Descargar la presentación
La descarga está en progreso. Por favor, espere

1
VIAS METABÓLICAS. ENZIMAS
TULB 2015

2
Metabolismo El conjunto de las reacciones químicas que tienen lugar en el seno de los tejidos constituye el metabolismo. Estas reacciones pueden estar dirigidos a: Obtener energía y poder reductor de los alimentos. Degradar los compuestos ingresados en productos más simples, utilizables como precursores para la síntesis de moléculas constituyentes de tejidos y órganos.

3
Procesos degradativos: CATABOLISMO
Metabolismo Procesos degradativos: CATABOLISMO Procesos de biosíntesis: ANABOLISMO

4
VIAS METABÓLICAS Las transformaciones metabólicas se realizan a través de series de reacciones catalizadas por enzimas, ordenadas en una secuencia definida, llamadas VÍAS METABÓLICAS. Cada una de estas convierte un precursor o sustrato inicial en un producto final. Cada uno de los productos intermedios se denominan metabolitos.

5
VÍAS METABÓLICAS El sustrato inicial por acción de una enzima específica, se convierte en un producto que sirve de sustrato para otra enzima de la reacción siguiente y así sucesivamente, hasta llegar al producto final, en una SECUENCIA LINEAL. A B C D E a b c d

6
VÍAS METABÓLICAS B tiene dos vías alternativas.
Existen vías que incluyen puntos de ramificación: C D E A B P Q R B tiene dos vías alternativas. c d b a p q b´

7
VÍAS METABÓLICAS Cuando todas las reacciones de la vía son reversibles, el camino puede recorrerse en ambos sentidos: A B C D E Cuando una o más reacciones de la vía son irreversibles, el camino de vuelta debe realizar desvíos: S

8
VÍAS METABÓLICAS Existen transformaciones que ocurren en forma cíclica: la secuencia ordenada de reacciones termina regenerando el compuesto final.

9
VÍAS METABÓLICAS CICLOS INTERCONECTADOS REACCIONES EN CASCADA

10
VÍAS METABÓLICAS Vías catabólicas: sustrato inicial es reducido a un compuesto más simple. Reacciones oxidativas, exergónicas, con producción de ATP y NADH. Glucólisis, beta-oxidación. Vías anabólicas: formación de nuevos enlaces químicos. Reacciones endergónicas, con hidrólisis de ATP y NADPH como donante de equivalentes de reducción. Gluconeogénesis , síntesis de ácidos grasos. Vías anfibólicas: funcionan como anabólicas o catabólicas de acuerdo a las necesidades celulares. Ciclo de Krebs.

11
ENZIMAS En los seres vivos se producen reacciones químicas para:
Transformar los nutrientes de los alimentos para obtener energía. Transformar nutrientes simples en moléculas complejas y viceversa. Obtener materia prima para la síntesis de nuevas moléculas. Degradar componentes celulares una vez cumplida su vida útil. Extraer energía desde combustibles por oxidación. Polimerizar subunidades para formar macromoléculas.

12
Las transformaciones bioquímicas en los seres vivos ocurren:
a gran velocidad con gran especificidad en condiciones muy moderadas de temperatura, pH, presión, etc. Un catalizador es una sustancia capaz de acelerar una reacción química sin formar parte de los productos finales ni desgastarse en el proceso.

13
ENZIMAS Los catalizadores biológicos son macromoléculas llamadas enzimas La mayoría de las enzimas son proteínas, con la excepción de un pequeño grupo de moléculas de RNA catalítico, llamadas ribozimas. Actividad catalítica: depende de la integridad de su conformación proteica nativa

