Descargar la presentación
La descarga está en progreso. Por favor, espere

1
CONSECUENCIAS COMUNITARIAS DE LA FRAGMENTACIÓN DEL HÁBITAT:
PATRONES, PROCESOS Y MECANISMOS

2
Fragmentación del hábitat
Efectos abióticos Efectos bióticos directos Efectos bióticos indirectos Biodiversidad (composicional, estructural y funcional) Modificado de Bustamante & Grez (1995)
 Modificado de Bustamante & Grez (1995)")
3
En esta clase, cómo la fragmentación del hábitat afecta:
1) Patrones de abundancia y riqueza de especies 2) Procesos (interacciones ecológicas): competencia, depredación (sensu lato), mutualismo, comensalismo 3) Mecanismos
 Patrones de abundancia y riqueza de especies. 2) Procesos (interacciones ecológicas): competencia, depredación (sensu lato), mutualismo, comensalismo. 3) Mecanismos.")
4
EFECTO DE LA FRAGMENTACION SOBRE ESTRUCTURA COMUNITARIA
PATRONES EFECTO DE LA FRAGMENTACION SOBRE ESTRUCTURA COMUNITARIA

5
Teoría de biogeografía de islas
EFECTO TAMAÑO Y DISTANCIA SOBRE RIQUEZA Teoría de biogeografía de islas (MacArthur & Wilson 1967)
")
6
Riqueza de especies disminuye en ambientes fragmentados y aislados
Gilbert et al. (1998) Local ( ) Global ( ) Riqueza de especies disminuye en ambientes fragmentados y aislados
 Local ( ) Global ( ) Riqueza de especies disminuye en ambientes fragmentados y aislados.")
7
Lo mismo ocurre en aves...pero depende...
Kattan (1996) Forman (1995)
 Forman (1995)")
8
Fragmentos Grandes (> 100 ha) Medianos (2 - 6 ha) Pequeños
En Chile (Chiloé) Willson et al. (1994) Fragmentos Grandes (> 100 ha) Medianos (2 - 6 ha) Pequeños (< 0,5 ha) Diversidad media 22,5 15,6 11,3 Abundancia 14,6 10,7 9,6
 Willson et al. (1994) Fragmentos. Grandes. (> 100 ha) Medianos. (2 - 6 ha) Pequeños. (< 0,5 ha) Diversidad. media. 22,5. 15,6. 11,3. Abundancia. 14,6. 10,7. 9,6.")
9
Decrecen Tienden a decrecer Incrementan Picaflor Torcaza Chincol
Chucao Hued-hued Torcaza Tienden a decrecer Fío-fío Picaflor Incrementan Chercán Chincol Zorzal

