Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Unidad 8: Morfología celular II

2
Índice Retículo endoplásmico Aparato de Golgi Lisosomas Peroxisomas
Mitocondrias Cloroplastos Pared celular Vacuolas

3
Retículo endoplásmico

4
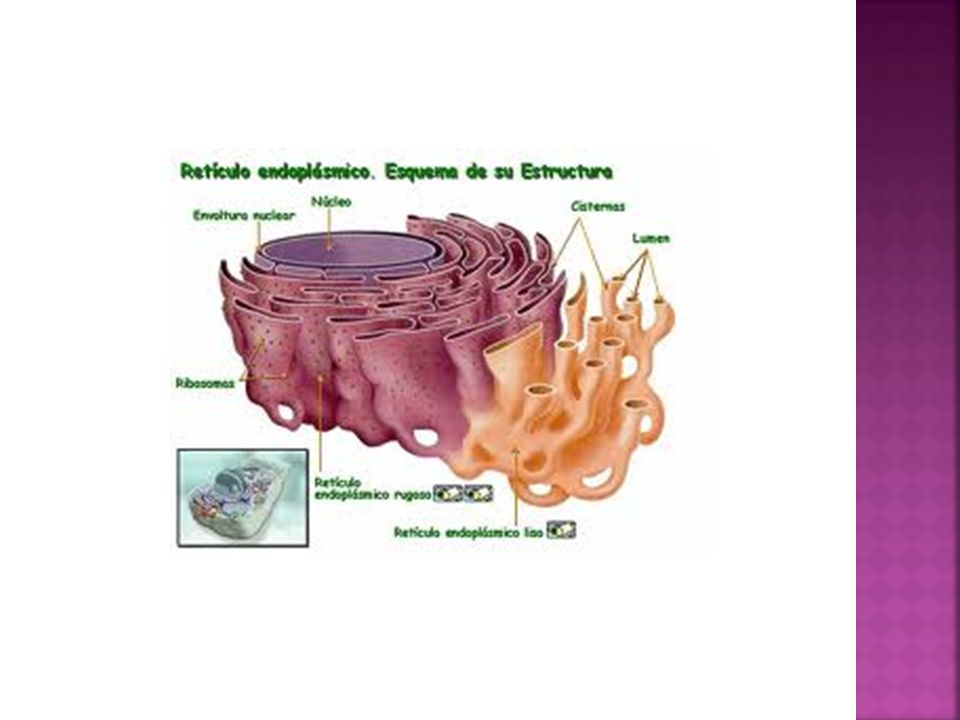
Retículo endoplasmático
Es una extensa red de sacos aplanados y túbulos intercomunicados que fabrican y transportan materiales dentro de las células eucariotas. La cantidad de retículo endoplasmático (RE) no es fija, sino que aumenta o disminuye de acuerdo a la actividad celular. Se halla en comunicación con la membrana nuclear externa. Este sistema constituye un único compartimiento con un espacio interno que recibe el nombre de lumen. Es el orgánulo más grande de la mayor parte de las células eucariotas. Se distinguen dos clases de retículo endoplasmático: Retículo endoplasmático rugoso (RER) con ribosomas en su cara externa (citosolica) Retículo endoplasmático liso (REL), que carece de ribosomas.
 no es fija, sino que aumenta o disminuye de acuerdo a la actividad celular. Se halla en comunicación con la membrana nuclear externa. Este sistema constituye un único compartimiento con un espacio interno que recibe el nombre de lumen. Es el orgánulo más grande de la mayor parte de las células eucariotas. Se distinguen dos clases de retículo endoplasmático: Retículo endoplasmático rugoso (RER) con ribosomas en su cara externa (citosolica) Retículo endoplasmático liso (REL), que carece de ribosomas.")
6
PROFESOR JANO - Estrategias de Trabajo y aprendizaje
23/03/2017 TIPOS RUGOSO LISO - Recursos culturales