14
CARACTERISTICAS DE LAS ENZIMAS
PROTEINAS y RNA (Ribozimas): Estructura terciaria y cuaternaria SITIO DE UNION AL SUSTRATO: Uniones no Covalentes (puente de hidrógeno, hidrofóbicas, electrostáticas) NECESITAN DE FACTORES: Inorgánicos (metales) y orgánicos (Coenzimas) ESPECIFICIDAD DE SUSTRATO: Estéreo-especificidad y especificidad geométrica SON REGULABLES: La síntesis de la proteína, su actividad y degradación.
: Estructura terciaria y cuaternaria. SITIO DE UNION AL SUSTRATO: Uniones no Covalentes (puente de hidrógeno, hidrofóbicas, electrostáticas) NECESITAN DE FACTORES: Inorgánicos (metales) y orgánicos (Coenzimas) ESPECIFICIDAD DE SUSTRATO: Estéreo-especificidad y especificidad geométrica. SON REGULABLES: La síntesis de la proteína, su actividad y degradación.")
15
DISTRIBUCION DE LAS ENZIMAS
COMPARTIMENTALIZACION: Diferente localización dentro de la célula. SISTEMAS MULTIENZIMATICOS: Enzimas relacionadas agrupadas formando verdaderos complejos. ENZIMAS MULTIFUNCIONALES: Una enzima que presenta varios sitios catalíticos distintos en una misma cadena polipeptídica.

16
Tipos de reacciones catalizadas por enzimas
Oxido-reducción Rotura y formación de enlaces C-C Reorganizaciones internas Transferencia de grupos Reacciones de condensación

17
NOMENCLATURA Y CLASIFICACIÓN DE ENZIMAS
Nombre común : nombre de uso cotidiano nombre del sustrato de la reacción + asa Ejemplo: sacarasa, ureasa, amilasa, etc Nombres arbitrarios Ej: ptialina salival, pepsina del jugo gástrico, etc. Nombres según el tipo de reacción catalizada. Ej: deshidrogenasas, carboxilasas, etc.

18
Nombre sistemático: Unión Internacional de Bioquímica y Biología Molecular (IUBMB) Se asigna a cada enzima un nombre descriptivo y un número que permite identificarla inequívocamente Nombre común: Hexoquinasa Nombre sistemático: ATP: glucosa fosfotransferasa Código de 4 dígitos: Clase 2: transferasa Subclase 7: fosfotransferasa Subsubclase 1: fosfotransferasa con un grupo hidroxilo como aceptor N° de orden de la enzima en su subclase 1: glucosa como aceptor del grupo fosfato

19
1- OXIDORREDUCTASAS: DESHIDROGENASAS, OXIDASAS, OXIGENASAS, REDUCTASAS, PEROXIDASAS
2- TRANSFERASAS: TRANSCARBOXILASAS, TRANSMETILASAS Y TRANSAMINASAS. 3- HIDROLASAS: ESTERASAS, FOSFATASAS, PEPTIDASAS. 4- LIASAS: DESCARBOXILASAS, HIDRATASAS, DESHIDRATASAS, DESAMINASAS. 5- ISOMERASAS: EPIMERASAS, MUTASAS. 6- LIGASAS: SINTETASAS, CARBOXILASAS