10
Mayoría de especies que declinaron son endémicas (6 de 7), usan nidos cubiertos (6 de 7) y son de sotobosque (5 de 7) Willson et al. (1994)
")
11
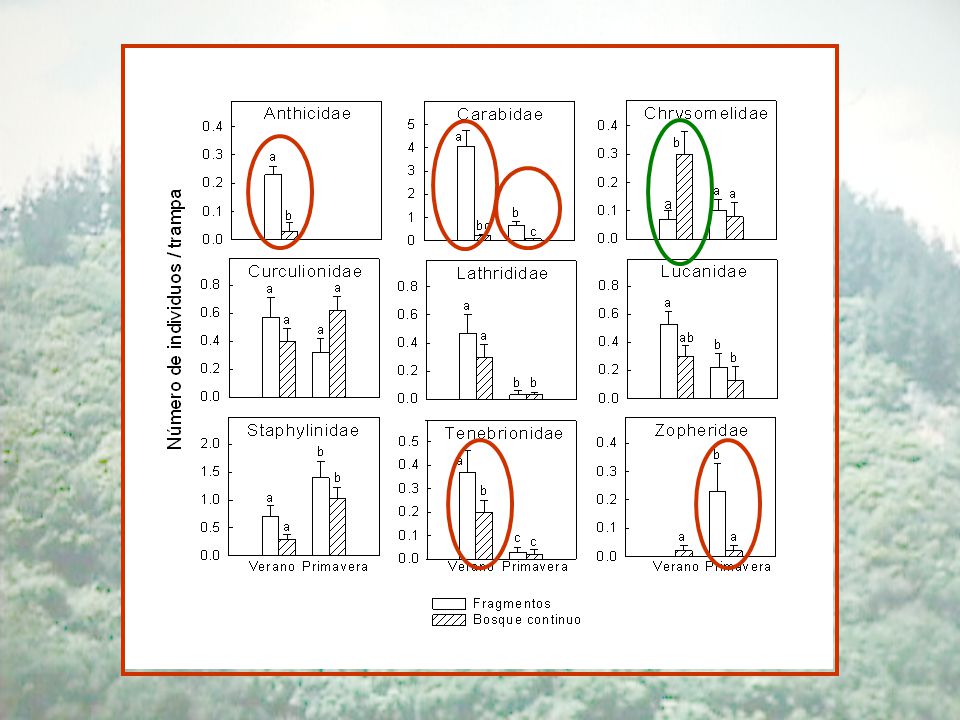
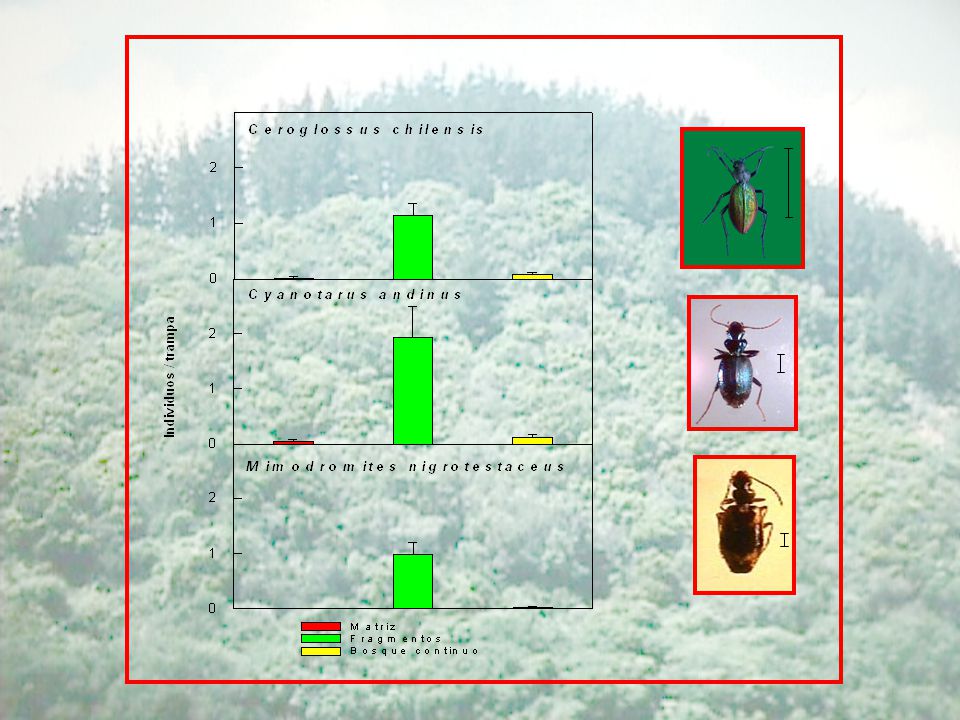
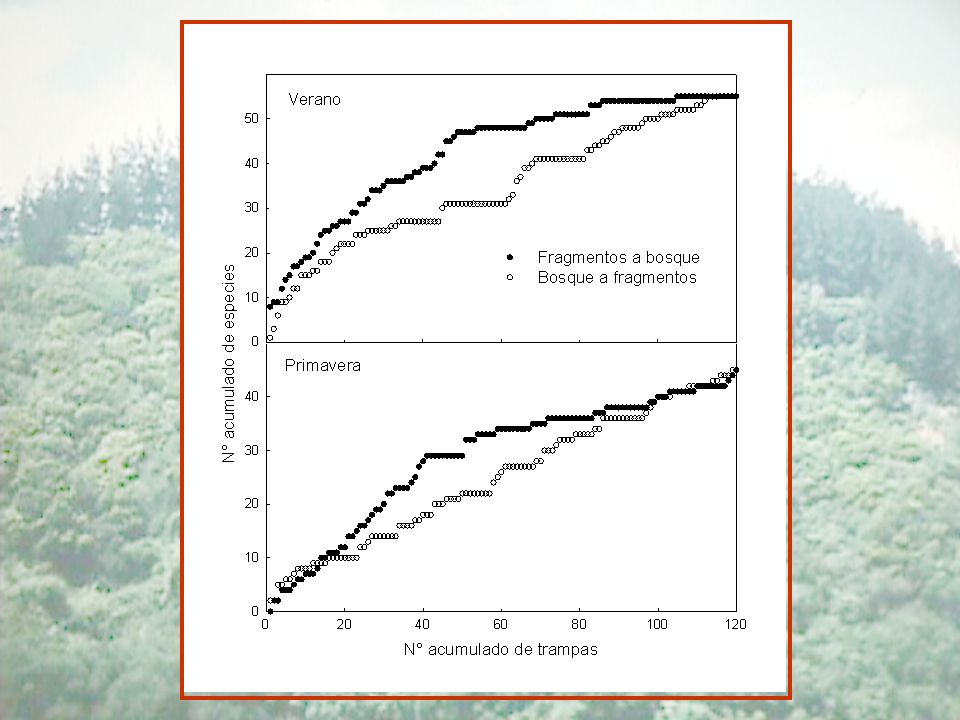
Coleópteros epigeos en el bosque Maulino
Enero Octubre Bosque continuo Fragmentos Abundancia (individuos/trampa) 2,3 0,3 a 2,4 3,3 0,4 Riqueza de especies (absoluta) 32 28 34 Riqueza de especies (especies/ trampa) 1,9 0,2 7,3 0,9 b 48 3,8 1,8 2,0 Coleópteros epigeos en el bosque Maulino
 2,3. 0,3. a. 2,4. 3,3. 0,4. Riqueza de especies (absoluta) Riqueza de especies (especies/ trampa) 1,9. 0,2. 7,3. 0,9. b ,8. 1,8. 2,0. Coleópteros epigeos en el bosque Maulino.")
15
Conclusiones La fragmentación del hábitat cambia la composición y estructura de la biota La riqueza de especies puede aumentar o disminuir Las respuestas son taxón-dependiente

