Correns Hugo De Vries Erich Tschermak Theodor Boveri Van Beneden Thomas H. Morgan Alfred H. Sturtevant Calvin Bridges

34
Introducción Entre finales del siglo XIX y principios del XX se establecieron las bases que acabarían por dar lugar a la teoría cromosómica de la herencia. Los avances citológicos permitieron el descubrimiento de los cromo-somas en el núcleo celular por Walter Flemming en 1879, así como sus patrones de comportamiento durante los procesos de la mitosis (Van Beneden, 1883) y de la meiosis (Sutton y Boveri, 1902). Por otro lado, en el 1900, tuvo lugar el redescubrimiento de las teorías de Mendel, gracias a las investigaciones de Correns, De Vries y Tschermak. Basándose en estas observaciones, y de manera independiente, Walter Sutton y Theodor Boveri relacionaron el comportamiento de los cromo-somas con los principios mendelianos de la segregación y de la trans-misión independiente, señalando que su separación durante la meiosis podría ser la base citológica de ambos fenómenos. Aunque pensaban que los factores hereditarios de Mendel eran los cromosomas, más que los genes en los cromosomas, sus hallazgos sirvieron de fundamento para posteriores investigaciones en genética. Es por ello que a ambos autores se les reconoce como los iniciadores de la teoría cromosómica de la herencia.
 y de la meiosis (Sutton y Boveri, 1902). Por otro lado, en el 1900, tuvo lugar el redescubrimiento de las teorías de Mendel, gracias a las investigaciones de Correns, De Vries y Tschermak. Basándose en estas observaciones, y de manera independiente, Walter Sutton y Theodor Boveri relacionaron el comportamiento de los cromo-somas con los principios mendelianos de la segregación y de la trans-misión independiente, señalando que su separación durante la meiosis podría ser la base citológica de ambos fenómenos. Aunque pensaban que los factores hereditarios de Mendel eran los cromosomas, más que los genes en los cromosomas, sus hallazgos sirvieron de fundamento para posteriores investigaciones en genética. Es por ello que a ambos autores se les reconoce como los iniciadores de la teoría cromosómica de la herencia.")
35
Posteriormente, los trabajos acerca del fenómeno del ligamiento en los genes de Drosophila realizados por Thomas H. Morgan, Alfred H. Sturtevant y Calvin Bridges demostraron, más allá de toda duda, la veracidad de esta teoría, que se concreta en los siguientes puntos: Los factores de la herencia mendelianos, ahora llamados genes, residen en lugares concretos (locus) dentro de los cromosomas. Los distintos loci de cada cromosoma estan dispuestos linealmente, unos a continuación de los otros. Los cromosomas homólogos poseen series idénticas de loci, y los genes que ocupan cada loci homólogo son los factores antagó-nicos o alelos. Existe una correspondencia directa entre el fenómeno genético de la herencia y el fenómeno citológico de intercambio de fragmen-tos cromosómicos durante la meiosis.
 dentro de los cromosomas. Los distintos loci de cada cromosoma estan dispuestos linealmente, unos a continuación de los otros. Los cromosomas homólogos poseen series idénticas de loci, y los genes que ocupan cada loci homólogo son los factores antagó-nicos o alelos. Existe una correspondencia directa entre el fenómeno genético de la herencia y el fenómeno citológico de intercambio de fragmen-tos cromosómicos durante la meiosis.")
36
Ligamiento y recombinación
Las investigaciones genéticas han puesto de manifiesto que, en muchos organismos, ciertos genes no se transmiten de acuerdo con la 3ª ley de Mendel o de la transmisión independiente, sino que parecen segregar como si estuviesen unidos, de manera que se alteran las proporciones esperadas en la descendencia. Experiencias, como las de Morgan, demostraron que tales genes formaban parte del mismo cromosoma y que se transmitían como si fuesen unidades únicas o sencillas. Se dice que los genes que forman parte del mismo cromosoma están ligados y muestran ligamiento en los cruces genéticos. De esta manera, los alelos de todos los loci de un cromosoma se trans-mitirán, en teoría, como una unidad en la formación de los gametos. Sin embargo, en muchos casos, durante la primera profase meiótica, cuando los homólogos se aparean (sinapsis), puede tener lugar un intercambio recíproco de fragmentos entre cromosomas (entrecruzamiento) y dar lugar a una mezcla o recombinación de los alelos entre homólogos. Esto se visualiza al microscopio como un contacto entre los cromosomas (quiasma).