20
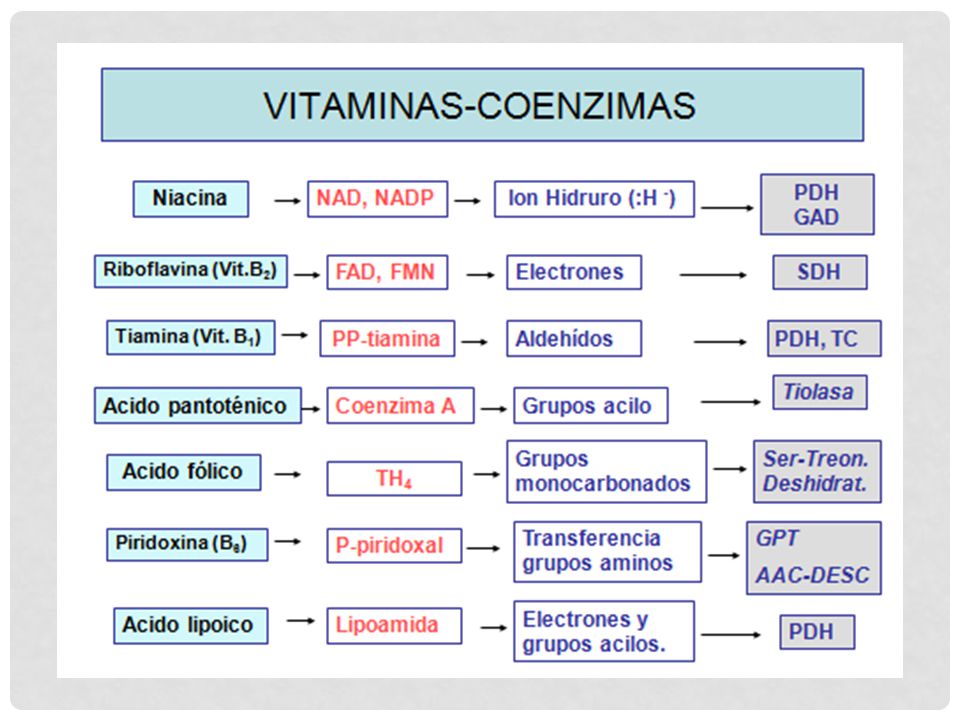
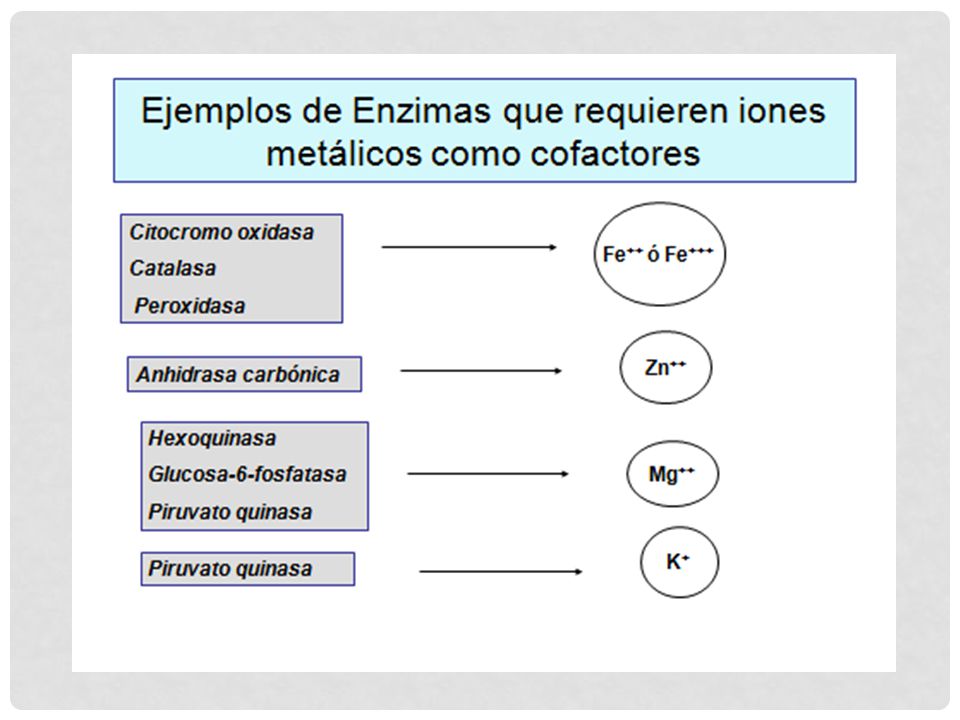
COFACTORES Según su modo de interacción con la enzima COENZIMAS
Unión laxa a la enzima Actuaría como co-sustrato Son moléculas orgánicas pequeñas Muchas se encuentran libres y solo se unen a la enzima en el momento de la catálisis GRUPO PROSTÉTICO Molécula orgánica no proteica Se une fuertemente a la enzima Solo se separa por desnaturalización IONES INORGÁNICOS Aniones - cationes