16
EFECTO DE LA FRAGMENTACION DEL HÁBITAT SOBRE INTERACCIONES ECOLÓGICAS
PROCESOS EFECTO DE LA FRAGMENTACION DEL HÁBITAT SOBRE INTERACCIONES ECOLÓGICAS

17
Zuidema et al. (1996) (N = 58)
 (N = 58)")
18
Competencia en ambientes fragmentados
- Pocos trabajos empíricos - Aumento de bordes: invasiones biológicas (Janzen 1983) - Modelos teóricos (sin recurrir a efecto borde)
 - Modelos teóricos (sin recurrir a efecto borde)")
19
Modelo metapoblacional de Nee & May (1992)
Dos especies: A (competidor superior) y B (competidor inferior) cA yz cA yx eA y cB zx eB z Parches con competidor superior (A) Parches vacíos inferior (B) z y x Supuestos: 1. B no puede colonizar a parches y (ocupados por especie A) 2. si A invade a parches z (ocupados por b) B se extingue 3. B puede coexistir con A a nivel de paisaje por mayor tasa de colonización o menor tasa de extinción o ambas
 y B (competidor inferior) cA yz. cA yx. eA y. cB zx. eB z. Parches con. competidor. superior (A) Parches vacíos. inferior (B) z. y. x. Supuestos: 1. B no puede colonizar a parches y (ocupados por especie A) 2. si A invade a parches z (ocupados por b) B se extingue. 3. B puede coexistir con A a nivel de paisaje por mayor tasa de colonización o menor tasa de extinción o ambas.")
20
Parches vacíos (x): dx/dt = -cAyx + eAy - cBzx + eBz
cA yz cA yx eA y cB zx eB z Parches con competidor superior (A) Parches vacíos inferior (B) z y x Parches vacíos (x): dx/dt = -cAyx + eAy - cBzx + eBz Parches con A (y): dy/dt = cAyx - eAy + cAyz Parches con B (z): dz/dt = cBzx - eBz - cAyz Parches habitables h = x + y + z
 Parches vacíos. inferior (B) z. y. x. Parches vacíos (x): dx/dt = -cAyx + eAy - cBzx + eBz. Parches con A (y): dy/dt = cAyx - eAy + cAyz. Parches con B (z): dz/dt = cBzx - eBz - cAyz. Parches habitables h = x + y + z.")
21
si h disminuye x* disminuye (disminuyen parches vacíos,
no destruídos) y* disminuye (disminuyen parches con A) z* aumenta (aumentan parches con B) para que B persista: cB/eB > cA/eA En equilibrio: x* = (1/cB) (hcA - eA + eB) y* = h - (eA/cA) z* = (eA (cA + cB)/cA cB) - eB/cB - hcA/cB
 y* disminuye (disminuyen parches con A) z* aumenta (aumentan parches con B) para que B persista: cB/eB > cA/eA. En equilibrio: x* = (1/cB) (hcA - eA + eB) y* = h - (eA/cA) z* = (eA (cA + cB)/cA cB) - eB/cB - hcA/cB.")
22
0,5 > 1-h > 0,75 sólo existe B, liberación competitiva
0 > 1-h > 0,5 A y B coexisten en proporciones dadas por ecuaciones 0,5 > 1-h > 0,75 sólo existe B, liberación competitiva 1-h > 0,75 desaparece también B (todos los parches quedan vacíos) A B
 A. B.")
23
En ausencia de remoción de parches: i) A aumenta directamente la extinción de B (al colonizar sus parches) y ii) disminuye el número de parches disponibles para que B colonice. Asi, la remoción de parches tiene un efecto indirecto positivo sobre la especie competidora inferior B. Competidor superior A inferior B Remoción de parches +

