Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Metabolismo: Tipos de reacciones metabólicas
Características de las reacciones metabólicas Catabolismo Respiración aerobia

2
Metabolismo El metabolismo es el conjunto de reacciones bioquímicas que permiten la transformación de los nutrientes en el interior de las células, bien para formar materia propia o bien para obtener energía. Se distinguen dos tipos de reacciones metabólicas: catabolismo y anabolismo. - El catabolismo es el conjunto de reacciones que sirven para descomponer o degradar sustancias y, al hacerlo, desprenden o liberan energía. Por tanto, transforma sustancias complejas en otras más sencillas. XY X + Y El anabolismo es el conjunto de reacciones que sirven para sintetizar sustancias y necesitan para ello energía. Por tanto, transforma sustancias sencillas en otras más complejas, que la célula puede emplear para formar materia propia o para descomponerlas de nuevo. X + Y → XY Catabolismo y anabolismo son inseparables y muchas de sus reacciones se hallan acopladas, dependiendo unas de otras. XY X + Y reacciones catabólicas energía 2H (2H+ + 2e-) X + Y → XY reacciones anabólicas
 X + Y → XY reacciones anabólicas.")
3
Algunas características de las reacciones metabólicas
Las reacciones metabólicas están acopladas energéticamente a través del ATP Las reacciones metabólicas son reacciones de oxidorreducción Las reacciones metabólicas tienen una secuencia encadenada y están catalizadas por enzimas Las reacciones metabólicas están compartimentadas.

4
XY X + Y reacciones catabólicas exergónicas
Las reacciones metabólicas están acopladas energéticamente a través del ATP Molécula rica en energía Moléculas pobres en energía XY X + Y reacciones catabólicas exergónicas energía ADP + P ATP energía X + Y → XY reacciones anabólicas endergónicas Moléculas pobres en energía Molécula rica en energía

5
La síntesis de ATP puede ser: Por fosforilación a nivel de sustrato
Una molécula (R-P) cede P al ADP: R-P + ADP R + ATP Por fosforilación oxidativa La energía para unir enlaces fosfato procede de reacciones de oxidorreducción: ADP + Pi + energía de reacciones redox ATP Por fotofosforilación La energía para la fosforilación procede de reacciones de oxidorreducción provocadas por la luz. ADP + Pi + energía de reacciones redox por la luz ATP
 cede P al ADP: R-P + ADP R + ATP. Por fosforilación oxidativa. La energía para unir enlaces fosfato procede de reacciones de oxidorreducción: ADP + Pi + energía de reacciones redox ATP. Por fotofosforilación. La energía para la fosforilación procede de reacciones de oxidorreducción provocadas por la luz. ADP + Pi + energía de reacciones redox por la luz ATP.")
6
La oxidación se produce por La reducción se produce por
2. Las reacciones metabólicas son reacciones de oxidorreducción Conviene tener en cuenta los distintos conceptos de oxidación y reducción: La oxidación se produce por La reducción se produce por Pérdida de electrones, por tanto, conlleva aumento del número de oxidación Ganancia de electrones, por tanto, conlleva disminución del número de oxidación Pérdida de hidrógeno Incorporación de hidrógeno Incorporación de oxígeno Pérdida de oxígeno