21
IONES INORGÁNICOS: Fe2+ Mg2+ Mn 2+ Zn2+
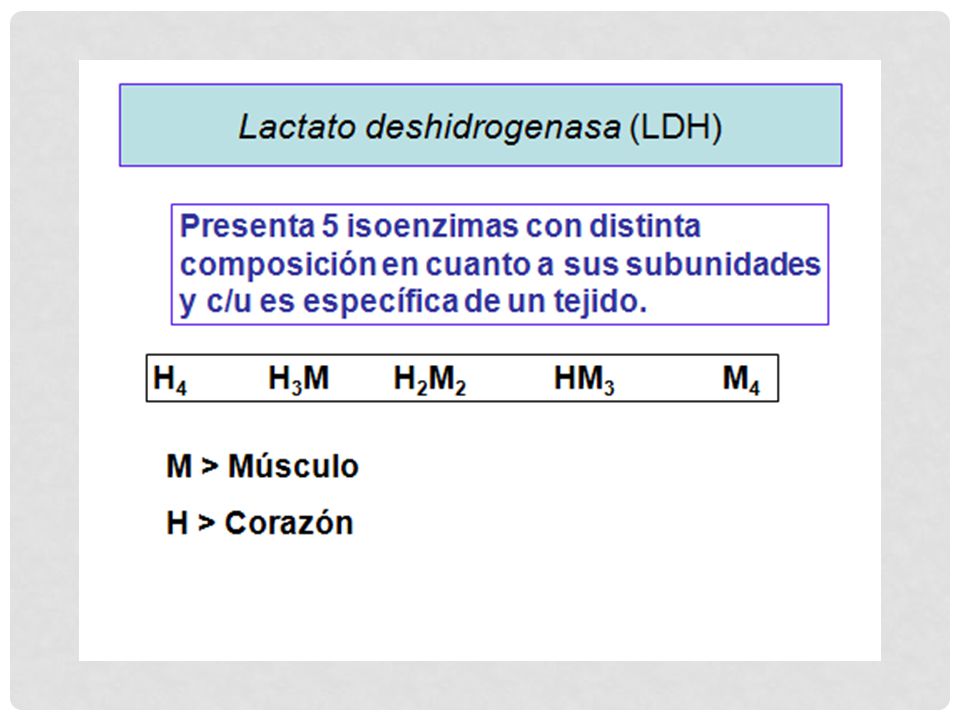
COFACTORES IONES INORGÁNICOS: Fe2+ Mg2+ Mn 2+ Zn2+ En el caso de las COENZIMAS se forma un complejo: HOLOENZIMA = Enzima total APOENZIMA + Proteína Termolábil No dializa COENZIMA No proteínica Termoestable Vitaminas especificidad fad g prostetico nad coenzima Oxidorreductasas, transferasas, isomerasas y ligasas requieren coenzimas

24
PROPIEDADES DE LAS ENZIMAS
LOCALIZACIÓN INTRACELULAR DE LAS ENZIMAS

25
PROPIEDADES DE LAS ENZIMAS
SITIO ACTIVO Región de la molécula enzimática a la que se une el sustrato por uniones no covalentes (puente de hidrógeno, hidrofóbicas, electrostáticas). Es una agrupación de un número no muy grande de aminoácidos distribuidos espacialmente de manera precisa.
. Es una agrupación de un número no muy grande de aminoácidos distribuidos espacialmente de manera precisa.")
26
Hipótesis de Koshland: estructuras adaptables
Hipótesis de Fischer: estructuras rígidas Sitio de unión Sitio catalítico Hipótesis de Koshland: estructuras adaptables

27
Determinación de la actividad enzimática
Se puede determinar, midiendo: La cantidad de producto formado O de sustrato consumido En un tiempo dado Como una sumatoria de todos los factores requeridos para la reacción

28
ACTIVIDAD ENZIMATICA Unidades Internacionales
Cantidad de enzima que cataliza la transformación de 1 μmol (10-6 mol) de Sustrato por minuto: E Sustrato Producto U.I. = mmol de S transformados minuto Siempre bajo condiciones definidas de pH y temperatura
 de Sustrato por minuto: E. Sustrato Producto. U.I. = mmol de S transformados. minuto. Siempre bajo condiciones definidas de pH y temperatura.")
29
ACTIVIDAD ESPECÍFICA Es una expresión que indica la pureza relativa de una preparación enzimática. Relaciona Actividad Enzimática, no con el volumen de muestra, sino con el total de proteínas existentes en la misma. Se define como: UNIDADES DE ENZIMA POR MG DE PROTEÍNAS PRESENTES EN LA MUESTRA

