Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Tema 8. Transcripción

2
Tema 8. Transcripción • dogma central de la biología molecular
• síntesis de RNA: RNA polimerasa 5’->3’, complementaria, antiparalela la transcripción es asimétrica • selección espacio, tiempo y cantidad: promotores (fuertes y débiles) y factores sigma • nomenclatura de la síntesis de RNA • el proceso de transcripción: iniciación, elongación y terminación (terminadores intrínsecos o dependientes de rho) • principales tipos de RNA: mRNA, rRNA, tRNA y los RNA ‘pequeños’
 y factores sigma. • nomenclatura de la síntesis de RNA. • el proceso de transcripción: iniciación, elongación y terminación (terminadores intrínsecos o dependientes de rho) • principales tipos de RNA: mRNA, rRNA, tRNA y los RNA ‘pequeños’")
3
dogma central de la biología molecular Crick, 1958
Figure: 13-01 How Genetic Information Produces Protein Flow chart illustrating how genetic information encoded in DNA produces protein. Why would the cell want to have an intermediate between DNA and the proteins it encodes? * The DNA can then stay pristine and protected, away from the caustic chemistry of the cytoplasm. * Gene information can be amplified by having many copies of an RNA made from one copy of DNA. * Regulation of gene expression can be effected by having specific controls at each element of the pathway between DNA and proteins. The more elements there are in the pathway, the more opportunities there are to control it in different circumstances.

5
DNA en el núcleo y síntesis de proteínas en citoplasma
los tres procesos de flujo de la información genética: replicación, transcripción y traducción DNA: núcleo y síntesis proteínas en citoplasma -> intermediario: RNA DNA en el núcleo y síntesis de proteínas en citoplasma RNA sintetizado en el núcleo RNA emigra al citoplasma Cantidad de RNA proporcional a la de proteínas

6
experimento de ‘pulso y caza’ con precursores radiactivos de RNA
resultado: el RNA se sintetiza en el núcleo y luego se exporta al citoplasma