7
En las reacciones bioquímicas o metabólicas la pérdida o ganancia de electrones va ligada a la de protones (hidrogeniones) en forma de hidrógeno: 2e- + 2H+ = 2H (H2) Por tanto, el concepto de oxidación-reducción que interesa considerar en biología es el relacionado con la cesión y captación de hidrógenos Estos hidrógenos son transportados por coenzimas de oxidorreducción que pasan de su forma oxidada a la reducida o viceversa. Por ejemplo: reducción NAD (oxidado) NAD. 2H (reducido) oxidación
 NAD. 2H (reducido) oxidación.")
8
XY X + Y reacciones catabólicas de oxidación
2. Las reacciones metabólicas son reacciones de oxidorreducción: se transfieren electrones ligados a protones en forma de hidrógeno (H+ + e-) por mediación de coenzimas de oxidorreducción Molécula reducida Moléculas oxidadas XY X + Y reacciones catabólicas de oxidación 2H (2H+ + 2e-) NAD (oxidado) NAD. 2H (reducido) 2H (2H+ + 2e-) X + Y XY reacciones anabólicas de reducción Moléculas oxidadas Molécula reducida
 por mediación de coenzimas de oxidorreducción. Molécula reducida. Moléculas oxidadas. XY X + Y reacciones catabólicas de oxidación. 2H (2H+ + 2e-) NAD (oxidado) NAD. 2H (reducido) 2H (2H+ + 2e-) X + Y XY reacciones anabólicas de reducción. Moléculas oxidadas. Molécula reducida.")
9
De ahí que, como hemos visto:
Además, en la transferencia de hidrógenos hay ligada una transferencia de energía ya que, las sustancias de potencial redox electronegativo (con un nivel energético elevado) ceden electrones a sustancias con potencial redox electropositivo (con un nivel energético inferior) y se libera energía. Compuesto con potencial redox alto (electronegativo) De ahí que, como hemos visto: En las reacciones catabólicas, en las que unos compuestos ceden hidrógeno a otros (oxidación), se libera energía. En las reacciones anabólicas, en las que unos compuestos captan hidrógeno de otros (reducción), se requiere energía. 2H (2H+ + 2e-) energía Compuesto con potencial redox bajo (electropositivo) Por tanto, Las reacciones del catabolismo son de oxidación por deshidrogenación y exergónicas Las reacciones del anabolismo son de reducción por hidrogenación y endergónicas
 ceden electrones a sustancias con potencial redox electropositivo (con un nivel energético inferior) y se libera energía. Compuesto con. potencial redox alto. (electronegativo) De ahí que, como hemos visto: En las reacciones catabólicas, en las que unos compuestos ceden hidrógeno a otros (oxidación), se libera energía. En las reacciones anabólicas, en las que unos compuestos captan hidrógeno de otros (reducción), se requiere energía. 2H (2H+ + 2e-) energía. Compuesto con. potencial redox bajo. (electropositivo) Por tanto, Las reacciones del catabolismo son de oxidación por deshidrogenación y exergónicas. Las reacciones del anabolismo son de reducción por hidrogenación y endergónicas.")
10
3. Las reacciones metabólicas…
… tienen una secuencia encadenada y… están catalizadas por enzimas El producto final de una sirve como producto inicial de la siguiente, como si se tratara de eslabones de una cadena Permite que se puedan realizar a las temperaturas ordinarias y a las velocidades adecuadas

11
4- Las reacciones metabólicas están compartimentadas.
Distintos tipos de vías metabólicas están separadas en compartimentos o “territorios” celulares diferentes, como consecuencia de la situación que ocupan las enzimas que las catalizan. Se evitan así interferencias y pueden realizarse distintos procesos a la vez. CITOPLASMA MITOCONDRIA

12
Diferencias entre metabolismo de células autótrofas y heterótrofas
Las células autótrofas tienen una etapa de anabolismo autótrofo y otra de anabolismo heterótrofo Las células heterótrofas sólo tienen una etapa de anabolismo heterótrofo Las reacciones del catabolismo son prácticamente idénticas

13
Catabolismo. Características y tipos
Reacciones de: Descomposición Oxidación Exergónicas Tipos de catabolismo Fermentación Se libera poca energía Respiración anaerobia Respiración aerobia Se libera mucha energía