32
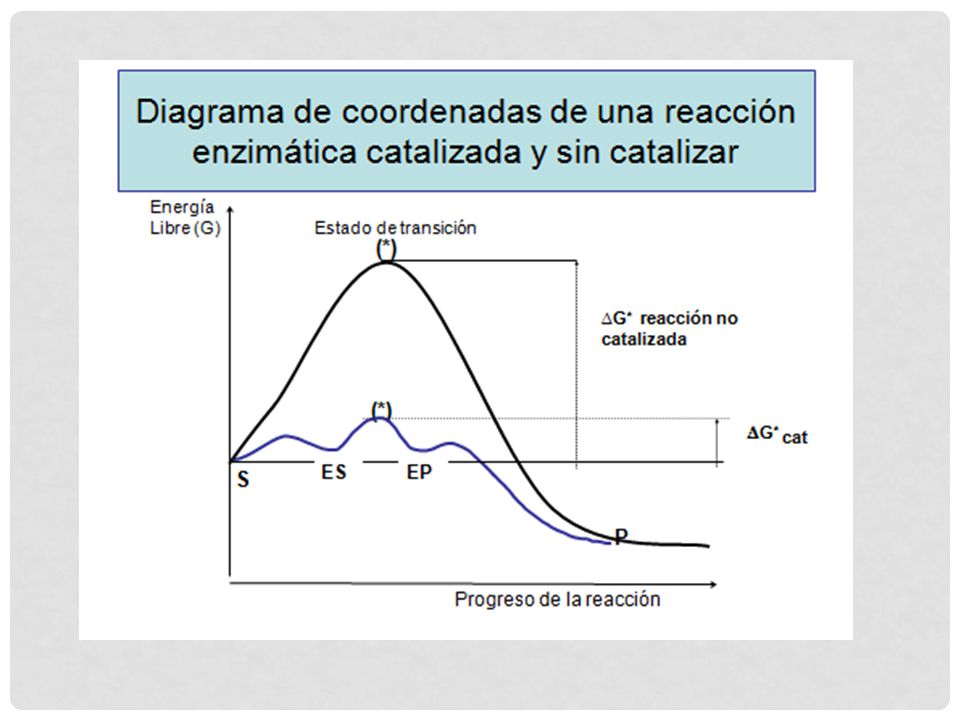
El estado de transición o barrera de activación, representa los cambios que se generan en la molécula del S. Las barreras de activación son importantes para la estabilidad de las biomoléculas. Es por esto que las reacciones sin catalizar tienden a ser muy lentas. Todas las moléculas biológicas son muy estables al pH neutro, temperatura suave y ambiente acuoso en el interior de las células. Todas las reacciones químicas celulares se llevan a cabo porque las enzimas actúan disminuyendo la barrera energética entre el S y el P

33
FACTORES QUE AFECTAN LA VELOCIDAD DE LA REACCIÓN
CONCENTRACIÓN DE ENZIMA Cantidad saturante de sustrato, pH y T°C constantes Velocidad de reacción directamente proporcional a la concentración de enzima ACTIVIDAD ENZIMÁTICA CONCENTRACIÓN DE ENZIMA

34
FACTORES QUE AFECTAN LA VELOCIDAD DE LA REACCIÓN
TEMPERATURA [E], [S], pH: constantes Temperatura óptima

35
Los cambios del pH del medio afectan el estado de ionización de grupos funcionales de las moléculas de E y S. Para la formación del [ES] es necesaria una correcta distribución de las cargas en ambas moléculas pH óptimo 7 6 8 PH ÓPTIMO ES AQUEL EN EL CUAL EL ESTADO DE DISOCIACIÓN DE LOS GRUPOS ESENCIALES ES EL MÁS APROPIADO PARA INTERACCIONAR EN EL COMPLEJO ES

36
FACTORES QUE AFECTAN LA VELOCIDAD DE LA REACCIÓN
EFECTO DEL pH [E], T°C, [S]: constantes

37
FACTORES QUE AFECTAN LA VELOCIDAD DE LA REACCIÓN
CONCENTRACIÓN DEL SUSTRATO [E], pH, T°C: constantes Reacción de primer orden: existe proporcionalidad entre velocidad y [S]. Curva hiperbólica Reacción de 1° orden