11
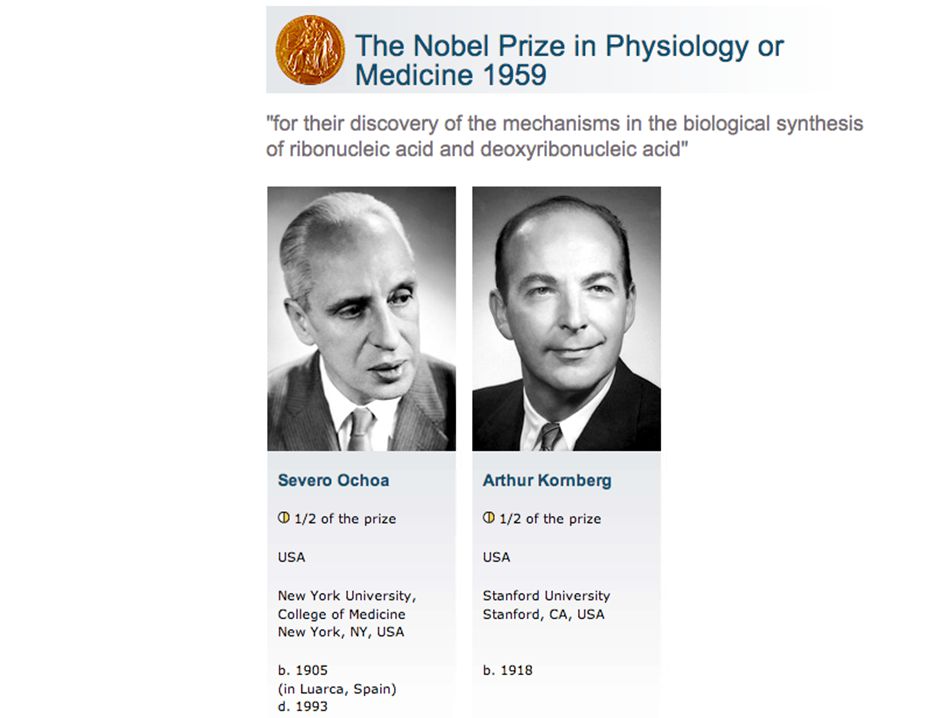
Severo Ochoa, 1955 (Premio Nobel, 1959)
In 1955, Marianne Grunberg-Manago and Severo Ochoa reported the isolation of an enzyme that catalyzed the synthesis of RNA. For this work, Ochoa shared the 1959 Nobel Prize in Medicine with Arthur Kornberg (who received the Prize for his work on DNA polymerase I). This enzyme could convert ribonucleoside diphosphates into RNA: (NMP)n + NDP --> (NMP)n+1 + Pi However, the enzyme had a number of unsettling properties. It did not need a template; and, it could use as little as 1 NDP or as many as 4 NDPs as substrate. In fact, the sequence of the product RNA depended entirely on the number and concentration of substrate NDPs. These are not the properties of an enzyme that must faithfully copy the genetic material for expression! We now know that Grunberg-Manago and Ochoa had isolated the enzyme polynucleotide phosphorylase which usually catalyzes the breakdown of RNA - not its synthesis!
. This enzyme could convert ribonucleoside diphosphates into RNA: (NMP)n + NDP --> (NMP)n+1 + Pi. However, the enzyme had a number of unsettling properties. It did not need a template; and, it could use as little as 1 NDP or as many as 4 NDPs as substrate. In fact, the sequence of the product RNA depended entirely on the number and concentration of substrate NDPs. These are not the properties of an enzyme that must faithfully copy the genetic material for expression! We now know that Grunberg-Manago and Ochoa had isolated the enzyme polynucleotide phosphorylase which usually catalyzes the breakdown of RNA - not its synthesis!")