, puede tener lugar un intercambio recíproco de fragmentos entre cromosomas (entrecruzamiento) y dar lugar a una mezcla o recombinación de los alelos entre homólogos. Esto se visualiza al microscopio como un contacto entre los cromosomas (quiasma).")
37
Acoplamiento Repulsión
El grado de entrecruzamiento entre dos loci cualesquiera de un cromo-soma es proporcional a la distancia que los separa. Así, el porcentaje de gametos recombinantes varía dependiendo de qué loci están siendo considerados. La manera de indicar que dos genes, con sus correspondientes alelos, por ejemplo, el A/a y el B/b, están ligados, es ponerlos en forma de quebrado: A B A b a b a B Acoplamiento Repulsión En el primer caso el individuo tiene los dos alelos dominantes en un cro-mosoma y los dos recesivos en el homólogo, esta situación se denomina de acoplamiento (posición cis) y la contraria de repulsión (trans). Un entrecruzamiento dado se produce sólo entre dos cromátidas no hermanas de una tétrada, por ello, cuando en una meiosis hay un entrecruzamiento entre dos locus dados, se producirán cuatro tipos de gametos: dos parentales y dos recombinantes.
 y la contraria de repulsión (trans). Un entrecruzamiento dado se produce sólo entre dos cromátidas no hermanas de una tétrada, por ello, cuando en una meiosis hay un entrecruzamiento entre dos locus dados, se producirán cuatro tipos de gametos: dos parentales y dos recombinantes.")
38
Frecuencias de recombinación y mapas cromosómicos
La frecuencia de recombinación es el resultado de dividir el número de descendientes recombinantes por el total de individuos obtenidos en un cruce prueba de la F1. FR = Tipos recombinantes / Total descendientes Morgan sugirió que las frecuencias de recombinación deberían ser pro-porcionales a las distancias entre los loci considerados. Cuanto más cerca estén dos loci entre sí, más difícil será que recombinen y, por tanto, menor será dicha frecuencia. Esta correlación sirve de base para la elaborción de mapas cromosó-micos, que proporcionan la situación relativa de los genes en los cromosomas. De este modo, la distancia entre genes también se puede indicar en unidades de mapa (u.m.). Una unidad de mapa equivale a un 1% de recombinación (p = 0’01) o a 1 centiMorgan (cM). 1 u.m. = 1 cM = 1% recombinación = probabilidad del 0’01
. Una unidad de mapa equivale a un 1% de recombinación (p = 0’01) o a 1 centiMorgan (cM). 1 u.m. = 1 cM = 1% recombinación = probabilidad del 0’01.")
39
Si dos loci están tan separados que la probabilidad de que se produzca un quiasma entre ellos es del 100%, entonces el 50% de los gametos serán recombinantes y el otro 50% serán parentales. Cuando en estos casos se realiza un cruzamiento prueba a los individuos F1, se espera una descendencia con proporciones 1:1:1:1, igual que si estuviesen en cromosomas distintos; por tanto, no podremos diferenciar el ligamiento de la transmisión independiente. De este modo, los porcentajes máximo y mínimo de gametos recombi-nados van desde el 50%, cuando entre dos genes dados siempre hay entrecruzamiento en la meiosis, hasta el 0%, cuando nunca hay entre-cruzamiento. Por ello, la frecuencia de entrecruzamiento (y la frecuencia de quiasmas) entre dos genes dados es el doble de su frecuencia de recombinación.
 entre dos genes dados es el doble de su frecuencia de recombinación.")
