Descargar la presentación
La descarga está en progreso. Por favor, espere

1
9 La membrana plasmática. Orgánulos membranosos

2
9 La membrana plasmática. Orgánulos membranosos
1.1. Composición de la membrana 1.2. Estructura. Modelo Singer-Nicholson 1972 1.3 Funciones de la membrana 1.4 Transporte a través de la membrana a) Transporte de moléculas pequeñas b) Transporte de macromoléculas 2. Retículo endolásmico 3. Aparato de Golgi 4. Lisosomas 5. Perroxisomas y glioxisomas 6. Mitocondrias 7. Cloroplastos y otros plastos
 Transporte de moléculas pequeñas. b) Transporte de macromoléculas. 2. Retículo endolásmico. 3. Aparato de Golgi. 4. Lisosomas. 5. Perroxisomas y glioxisomas. 6. Mitocondrias. 7. Cloroplastos y otros plastos.")
3
La membrana plasmática. Orgánulos membranosos
Esquema La membrana plasmática. Orgánulos membranosos 1. La membrana plasmática El retículo endoplasmático El transporte a través de la membrana El aparato de Golgi Las vacuolas Endocitosis Los lisosomas Uniones intercelulares Los peroxisomas y los glioxisomas Las mitocondrias Los cloroplastos

4
Recursos para la explicación de la unidad
La membrana plasmática El transporte a través de membrana Endocitosis Uniones intercelulares El retículo endoplasmático El aparato de Golgi Las vacuolas Los lisosomas Los peroxisomas y los glioxisomas Las mitocondrias Los cloroplastos

5
Estructura de la Membrana Celular
El grosor de la membrana es de 7.5 a 10 nanómetros (nm). No es visible en el microscopio óptico. Límite entre el medio extracelular e intracelular. Mitocondria Membrana plasmática Núcleo Membrana plasmática
. No es visible en el microscopio óptico. Límite entre el medio extracelular e intracelular. Mitocondria. Membrana plasmática. Núcleo. Membrana plasmática.")
6
Composición química La membrana se compone, casi completamente, de lípidos y proteínas, adicionalmente presenta colesterol y glúcidos.

7
Lípidos de Membrana Encontramos: Fosfolípidos Glucolípidos Colesterol
Cabeza Símbolo Colas Encontramos: Fosfolípidos Glucolípidos Colesterol Nota: La membrana plasmática de procariotas no presenta esteroles y el porcentaje de fosfolípidos es distinto. En arqueas en vez de ácidos grasos hay unidades repetitivas de isopreno (unidos por enlace éster a la glicerina).
.")
8
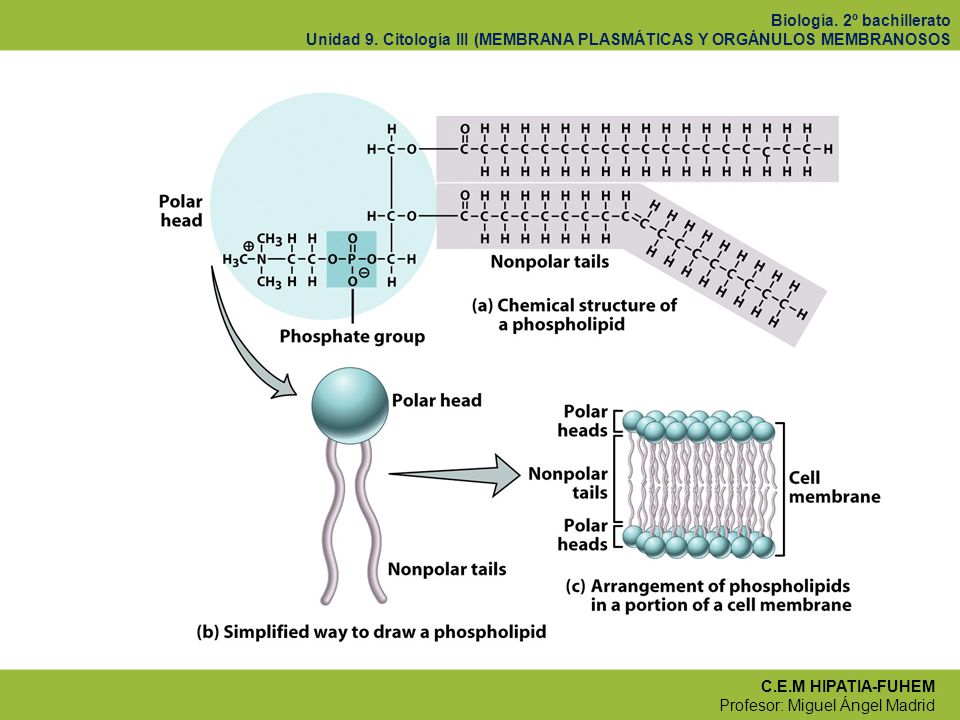
Lípidos de Membrana Cabeza Símbolo Colas Los fosfolípidos son el principal componente estructural de todas las membranas celulares. Carácter anfipático. Formación de micelas bicapas Distribución asimétrica Cabeza polar hidrofílica: (glicerol + fosfato + colina, o serina, etc. depende del tipo) Dos colas no polares: (dos ácidos grasos) que son hidrofóbicas o anfipáticas.
 Dos colas no polares: (dos ácidos grasos) que son hidrofóbicas o anfipáticas.")
11
Fosfolípidos mayoritarios en las membranas eucariotas
Hay cuatro tipos de fosfolípidos en la membrana celular: fosfatidilcolina esfingomielina (en este fosfolípido la glicerina ha sido sustituída por un aminoalcohol llamado D-4-esfingenina) fosfatidilserina fosfatidiletanolamina La zona media hidrófoba de la membrana es la que impide el paso de moléculas e iones hidrosulobles. Pero sin embargo sabemos que existen moléculas polares que atraviesan la membrana. Esta propiedad selectiva y especializada del transporte de membrana se deben al componente proteico de las mismas. Note that only phosphatidylserine carries a net negative charge, the importance of which we discuss later; the other three are electrically neutral at physiological pH, carrying one positive and one negative charge. Together these four phospholipids constitute more than half the mass of lipid in most membranes. Other phospholipids, such as the inositol phospholipids, are present in smaller quantities but are functionally very important. The inositol phospholipids, for example, have a crucial role in cell signaling.
 fosfatidilserina. fosfatidiletanolamina La zona media hidrófoba de la membrana es la que impide el paso de moléculas e iones hidrosulobles. Pero sin embargo sabemos que existen moléculas polares que atraviesan la membrana. Esta propiedad selectiva y especializada del transporte de membrana se deben al componente proteico de las mismas. Note that only phosphatidylserine carries a net negative charge, the importance of which we discuss later; the other three are electrically neutral at physiological pH, carrying one positive and one negative charge. Together these four phospholipids constitute more than half the mass of lipid in most membranes. Other phospholipids, such as the inositol phospholipids, are present in smaller quantities but are functionally very important. The inositol phospholipids, for example, have a crucial role in cell signaling.")
12
Colesterol Las moléculas de colesterol se encuentran intercaladas entre los fosfolípidos, y su función principal es la de regular la fluidez de la bicapa inmovilizando las colas hidrofóbicas próximas a la regiones polares. Es decir, endurece las membranas, reduce su fluidez y permeabilidad. Carácter anfipático. Distribución asimétrica