12
- ¡requiere un molde de DNA! - pero NO cebador
+ Mg2+ la RNA polimerasa: - ¡requiere un molde de DNA! - pero NO cebador incorporación secuencial, uno a uno, de ribonucleósidos trifosfatos a una cadena en crecimiento por la RNA polimerasa dirección 5’->3’ y manteniendo antiparalelismo y complementariedad con la cadena molde la RNA polimerasa NO necesita cebador! In 1960, the true enzyme was identified by 4 separate groups: Sam Weiss at the University of Chicago, Jerard Hurwitz, A. Stevens and J. Bonner. This enzyme required a template, used all four rNTPs as substrates and synthesized a product with a composition similar to that of the template, and it required Mg++. DNA-directed RNA polymerase catalyzes the synthesis of RNA on a DNA template following similar nucleophilic attack chemistry as DNA polymerase. For a long time, E. coli was thought to contain just a single RNA polymerase. We now know that, although it has one major or principal RNA polymerase, it also contains (at least) six minor forms that are required or are used in special circumstances.
 six minor forms that are required or are used in special circumstances.")
13
RNA polimerasa de E. coli
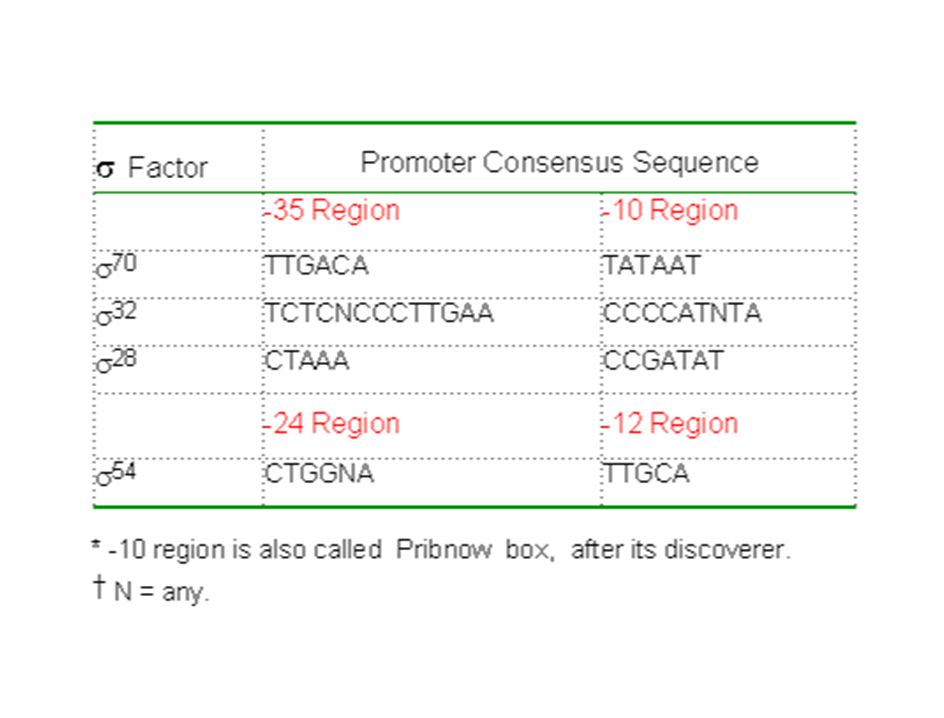
Classes of RNA polymerases E. coli An E. coli RNA polymerase is composed of five subunits: two a subunits, and one for each b, b', and s subunit. b (151 kD) and b' (156 kD) are significantly larger than a (37 kD). Several different forms of s subunits have been identified, with molecular weights ranging