7
PROFESOR JANO - Estrategias de Trabajo y aprendizaje
23/03/2017 Retículo endoplasmático rugoso Presenta ribosomas en la cara externa, la llamada cara citoplasmática. Está formado por sáculos aplastados comunicados entre sí. Se encuentra comunicado con el REL y con la membrana externa de la envoltura nuclear. El tamaño depende de la actividad celular (mayor en células muy activas) - Recursos culturales
 - Recursos culturales.")
8
PROFESOR JANO - Estrategias de Trabajo y aprendizaje
23/03/2017 Funciones RER Síntesis y almacenamiento de proteínas. En el RER tiene lugar la síntesis de proteínas que pueden tener dos destinos: Ser secretadas por las células Formar parte de los orgánulos (RE, Aparato de Golgi, lisosomas, membrana...) Glucosilación de proteínas. Adicción de azúcares a las proteínas. Proceso que tienen lugar en el lumen de forma previa al transporte a otro destino. 3. Plegamiento de proteínas. Proceso que tiene lugar en el lumen - Recursos culturales
 Glucosilación de proteínas. Adicción de azúcares a las proteínas. Proceso que tienen lugar en el lumen de forma previa al transporte a otro destino. 3. Plegamiento de proteínas. Proceso que tiene lugar en el lumen. - Recursos culturales.")
9
Síntesis de proteínas en la membrana del RER
Se inicia en el citosol. Se une el ARNm a la subunidad pequeña del ribosoma. Se unen ambas subunidades del ribosoma. Comienza la formación de la proteína que presenta en su extremo un péptido de señalización. Este péptido es reconocido por la membrana del RER que permite al ribosoma unirse a receptores de la membrana. La proteína es introducida a través de proteínas transmembranosas en el lumen, donde pierde el péptido de señalización. Al tiempo que la proteína se sintetiza en la luz del retículo, se le unen los oligosacáridos (glucosilación). Se comienza a plegar la proteína y cuando finaliza su síntesis, el ribosoma se suelta y vuelve al citosol y los canales de membrana se cierran.
. Se comienza a plegar la proteína y cuando finaliza su síntesis, el ribosoma se suelta y vuelve al citosol y los canales de membrana se cierran.")
11
Las proteínas no salen del RER si no están perfectamente plegadas y ensambladas.
Las proteínas que no están en condiciones son degradadas en el propio RE. Las proteínas propias del RER llevan una corta señal que las identifica; si son erróneamente empaquetadas en una vesícula y dirigidas al Golgi, la señal es reconocida y son enviadas de retorno desde el aparato de Golgi al RE, donde son destruidas.

12
Retículo endoplasmático liso (REL)
Es un tipo de retículo endoplasmático que carece de ribosomas. Está constituido por una red de túbulos unidos al retículo endoplasmático rugoso y que se expande por todo el citoplasma. La membrana del retículo endoplasmático liso posee gran cantidad de enzimas cuya principal actividad es la síntesis de lípidos. Es muy abundante en células hepáticas, musculares, ováricas, de los testículos, y en la corteza suprarrenal.

13
Funciones del retículo endoplasmático liso
Síntesis, almacenamiento y transporte de lípidos y derivados lipídicos. Se sintetizan casi todos los lípidos constituyentes de las membranas: colesterol, fosfolípidos. En ciertos tipos celulares se sintetizan los derivados lipídicos de las hormonas esteroideas (células de Sertoli, células de Leydig) o los ácidos biliares (hepatocitos) Detoxificación de sustancias tóxicas liposolubles. Contiene enzimas desintoxicantes que degradan sustancias químicas como carcinógenos y los conviertan en moléculas solubles fácilmente excretables por el organismo. Muy importante en el hígado, también en riñones, piel, pulmones. Regulación de los niveles de calcio intracelular. La membrana del REL posee bombas de Calcio, dependientes de ATP, que permiten captar calcio desde el citosol. También posee canales de calcio, que permiten liberar calcio por parte del REL hacia el citosol . (contracción muscular) Liberación de glucosa a partir del glucógeno (en hepatocitos). Elimina el grupo fosfato de la G-6-P y la convierte en glucosa lista para ser exportada al torrente sanguíneo.
 o los ácidos biliares (hepatocitos) Detoxificación de sustancias tóxicas liposolubles. Contiene enzimas desintoxicantes que degradan sustancias químicas como carcinógenos y los conviertan en moléculas solubles fácilmente excretables por el organismo. Muy importante en el hígado, también en riñones, piel, pulmones. Regulación de los niveles de calcio intracelular. La membrana del REL posee bombas de Calcio, dependientes de ATP, que permiten captar calcio desde el citosol. También posee canales de calcio, que permiten liberar calcio por parte del REL hacia el citosol . (contracción muscular) Liberación de glucosa a partir del glucógeno (en hepatocitos). Elimina el grupo fosfato de la G-6-P y la convierte en glucosa lista para ser exportada al torrente sanguíneo.")
14
Aparato de Golgi