24
Conclusiones de Nee & May (1992)
1. Resultado contraintuitivo: a medida que se remueven parches A disminuye y B aumenta. 2. No es necesario recurrir a efectos de bordes sino a la remoción de hábitat per se para el éxito de especies competitivamente inferiores. 3. Remoción de parches aumenta la biodiversidad.

25
Modelo celular de Dytham (1994)
B vacíos 0.2 - Cuando h < 0.2 (80% de destrucción) B ocupa más parches que A. En Nee & May esto ocurre antes) - A y B presentes hasta el 96% de remoción de parches (en Nee & May sólo coexisten hasta el 50% de remoción) - Al igual que Nee & May, destrucción del hábitat aumenta la diversidad
 B ocupa más parches que A. En Nee & May esto ocurre antes) - A y B presentes hasta el 96% de remoción de parches (en Nee & May sólo coexisten hasta el 50% de remoción) - Al igual que Nee & May, destrucción del hábitat aumenta la diversidad.")
26
Modelo celular de Tilman et al. (1994)
A mayor destrucción mayor pérdida de especies (desde A hasta B) Modelo celular de Tilman et al. (1994)
 Modelo celular de Tilman et al. (1994)")
27
Más especies se extinguirán cuando el competidor dominante es raro (q = 0.05, trópico) que abundante (q = 0.5, templado)
 que abundante (q = 0.5, templado)")
28
A medida que más hábitat ya ha sido destruído mayor es el efecto de la fragmentación adicional sobre la extinción. Debido a que las extinciones se producen con un tiempo de retardo, existe una deuda: un costo futuro de la actual destrucción del hábitat.

29
Conclusiones generales
: 1. favorece a especies con baja capacidad competitiva (usualmente las con mayor capacidad de dispersión). 2. Esto, unido al efecto borde, implica que la fragmentación del hábitat aumenta la biodiversidad (mayor coexistencia de especies). Pero de que tipo? 3. Se requiere poner a prueba estos modelos (Gilbert et al. 1998) Los tres modelos coinciden en que la fragmentación del hábitat
. 2. Esto, unido al efecto borde, implica que la fragmentación del. hábitat aumenta la biodiversidad (mayor coexistencia de. especies). Pero de que tipo 3. Se requiere poner a prueba estos modelos (Gilbert et al. 1998) Los tres modelos coinciden en que la fragmentación del hábitat.")
30
Depredación en ambientes fragmentados