14
Respiración aerobia La respiración aerobia es un tipo de catabolismo total de cualquier forma de materia orgánica que se oxida hasta dar CO2, liberando energía e hidrógenos, los cuales son aceptados por el oxígeno, que se reduce formando agua. La materia orgánica utilizada como sustrato suele ser glúcidos y lípidos Suele identificarse el catabolismo total de los glúcidos en presencia de oxígeno con la respiración aerobia La glucosa es la molécula más utilizada por las células para obtener energía. Por ello, se emplea la reacción química que expresa su degradación total en presencia de oxígeno para representar en conjunto todo el proceso de la respiración aerobia C6H12O O2 6CO2 + 6H2O + energía (en ATP y calor) (glucosa) Esta “reacción global” no es más que una representación simplificada del proceso En el metabolismo celular real no es posible que los H de la glucosa se unan al O2
 (glucosa) Esta reacción global no es más que una representación simplificada del proceso. En el metabolismo celular real no es posible que los H de la glucosa se unan al O2.")
15
Catabolismo de glúcidos
Respiración aerobia Catabolismo de glúcidos Podemos considerar el proceso dividido en etapas, aunque en la célula suceden de forma consecutiva e ininterrumpida, y pueden hacerlo en compartimentos diferentes: Etapa preliminar: las grandes moléculas de polisacáridos llegan a descomponerse en monosacáridos, fundamentalmente, en glucosa. Se desarrollan en el hialoplasma. Glucolisis: serie de reacciones de oxidación por las que la glucosa se transforma en ácido pirúvico. Tiene lugar también en el hialoplasma Como se ha advertido, algunos autores restringen el término respiración a las dos últimas etapas consideradas aquí (ciclo de Krebs y cadena respiratoria), ya que éstas sólo se realizan si la célula dispone de oxígeno. Ciclo de Krebs. transformación previa del ácido pirúvico en Acetil-coenzima A (acetilCoA) y la serie de reacciones cíclicas (llamadas también ciclo de los ácidos tricarboxílicos), en las que se producen descarboxilaciones oxidativas, que liberan CO2 e hidrógenos. Tiene lugar en la matriz de la mitocondria. Fosforilación oxidativa. se sintetiza ATP Los hidrógenos de las reacciones anteriores llegan a la cadena respiratoria, para que se transporten hasta el oxígeno, al que se unen para formar agua. En el transporte por reacciones redox se libera energía que se emplea en formar los enlaces fosfato del ATP Tiene lugar en las crestas mitocondriales.
, ya que éstas sólo se realizan si la célula dispone de oxígeno. Ciclo de Krebs. transformación previa del ácido pirúvico en Acetil-coenzima A (acetilCoA) y la serie de reacciones cíclicas (llamadas también ciclo de los ácidos tricarboxílicos), en las que se producen descarboxilaciones oxidativas, que liberan CO2 e hidrógenos. Tiene lugar en la matriz de la mitocondria. Fosforilación oxidativa. se sintetiza ATP Los hidrógenos de las reacciones anteriores llegan a la cadena respiratoria, para que se transporten hasta el oxígeno, al que se unen para formar agua. En el transporte por reacciones redox se libera energía que se emplea en formar los enlaces fosfato del ATP. Tiene lugar en las crestas mitocondriales.")
16
Etapa preliminar a la sangre (hígado) glucosa + Pi Glucógeno G1P (n-glucosas) (músculo ) G6P Glucógeno glucolisis (n-1glucosas) energía
 energía.")
17
Glucolisis Glucosa 2 pirúvico NAD.2H ATP

18
De forma simplificada podemos representar la glucolisis así:
Glucosa 2 pirúvico NAD.2H ATP

20
Descarboxilación oxidativa del pirúvico y ciclo de Krebs
oxidación Descarboxilación oxidativa oxidación Fosforilación a nivel de sustrato

21
Resumen del ciclo de Krebs
Ácido pirúvico CO2 NAD.2H Consideraremos que el GTP es equivalente al ATP, ya que se puede transformar así: GTP + ADP GDP + ATP glucosa 6 CO NAD.2H FAD.2H ATP El ciclo de Krebs tampoco consume oxígeno, pero sólo funciona si hay disponibilidad de éste, ya que de esa forma las coenzimas ceden los hidrógenos y se regeneran para seguir realizando oxidaciones en el ciclo.