13
El colesterol amortigua la fluidez de la MP (= menos deformable)
La fluidez de la membrana depende de su composición. En este aspecto el colesterol actúa como un amortiguador de la fluidez. El colesterol es un constituyente fundamental en las membranas celulares. Está constituido por una cabeza polar, un núcleo esteroideo, el cual se dispone paralelo a las cadenas de ácidos grasos de los fosfolípidos de la membrana y una cola hidrocarbonada. Si nos fijamos en el esquema veremos como las moléculas de colesterol se encuentran orientadas en la bicapa de forma que los grupos hidroxilos se encuentran próximos a las cabezas polares de los fosfolípidos. En esta posición el núcleo de anillos esteroideos interacciona e inmoviliza parcialmente las cadenas de hidrocarbono más cercanas a las cabezas polares. Al disminuir la movilidad de estos primeros grupos de CH2 de las cadenas de fosfolípidos el colesterol hace que, en esta región la membrana sea menos deformable y disminuye la permeabilidad de la bicapa a las moléculas de agua. De este modo puede decirse que el colesterol actúa como una amortiguador de la fluidez de la membrana. Las membranas eucariotas presentan grandes cantidades de colesterol (hasta una molécula de colesterol por cada una de fosfolípido, Although cholesterol tends to make lipid bilayers less fluid, at the high concentrations found in most eucaryotic plasma membranes, it also prevents the hydrocarbon chains from coming together and crystallizing. In this way, it inhibits possible phase transitions.) Además de los fosfolípidos, las proteínas y el colesterol existe otro integrante de las membranas celulares, los glicolípidos. El colesterol amortigua la fluidez de la MP (= menos deformable) Disminuye la permeabilidad de la MP al agua
 Además de los fosfolípidos, las proteínas y el colesterol existe otro integrante de las membranas celulares, los glicolípidos. El colesterol amortigua la fluidez de la MP (= menos deformable) Disminuye la permeabilidad de la MP al agua.")
14
La fluidez de la membrana depende de:
Temperatura: Al aumentar la T aumenta la fluidez Naturaleza de los lípidos: Dobles enlaces: A mayor nº de dobles enlaces mayor fluidez (menor FVW). Longitud de la cadena: A mayor longitud de la cadena menor fluidez.Los lípidos insaturados y de cadena corta favorece el aumento de fluidez. Presencia de colesterol. Endurece las membranas reduciendo su fluidez y permeabilidad
. Longitud de la cadena: A mayor longitud de la cadena menor fluidez.Los lípidos insaturados y de cadena corta favorece el aumento de fluidez. Presencia de colesterol. Endurece las membranas reduciendo su fluidez y permeabilidad.")
15
Movimientos que pueden realizar los lípidos

16
Proteínas de membrana Determinan la función de la membrana biológica. Confieren a la membrana sus funciones específicas. Son características de cada especie. La mayoría globulares. Se mueven por difusión lateral. Distribución asimétrica