38
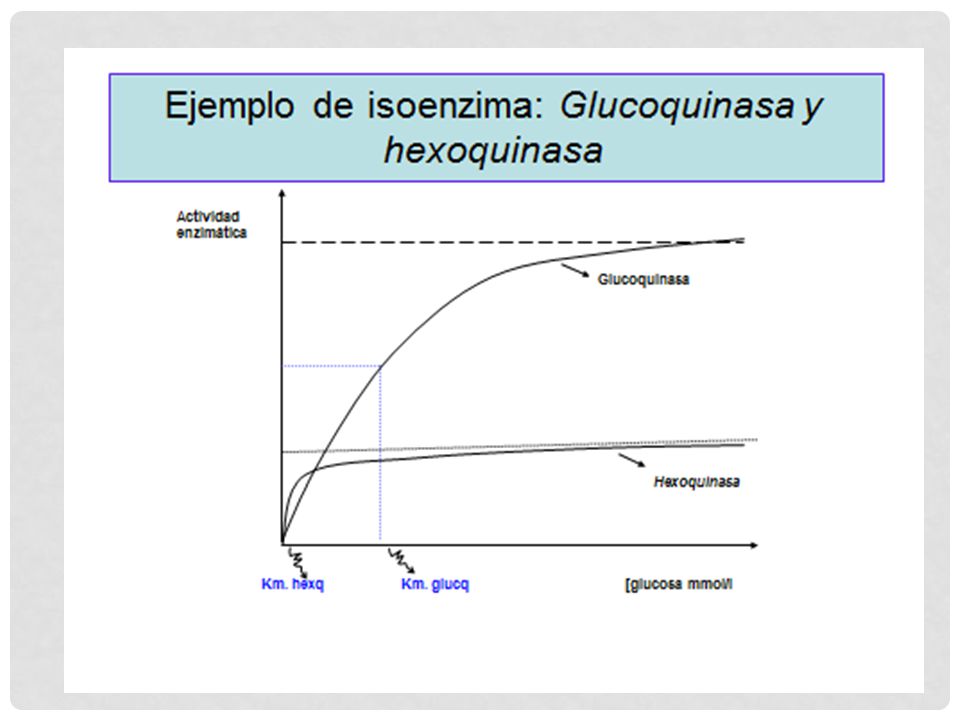
Orden de la reacción Cuando [S] es menor a Km, la velocidad de reacción es proporcional a la [S] por lo tanto la velocidad de reacción es de primer orden con respecto al sustrato. Cuando [S] es mayor a Km, la velocidad de reacción es constante e igual a Vmax por lo tanto la velocidad de reacción es de orden cero con respecto a la concentración del sustrato.
![Orden de la reacción Cuando [S] es menor a Km, la velocidad de reacción es proporcional a la [S]](http://slideplayer.es/slide/4167699/13/images/38/Orden+de+la+reacci%C3%B3n+Cuando+%5BS%5D+es+menor+a+Km%2C+la+velocidad+de+reacci%C3%B3n+es+proporcional+a+la+%5BS%5D.jpg "por lo tanto la velocidad de reacción es de primer orden con respecto al sustrato. Cuando [S] es mayor a Km, la velocidad de reacción es constante e igual a Vmax. por lo tanto la velocidad de reacción es de orden cero con respecto a la concentración del sustrato.")
39
E + P ES E + S ECUACIÓN DE MICHAELIS- MENTEN
Cinética del estado estacionario E + P ES E + S k1 k -1 k2 Reacción de orden cero Reacción de 1° orden Hablar de Km y orden de la reaccion V0= Velocidad inicial de la reacción Vmáx= Velocidad Máxima Km= constante de Michaelis [S]= concentración del sustrato

40
La ecuación de Michaelis- Menten puede ser transformada algebraicamente en ecuaciones utilizables para la determinación práctica del valor de Km. Es decir, transformamos matemáticamente una curva hiperbólica en una recta. Al invertir la ecuación de Michaelis- Menten se obtiene la ecuación de una recta llamada: LINEWEAVER-BURK

41
ECUACIÓN DE LINEWEAVER-BURK
Ordenada al origen = 1/Vmáx. Intersección c/eje x = - 1/Km El valor de km guarda relación inversa con la afinidad de la E por el S A MAYOR AFINIDAD, MENOR VALOR DE Km

42
Los valores de Km y Vmax de una enzima se determinan experimentalmente.
Midiendo las velocidades iniciales de reacción a diferentes [S]. Esta representación de dobles recíprocas tiene la ventaja de permitir una determinación más precisa de Vmax, valor que solo puede obtenerse a partir de la fórmula de Vo frente a [S].