22
Cadena respiratoria, transporte de electrones y fosforilación oxidativa
Terminada la glucolisis y el ciclo de Krebs el rendimiento de una molécula de glucosa es: glucosa 6 CO NAD.2H FAD.2H ATP Cadena respiratoria Ahora todas las coenzimas reducidas, tanto las producidas en el citoplasma por la glucolisis, como las que se hallan en la matriz mitocondrial,, han de oxidarse en la cadena respiratoria

23
En el transporte se libera energía que se almacena en ATP,
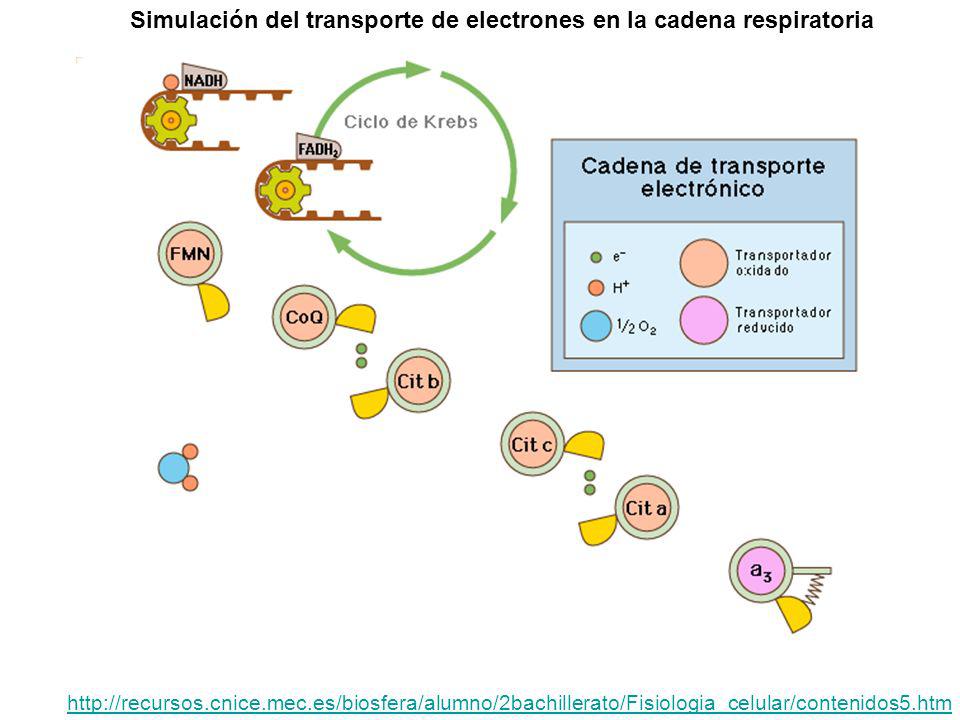
Cadena respiratoria, transporte de electrones y fosforilación oxidativa La cadena respiratoria es un conjunto de moléculas que se encuentran en la membrana interna de la mitocondria. Permiten, mediante una serie de reacciones de oxidorreducción, transportar los H (H+ y e-) desde las coenzimas reducidas hasta el aceptor final (oxígeno) En el transporte se libera energía que se almacena en ATP, es decir, se produce la fosforilación oxidativa. NAD.2H (2H+ + 2e-) Transporte de e- 1/2 O2 Aceptor final de e- 2H 2H 2H 2H 2H 2H cadena respiratoria NAD H2O energía ADP + Pi ATP Fosforilación oxidativa
 desde las coenzimas reducidas hasta el aceptor final (oxígeno) En el transporte se libera energía que se almacena en ATP, es decir, se produce la fosforilación oxidativa. NAD.2H (2H+ + 2e-) Transporte de e- 1/2 O2. Aceptor. final de e- 2H. 2H. 2H. 2H. 2H. 2H. cadena respiratoria. NAD. H2O. energía. ADP + Pi. ATP. Fosforilación oxidativa.")
24
Los coenzimas reducidos y el oxígeno tienen niveles de potencial redox muy alejados, lo que impide la cesión directa entre unos y otro. Los componentes de la cadena se agrupan en complejos moleculares con distintos potenciales redox que hacen el papel de escalones intermedios entre los coenzimas reducidos y el oxígeno. De este modo es como si los electrones fueran cayendo por una serie de peldaños, y en cada salto liberan sólo cierta cantidad de energía. Así, se evita el salto directo de electrones y la liberación brusca de energía entre dos niveles con potenciales redox muy alejados (el del coenzima y el del oxígeno). Representación de los componentes de la cadena respiratoria según sus potenciales redox y niveles energéticos