17
Muchas proteínas de membrana son glucoproteínas.
Tipos (por la forma en la que están dispuestas en la MP): - Periféricas o extrínsecas: incluidas de manera parcial en una de las superficies de la membrana, unidas covalentemente a lípidos (lipoproteínas) o asociadas a ellos. - Integrales o intrínsecas: Inmersas en la bicapa. Si atraviesan totalmente la membrana (pudiendo sobresalir) se denominan transmembrana. Aunque la estructura básica de la membrana celular sea la bicapa lipídica la mayoría de funciones específicas se llevan a cabo mediante la acción de proteínas. Su tipo y cantidad son muy variables en función del tipo celular. Como los lípidos, las proteínas de membrana a menudo se encuentran asociadas a cadenas de oligosacáridos (presentes en el exterior celular). De este modo la superficie celular presenta una cubierta rica en carbohidratos, el glicocálix, que veremos posteriormente. Existen diferentes formas de asociación entre la bicapa lipídica y las proteínas: Proteínas transmembrana (1,2,3,): Pueden atravesar completamente la bicapa, teniendo porciones de la proteína en contacto con el exterior y el interior celular. Al igual que sus vecinos los lípidos, estas proteínas transmembrana son moléculas anfipáticas (la región hidrofóbica se localiza en el interior de la membrana e interacciona con las colas hidrofóbicas de los lípidos). La hidrofobicidad de algunas de estas proteínas se aumenta por la formación de interacciones covalentes con cadenas de ácidos grasos que se insertan desde la monocapa citosólica (1). Proteínas localizadas en el citosol (4, 5, 7): ya sea por asociación con la monocapa lipídica por una hélice alfa anfipática (4) o por una o más uniones covalentes lipídicas por ejemplo por cadena de ácidos grasos (5) (son proteínas traducidas en citosol). Otras proteínas sólo se localizan en la superficie celular unidas mediante un oligosacárido por un puente covalente al fosfatidilinositol (6)(son proteinas traducidas en retículo endoplasmático). Otras proteínas se unen a la membrana mediante interacciones no covalentes con otras proteínas. Se les deomina proteínas periféricas de membrana (pueden liberarse de la membrana exponiendo a las células a medios hipo o hipertónicos o modificando el pH). El resto de proteínas transmembrana o ancladas a la bicapa mediante grupos lipídicos o con otras proteínas mediante inusuales uniones estrechas (pero que no pueden separarse por estos medios) se denominan proteínas integrales. El tipo de unión en la membrana refleja la función de la proteína. Solo las transmembrana actúan a ambos lados de la membrana o pueden transportar moléculas a su través. Los receptores de superficie son proteínas transmembrana que unen moléculas señal en el exterior celular y generan señales intracelulares. Las proteínas que sólo funcionan en uno de los lados normalmente se asocian con otras proteínas o lípidos que también actúan en ese lado. (esto es una tontería). La mayoría de las proteínas transmembrana atraviensan la bicapa como una alpha hélice simple (1), a hélice multiple (2) o como barriles β (3). Además las proteínas siempre tiene una única orientación en la membrana. Esta asimetría refleja el proceso de síntesis e inserción desde RE y las diferentes funciones de dominios extra e intracelulares. Los residuos proteicos que atraviesan la bicapa lipídica son aminoácidos con cadenas laterales no polares. Debido a que en enlace peptídico es polar y al ambiente hidrofóbico en que se encuentran estos enlaces peptídicos están dirigidos por puentes de hidrógeno, función de los cuales se optimiza al formarse la alpha hélice. Los barriles beta son más rígidos y cristalizan más rápido que las alpha hélices. El número de cadenas que atraviesan la membran puede oscilar de 8 a 22 y son abundantes en mitocondria, Cloroplastos y bacterias. Algunas forman poros llenos de agua que posibilitan el paso de sustancias hidrofílicas, otras actúan como receptores o incluso como enzimas (mayoritariamente restringidos a mitocondria, Cloroplastos y bacterias). En células eucariotas la mayoría de proteínas transmembrana son alpha hélice. The helices within these proteins can slide against each other, allowing the protein to undergo conformational changes that can be exploited to open and shut ion channels, transport solutes, or transduce extracellular signals into intracellular ones. In β barrel proteins, by contrast, each β strand is bound rigidly to its neighbors by hydrogen bonds, making conformational changes of the barrel itself unlikely. Al igual que los lípidos de membrana las proteínas pueden estar glicosiladas. De hecho, la gran mayoría de las proteínas en células animaleslo están. Al igual que ocurría en lípidos los oligosacátidos se añaden en el lumen de retículo y golgi. Por esta razón los oligos siempre se encuentran en la cara externa de la membrana. Otra diferencia que existe entre la cara citosólica y la externa en las proteínas de membrana es que el citosol es un ambiente reductor por lo que en la cara externa se formarán intra y entre proteínas puentes disulfuro entre la cys. Estos puentes son muy importantes en el mantenimiento de la conformación estructural o en la asociación con otros polipéptidos.
: - Periféricas o extrínsecas: incluidas de manera parcial en una de las superficies de la membrana, unidas covalentemente a lípidos (lipoproteínas) o asociadas a ellos. - Integrales o intrínsecas: Inmersas en la bicapa. Si atraviesan totalmente la membrana (pudiendo sobresalir) se denominan transmembrana. Aunque la estructura básica de la membrana celular sea la bicapa lipídica la mayoría de funciones específicas se llevan a cabo mediante la acción de proteínas. Su tipo y cantidad son muy variables en función del tipo celular. Como los lípidos, las proteínas de membrana a menudo se encuentran asociadas a cadenas de oligosacáridos (presentes en el exterior celular). De este modo la superficie celular presenta una cubierta rica en carbohidratos, el glicocálix, que veremos posteriormente. Existen diferentes formas de asociación entre la bicapa lipídica y las proteínas: Proteínas transmembrana (1,2,3,): Pueden atravesar completamente la bicapa, teniendo porciones de la proteína en contacto con el exterior y el interior celular. Al igual que sus vecinos los lípidos, estas proteínas transmembrana son moléculas anfipáticas (la región hidrofóbica se localiza en el interior de la membrana e interacciona con las colas hidrofóbicas de los lípidos). La hidrofobicidad de algunas de estas proteínas se aumenta por la formación de interacciones covalentes con cadenas de ácidos grasos que se insertan desde la monocapa citosólica (1). Proteínas localizadas en el citosol (4, 5, 7): ya sea por asociación con la monocapa lipídica por una hélice alfa anfipática (4) o por una o más uniones covalentes lipídicas por ejemplo por cadena de ácidos grasos (5) (son proteínas traducidas en citosol). Otras proteínas sólo se localizan en la superficie celular unidas mediante un oligosacárido por un puente covalente al fosfatidilinositol (6)(son proteinas traducidas en retículo endoplasmático). Otras proteínas se unen a la membrana mediante interacciones no covalentes con otras proteínas. Se les deomina proteínas periféricas de membrana (pueden liberarse de la membrana exponiendo a las células a medios hipo o hipertónicos o modificando el pH). El resto de proteínas transmembrana o ancladas a la bicapa mediante grupos lipídicos o con otras proteínas mediante inusuales uniones estrechas (pero que no pueden separarse por estos medios) se denominan proteínas integrales. El tipo de unión en la membrana refleja la función de la proteína. Solo las transmembrana actúan a ambos lados de la membrana o pueden transportar moléculas a su través. Los receptores de superficie son proteínas transmembrana que unen moléculas señal en el exterior celular y generan señales intracelulares. Las proteínas que sólo funcionan en uno de los lados normalmente se asocian con otras proteínas o lípidos que también actúan en ese lado. (esto es una tontería). La mayoría de las proteínas transmembrana atraviensan la bicapa como una alpha hélice simple (1), a hélice multiple (2) o como barriles β (3). Además las proteínas siempre tiene una única orientación en la membrana. Esta asimetría refleja el proceso de síntesis e inserción desde RE y las diferentes funciones de dominios extra e intracelulares. Los residuos proteicos que atraviesan la bicapa lipídica son aminoácidos con cadenas laterales no polares. Debido a que en enlace peptídico es polar y al ambiente hidrofóbico en que se encuentran estos enlaces peptídicos están dirigidos por puentes de hidrógeno, función de los cuales se optimiza al formarse la alpha hélice. Los barriles beta son más rígidos y cristalizan más rápido que las alpha hélices. El número de cadenas que atraviesan la membran puede oscilar de 8 a 22 y son abundantes en mitocondria, Cloroplastos y bacterias. Algunas forman poros llenos de agua que posibilitan el paso de sustancias hidrofílicas, otras actúan como receptores o incluso como enzimas (mayoritariamente restringidos a mitocondria, Cloroplastos y bacterias). En células eucariotas la mayoría de proteínas transmembrana son alpha hélice. The helices within these proteins can slide against each other, allowing the protein to undergo conformational changes that can be exploited to open and shut ion channels, transport solutes, or transduce extracellular signals into intracellular ones. In β barrel proteins, by contrast, each β strand is bound rigidly to its neighbors by hydrogen bonds, making conformational changes of the barrel itself unlikely. Al igual que los lípidos de membrana las proteínas pueden estar glicosiladas. De hecho, la gran mayoría de las proteínas en células animaleslo están. Al igual que ocurría en lípidos los oligosacátidos se añaden en el lumen de retículo y golgi. Por esta razón los oligos siempre se encuentran en la cara externa de la membrana. Otra diferencia que existe entre la cara citosólica y la externa en las proteínas de membrana es que el citosol es un ambiente reductor por lo que en la cara externa se formarán intra y entre proteínas puentes disulfuro entre la cys. Estos puentes son muy importantes en el mantenimiento de la conformación estructural o en la asociación con otros polipéptidos.")
18
Las proteínas de la membrana pueden actuar: Enzimas: catalizan
Túnel: permiten el paso de sustancias. Bombas: Transportan iones a ambos lados de la membrana (Na+., K+ …) Receptores de mensajeros químicos. Aunque la estructura básica de la membrana celular sea la bicapa lipídica la mayoría de funciones específicas se llevan a cabo mediante la acción de proteínas. Su tipo y cantidad son muy variables en función del tipo celular. Como los lípidos, las proteínas de membrana a menudo se encuentran asociadas a cadenas de oligosacáridos (presentes en el exterior celular). De este modo la superficie celular presenta una cubierta rica en carbohidratos, el glicocálix, que veremos posteriormente. Existen diferentes formas de asociación entre la bicapa lipídica y las proteínas: Proteínas transmembrana (1,2,3,): Pueden atravesar completamente la bicapa, teniendo porciones de la proteína en contacto con el exterior y el interior celular. Al igual que sus vecinos los lípidos, estas proteínas transmembrana son moléculas anfipáticas (la región hidrofóbica se localiza en el interior de la membrana e interacciona con las colas hidrofóbicas de los lípidos). La hidrofobicidad de algunas de estas proteínas se aumenta por la formación de interacciones covalentes con cadenas de ácidos grasos que se insertan desde la monocapa citosólica (1). Proteínas localizadas en el citosol (4, 5, 7): ya sea por asociación con la monocapa lipídica por una hélice alfa anfipática (4) o por una o más uniones covalentes lipídicas por ejemplo por cadena de ácidos grasos (5) (son proteínas traducidas en citosol). Otras proteínas sólo se localizan en la superficie celular unidas mediante un oligosacárido por un puente covalente al fosfatidilinositol (6)(son proteinas traducidas en retículo endoplasmático). Otras proteínas se unen a la membrana mediante interacciones no covalentes con otras proteínas. Se les deomina proteínas periféricas de membrana (pueden liberarse de la membrana exponiendo a las células a medios hipo o hipertónicos o modificando el pH). El resto de proteínas transmembrana o ancladas a la bicapa mediante grupos lipídicos o con otras proteínas mediante inusuales uniones estrechas (pero que no pueden separarse por estos medios) se denominan proteínas integrales. El tipo de unión en la membrana refleja la función de la proteína. Solo las transmembrana actúan a ambos lados de la membrana o pueden transportar moléculas a su través. Los receptores de superficie son proteínas transmembrana que unen moléculas señal en el exterior celular y generan señales intracelulares. Las proteínas que sólo funcionan en uno de los lados normalmente se asocian con otras proteínas o lípidos que también actúan en ese lado. (esto es una tontería). La mayoría de las proteínas transmembrana atraviensan la bicapa como una alpha hélice simple (1), a hélice multiple (2) o como barriles β (3). Además las proteínas siempre tiene una única orientación en la membrana. Esta asimetría refleja el proceso de síntesis e inserción desde RE y las diferentes funciones de dominios extra e intracelulares. Los residuos proteicos que atraviesan la bicapa lipídica son aminoácidos con cadenas laterales no polares. Debido a que en enlace peptídico es polar y al ambiente hidrofóbico en que se encuentran estos enlaces peptídicos están dirigidos por puentes de hidrógeno, función de los cuales se optimiza al formarse la alpha hélice. Los barriles beta son más rígidos y cristalizan más rápido que las alpha hélices. El número de cadenas que atraviesan la membran puede oscilar de 8 a 22 y son abundantes en mitocondria, Cloroplastos y bacterias. Algunas forman poros llenos de agua que posibilitan el paso de sustancias hidrofílicas, otras actúan como receptores o incluso como enzimas (mayoritariamente restringidos a mitocondria, Cloroplastos y bacterias). En células eucariotas la mayoría de proteínas transmembrana son alpha hélice. The helices within these proteins can slide against each other, allowing the protein to undergo conformational changes that can be exploited to open and shut ion channels, transport solutes, or transduce extracellular signals into intracellular ones. In β barrel proteins, by contrast, each β strand is bound rigidly to its neighbors by hydrogen bonds, making conformational changes of the barrel itself unlikely. Al igual que los lípidos de membrana las proteínas pueden estar glicosiladas. De hecho, la gran mayoría de las proteínas en células animaleslo están. Al igual que ocurría en lípidos los oligosacátidos se añaden en el lumen de retículo y golgi. Por esta razón los oligos siempre se encuentran en la cara externa de la membrana. Otra diferencia que existe entre la cara citosólica y la externa en las proteínas de membrana es que el citosol es un ambiente reductor por lo que en la cara externa se formarán intra y entre proteínas puentes disulfuro entre la cys. Estos puentes son muy importantes en el mantenimiento de la conformación estructural o en la asociación con otros polipéptidos.
 Receptores de mensajeros químicos. Aunque la estructura básica de la membrana celular sea la bicapa lipídica la mayoría de funciones específicas se llevan a cabo mediante la acción de proteínas. Su tipo y cantidad son muy variables en función del tipo celular. Como los lípidos, las proteínas de membrana a menudo se encuentran asociadas a cadenas de oligosacáridos (presentes en el exterior celular). De este modo la superficie celular presenta una cubierta rica en carbohidratos, el glicocálix, que veremos posteriormente. Existen diferentes formas de asociación entre la bicapa lipídica y las proteínas: Proteínas transmembrana (1,2,3,): Pueden atravesar completamente la bicapa, teniendo porciones de la proteína en contacto con el exterior y el interior celular. Al igual que sus vecinos los lípidos, estas proteínas transmembrana son moléculas anfipáticas (la región hidrofóbica se localiza en el interior de la membrana e interacciona con las colas hidrofóbicas de los lípidos). La hidrofobicidad de algunas de estas proteínas se aumenta por la formación de interacciones covalentes con cadenas de ácidos grasos que se insertan desde la monocapa citosólica (1). Proteínas localizadas en el citosol (4, 5, 7): ya sea por asociación con la monocapa lipídica por una hélice alfa anfipática (4) o por una o más uniones covalentes lipídicas por ejemplo por cadena de ácidos grasos (5) (son proteínas traducidas en citosol). Otras proteínas sólo se localizan en la superficie celular unidas mediante un oligosacárido por un puente covalente al fosfatidilinositol (6)(son proteinas traducidas en retículo endoplasmático). Otras proteínas se unen a la membrana mediante interacciones no covalentes con otras proteínas. Se les deomina proteínas periféricas de membrana (pueden liberarse de la membrana exponiendo a las células a medios hipo o hipertónicos o modificando el pH). El resto de proteínas transmembrana o ancladas a la bicapa mediante grupos lipídicos o con otras proteínas mediante inusuales uniones estrechas (pero que no pueden separarse por estos medios) se denominan proteínas integrales. El tipo de unión en la membrana refleja la función de la proteína. Solo las transmembrana actúan a ambos lados de la membrana o pueden transportar moléculas a su través. Los receptores de superficie son proteínas transmembrana que unen moléculas señal en el exterior celular y generan señales intracelulares. Las proteínas que sólo funcionan en uno de los lados normalmente se asocian con otras proteínas o lípidos que también actúan en ese lado. (esto es una tontería). La mayoría de las proteínas transmembrana atraviensan la bicapa como una alpha hélice simple (1), a hélice multiple (2) o como barriles β (3). Además las proteínas siempre tiene una única orientación en la membrana. Esta asimetría refleja el proceso de síntesis e inserción desde RE y las diferentes funciones de dominios extra e intracelulares. Los residuos proteicos que atraviesan la bicapa lipídica son aminoácidos con cadenas laterales no polares. Debido a que en enlace peptídico es polar y al ambiente hidrofóbico en que se encuentran estos enlaces peptídicos están dirigidos por puentes de hidrógeno, función de los cuales se optimiza al formarse la alpha hélice. Los barriles beta son más rígidos y cristalizan más rápido que las alpha hélices. El número de cadenas que atraviesan la membran puede oscilar de 8 a 22 y son abundantes en mitocondria, Cloroplastos y bacterias. Algunas forman poros llenos de agua que posibilitan el paso de sustancias hidrofílicas, otras actúan como receptores o incluso como enzimas (mayoritariamente restringidos a mitocondria, Cloroplastos y bacterias). En células eucariotas la mayoría de proteínas transmembrana son alpha hélice. The helices within these proteins can slide against each other, allowing the protein to undergo conformational changes that can be exploited to open and shut ion channels, transport solutes, or transduce extracellular signals into intracellular ones. In β barrel proteins, by contrast, each β strand is bound rigidly to its neighbors by hydrogen bonds, making conformational changes of the barrel itself unlikely. Al igual que los lípidos de membrana las proteínas pueden estar glicosiladas. De hecho, la gran mayoría de las proteínas en células animaleslo están. Al igual que ocurría en lípidos los oligosacátidos se añaden en el lumen de retículo y golgi. Por esta razón los oligos siempre se encuentran en la cara externa de la membrana. Otra diferencia que existe entre la cara citosólica y la externa en las proteínas de membrana es que el citosol es un ambiente reductor por lo que en la cara externa se formarán intra y entre proteínas puentes disulfuro entre la cys. Estos puentes son muy importantes en el mantenimiento de la conformación estructural o en la asociación con otros polipéptidos.")
19
Proteínas de Membrana