43
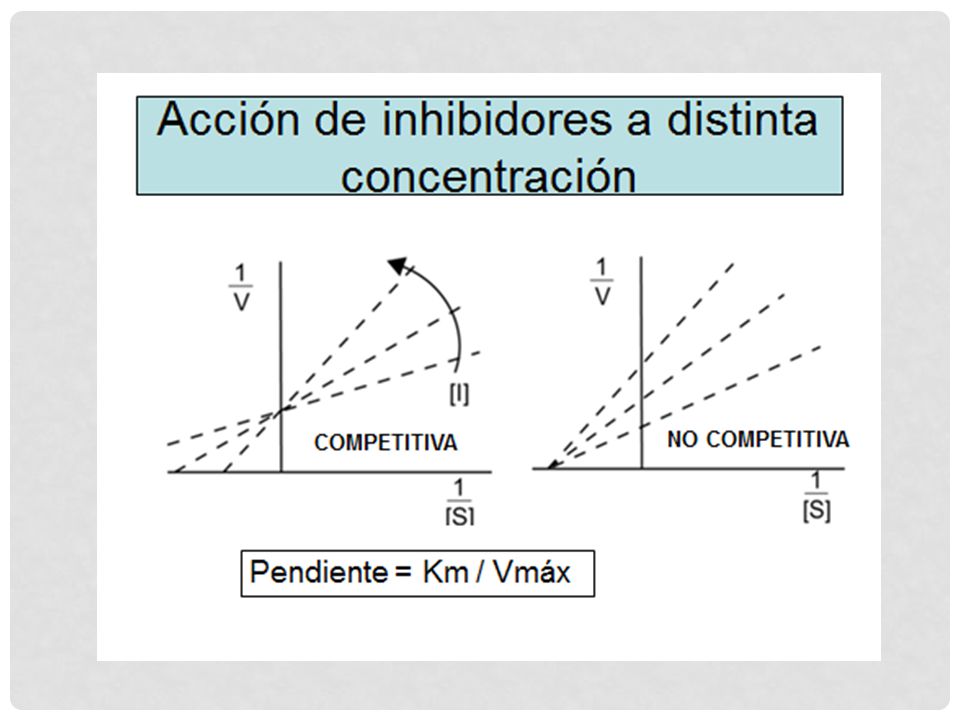
INHIBIDORES ENZIMÁTICOS
Agentes químicos que inhiben la acción catalítica de enzimas. Algunos ejercen su acción uniéndose a sitios o grupos funcionales esenciales de la E. Estas sustancias son de gran utilidad para estudiar algunos aspectos de la acción enzimática. Por ejemplo, que grupos funcionales son los que participan en la catálisis o son responsables de asegurar los requerimientos estructurales para la unión E-S. La inhibición puede ser reversible o irreversible.

44
INHIBICIÓN DE LA ACTIVIDAD ENZIMÁTICA
INHIBIDORES IRREVERSIBLES Sustancias que producen un cambio permanente en la molécula de enzima. Producen un deterioro de su capacidad catalítica. Ej. Insecticidas, «inhibidores suicidas» (semejantes estructuralmente al S y ocupan el sitio activo, caso del allopurinol- xantina oxidasa como tratamiento de la Gota)
")
45
INHIBIDORES REVERSIBLES COMPETITIVOS
Reacción de succinato deshidrogenas en pizarron

46
Similitud Estructural del I con el S Ambos compiten por el sitio activo de E

47
Semejanza estructural

48
INHIBIDORES REVERSIBLES NO COMPETITIVOS
La unión S-E no esta afectada, porque el I se une a la E libre o al [ES]. Una vez formado el [ESI] la enzima se inactiva. El valor de Km no se modifica, pues el S reacciona con la E igual que en ausencia del I. Ejemplos: reactivos que se unen a grupos –SH de cisteína, indispensables para algunas enzimas, iones metálicos (Cu2+, Hg2+ y Ag2+). Otros I se unen a los metales componentes de la E, por ejemplo: cianuro se une fuertemente al Fe de los citocromos, catalasas y así bloquean su actividad.
. Otros I se unen a los metales componentes de la E, por ejemplo: cianuro se une fuertemente al Fe de los citocromos, catalasas y así bloquean su actividad.")
49
INHIBIDORES REVERSIBLES NO COMPETITIVOS
Se unen a la E en un sitio diferente al sitio activo Este tipo de inhibición no es revertida por aumento de la [S]