40
Mapas de tres puntos Para los genes más apartados, es conveniente emplear un tercer marca-dor entre los otros dos loci, con el fin de detectar cualquier doble entre-cruzamiento entre ambos. Supóngase un cruce prueba con trihíbridos. Los dos fenotipos no recombinantes (parentales) son los que se encuen-tran en mayor proporción, mientras que los dobles recombinantes (DR), debido a su probabilidad de aparición, son los que se encuentran en menor número. Las cuatro clases fenotípicas restantes representan a dos categorías que resultan de entrecruzamientos sencillos, RS (en la zona I y en la zona II, respectivamente). Para calcular las distancias entre dos loci hay que tener en cuenta a los dobles recombinantes (DR) de la siguiente forma: RSI + DR FRA-B = Total RSII + DR FRB-C = Total FRA-C = FRA-B + FRB-C
 son los que se encuen-tran en mayor proporción, mientras que los dobles recombinantes (DR), debido a su probabilidad de aparición, son los que se encuentran en menor número. Las cuatro clases fenotípicas restantes representan a dos categorías que resultan de entrecruzamientos sencillos, RS (en la zona I y en la zona II, respectivamente). Para calcular las distancias entre dos loci hay que tener en cuenta a los dobles recombinantes (DR) de la siguiente forma: RSI + DR. FRA-B = Total. RSII + DR. FRB-C = Total. FRA-C = FRA-B + FRB-C.")
41
CC = % DR esperados Interferencia y coeficiente de coincidencia
En la mayor parte de los organismos superiores, la formación de un quiasma reduce en forma efectiva la probabilidad de la formación de otro en una región cromosómica inmediatamente adyacente, posiblemente por la reducción de la capacidad física de las cromátidas para doblarse sobre si mismas hacia atrás. El resultado de esta interferencia es la observación de un número menor de dobles recombinantes de los que cabría esperar de acuerdo con la distancia de mapeo. El porcentaje entre frecuencias observadas y las esperadas se conoce como coeficiente de coincidencia (CC). % DR observados CC = % DR esperados
. % DR observados. CC = % DR esperados.")
42
Los DR esperados se calculan como FRA-B x FRB-C expresado en tanto por uno (probabilidad).
De este modo, la interferencia (I) se calcula como: I = 1 – CC Cuando la interferencia es completa no se observan entrecruzamientos de intercambio doble y la coincidencia será cero. Cuando observamos el total de cruzamientos de intercambio dobles esperados, la coincidencia es total y la interferencia es igual a cero.
 se calcula como: I = 1 – CC. Cuando la interferencia es completa no se observan entrecruzamientos de intercambio doble y la coincidencia será cero. Cuando observamos el total de cruzamientos de intercambio dobles esperados, la coincidencia es total y la interferencia es igual a cero.")
43
3. Las mutaciones Concepto de mutación
Una de las características del material hereditario, el ADN, es la gran fidelidad con que se transmite de generación en generación; sin embar-go, en ocasiones puede sufrir cambios al azar que se pueden transmitir a la descendencia. Estos cambios reciben el nombre de mutaciones. Normalmente suponen deficiencias y pueden llegar a ser letales aunque, por lo general, son recesivas y permanecen ocultas. Pese a ser casi siempre negativas para el individuo, comportan un aspecto positivo para la especie, ya que aportan variabilidad a la población. Ello permite que, si se produce un cambio en el ambiente y las nuevas condiciones son muy adversas para los individuos normales, la existencia de individuos mutantes hace que pueda haber algunos que soporten esas condiciones y que, gracias a ellos, por selección natural, la especie no se extinga. De este modo, las mutaciones pueden permitir la evolución de las especies.

44
Las mutaciones pueden darse en células somáticas (mutaciones somá-ticas) y en células reproductoras (mutaciones germinales). Las que se dan en las células somáticas, salvo que las conviertan en células cance-rosas, carecen de importancia, ya que si las células no son viables, pueden ser sustituidas por otras células. Sin embargo, las mutaciones germinales sí son trascendentales, ya que todas las células del nuevo organismo tendrán la misma información que la célula cigoto. Las mutaciones pueden aparecer espontáneamente (mutaciones natu-rales) o pueden ser provocadas artificialmente (mutaciones inducidas) mediante radiaciones y ciertas sustancias químicas, los denominados agentes mutágenos. En los humanos la tasa de mutación espontánea es de un gen mutado por cada genes. De esto resulta que, por término medio, haya un gen mutado en cada gameto y, en consecuencia, se incorporen dos genes mutados por individuo (zigoto). Este valor es conocido como «carga genética de efectos negativos». Esta “carga” va aumentando en cada generación, sobre todo en las poblaciones más desarrolladas, ya que en ellas el uso de fármacos permite la supervivencia de los afectados.