20
Glúcidos En forma de glucolípidos y glucoproteínas.
Localizados en cara externa. Constituyen el glucocálix (solo en células animales) Distribución asimétrica
 Distribución asimétrica.")
21
Funciones glucocálix Protección contra daños químicos y/o físicos.
Interviene en fenómenos de reconocimiento celular. Confiere viscosidad a la membrana. Interviene en la comunicación intracelular. Presenta propiedades inmunitarias (ejemplo el glucocálix de los eritrocitos representan los antígenos de los grupos sanguíneos). Contribuye al reconocimiento y fijación de determinadas sustancias que la célula incorpora mediante fagocitosis y pinocitosis.
. Contribuye al reconocimiento y fijación de determinadas sustancias que la célula incorpora mediante fagocitosis y pinocitosis.")
22
Membrana plasmática La mayoría de las membranas celulares constituyen un “mosaico fluido” de fosfolípidos y proteínas. Modelo Singer-Nicholson 1972 Los componentes proteicos de la membrana pueden dividirse en proteínas periféricas y proteínas integrales. Debido a que la membrana celular no es sólida y en su interior, fosfolípidos y proteínas pueden desplazarse lateralmente. Ello origina que las proteínas no presenten una distribución uniforme, sino que la distribución de las mismas presenta un patrón en mosaico que cambia constantemente la que se ha denominado modelo de mosaico fluido. Aunque la mayoría de lípidos y proteínas pueden moverse libremente por en el plano de la membrana el intercambio entre las monocapas es poco frecuente (es poco probable que una porción hidrofílica grande cambie de monocapa si tiene que ser arrastrada a través del interior no polar de la bicapa lipídica). Existen excepciones en los que los componentes de la membrana no pueden difundir libremente, por ejemplo, el secuestro de los receptores de acetilcolina (proteína integral de membrana) en la placa motora terminal de músculo esquelético. En este tipo de distribución el citoesqueleto parece fijar ciertas proteínas de membrana. Pero no sólo las proteínas desempeñan funciones celulares importantes. Determinados fosfolípidos presentes en diminutas proporciones en la membrana plasmática tienen importantes funciones en la transducción de señales. El bifosfato de fosfatidilinositol, cuando se hidroliza por una fosfolipasa C activada por un receptor, libera inositol 1,4,5-trifosfato (IP3) y diacilglicerol. El IP3 se libera en el citosol, donde actúa sobre receptores del retículo endoplásmico para provocar la liberación de Ca2+, lo que afecta a muchos procesos celulares. El diacilglicerol permanece en la membrana, donde junto con el Ca2+ interviene en la activación de la proteinkinasa C, una proteína muy importante en la transducción de señales. La fluidez de la membrana depende de su composición. En este aspecto el colesterol actúa como un amortiguador de la fluidez.
. Existen excepciones en los que los componentes de la membrana no pueden difundir libremente, por ejemplo, el secuestro de los receptores de acetilcolina (proteína integral de membrana) en la placa motora terminal de músculo esquelético. En este tipo de distribución el citoesqueleto parece fijar ciertas proteínas de membrana. Pero no sólo las proteínas desempeñan funciones celulares importantes. Determinados fosfolípidos presentes en diminutas proporciones en la membrana plasmática tienen importantes funciones en la transducción de señales. El bifosfato de fosfatidilinositol, cuando se hidroliza por una fosfolipasa C activada por un receptor, libera inositol 1,4,5-trifosfato (IP3) y diacilglicerol. El IP3 se libera en el citosol, donde actúa sobre receptores del retículo endoplásmico para provocar la liberación de Ca2+, lo que afecta a muchos procesos celulares. El diacilglicerol permanece en la membrana, donde junto con el Ca2+ interviene en la activación de la proteinkinasa C, una proteína muy importante en la transducción de señales. La fluidez de la membrana depende de su composición. En este aspecto el colesterol actúa como un amortiguador de la fluidez.")
23
La membrana celular