. Representación de los componentes de la cadena respiratoria según sus potenciales redox y niveles energéticos.")
25
La cadena respiratoria y el transporte de electrones
(Descripción del transporte de H+ y e-) Transporte de H+ y e- E E Transporte sólo de e- E El oxígeno acepta los e- y los H+ y se forma H2O Cada NAD.2H: 3ATP Cada FAD.2H: 2ATP E saltos en los que se libera energía para ATP
 Transporte de H+ y e- E. E. Transporte sólo de e- E. El oxígeno acepta los e- y los H+ y se forma H2O. Cada NAD.2H: 3ATP. Cada FAD.2H: 2ATP. E. saltos en los que se. libera energía para ATP.")
26
Cadena respiratoria y fosforilación oxidativa
(representación en su territorio celular: la membrana interna de la mitocondria) E E E E E E Teoría quimiosmótica de la fosforilación oxidativa: La energía del transporte de e− no se emplea directamente en la fosforilación (síntesis de ATP) La energía del transporte de e− se emplea primero en realizar un transporte activo de H+ desde la matríz hacia el espacio intermembrana (Los complejos I, III y IV actúan como bombas de H+). Los H+ allí acumulados liberarán energía si vuelven a la matriz a favor de gradiente (lo que sólo pueden hacer a través de las partículas F) La energía liberada en las partículas F se empleará ahora en la fosforilación: ADP+Pi =ATP
 E. E. E. E. E. E. Teoría quimiosmótica de la fosforilación oxidativa: La energía del transporte de e− no se emplea directamente en la fosforilación (síntesis de ATP) La energía del transporte de e− se emplea primero en realizar un transporte activo de H+ desde la matríz hacia el espacio intermembrana (Los complejos I, III y IV actúan como bombas de H+). Los H+ allí acumulados liberarán energía si vuelven a la matriz a favor de gradiente (lo que sólo pueden hacer a través de las partículas F) La energía liberada en las partículas F se empleará ahora en la fosforilación: ADP+Pi =ATP.")
27
Simulación del transporte de electrones en la cadena respiratoria
28
H+ H+ H+ Estructura de las partículas F (factores de acoplamiento)
Fosforilación oxidativa H+ H+ H+ ATPsintetasas ADP + Pi ATP Fosforilación oxidativa energía H+ H+ H+