 o pueden ser provocadas artificialmente (mutaciones inducidas) mediante radiaciones y ciertas sustancias químicas, los denominados agentes mutágenos. En los humanos la tasa de mutación espontánea es de un gen mutado por cada genes. De esto resulta que, por término medio, haya un gen mutado en cada gameto y, en consecuencia, se incorporen dos genes mutados por individuo (zigoto). Este valor es conocido como «carga genética de efectos negativos». Esta carga va aumentando en cada generación, sobre todo en las poblaciones más desarrolladas, ya que en ellas el uso de fármacos permite la supervivencia de los afectados.")
45
Según la extensión del material genético afectado, se distinguen tres tipos de mutaciones:
Mutaciones génicas (alteraciones de la secuencia de nucleótidos de un gen). Mutaciones cromosómicas (alteraciones de la secuencia de genes de un cromosoma). Mutaciones genómicas (alteraciones en el número de cromosomas).
. Mutaciones cromosómicas (alteraciones de la secuencia de genes de un cromosoma). Mutaciones genómicas (alteraciones en el número de cromosomas).")
46
Las mutaciones génicas
Son alteraciones en la secuencia de nucleótidos de un gen. Por ello tam-bién se las denomina mutaciones puntuales. Según el tipo de altera-ción se clasifican en mutaciones por sustitución de bases y en mutacio-nes por pérdida o inserción de nucleótidos. a) Mutaciones por sustitución de bases Son cambios de una base por otra. Como hay dos tipos de bases, las púricas (A y G), y las pirimidínicas (T y C), se distinguen dos tipos de sustituciones de bases: las transiciones, que consisten en la sustitución de una purina por otra, o de una pirimidina por otra, y las transversio-nes, que son sustituciones de una purina por una pirimidina, o viceversa. Las sustituciones provocan la alteración de un único triplete y, por tanto, salvo que indiquen un triplete de parada, o un aminoácido del centro activo de una enzima, no suelen ser perjudiciales. Constituyen solamen-te el 20 % de las mutaciones génicas espontáneas.
 Mutaciones por sustitución de bases. Son cambios de una base por otra. Como hay dos tipos de bases, las púricas (A y G), y las pirimidínicas (T y C), se distinguen dos tipos de sustituciones de bases: las transiciones, que consisten en la sustitución de una purina por otra, o de una pirimidina por otra, y las transversio-nes, que son sustituciones de una purina por una pirimidina, o viceversa. Las sustituciones provocan la alteración de un único triplete y, por tanto, salvo que indiquen un triplete de parada, o un aminoácido del centro activo de una enzima, no suelen ser perjudiciales. Constituyen solamen-te el 20 % de las mutaciones génicas espontáneas.")
47
b) Mutaciones por pérdida o inserción de nucleótidos
Estas mutaciones se denominan deleciones o adiciones, respectiva-mente. Como el mensaje genético se traduce de triplete en triplete, las deleciones y adiciones, salvo que se compensen entre sí, producen un corrimiento en el orden de lectura y, por tanto, alteran todos los tripletes siguientes. Sus consecuencias suelen ser graves. Constituyen el 80 % de las mutaciones génicas espontáneas. c) Causas de las mutaciones génicas Las mutaciones génicas pueden producirse por tres causas: por errores de lectura durante la replicación del ADN, por lesiones fortuitas o por transposiciones (cambios de posición) de ciertos segmentos del gen.
 Causas de las mutaciones génicas. Las mutaciones génicas pueden producirse por tres causas: por errores de lectura durante la replicación del ADN, por lesiones fortuitas o por transposiciones (cambios de posición) de ciertos segmentos del gen.")