from 28 kD to 70 kD. The s subunit is also known as the s factor. It plays an important role in recognizing the transcriptional initiation site, and also possesses the helicase activity to unwind the DNA double helix. Nucleotide synthesis is carried out by other four subunits, which together are called the core polymerase. The term "holoenzyme" refers to a complete and fully functional enzyme. In this case, the holoenzyme includes the core polymerase and the s factor.
 and b (156 kD) are significantly larger than a (37 kD). Several different forms of s subunits have been identified, with molecular weights ranging from 28 kD to 70 kD. The s subunit is also known as the s factor. It plays an important role in recognizing the transcriptional initiation site, and also possesses the helicase activity to unwind the DNA double helix. Nucleotide synthesis is carried out by other four subunits, which together are called the core polymerase. The term holoenzyme refers to a complete and fully functional enzyme. In this case, the holoenzyme includes the core polymerase and the s factor.")
15
síntesis de RNA: 5’->3’ complementaria antiparalela
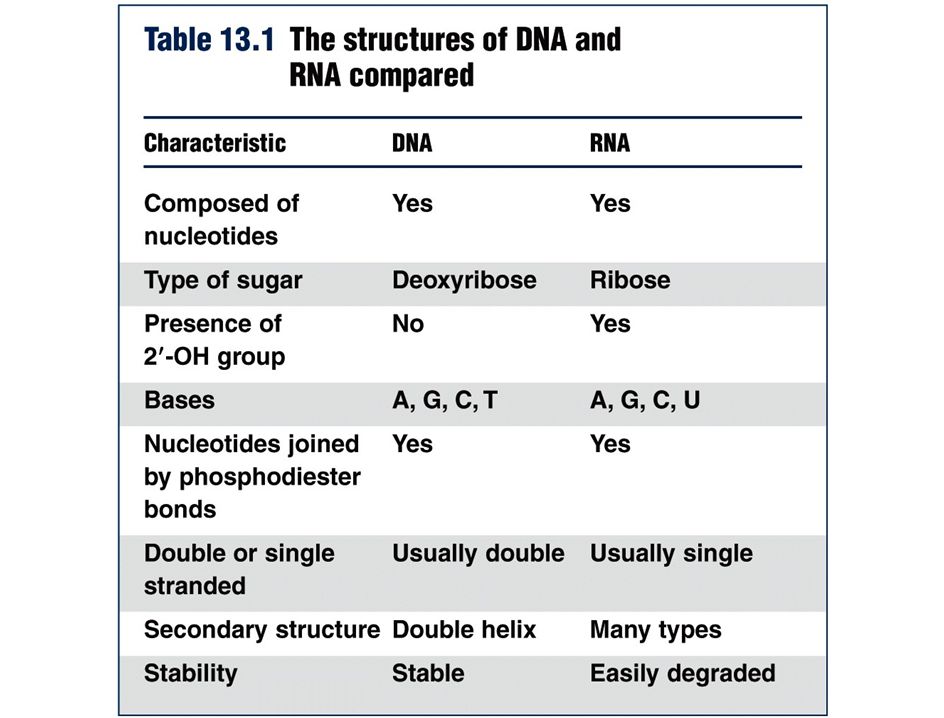
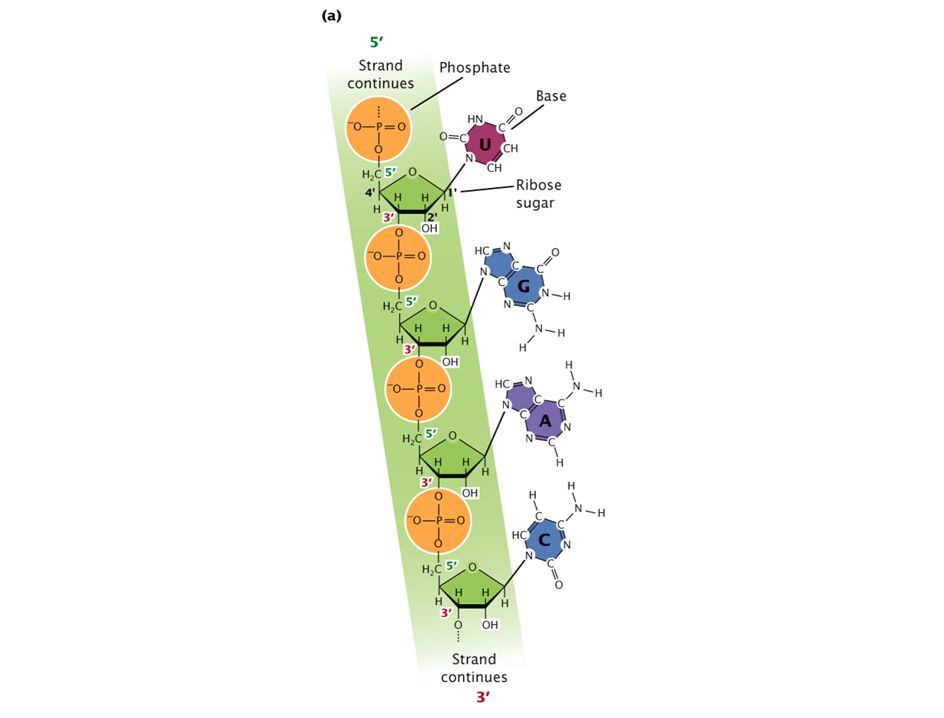
polimerización de RNA en la dirección 5’ -> 3’, siendo antiparalela a la cadena molde de DNA y manteniendo la complementariedad (ojo! en RNA A=U)
")
16
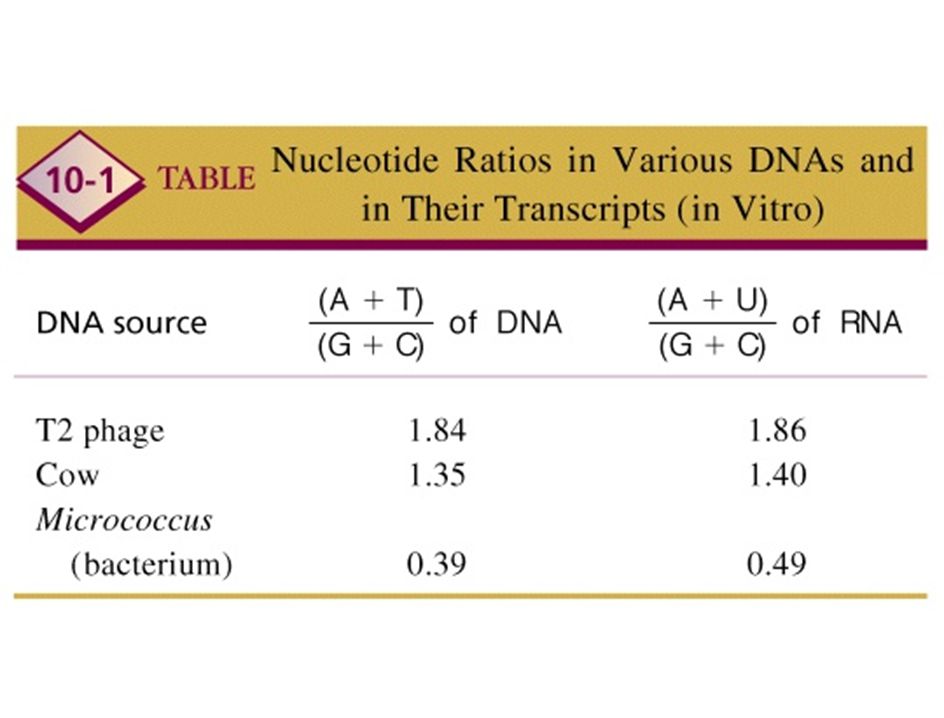
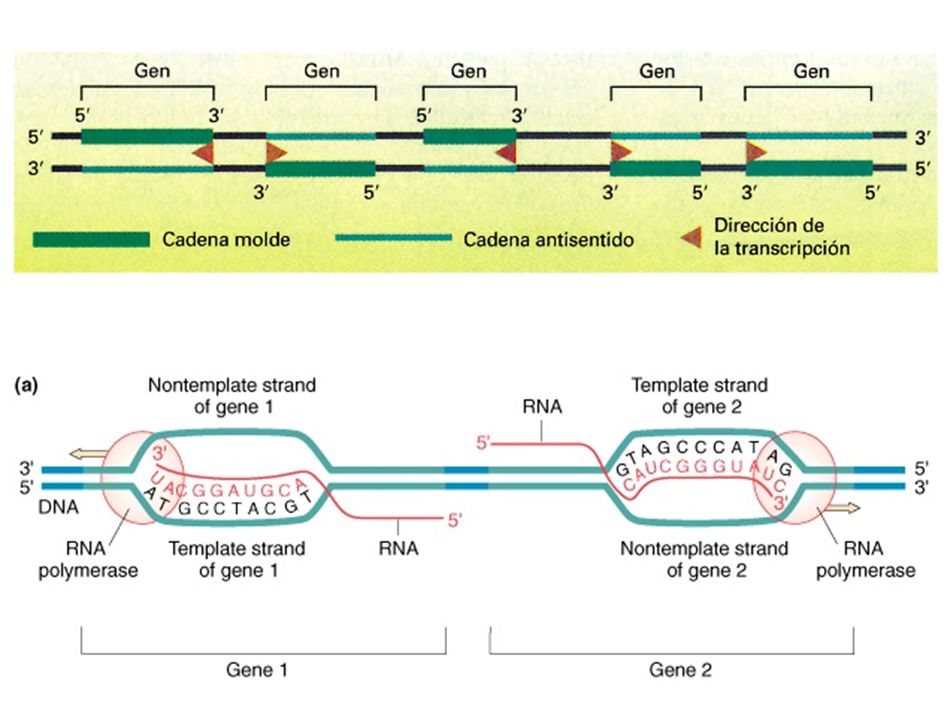
la transcripción es asimétrica
la transcripción es asimetrica experimentos de hibridación DNA-RNA