15
Está localizado cerca del núcleo celular
Aparato de Golgi Fue descubierto por Camilo Golgi en 1898 gracias a una nueva técnica de tinción con sales de plata. Está localizado cerca del núcleo celular El aparato de Golgi está formado por uno o varios dictiosomas (agrupación en paralelo de cinco a ocho sáculos aplanados o cisternas), acompañados de vesículas de secreción. Los sáculos o cisternas están dilatados en los extremos y asociados se localizan las vesículas. Camilo Golgi
, acompañados de vesículas de secreción. Los sáculos o cisternas están dilatados en los extremos y asociados se localizan las vesículas. Camilo Golgi.")
16
La agrupación de sáculos se denomina dictiosoma
En este esquema se aprecian los sáculos aplanados y con los extremos dilatados. La agrupación de sáculos se denomina dictiosoma

17
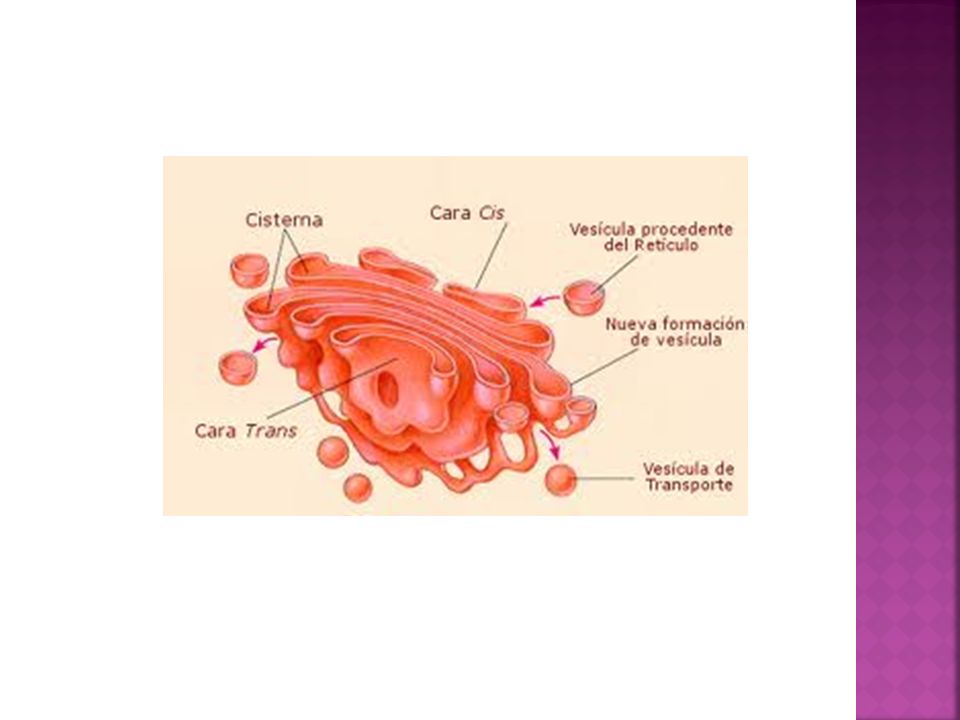
Estructura del aparato de Golgi
El aparato de Golgi está estructural y fisiológicamente polarizado. Cada uno de los dictiosmas presenta tres regiones diferenciadas: Cara cis o cara de formación: se encuentra más cercana al núcleo. Convexa. La cara cis recibe vesículas (vesículas de transición) procedentes del Retículo endoplásmico (lípidos y proteínas), que se fusionan con las membranas de dicha cara. Cara media: sáculos de la parte central del dictiosoma. Tras pasar por la cara cis, pasan a la cara media, donde tienen lugar la mayor parte de las actividades metabólicas del aparato de Golgi. Cara trans o cara de maduración: sáculos más alejados del núcleo y por tanto más cercanos a la membrana. Cóncava. Una vez que han sido modificadas pasan a la cara trans donde se empaquetan en vesículas de transporte (vesículas de secreción) hacia su destino final (lisosomas, membrana o exterior celular).
 procedentes del Retículo endoplásmico (lípidos y proteínas), que se fusionan con las membranas de dicha cara. Cara media: sáculos de la parte central del dictiosoma. Tras pasar por la cara cis, pasan a la cara media, donde tienen lugar la mayor parte de las actividades metabólicas del aparato de Golgi. Cara trans o cara de maduración: sáculos más alejados del núcleo y por tanto más cercanos a la membrana. Cóncava. Una vez que han sido modificadas pasan a la cara trans donde se empaquetan en vesículas de transporte (vesículas de secreción) hacia su destino final (lisosomas, membrana o exterior celular).")