48
Cambio de fase que da lugar a la adición de T
Errores de lectura. Los errores de lectura que pueden aparecer durante la replicación del ADN pueden deberse a dos causas: Cambios tautoméricos. Cada base nitrogenada puede presentarse en dos formas diferentes denominadas tautómeros, siendo una normal y la otra rara. Ambas formas están en equilibrio, y espontáneamente se pasa de una a la otra, lo que se denomina cambio tautomérico. Esto, si sucede durante la replicación, implica mutaciones, ya que cambia la base complementaria en la nueva hebra de ADN. Por ejemplo, la forma normal de la G se complementa con la C, mientras que la forma rara de G lo hace con la T. Cambios de fase. Son deslizamientos de la hebra que se está for-mando sobre la hebra molde, de forma que quedan bucles al volverse a emparejar. El crecimiento sigue y la diferencia queda fijada, origi-nándose así la mutación. Cambio de fase que da lugar a la adición de T

49
Lesiones fortuitas. Son alteraciones de la estructura de uno o de varios nucleótidos, que aparecen de forma natural. Las más frecuentes son: Despurinización. Pérdida de purinas por rotura del enlace entre éstas y las desoxirribosas. Se producen a razón de unas a por día, en cada célula humana. Desaminación. Pérdida de grupos amino en las bases nitrogenadas, que entonces se emparejan con una distinta de la normal. Dímeros de timina. Se forman enlaces entre dos timinas contiguas, generalmente debido a los rayos ultravioleta de la radiación solar, que alteran la formación de las cadenas complementarias de ADN. Transposiciones. Son cambios de lugar espontáneos de determinados segmentos de ADN. Estos pueden ser menores que un gen (secuencias de inserción), un gen, o un grupo de genes (transposones). Las trans-posiciones pueden producir mutaciones génicas si el elemento genético transpuesto se sitúa dentro de un gen, o mutaciones cromosómicas si pasa a un lugar donde no hay un gen, ya sea dentro del mismo cromo-soma o incluso a otro cromosoma.
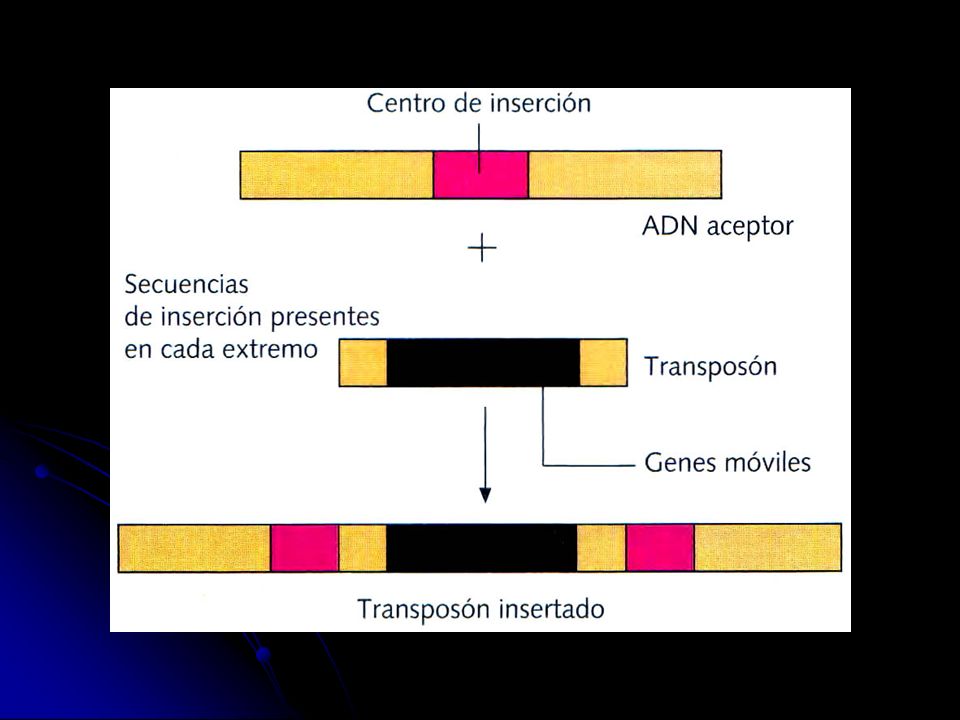
, un gen, o un grupo de genes (transposones). Las trans-posiciones pueden producir mutaciones génicas si el elemento genético transpuesto se sitúa dentro de un gen, o mutaciones cromosómicas si pasa a un lugar donde no hay un gen, ya sea dentro del mismo cromo-soma o incluso a otro cromosoma.")