31
Modelo metapoblacional de May (1994)
Dos especies: v (presa) y p (depredador) Parches con depredadores y presas (p, v) cp zy cv yx ev y ep z presas (v) Parches vacíos z y x Supuestos: 1) p pueden colonizar sólo los parches que tienen presas. 2) v en paches con depredadores y presas (z) no colonizan parches vacíos. 3) Parches con p y v se extinguen a una tasa ep 4) v y p pueden coexistir como metapoblaciones en el paisaje mientras dinámicas sean asincrónicas.
 y p (depredador) Parches con. depredadores y presas (p, v) cp zy. cv yx. ev y. ep z. presas (v) Parches vacíos. z. y. x. Supuestos: 1) p pueden colonizar sólo los. parches que tienen presas. 2) v en paches con depredadores y. presas (z) no colonizan parches. vacíos. 3) Parches con p y v se extinguen a una tasa ep. 4) v y p pueden coexistir como metapoblaciones en el paisaje. mientras dinámicas sean asincrónicas.")
32
Parches vacíos (x): dx/dt = evy + epz - cvyx
Parches con depredadores y presas (p, v) cp zy cv yx ev y ep z presas (v) Parches vacíos z y x Parches vacíos (x): dx/dt = evy + epz - cvyx Parches con v (y): dy/dt = cvyx - cpzy - evy Parches con p y v (z): dz/dt = cpzy - epz
 cp zy. cv yx. ev y. ep z. presas (v) Parches vacíos. z. y. x. Parches vacíos (x): dx/dt = evy + epz - cvyx. Parches con v (y): dy/dt = cvyx - cpzy - evy. Parches con p y v (z): dz/dt = cpzy - epz.")
33
x* = h - y* - z* y* = ep/cp z* = (cv/(cv + cp))(h - (ep/cp) - (ev/cv)) 1. La destrucción del hábitat no tiene un efecto en el número en equilibrio de parches de presas (y) hasta llegar a un nivel en que se extinguen los depredadores. 2. La abundancia de depredadores declina a mayor destrucción de hábitat, aún cuando no hay cambios en la abundancia de presas.
 hasta llegar a un nivel en que se extinguen los depredadores. 2. La abundancia de depredadores declina a mayor destrucción de hábitat, aún cuando no hay cambios en la abundancia de presas.")
34
TRABAJOS EMPÍRICOS DE RELACIÓN DEPREDADOR/PRESA EN AMBIENTES FRAGMENTADOS

35
Huffaker (1958) Refugios espaciales (temporales)
 Refugios espaciales (temporales)")
36
Janssen et al. (1997), tomado de McCauley et al. (2000)
Super-isla Metapoblaciones Persistencia por más de 30 generaciones Inestabilidad

37
Refugios espaciales más permanentes para las presas (escape).
Kareiva (1987) Refugios espaciales más permanentes para las presas (escape). Fragmentación desestabiliza relación depredador-presa.
 Refugios espaciales más permanentes para las presas. (escape). Fragmentación desestabiliza relación depredador-presa.")
38
Herbívoros escapan de enemigos naturales en los bordes
Roland (1993) Malacosoma distria Populus tremuloides
 Malacosoma distria. Populus tremuloides.")
39
A nivel de mosaicos, la fragmentación puede estabilizar o desestabilizar la relación depredador-presa dependiendo de cómo la dispersión y colonización de interactuantes sean afectadas

40
DEPREDACIÓN DE HUEVOS

41
DEPREDACIÓN DE HUEVOS

42
DEPREDACIÓN DE HUEVOS Angelstam (1988)
")
43
EFECTO BORDE SEGÚN USO DEL HÁBITAT
Andrén (1995)
")
44
EFECTO BORDE SEGÚN CONTRASTE CON MATRIZ
Angelstam (1986)
")
45
DEPREDACIÓN SEGÚN CONTRASTE CON MATRIZ
Andrén (1995) DEPREDACIÓN SEGÚN CONTRASTE CON MATRIZ
 DEPREDACIÓN SEGÚN CONTRASTE CON MATRIZ.")
46
Chalfoun et al. (2002) La respuesta de los depredadores varía de acuerdo al tipo de paisaje: mayor cuando el paisaje adyacente es agrícola que cuando es forestal. 41,2% 12,9%

47
EFECTO DE LA FRAGMENTACIÓN DEL HÁBITAT SOBRE LA GRANIVORÍA

48
Burkey (1993)
")
49
¡¡En bosque Maulino ocurre algo similar!!
Tellería et al. (1991) ¡¡En bosque Maulino ocurre algo similar!! (Donoso et al. 2003)
 ¡¡En bosque Maulino ocurre algo similar!! (Donoso et al. 2003)")
50
CONSECUENCIAS COMUNITARIAS DE CAMBIOS EN LA GRANIVORÍA
(Leigh et al. 1993) ISLAS EN LAGO GATUN, PANAMÁ CONSECUENCIAS COMUNITARIAS DE CAMBIOS EN LA GRANIVORÍA (+)
 ISLAS EN LAGO GATUN, PANAMÁ. CONSECUENCIAS COMUNITARIAS DE CAMBIOS EN LA GRANIVORÍA. (+)")
51
(Leigh et al. 1993) Islas más chicas
 Islas más chicas")
52
(Leigh et al. 1993) Ausencia de granívoros en islas pequeñas modificaría las interacciones competitivas entre las especies de plantas, alterando la estructura comunitaria