24
Proteína transmembranosa
La membrana plasmática Proteína integral Proteína transmembranosa Fosfolípido Glicolípido Glucoproteína Proteína periférica Colesterol Líquido extracelular Citosol

25
Funciones de la Membrana Plasmática
Protegen la célula o a la organelas del medio externo. Mantienen una forma estable de la célula u organela. Regulan el transporte de sustancias y energía hacia adentro o hacia afuera de la célula u orgánulo Permite la comunicación entre las células adyacentes. Permiten el reconocimiento celular. Permiten la motilidad de algunas células u orgánulos

26
La membrana plasmática
Microvellosidades Membrana

27
MECANISMO DE TRANSPORTE
Transporte de moléculas de bajo peso molecular Transporte de moléculas de elevada masa molecular T. pasivo T. activo Exocitosis Endocitosis Fagocitosis Pinocitosis Mediada por receptor Difusión simple Difusión facilitada Bomba sodio-potasio

28
El transporte de moléculas de bajo peso molecular
Puede ser de dos tipos: Transporte activo Transporte pasivo

29
Transporte Pasivo No requiere el consumo de energía (ATP).
A favor de gradiente. El movimiento ocurre por diferencias en la concentración y en las cargas eléctricas de las sustancias en ambos lados de la membrana. Tenemos los siguientes mecanismos: Difusión simple Ósmosis Difusión facilitada EQUILIBRIO Moléculas de colorante Membrana

30
Difusión Simple Las moléculas atraviesan directamente la mb o por canales membranosos (proteínas en canal) El movimiento de moléculas se da a través de la membrana de fosfolípidos, de un gradiente de alta concentración a baja concentración. Cuando mayor es el gradiente de concentración, más rápida es la velocidad de difusión. Moléculas solubles en lípidos como etanol, y moléculas pequeñas como H2O, N2, CO2 y O2. Citoplasma Exterior de la Célula O2 CO2 Mayor concentración Menor concentración

31
(Proteínas Transmembrana) H. esteroideas, Éter, cloroformo
DIFUSIÓN SIMPLE A través de la bicapa Por canales (Proteínas Transmembrana) H. esteroideas, Éter, cloroformo Apolares: O2, N2, H2O, CO2 Na+, K+, Ca2+, Cl- Difusión por ligando y receptor
 H. esteroideas, Éter, cloroformo. Apolares: O2, N2, H2O, CO2. Na+, K+, Ca2+, Cl- Difusión por ligando y receptor.")
32
Difusión Facilitada Algunas moléculas por su tamaño o carga no difunden libremente a través de la membrana. Se efectúa mediante proteínas transportadoras (permeasas o carrier transmmembranas) capaces de trasladar moléculas polares o iones. Dichas proteínas sufren cambios conformacionales Estos canales son usados para la glucosa, aminoácidos y para iones pequeños y con carga tales como K+, Na+, Cl-.
 capaces de trasladar moléculas polares o iones. Dichas proteínas sufren cambios conformacionales. Estos canales son usados para la glucosa, aminoácidos y para iones pequeños y con carga tales como K+, Na+, Cl-.")
33
RESUMIENDO: El transporte pasivo
Difusión simple Difusión por variación de potencial eléctrico Membrana polarizada Membrana despolarizada Difusión facilitada por permeasa Difusión por ligando Ligando Permeasa

34
Transporte Activo Las células utilizan energía (ATP) durante el transporte. Se realiza en contra gradiente químico y eléctrico (electroquímico) La sustancia transportada se une a la proteína transportadora y es liberada en el interior de la célula sin sufrir modificación.