29
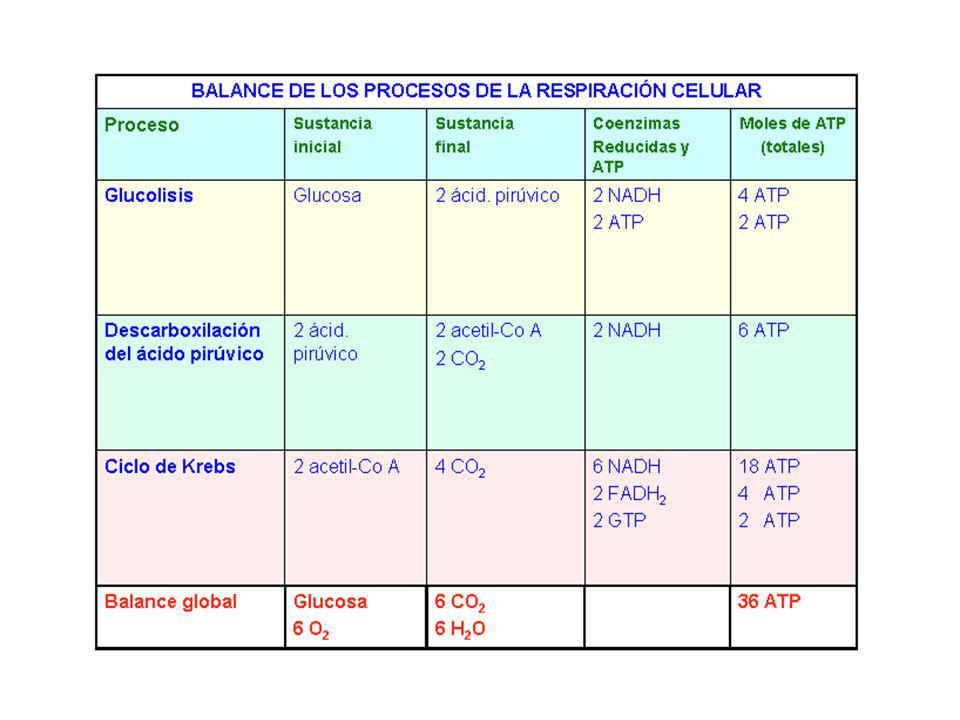
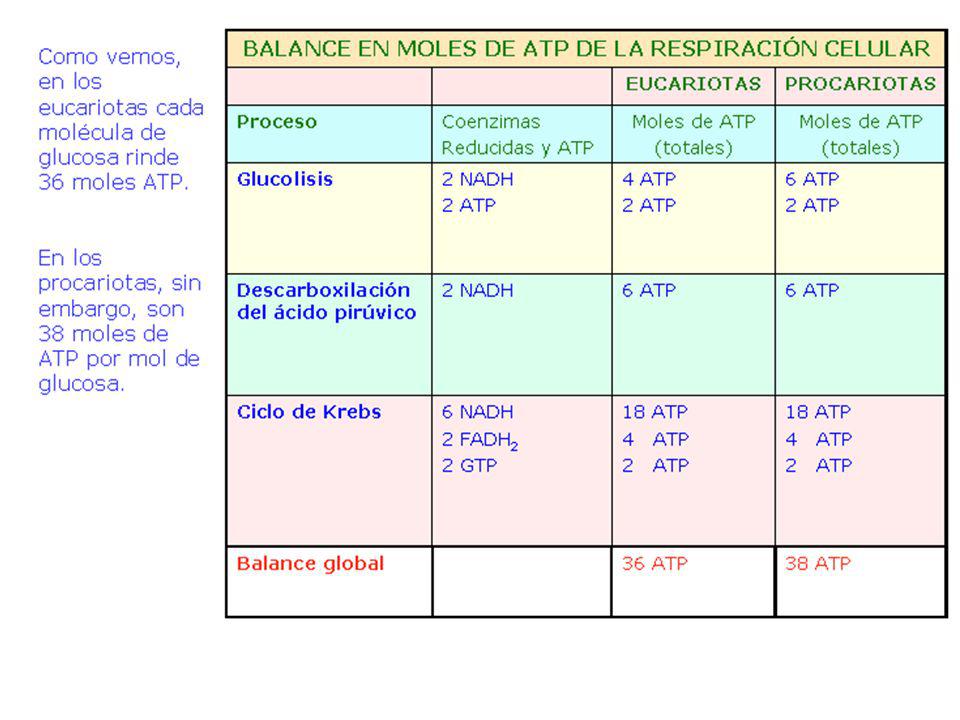
A partir de un mol de glucosa
Balance energético del catabolismo total de la glucosa A partir de un mol de glucosa nº de coenzimas reducidas nº de ATP por fosforilación oxidativa nº de ATP por fosforilación a nivel de sustrato TOTAL Citoplasma Glucolisis 2 NAD.2H 2 x 3 = 6 ATP 2 ATP 8 ATP(*) Mitocondria 2 Pirúvico a 2 AcetilCoA 2 x 3 = 6 ATP 6 ATP Ciclo Krebs (2 veces) 6 NAD.2H 2 FAD.2H 6 x 3 = 18 ATP 2 x 2 = 4 ATP 2ATP 24 ATP 10 NAD.2H 2FAD.2H 34 ATP 4 ATP 38 ATP (*) En las células eucarióticas el paso de los NAD.2H del citoplasma a la mitocondria suponen gasto de ATP. Concretamente, en las células musculares se gastan 2 ATP. Por tanto, un mol de glucosa produce 36 ATP Animación balance energético de la glucosa:
 Mitocondria. 2 Pirúvico a. 2 AcetilCoA. 2 x 3 = 6 ATP. 6 ATP. Ciclo Krebs. (2 veces) 6 NAD.2H. 2 FAD.2H. 6 x 3 = 18 ATP. 2 x 2 = 4 ATP. 2ATP. 24 ATP. 10 NAD.2H. 2FAD.2H. 34 ATP. 4 ATP. 38 ATP. (*) En las células eucarióticas el paso de los NAD.2H del citoplasma a la mitocondria. suponen gasto de ATP. Concretamente, en las células musculares se gastan 2 ATP. Por tanto, un mol de glucosa produce 36 ATP. Animación balance energético de la glucosa: title=Animaci%C3%B3n:_visi%C3%B3n_general_de_la_oxidaci%C3%B3n_de_la_glucosa.")
30
Balance energético del catabolismo total de la glucosa
La hidrólisis de un ATP a ADP + Pi libera 7,3 Kcal/mol (esta es la energía contenida en cada enlace fosfato) Por tanto, 38 ATP suponen a la célula unas 277 kcal por cada 180 g de glucosa (1 mol) La combustión total artificial de la glucosa se sabe que liberaría 686 Kcal. Se puede calcular el rendimiento energético en %: 277 kcal x 100 / 686 Kcal = 40 El rendimiento energético para la célula es del 40%; el resto se pierde en forma de calor
 Por tanto, 38 ATP suponen a la célula unas 277 kcal por cada 180 g de glucosa (1 mol) La combustión total artificial de la glucosa se sabe que liberaría 686 Kcal. Se puede calcular el rendimiento energético en %: 277 kcal x 100 / 686 Kcal = 40. El rendimiento energético para la célula es del 40%; el resto se pierde en forma de calor.")