18
transcripción: selección espacio, tiempo y cantidad

19
promotores espacio secuencia Pribnow

20
tiempo factores sigma

22
cantidad promotores fuertes: 1 inicio transcripción / seg
promotores débiles: 1 inicio transcripción / min-h

23
nomenclatura síntesis RNA
cadenas DNA: complementaria molde molde = con sentido = codificante complementaria = no molde = anti-sentido = no codificante = acompañante por convención: mostrar cadena complementaria (idem secuencia RNA) 5’-P-transcrito-3’ gen de la Beta-galactosa
 5’-P-transcrito-3’ gen de la Beta-galactosa.")
24
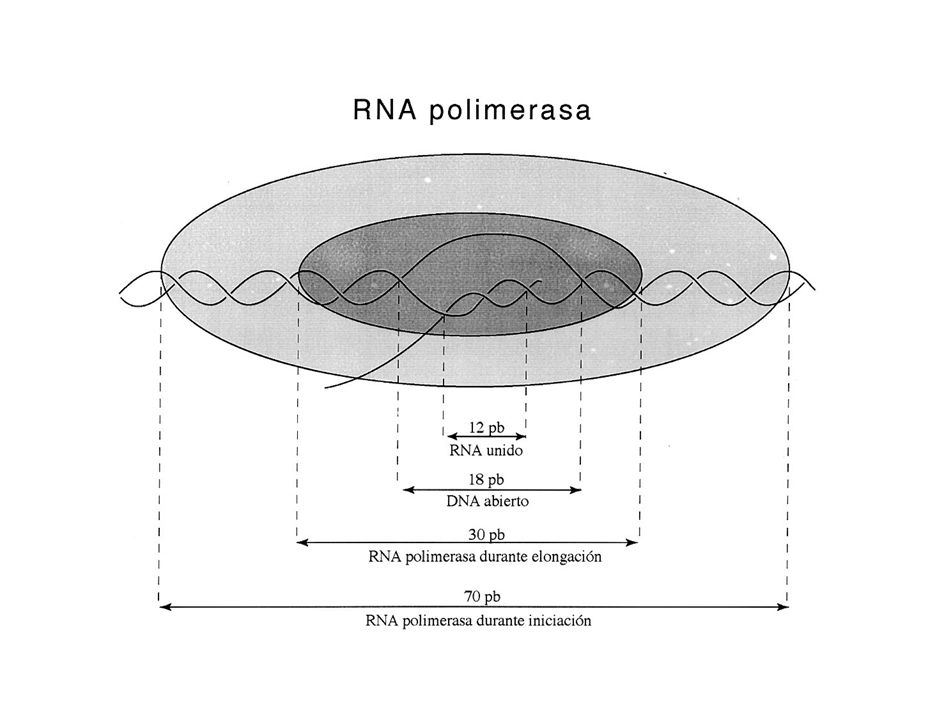
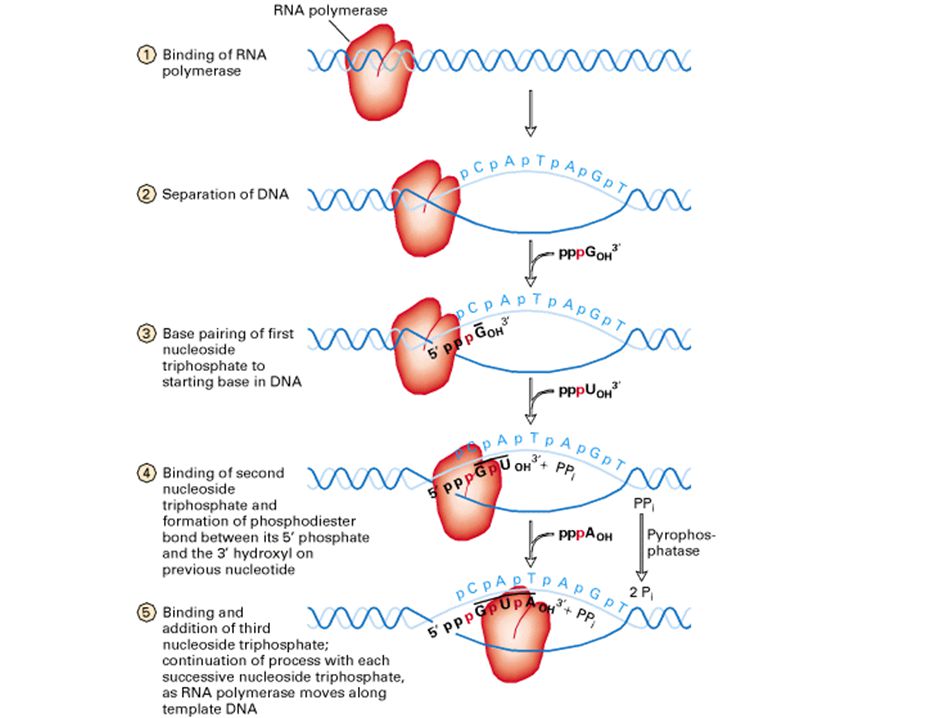
la transcripción: proceso
Figure: 13-08a The Early Stages of Transcription in Prokaryotes The early stages of transcription in prokaryotes, showing the components of the process.