20
Funciones del aparato de Golgi
Finaliza la glucosilación de las proteínas: que había sido iniciada en el RER. Síntetiza glucolípidos y esfingomielina: lípidos característicos de la membrana plasmática de las células. Sintetiza los polisacáridos (hemicelulosa y pectina) que se situarán en la pared celular y que son transportados hasta ella a través de vesículas. Dirigir la distribución y exportación de proteínas, excepto aquellas que se quedan en el propio orgánulo.
 que se situarán en la pared celular y que son transportados hasta ella a través de vesículas. Dirigir la distribución y exportación de proteínas, excepto aquellas que se quedan en el propio orgánulo.")
21
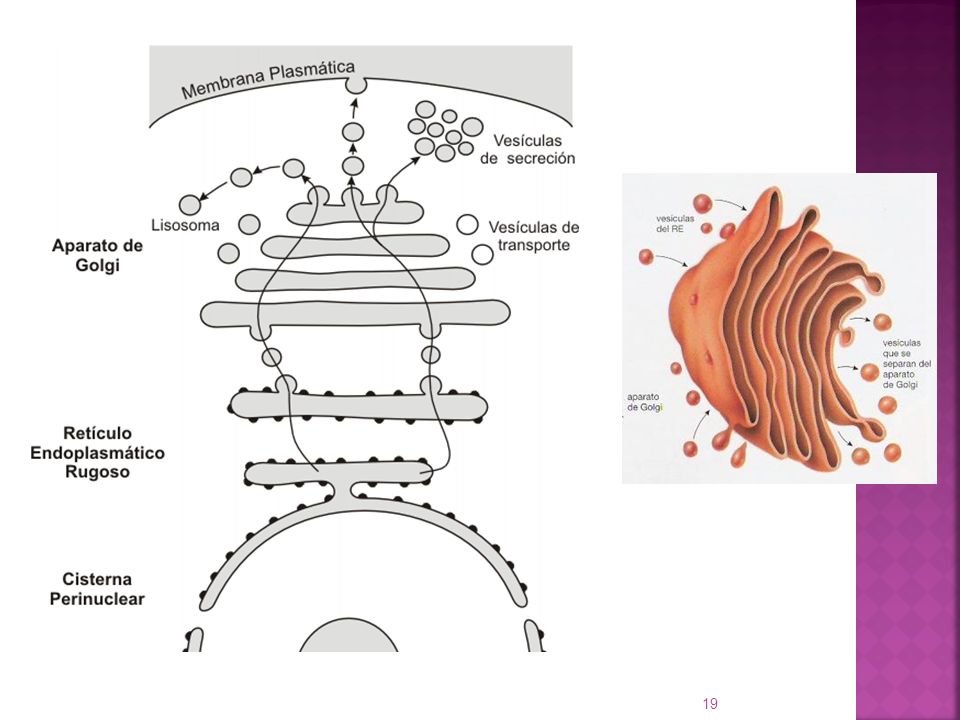
Transporte de proteínas de secreción desde la cara trans
del Aparato de Golgi Todas las proteínas que pasan por el Aparato de Golgi llegan a la cara trans. Una vez que llegan a esta cara son clasificadas y agrupadas según su destino final. Las proteínas pueden ser secretadas por dos vías: Vía de secreción constitutiva: Presente en todas las células Se utiliza para renovar los constituyentes de la membrana plasmática. Salen vesículas de transporte o de secreción que de forma continua se fusionan con la membrana Vía de secreción regulada: Está presente en determinadas células secretoras especializadas. Los materiales que siguen esta vía (hormonas, neurotransmisores, enzimas digestivas) salen de la cara trans en vesículas, pero estas quedan almacenadas en el citoplasma hasta que se produce el estímulo adecuado.
 salen de la cara trans en vesículas, pero estas quedan almacenadas en el citoplasma hasta que se produce el estímulo adecuado.")
22
Formación del acrosoma de los espermatozoides
Otras funciones del aparato de Golgi Síntesis de proteoglucanos, que son parte esencial de la matriz extracelular. Formación del acrosoma de los espermatozoides Formación del fragmoplasto en las células vegetales