35
Bomba de Sodio (Na) y Potasio (K)
Es una proteína presente en todas las membranas plasmáticas de las células animales, cuyo objetivo es eliminar sodio de la célula e introducir potasio en el citoplasma.

36
Funciones de la Bomba de Sodio (Na) y Potasio (K)
Mantenimiento de la osmolaridad y del volumen celular Mantiene un potencial eléctrico de membrana Favorece la trasmisión de impulsos nerviosos Mantenimiento de los gradientes de sodio y potasio

37
Bomba de sodio-potasio
RESUMIENDO: El transporte activo ATP Bomba de sodio-potasio Na+ ADP + Pi K+ Se produce un cambio conformacional y se bombean dos iones de potasio hacia el interior. Se produce un cambio conformacional de la proteína y se bombean tres iones de sodio hacia el exterior.

38
Transporte de macromoléculas
Endocitosis Exocitosis Se basan en la formación de vesículas membranosas.

39
Exocitosis Una vesícula membranosa se desplaza hasta la membrana, se fusiona con la membrana y el contenido se vacía fuera de la célula. Se consigue expulsan fuera residuos del metabolismo… Fluido celular externo Citoplasma

40
Exocitosis Organismos unicelulares por ejemplo desechan sus residuos metabólicos mediante la formación de vesículas que expulsan al exterior

41
Tipos de Exocitosis Secreción Constitutiva
Reponer membrana o proteínas Secreción Reguladora Secreción de enzimas u hormonas

42
Endocitosis Mediante la formación de vesículas o vacuolas a partir de la membrana plasmática la célula incorpora macromoléculas u otras partículas. Se induce la formación de un sistema reticular de clatrina (proteína filamentosa), posteriormente un relieve membranoso, para formar la vesícula. Así, por ejemplo, se capta colesterol. Tipos: Fagocitosis, Pinocitosis y Endocitosis mediada por receptores. Citoplasma Líquido intersticial Vesícula Membrana Plasmática
, posteriormente un relieve membranoso, para formar la vesícula. Así, por ejemplo, se capta colesterol. Tipos: Fagocitosis, Pinocitosis y Endocitosis mediada por receptores. Citoplasma. Líquido intersticial. Vesícula. Membrana Plasmática.")
43
Tipos de Endocitosis: Fagocitosis
La membrana plasmática forma prolongaciones celulares (pseudópodos) que envuelven la partícula sólida, englobándola en una vesícula (fagosoma). Luego, uno o varios lisosomas se fusionan con la vacuola y vacían sus enzimas hidrolíticas en el interior de la vacuola. Pseudópodo Alimento a ser ingerido FAGOCITOSIS
 que envuelven la partícula sólida, englobándola en una vesícula (fagosoma). Luego, uno o varios lisosomas se fusionan con la vacuola y vacían sus enzimas hidrolíticas en el interior de la vacuola. Pseudópodo. Alimento a ser ingerido. FAGOCITOSIS.")
44
Pinocitosis Membrana celular PINOCITOSIS La membrana celular se invagina, formando una vesícula alrededor del líquido del medio externo que será incorporado a la célula. Luego se libera en el citoplasma.

45
Endocitosis mediada por receptor
Las sustancias que serán transportadas al interior deben primero acoplarse a las moléculas receptoras específicas. concentrados en zonas particulares de la membrana (depresiones). Cuando los receptores están unidos con sus moléculas especificas, se ahuecan y se cierran formando una vesícula. Material unido a las proteínas receptoras ENDOCITOSIS MEDIADA POR RECEPTORES Membrana celular CAVIDAD citoplasma
. Cuando los receptores están unidos con sus moléculas especificas, se ahuecan y se cierran formando una vesícula. Material unido a las proteínas receptoras. ENDOCITOSIS MEDIADA. POR RECEPTORES. Membrana celular. CAVIDAD. citoplasma.")
46
Endocitosis por receptor
Clatrina Pinocitosis receptor Clatrina ligando Complejo receptor-ligando Vesícula pinocítica Fagocitosis Endocitosis por receptor Clatrina Fagosoma Vesícula endocítica

47
Repaso

49
Uniones intercelulares
Son regiones especializadas de la membrana plasmática en las que se concentran proteínas transmembranas especiales, mediante las cuales se establecen conexiones entre dos células o entre una célula y la matriz extracelular. Aparecen en todos los tejidos, pero son abundantes en los epiteliales. Por su función se clasifican en: Uniones íntimas o de oclusión. Uniones adherentes o desmosomas Uniones de comunicación tipo gap

51
UNIONES OCLUYENTES O ÍNTIMAS
Uniones intercelulares UNIONES OCLUYENTES O ÍNTIMAS Aparecen en la región lateral de células epiteliales que separan medios de composición diferentes (pared vasos s, pared vejiga…). Permiten cerrar el espacio intercelular (por tanto no dejan espacio) entre las células impidiendo el paso de moléculas entre ellas. Se forman de proteínas transmembrana dispuestas en hilera que sueldan membranas.
. Permiten cerrar el espacio intercelular (por tanto no dejan espacio) entre las células impidiendo el paso de moléculas entre ellas. Se forman de proteínas transmembrana dispuestas en hilera que sueldan membranas.")
52
Uniones intercelulares
Unión íntima Desmosoma Unión tipo GAP Canal Proteína transmembranosa Espacio intercelular Proteínas transmembranosas Proteínas transmembranosas Placa Filamentos de queratina

53
Uniones intercelulares
Unión íntima Desmosoma Unión tipo GAP Canal Proteína transmembranosa Canal Espacio intercelular Proteínas transmembranosas Proteínas transmembranosas Placa Filamentos de queratina Proteína transmembranosa

54
DESMOSOMAS O UNIONES ADHERENTES
Uniones intercelulares DESMOSOMAS O UNIONES ADHERENTES Uniones de adherencia entre las células. Unen células pero no impiden el paso de sustancias.. Función: aumentan la resistencia de las células frente a tensiones mecánicas fuertes.

55
UNIONES DE COMUNICACIÓN TIPO GAP
Uniones intercelulares UNIONES DE COMUNICACIÓN TIPO GAP Uniones que no dejan espacio intercelular, pero si un pequeño espacio de comunicación entre citoplasmas. Función: establecer puntos de comunicación directa entre el citoplasma de células adyacentes. A través de ellos pasan iones y pequeñas moléculas. Las proteínas transmembrana llamadas conexinas forman canales denominados conexones, que las atraviesan

56
Sacos aplanados o cisternas, túbulos sinuosos
El retículo endoplasmático Estructura: Sacos aplanados o cisternas, túbulos sinuosos Localización: Células eucariotas Tipos: REr, REl Retículo endoplasmático liso (REl) Retículo endoplasmático rugoso (REr)
 Retículo endoplasmático rugoso (REr)")
57
El retículo endoplasmático
VER SÍNTESIS DE PROTEÍNAS El retículo endoplasmático Ribosomas Retículo endoplasmático rugoso (REr) Retículo endoplasmático liso (REl)
 Retículo endoplasmático liso (REl)")
58
RErugoso Estructura: Ribosomas unidos por sub mayor (riboforinas)
Sacos aplanados o cisternas. Ribosomas unidos por sub mayor (riboforinas)
")
59
El retículo endoplasmático
REr: Muy desarrollado en aquellas células que participan muy activamente en la síntesis de proteínas Ribosomas Retículo endoplasmático rugoso (REr) Retículo endoplasmático liso (REl)
 Retículo endoplasmático liso (REl)")
60
Retículo endoplasmático