33
Catabolismo de los lípidos
Ácido de nC FAD.2H + NAD.2H + acetilCoA (2C) + ácido de (n-2)C
 + ácido de (n-2)C.")
34
Hélice de Lynen Se puede generalizar que para un ácido graso saturado de n átomos de C, siendo n par, la hélice da (n/2)-1 vueltas y se obtienen: (n/2) unidades de acetilCoA, (n/2)-1 de NAD.2H (n/2)-1 de FAD.2H. Cadena respiratoria Ciclo de Krebs
 unidades de acetilCoA, (n/2)-1 de NAD.2H. (n/2)-1 de FAD.2H. Cadena. respiratoria. Ciclo de. Krebs.")
35
Balance energético del catabolismo total de un ácido graso
La -oxidación del ácido caproico de 6 átomos de C da lugar a: CH3-(CH2)4-COOH 3 AcetilCoA + 2 NAD.2H + 2 FAD.2H ácido caproico β-oxidación de ácido graso de nC nº de acetilCoA n/2 nº de coenzimas reducidas (n/2)-1 nº de ATP por fosforilación oxidativa y nivel sustrato TOTAL A partir de un mol del ác. caproico (6C) (6/2)-1=2 NAD.2H (6/2)-1=2 FAD.2H 2 x 3ATP = 6 ATP 2 x 2ATP = 4 ATP 10 ATP 6/2= 3 acetilCoA 3 x 3= 9 NAD.2H 3 x 1=3 FAD.2H 3 veces el ciclo Krebs (una vez por cada acetilCoA) 9x3ATP=27 ATP 3x2ATP=6 ATP 3 ATP(de sustrato) 36 ATP 11 NAD.2H 5 FAD.2H 11x3ATP=33 ATP 5x2ATP=10 ATP 3 ATP (de sustrato) 46 ATP El rendimiento del ácido caproico es de 8 ATP más que el de la glucosa. En general, a mayor longitud de la cadena de ácido graso, mayor rendimiento energético
4-COOH 3 AcetilCoA + 2 NAD.2H + 2 FAD.2H. ácido caproico. β-oxidación de ácido graso de nC. nº de acetilCoA. n/2. nº de coenzimas reducidas. (n/2)-1. nº de ATP por fosforilación oxidativa y nivel sustrato. TOTAL. A partir de un mol del ác. caproico. (6C) (6/2)-1=2 NAD.2H. (6/2)-1=2 FAD.2H. 2 x 3ATP = 6 ATP. 2 x 2ATP = 4 ATP. 10 ATP. 6/2= 3 acetilCoA. 3 x 3= 9 NAD.2H. 3 x 1=3 FAD.2H. 3 veces el ciclo Krebs (una vez por cada acetilCoA) 9x3ATP=27 ATP. 3x2ATP=6 ATP. 3 ATP(de sustrato) 36 ATP. 11 NAD.2H. 5 FAD.2H. 11x3ATP=33 ATP. 5x2ATP=10 ATP. 3 ATP (de sustrato) 46 ATP. El rendimiento del ácido caproico es de 8 ATP más que el de la glucosa. En general, a mayor longitud de la cadena de ácido graso, mayor rendimiento energético.")
36
Los lípidos son los mejores combustibles para las células animales.