51
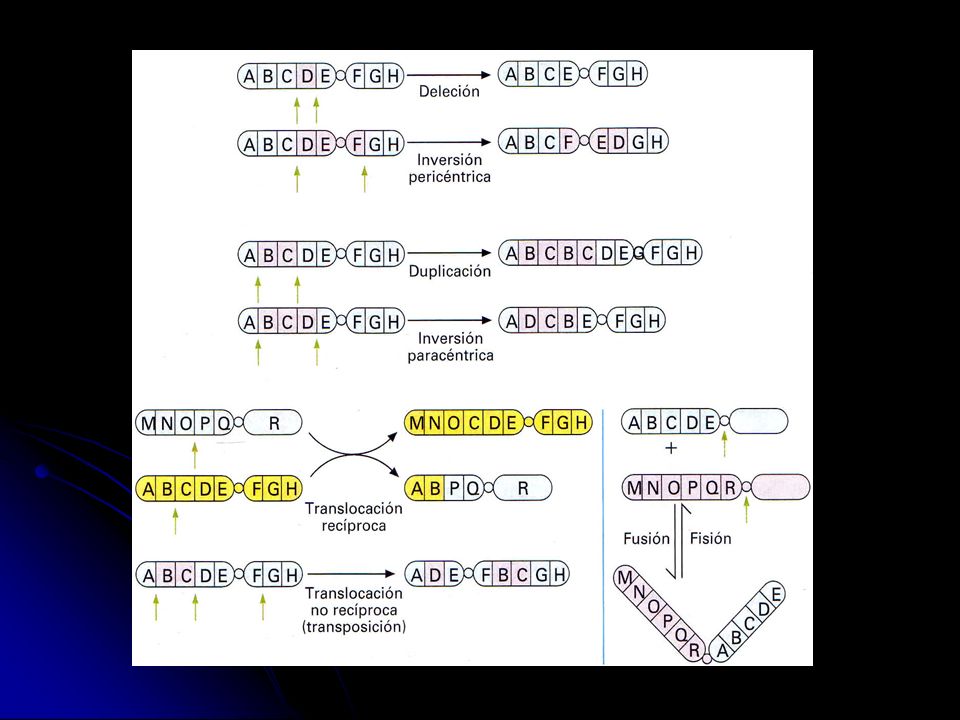
Las mutaciones cromosómicas
Son mutaciones que provocan cambios en la estructura interna de los cromosomas. Se distinguen los siguientes tipos: Deleción. Es la pérdida de un fragmento del cromosoma. Si el frag-mento contiene muchos genes, la deleción puede tener consecuen-cias patológicas o incluso letales. Si una deleción afecta a los dos cromosomas homólogos, suele ser letal. Duplicación. Es la repetición de un segmento de un cromosoma. La réplica puede hallarse en el mismo cromosoma, haberse unido a un cromosoma no homólogo, o incluso haberse independizado con su propio centrómero. Las duplicaciones permiten aumentar el material genético y, gracias a posteriores mutaciones, pueden determinar la aparición de nuevos genes durante el proceso evolutivo. Inversión. Es el cambio de sentido de un fragmento en el cromoso-ma. No suelen comportar perjuicios al individuo pero sí a los descen-dientes si durante la meiosis se produce un entrecruzamiento dentro de la inversión.