61
Retículo endoplasmático

62
El retículo endoplasmático
ARN mensajero 1l Ribosoma 2l Citosol Retículo endoplasmático rugoso 5l 4l Lumen 3l Péptido de señalización Proteína

63
Túbulos interconectados entre ellos y con REr.
El retículo endoplasmático liso Estructura: Túbulos interconectados entre ellos y con REr. Muy desarrollado en: Células musculares estriadas. (retículo sarcoplásmico). Células interticiales de ovarios y testículos. Hepatocitos Retículo endoplasmático liso (REl)
. Células interticiales de ovarios y testículos. Hepatocitos. Retículo endoplasmático liso (REl)")
64
Retículo endoplasmático liso (REl)
El retículo endoplasmático liso Funciones: Síntesis de lípidos de mb (fosfolípidos, glicolípidos, colesterol). Almacén de lípidos. Transporte de lípidos. Detoxificación de sustancias. Contracción muscular. Retículo endoplasmático liso (REl)
. Almacén de lípidos. Transporte de lípidos. Detoxificación de sustancias. Contracción muscular. Retículo endoplasmático liso (REl)")
65
RETICULO ENDOPLASMATICO Y APARATO DE GOLGI

66
Aparato de Golgi Cara cis Cara trans

67
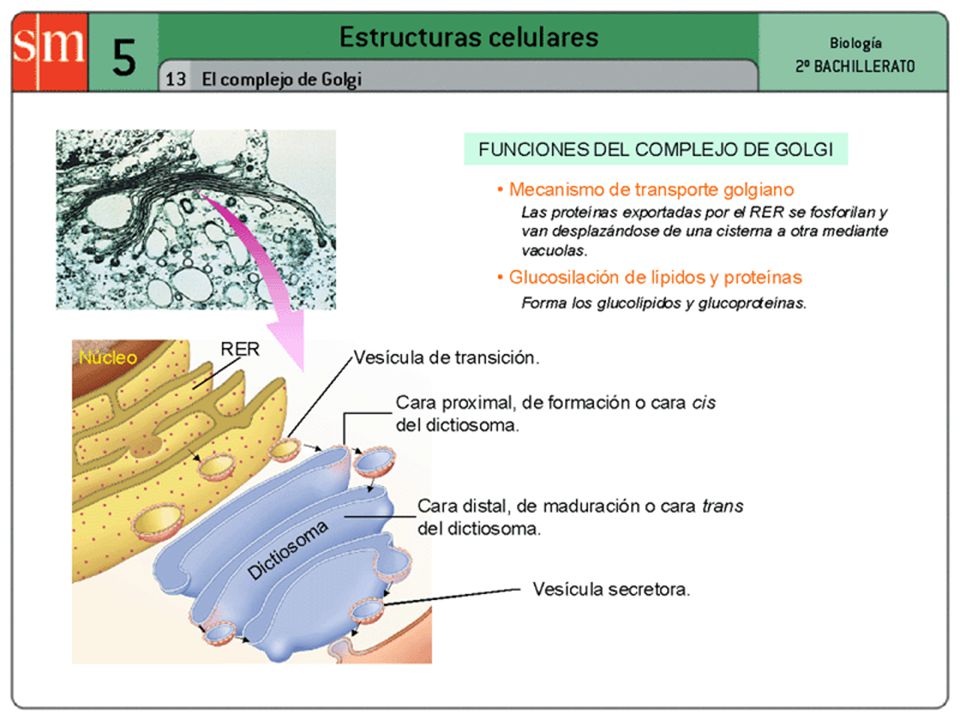
El aparato de Golgi Vesículas de secrección
CARA CIS, PROXIMAL O FORMACIÓN 1. Las vesículas de transición, procedentes de la envoltura nuclear y del retículo endoplasmático, se unen a la cara cis del dictiosoma. 2. El contenido molecular se incorpora al dictiosoma. 3. Las vesículas intercisternas pasan el contenido de cisterna a cisterna, y al llegar a la cara trans, se concentra y se acumula en el interior de las vesículas. 4. Las vesículas de secreción se dirigen hacia la membrana plasmática, se fusionan con ella y vierten su contenido al medio externo. 5. La superficie de las vesículas que se forman están revestidas de clatrina. Este revestimiento se pierde una vez formada la vesícula. 3 1 2 4 5 Vesículas de transición CARA TRANS, DISTAL O MADURACIÓN

68
El aparato de Golgi

70
El aparato de Golgi Funciones:
Transporte de sustancias. (formación de vesículas) Maduración. Acumulación y secreción de proteínas. Glucosidación de lípidos y proteínas. Síntesis de polisacáridos (celulosa, proteoglucanos de matriz). Formación del tabique telofásico en células vegetales.
 Maduración. Acumulación y secreción de proteínas. Glucosidación de lípidos y proteínas. Síntesis de polisacáridos (celulosa, proteoglucanos de matriz). Formación del tabique telofásico en células vegetales.")
71
Vacuola en célula vegetal
Las vacuolas Vacuola en célula vegetal

72
¿Quiénes las presentan?
Las vacuolas ¿Qué son? Vesículas membranosas con interior acuoso Origen REr, A.G e invaginaciones de membrana Tonoplasto ¿Quiénes las presentan? Células animales (vesículas) Células vegetales Funciones Mantenimiento turgencia celular. Digestión celular (en vegetales) Almacenamiento de sustancias diversas (pétalos, alcaloides, cristales…)
 Células vegetales. Funciones. Mantenimiento turgencia celular. Digestión celular (en vegetales) Almacenamiento de sustancias diversas (pétalos, alcaloides, cristales…)")
75
LISOSOMAS Orgánulos esféricos con membrana simple
son Aparato de Golgi Orgánulos esféricos con membrana simple contienen Bomba De protones por HIDROLASAS ÁCIDAS (pH = 4’6) RER (más de 50 enzimas diferentes) son ES EL SISTEMA DIGESTIVO CELULAR
 RER. (más de 50 enzimas diferentes) son. ES EL SISTEMA DIGESTIVO CELULAR.")
76
Los lisosomas Origen Aparato Golgi
Las enzimas se forman en R.E y pasan al Golgi FUNCIÓN DIGESTIÓN CELULAR (extracelular e intracelular)
")
77
LISOSOMAS SECUNDARIOS PRIMARIOS Contenido homogéneo
Aún sin unirse a vacuolas de pinocitosis o fagocitosis Contenido heterogéneo. - Vacuolas digestivas (Heterofagosoma) - Vacuolas autofágicas (Autofagosoma) - Cuerpos residuales
 - Vacuolas autofágicas. (Autofagosoma) - Cuerpos residuales.")
78
¿Por qué un lisosoma no se digiere a si mismo?
Porque las proteínas de la cara interna de la membrana lisosómica están altamente glucosiladas, los cual les sirve de protección frente a sus propias enzimas y la acidez del medio