25
iniciación de la transcripción

26
elongación de la transcripción

29
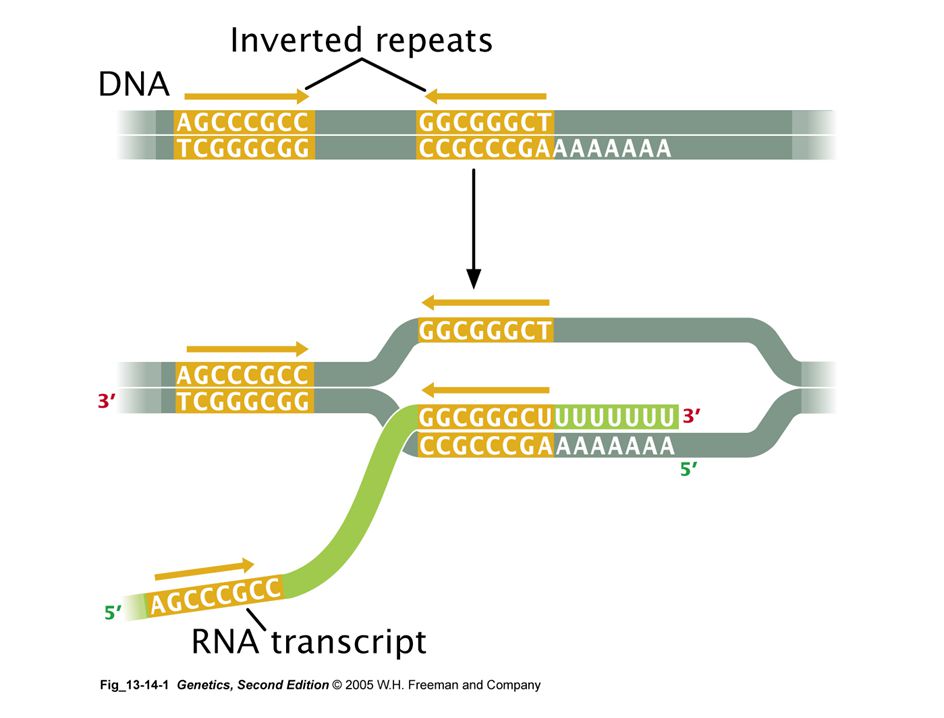
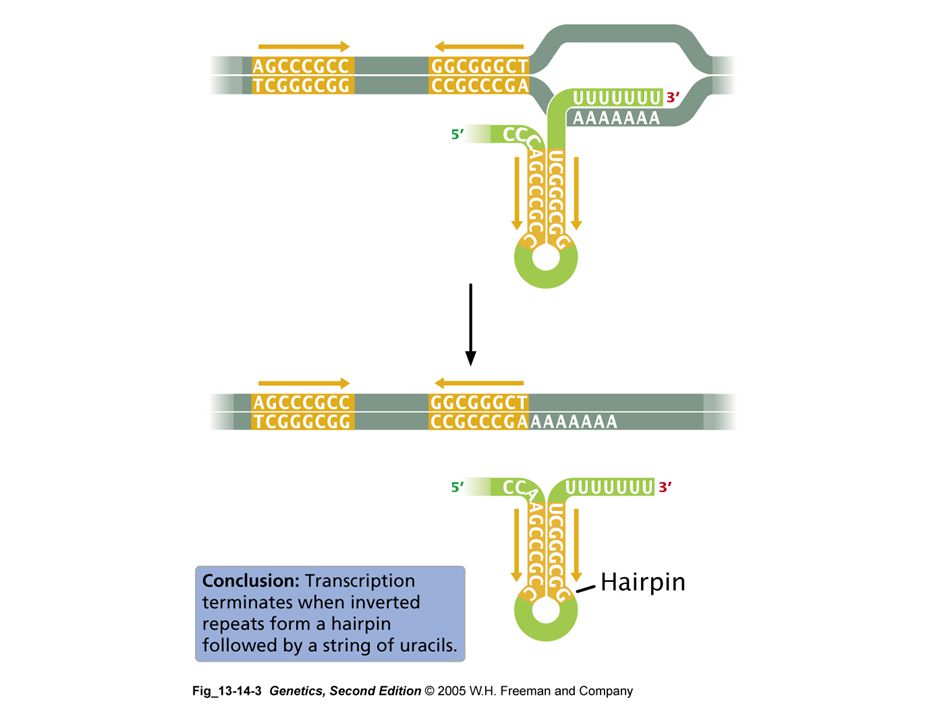
terminación de la transcripción
terminadores intrínsecos