51
REGULACIÓN DE LA ACTIVIDAD ENZIMÁTICA
La AE en las células se ajustan a las necesidades fisiológicas, cambiantes permanentemente. A [S] bajas, la AE es proporcional a los niveles de S. En condiciones fisiológicas, la [S] se encuentra por debajo o próxima al Km, así los niveles de S, determinarán la mayor o menor AE. A mayor [S] mayor AE La E que cataliza la primera etapa de una vía metabólica suele ser reguladora.

52
Los seres vivos han generado mecanismos para regular las rutas bioquímicas.
La regulación es esencial por las siguientes razones: Mantenimiento de un estado ordenado no hay desperdicio de recursos. Conservación de la energía utilización de la E suficiente en las células, para consumir los nutrientes según sus necesidades energéticas. Respuesta a variaciones ambientales son los ajustes rápidos que deben realizar las células (pH, [S], T°C), por su capacidad de aumentar o disminuir la velocidad de las reacciones específicas.
, por su capacidad de aumentar o disminuir la velocidad de las reacciones específicas.")
53
Regulación por retroinhibición o Feed- back negativo
ENZIMAS ALOSTÉRICAS Regulación por retroinhibición o Feed- back negativo Tanto la inhibición como la activación son reversibles. El agente modificador actúa uniéndose a la E en un sitio diferente al sitio catalítico Estas Enzimas Alostéricas, además del sitio catalítico, presentan otros sitios reguladores donde se unirán moléculas que actúan sobre su AE. Moduladores, modificadores o efectores alostéricos Positivos o negativos para la AE. Efector Homotrópico: el propio sustrato actúa como efector positivo. Efector Heterotrópico: efector diferente del sustrato.

54
Modifican la conformación de la E
Los moduladores alostéricos (-) no deben ser confundidos con los Inhibidores. Forma T Modifican la conformación de la E Forma R
 no deben ser confundidos con los Inhibidores. Forma T. Modifican la conformación de la E. Forma R.")
55
REGULACIÓN DE LA ACTIVIDAD ENZIMÁTICA
ENZIMAS ALOSTÉRICAS Modificación enzimática inmediata Curvas sigmoideas: las enzimas con este tipo de cinética pueden regular la [S]

58
MODIFICACIÓN COVALENTE
REGULACIÓN DE LA ACTIVIDAD ENZIMÁTICA MODIFICACIÓN COVALENTE (modificación enzimática inmediata o minutos ) QUINASA DE PROTEÍNAS ATP ADP ENZIMA OH OPO3= HPO4= H2O FOSFATASA DE PROTEÍNAS Adición o remoción de grupos fosfato sobre residuos de Serina, Treonina o Tirosina específicos de la enzima. Quinasas de proteínas familia de enzimas que catalizan reacciones de fosforilación utilizan como dador del grupo fosfato al ATP. Fosfatasas de proteínas separan los grupos fosfatos de las enzimas fosforiladas
 QUINASA DE PROTEÍNAS. ATP. ADP. ENZIMA. OH. OPO3= HPO4= H2O. FOSFATASA DE PROTEÍNAS. Adición o remoción de grupos fosfato sobre residuos de Serina, Treonina o Tirosina específicos de la enzima. Quinasas de proteínas familia de enzimas que catalizan reacciones de fosforilación utilizan como dador del grupo fosfato al ATP. Fosfatasas de proteínas separan los grupos fosfatos de las enzimas fosforiladas.")
59
Glucógeno fosforilasa

Presentaciones similares






















![1 KM 1 1 Vo= Vmax [S] + Vmax Y = m x + b = m = b.](/2/161459/big_thumb.jpg "1 KM 1 1 Vo= Vmax [S] + Vmax Y = m x + b = m = b.>")