53
EFECTO DE LA FRAGMENTACIÓN DEL HÁBITAT SOBRE LA FOLIVORÍA

54
Benites-Malvido et al. (1999)
En bosques Amazónicos (Manaos) herbivoría de plántulas es menor en fragmentos pequeños, pero esta diferencia desaparece con el tiempo. Pouteria caimito
 herbivoría de plántulas es menor en fragmentos pequeños, pero esta diferencia desaparece con el tiempo. Pouteria caimito.")
55
Herbivoría en plántulas de peumo en Bosque Maulino
F= 5,04; gl= 1; P= 0,02 * F= 5,43; gl= 1; P= 0,02 Herbivoría en plántulas de peumo en Bosque Maulino Vega (2001)
")
56
Herbivoría en maqui Vásquez (2003)
")
57
EFECTO DE LA FRAGMENTACIÓN SOBRE POLINIZACIÓN

58
Aizen & Feinsinger (1994) Polinización
 Polinización")
59
Aizen & Feinsinger (1994)
")
61
Palaemonella sp. Trapezia cymodoce Caley et al. (2001) Comensalismo
 Comensalismo")
62
Mecanismos (no excluyentes) mediante los cuales la fragmentación (bordes) altera las interacciones ecológicas (Fagan et al. 1999) a) Bordes afectan diferencialmente el movimiento de organismos, facilitando o restringiendo la dispersión (permeabilidad). Dispersión de semillas - interacciones competitivas entre plantas - composición vegetal (Janzen 1983) Dispersión de depredadores - cambios en estabilización de relación D/P (Kareiva 1987) Dispersión de polinizadores - polinización - tamaño reproductivo - cambios en composición vegetal (Aizen & Feinsinger, 1994)
 Bordes afectan diferencialmente el movimiento de organismos, facilitando o restringiendo la dispersión (permeabilidad). Dispersión de semillas - interacciones competitivas entre plantas - composición vegetal (Janzen 1983) Dispersión de depredadores - cambios en estabilización de relación D/P (Kareiva 1987) Dispersión de polinizadores - polinización - tamaño reproductivo - cambios en composición vegetal (Aizen & Feinsinger, 1994)")
63
b) Bordes afectan diferencialmente la mortalidad.
Cambios microclimáticos afectan sobrevivencia u otros procesos demográficos de semillas o plántulas (e.g., germinación): - reclutamiento - cambios en interacciones competitivas. Consumo preferencial por depredadores en bordes - aumenta mortalidad de presas (o viceversa, e.g., Roland 1993)
: - reclutamiento - cambios en interacciones competitivas. Consumo preferencial por depredadores en bordes - aumenta mortalidad de presas (o viceversa, e.g., Roland 1993)")
64
c) “Subsidios” a través de los bordes
Poblaciones de algunas especies mantienen altos niveles de abundancia al reproducirse o alimentarse en otras áreas, pero al dispersarse a través de los bordes deprimen poblaciones residentes (e.g., especies malezas). “Hipótesis de la trampa ecológica” (Gates & Gysel 1972): aves prefieren bordes como hábitat para anidar aún cuando allí los depredadores generalistas de hábitat (i.e., subsidios) las depredan más.
. Hipótesis de la trampa ecológica (Gates & Gysel 1972): aves prefieren bordes como hábitat para anidar aún cuando allí los depredadores generalistas de hábitat (i.e., subsidios) las depredan más.")
65
d) Bordes como hábitat para nuevas interacciones
Cambios estructurales en los bordes generan efectos en cascada, apareciendo allí nuevas interacciones. Bordes pueden transformarse en hábitat sumidero para ciertas especies o viceversa

66
Noss (1990)
")
67
Desigualdad en estudios de diferentes interacciones
Variabilidad enorme de las respuestas Para buscar generalidades se requiere conocer más de la historia natural de los organismos y distinguir los atributos de historia de vida (e.g., dispersión, uso del hábitat) que los hacen más susceptibles a la fragmentación
 que los hacen más susceptibles a la fragmentación.")
Presentaciones similares







 Problemas que afectan la biodiversidad>")

>")