32
terminadores dependientes de rho
Rho-dependent termination of transcription in E. coli RNA polymerase is stalled at a pause site, where rho binds to newly synthesized RNA. This binding is accompanied by ATP hydrolysis. Rho probably wraps the nascent RNA chain around itself, thereby destabilizing the RNA-DNA hybrid and terminating transcription. [Adapted from Platt, T. (1986). Transcription termination and the regulation of gene expression. Annu. Rev. Biochem. 55:339®¢372.] Notes Rho cannot bind to the single stranded RNA where ribosomes are bound but does in the region after the translation stop codon, where no ribosomes are present.Rho independent termination may use pause structures such as hairpins. 21.10
. Transcription termination and the regulation of gene expression. Annu. Rev. Biochem. 55:339®¢372.] Notes. Rho cannot bind to the single stranded RNA where ribosomes are bound but does in the region after the translation stop codon, where no ribosomes are present.Rho independent termination may use pause structures such as hairpins")
33
RNA polimerasa no cebador y sí molde! tamaño aprox: 500 KDa
polimerización: 43 n/seg y estable (hasta h) error: 1/ n no cebador y sí molde! tamaño aprox: 500 KDa RNA polimerasa
 error: 1/ n. no cebador y sí molde! tamaño aprox: 500 KDa. RNA polimerasa.")
34
RNA polimerasa - requiere un molde de DNA - pero no requiere cebador No todos los genes se expresan por igual - rRNA = 50% del RNA celular - mRNA = 50% del RNA celular - genes de rRNA = 0,32% del DNA - genes de mRNA = 99,68% del DNA 0,0032 x Activ. = 0,9968 Actividad = 311 veces más activos Frecuencia de fabricación - 1/seg : rRNA - 1/ciclo (1h) : ftsZ
 : ftsZ.")
35
demostración citológica de la transcripción
Transcripción en el nucléolo de Triturus viridiscens (tritón, un anfibio) de los genes repetidos en tándem que determinan el rRNA (aspecto "arbol de Navidad"). nucléolo de Triturus viridiscens
 de los genes. repetidos en tándem que determinan el rRNA (aspecto arbol de Navidad ). nucléolo de Triturus viridiscens.")
36
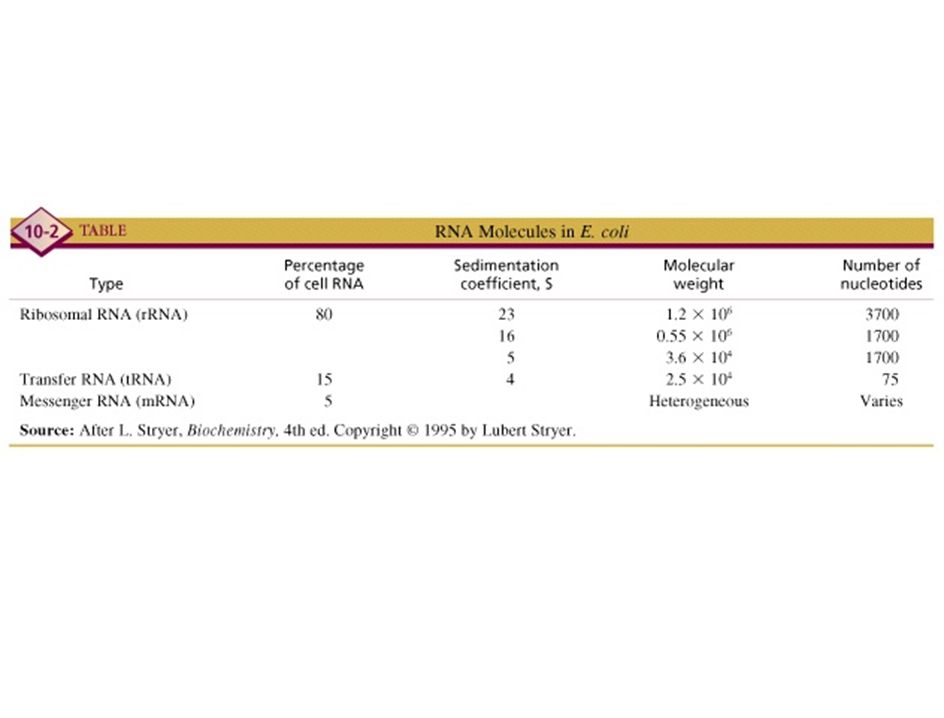
mRNA

37
procariotas eucariotas

38
rRNA Figure: 14-01 A Comparison of the Components in Prokaryotic and Eukaryotic Ribosomes A comparison of the components in prokaryotic and eukaryotic ribosomes.