Sin embargo, las células utilizan los glúcidos preferentemente antes que las grasas por varias razones: La movilización de las reservas de grasas es un proceso más lento (los músculos movilizan el glucógeno antes que la grasa de los tejidos adiposos) - En condiciones anaerobias las grasas no pueden catabolizarse, ya que en tal situación no funciona el ciclo de Krebs, mientras que los glúcidos sí tienen la vía alternativa anaerobia de las fermentaciones a nivel del ácido pirúvico >Las células vegetales cuentan en los glioxisomas con una vía metabólica especial, llamada ciclo del glioxilato, por la que pueden transformar los ácidos grasos en glúcidos Los ácidos grasos no pueden transformarse en glucosa en las células animales, porque no existe tal vía metabólica, por lo que las reservas de grasa no sirven para mantener los niveles de glucosa en sangre - Los ácidos grasos necesitan moléculas especiales para ser transportadas por la sangre desde las células del tejido adiposo (donde se almacenan), hasta las que necesitan energía, mientras la glucosa circula sin dificultad dada su solubilidad.
 - En condiciones anaerobias las grasas no pueden catabolizarse, ya que en tal situación no funciona el ciclo de Krebs, mientras que los glúcidos sí tienen la vía alternativa anaerobia de las fermentaciones a nivel del ácido pirúvico. >Las células vegetales cuentan en los glioxisomas con una vía metabólica especial, llamada ciclo del glioxilato, por la que pueden transformar los ácidos grasos en glúcidos. Los ácidos grasos no pueden transformarse en glucosa en las células animales, porque no existe tal vía metabólica, por lo que las reservas de grasa no sirven para mantener los niveles de glucosa en sangre. - Los ácidos grasos necesitan moléculas especiales para ser transportadas por la sangre desde las células del tejido adiposo (donde se almacenan), hasta las que necesitan energía, mientras la glucosa circula sin dificultad dada su solubilidad.")
37
Catabolismo de las proteínas

38
Visión de conjunto del catabolismo

Presentaciones similares

















: Autótrofos: A partir de la materia inorgánica y del CO2 de la atmósfera.>")