52
Translocación. Es el cambio de posición de un segmento de cromosoma
Translocación. Es el cambio de posición de un segmento de cromosoma. Cuando se produce por intercambio de segmentos entre dos cromosomas no homólogos, que es lo más frecuente, se denomina translocación recíproca. Cuando sólo hay traslación de un segmento a otro lugar del mismo cromosoma o de otros cromosomas, sin reciprocidad, se denomina transposición. Las translocaciones no suelen perjudicar al individuo que las ha sufrido pero sí a su descendencia, ya que puede heredar un cromosoma incompleto o con duplicaciones. Estos descendientes heredan de cada progenitor un solo cromosoma de cada tipo y éste, con más posibilidades todavía si ha habido recombinación, puede ser

54
Las mutaciones genómicas
Son las alteraciones en el número de cromosomas propio de una espe-cie. Se distinguen estos tipos: aneuploidías y euploidías. Aneuploidía. Son alteraciones en el número normal de ejemplares de uno o más tipos de cromosomas, sin llegar a afectar al juego completo. Pueden ser nulisomías, monosomías, trisomías, tetraso-mías, etc., cuando en lugar de dos cromosomas de cada par no hay ninguno, o hay uno, tres, cuatro, etc. Un ejemplo de monosomía en humanos es el síndrome de Turner, en el que las mujeres, en lugar de tener dos cromosomas X, sólo tienen uno (mujeres X0); en total sólo tienen 45 cromosomas. Un ejemplo de trisomía en humanos es el síndrome de Down o trisomía del cromosoma 21, en el que los individuos, en vez de tener dos cromosomas tipo 21, tienen tres; en total tienen 47 cromosomas.
; en total sólo tienen 45 cromosomas. Un ejemplo de trisomía en humanos es el síndrome de Down o trisomía del cromosoma 21, en el que los individuos, en vez de tener dos cromosomas tipo 21, tienen tres; en total tienen 47 cromosomas.")
55
Euploidía. Es la alteración en el número normal de dotaciones haploides (juegos de cromosomas) de un individuo. Incluye dos tipos: La monoploidía o haploidía es la existencia de una sola dotación cromosómica, es decir, un solo ejemplar de cada tipo de cromosoma. La poliploidía es la existencia de más de dos ejemplares de cada tipo de cromosomas o, dicho de otro modo, de más de dos juegos completos de cromosomas. Pueden ser triploidías, tetraploidías, etcétera. Son frecuentes en algunas plantas y raras en los animales. Las formas poliploides tienen hojas y frutos de mayor tamaño, por lo que resultan interesantes para la producción agrícola. La unión de dos de estos gametos origina individuos 4n. El 47% de las angiospermas que consumimos son poliploides.

56
Las causas de las mutaciones genómicas son:
Fusión céntrica. Es la unión de dos cromosomas no homólogos, con pérdida del centrómero de uno de ellos. Ejemplo de fusión es el origen de las dotaciones genéticas de varias especies de Drosophila (fig. 9) y probablemente el origen del cromosoma 2 humano a partir de dos cromosomas de primate: en humanos n es 23, mientras que en chimpancés y orangutanes n es 24. Fisión céntrica. Es la escisión de un cromosoma en dos. Da lugar a un nuevo centrómero. Segregación errónea durante la meiosis. Es la distribución erró-nea de las cromátidas homólogas entre las células hijas durante la meiosis. A una célula irán las dos cromátidas y la otra se quedará sin ninguna.
 y probablemente el origen del cromosoma 2 humano a partir de dos cromosomas de primate: en humanos n es 23, mientras que en chimpancés y orangutanes n es 24. Fisión céntrica. Es la escisión de un cromosoma en dos. Da lugar a un nuevo centrómero. Segregación errónea durante la meiosis. Es la distribución erró-nea de las cromátidas homólogas entre las células hijas durante la meiosis. A una célula irán las dos cromátidas y la otra se quedará sin ninguna.")
57
Origen de los cromosomas de Drosophila
Origen de los cromosomas de Drosophila. Todas las especies de este género tienen un número de cromosomas muy bajo. El origen de las dotaciones cromosómicas puede deberse a la fusión de cromosomas a partir de un patrón original, la dotación cromosómica de Drosophila virilis.

Presentaciones similares







 con información.>")









. Cuando se cruzan dos individuos (P) de raza pura ambos (homocigotos ) para un.>")