39
operón rrn de E. coli (rDNA)
5’ 3’ 16S 23S 5S tRNA operón rrn de E. coli (rDNA) Transcription of E. coli ribosomal RNA genes The genes are being transcribed from left to right. The RNA product associates with proteins and is processed by lucleolytic cleavage before transcription is complete. Notes In this micrograph it can be seen that many transcripts are generated from the same E. coli gene at the same time. 1.1 How are rRNA genes organized on the prokaryotic genome? Clustering of rRNA genes into operons is common among prokaryotes and permits transcription of equimolar quantities of each rRNA gene. However, alternative organizations of rRNA genes are observed in some prokaryotes indicating that organization into operons is not a strict requirement. Genes encoding transfer RNAs (tRNAs) are often located in the internally transcribed spacer (ITS) region and distal to the 5S rRNA gene. The ITS region is variable in length and contributes to the majority of sequence diversity between multiple rRNA operons in many species of bacteria. The rRNA operon is transcribed as a single RNA molecule and subsequently cleaved by ribonucleases into component rRNA and tRNA molecules. Organization of rRNA operons around the origin of replication is a common structural feature among phylogenetically diverse bacteria. Localization of rRNA operons near the origin of replication provides a gene dosage effect during rapid growth where the effective number of rRNA operons in E.coli can be as high as 36 copies during exponential growth with multiple replications forks.
 Transcription of E. coli ribosomal RNA genes The genes are being transcribed from left to right. The RNA product associates with proteins and is processed by lucleolytic cleavage before transcription is complete. Notes In this micrograph it can be seen that many transcripts are generated from the same E. coli gene at the same time. 1.1 How are rRNA genes organized on the prokaryotic genome Clustering of rRNA genes into operons is common among prokaryotes and permits transcription of equimolar quantities of each rRNA gene. However, alternative organizations of rRNA genes are observed in some prokaryotes indicating that organization into operons is not a strict requirement. Genes encoding transfer RNAs (tRNAs) are often located in the internally transcribed spacer (ITS) region and distal to the 5S rRNA gene. The ITS region is variable in length and contributes to the majority of sequence diversity between multiple rRNA operons in many species of bacteria. The rRNA operon is transcribed as a single RNA molecule and subsequently cleaved by ribonucleases into component rRNA and tRNA molecules. Organization of rRNA operons around the origin of replication is a common structural feature among phylogenetically diverse bacteria. Localization of rRNA operons near the origin of replication provides a gene dosage effect during rapid growth where the effective number of rRNA operons in E.coli can be as high as 36 copies during exponential growth with multiple replications forks.")
40
organizador nucleolar en Drosophila
Suzuki 3-43 eu- y heterocromatina en Drosophila eucromatina: amarillo; heterocromatina: azul C: centrómero; NO: organizador nucleolar

41
tRNA

42
Figure: 14-02 Ribonucleotides Ribonucleotides containing unusual nitrogenous bases found in transfer RNA.

43
tRNA estructura terciaria

45
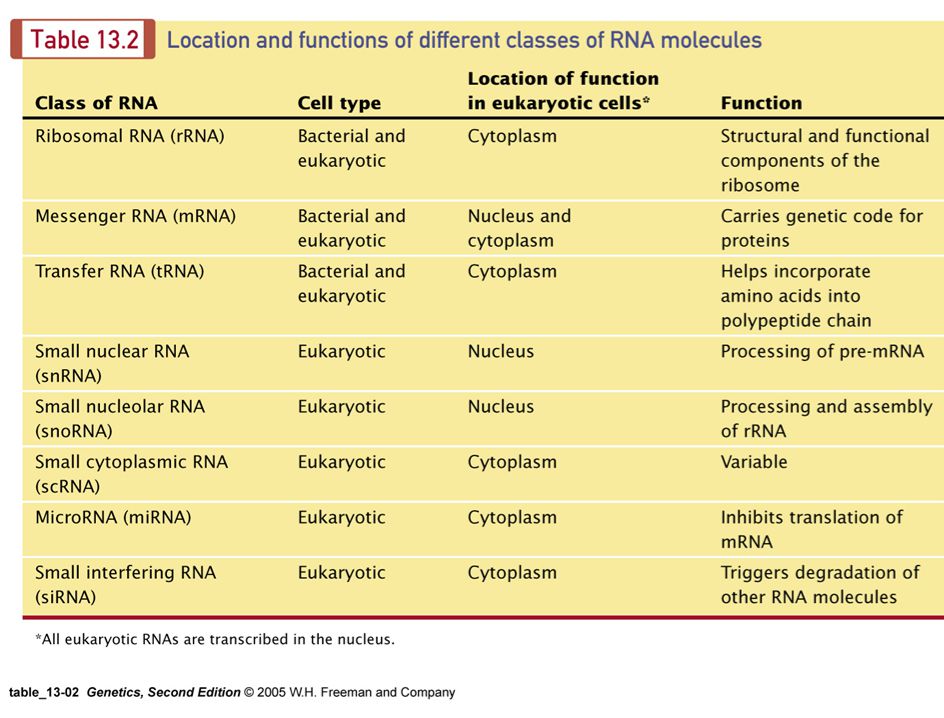
Figure 13.2 All cellular types of RNA are transcribed from DNA.

48
problemas capítulo 2 2ª ed.: 2.7, 2.9 y 2.29 3ª ed.: 2.10, 2.11 y 2.30
1 h de clase: aprox. 4-diciembre-2009 (junto a los problemas de los temas 9 y 10)
")
Presentaciones similares














 comprueban en Neurospora.>")




 Falta el enzima Phehidroxilasa No se produce Tyr.>")