81
Los lisosomas Autofagia Lisosoma secundario Lisosoma primario
Heterofagia Lisosoma secundario

83
Peroxisomas Proceden del R.E Contienen enzimas oxidasas y catalasas
Abundantes en las células de riñón e hígado

84
Actividad oxidativa de los peroxisomas
Los peroxisomas y los glioxisomas Actividad oxidativa de los peroxisomas Oxidasa Catalasa Sustrato (aa, a.g, á úrico) H2O2 H2O + ½ O2 Peroxisoma Las oxidasas son responsables de la oxidación de diferentes sustancias (aminoácidos, ácidos grasos, ácido úrico…), como producto se origina agua oxigenada (altamente tóxico), que es descompuesta por la catalasa en el mismo orgánulo.
 H2O2. H2O + ½ O2. Peroxisoma. Las oxidasas son responsables de la oxidación de diferentes sustancias (aminoácidos, ácidos grasos, ácido úrico…), como producto se origina agua oxigenada (altamente tóxico), que es descompuesta por la catalasa en el mismo orgánulo.")
85
Los peroxisomas y los glioxisomas
En las células de las semillas en germinación los peroxisomas son los responsables del ciclo del glioxilato (conversión de ácidos grasos a glúcidos, que proporcionan energía para la geminación y el crecimiento Glioxisomas

86
Mitocondria

87
Las mitocondrias Distribución: uniforme por todo el citoplasma. (su nº depende del tipo de célula)
")
88
Las mitocondrias Condrioma: conjunto de mitocondrias de una célula
ADN mitocondrial Circular bicatenario y diferente al ADN nuclear Condrioma: conjunto de mitocondrias de una célula Matriz mitocondrial Espacio intermembranoso. Contenido similar al citosol Enzimas (duplicación, transcricpicón, traducción, ciclo Krebs, beta oxidación) Cresta mitocondrial. Se disponen transversalmente al eje mitocondrial Membrana externa. Similar resto de membranas. Con porinas, da permeablidad Membrana interna. Presenta crestas mitocondriales Mitorribosomas (70 S = )
 Cresta mitocondrial. Se disponen transversalmente al eje mitocondrial. Membrana externa. Similar resto de membranas. Con porinas, da permeablidad. Membrana interna. Presenta crestas mitocondriales. Mitorribosomas. (70 S = )")
89
Espacio intermembranoso
Las ATP asas se encuentran en la cara externa de las crestas, orientadas hacia la matriz Las mitocondrias ADN mitocondrial Matriz mitocondrial Espacio intermembranoso Cresta mitocondrial ATP-sintetasa Membrana externa Membrana interna Mitorribosomas

90
Complejos ATP-sintetasas Las mitocondrias. Partículas elementales F
VOLVER Las mitocondrias. Partículas elementales F H+ ADP + Pi ATP Fo o pedúnculo (inmerso en la membrana, constituido de 3 /4 polipéptidos) Cabeza esférica o partícula F1 proteína globular
 Cabeza esférica o partícula F1 proteína globular.")
91
FUNCIONES Ciclo de Krebs (matriz) Cadena respiratoria (mb interna) Fosforilación oxidativa (ATP sintetasa) Beta oxidación de ácidos grasos (matriz) Concentración de sustancias Duplicación ADN
 Concentración de sustancias. Duplicación ADN.")
92
Las mitocondrias Bacterias aerobias Mitocondria Célula primitiva
Endosimbiosis Célula eucariota

94
Cloroplastos

95
Cloroplastos

96
Los cloroplastos ADN plastidial Membrana externa. Estructura como el resto de membranas. Más permeable Grana. Apilamientos con nº variable de sacos Membrana interna. Estructura como el resto de membranas. Casi impermeable, con proteínas transportadoras. Ribosomas (plastorribosomas) Rubisco. Enzima encargada de la fijación del CO2 Estroma Tilacoide del estroma
 Rubisco. Enzima encargada de la fijación del CO2. Estroma. Tilacoide del estroma.")
97
Los cloroplastos Bacterias aerobias Cianobacterias Célula primitiva
Mitocondria Cloroplasto

98
FUNCIONES Duplicación del ADN Reducción de nitratos a nitritos (los nitritos se reducen a amoníaco, que es la fuente de N para la síntesis de aminoácidos y nucleótidos) Fotosíntesis Fase dependiente de la luz (tilacoides) Fase independiente (estroma)
 Fotosíntesis. Fase dependiente de la luz (tilacoides) Fase independiente (estroma)")
99
Los cloroplastos

100
Aspecto al microscopio electrónico
Cloroplastos Aspecto al microscopio electrónico Las estructuras de los cloroplastos pueden observarse con facilidad al microscopio electrónico.

101
Cloroplastos Funaria hygrometrica Zygnema Spirogyra
Además a diferencia de otras estructuras de la célula, el tamaño y el color de los cloroplastos permite que se puedan observar con facilidad en el microscopio óptico. De hecho, no es necesario realizar fijaciones, ni tinciones, lo que permite su observación en una célula viva. El número de cloroplastos que aparece en los organismos fotosintéticos es muy variable una célula del tejido fotosintético de una hoja de angiosperma, pude albergar entre 40 o 50 cloroplastos, mientras que las antocerotas, un grupo de briofitos, cuenta con sólo cloroplasto por célula. Su forma también es bastante variable, si bien predomina la forma de disco. Estos cloroplastos no están fijos dentro de la célula, sino que gozan de movilidad, tomando diferentes patrones en función del origen o intensidad de la luz.

102
SEMEJANZAS DIFERENCIAS
Orgánulos energéticos de las células eucariotas. Poseen gran cantidad de mb interna. Son orgánulos semiautónomos (contienen los componentes necesarios para ello: ADNdc y ribosomas) Se reproducen por división binaria. Evolucionaron a partir de procariotas primitivas. Poseen ATPasas MITOCONDRIAS CLOROPLASTOS Tanto en vegetales como en animales. Menor tamaño. Posee dos membranas diferentes y 2 compartimentos. Función: Respiración celular. No poseen pigmentos. Proceden de primitivas bacterias aerobias Exclusivos de células vegetales. Tamaño mayor. Posee 3 membranas y 3 compartimentos internos separados. Función: Fotosíntesis. Poseen pigmentos. Proceden de primitivas cianobacterias Las mitocondrias tienen tamaños más reducidos que los plastos, presentándose en un número mucho más elevado. Son más numerosas en aquellas células con metabolismo activo, pudiendo llegando a haber hasta decenas de millares por célula. Al igual que los cloroplastos, las mitocondrias se desplazan por la célula, acercándose a aquellas partes que estén consumiendo cantidades elevadas. Así, se colocan en las base de los flagelos cuando estos están activos.
 Se reproducen por división binaria. Evolucionaron a partir de procariotas primitivas. Poseen ATPasas. MITOCONDRIAS. CLOROPLASTOS. Tanto en vegetales como en animales. Menor tamaño. Posee dos membranas diferentes y 2 compartimentos. Función: Respiración celular. No poseen pigmentos. Proceden de primitivas bacterias aerobias. Exclusivos de células vegetales. Tamaño mayor. Posee 3 membranas y 3 compartimentos internos separados. Función: Fotosíntesis. Poseen pigmentos. Proceden de primitivas cianobacterias. Las mitocondrias tienen tamaños más reducidos que los plastos, presentándose en un número mucho más elevado. Son más numerosas en aquellas células con metabolismo activo, pudiendo llegando a haber hasta decenas de millares por célula. Al igual que los cloroplastos, las mitocondrias se desplazan por la célula, acercándose a aquellas partes que estén consumiendo cantidades elevadas. Así, se colocan en las base de los flagelos cuando estos están activos.")
Presentaciones similares




















.>")





.>")