23
Lisosomas

24
Lisosomas Los lisosomas son vesículas procedentes del aparato de Golgi que contienen enzimas digestivas. Son los encargados de la digestión intracelular. Las enzimas son hidrolasas ácidas (actúan a pH óptimo de 5) Para mantener el pH próximo a 5, en la membrana de los lisosomas hay bombas de protones dependientes de ATP que bombean protones al interior. También poseen en la membrana proteínas de transporte para pasar los productos resultantes de la digestión intracelular al citosol. Los lisosomas abundan en las células encargadas de combatir las enfermedades, como los leucocitos.
 Para mantener el pH próximo a 5, en la membrana de los lisosomas hay bombas de protones dependientes de ATP que bombean protones al interior. También poseen en la membrana proteínas de transporte para pasar los productos resultantes de la digestión intracelular al citosol. Los lisosomas abundan en las células encargadas de combatir las enfermedades, como los leucocitos.")
26
Estructura de los lisosomas
Su tamaño es muy variable. Los lisosomas poseen una membrana plasmática con las proteínas de su cara interna muy glucosiladas. Estas glucoproteínas impiden que las enzimas hidrolasas ataquen a la propia membrana del lisosoma.

27
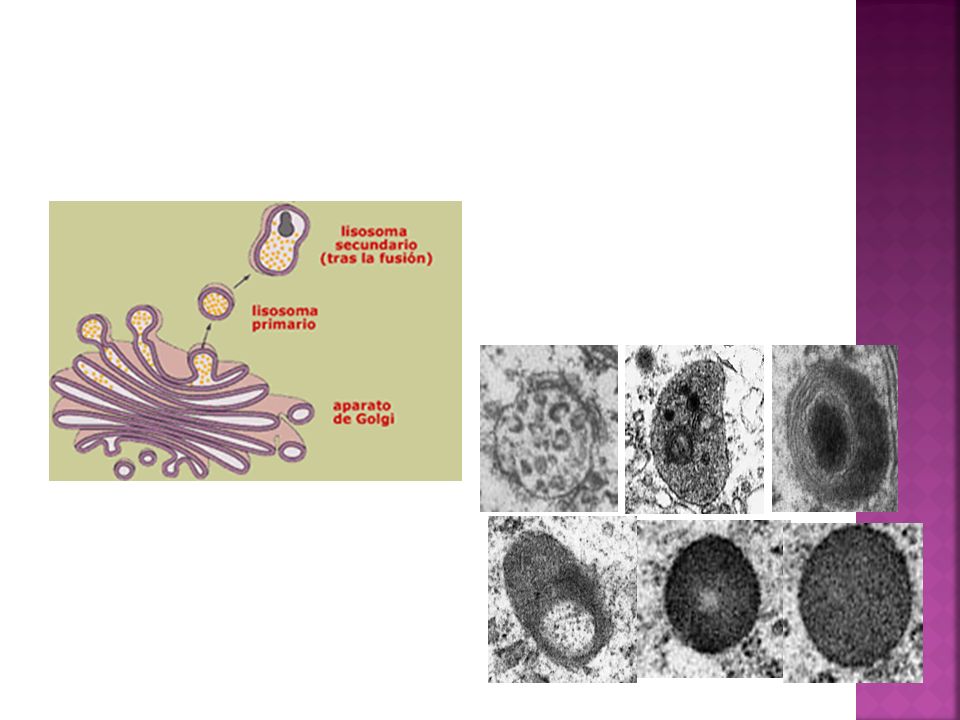
Clasificación de los lisosomas
Lisosomas primarios: lisosomas que acaban de salir de la cara trans del Aparato de Golgi. Contienen solamente enzimas hidrolíticas, pues todavía no han participado de ningún proceso digestivo. Se observan como vesículas de pequeño tamaño con el contenido homogéneo. Lisosomas secundarios: se forman cuando los lisosomas primarios se fusionan con vesículas que contienen los materiales a digerir (que proceden del interior o exterior celular). Son de mayor tamaño y de contenido heterogéneo. Cuerpos residuales: lisosomas secundarios que han finalizado el proceso de digestión intracelular y que contienen material no digerible. Son de tamaño variable y contenido heterogéneo
. Son de mayor tamaño y de contenido heterogéneo. Cuerpos residuales: lisosomas secundarios que han finalizado el proceso de digestión intracelular y que contienen material no digerible. Son de tamaño variable y contenido heterogéneo.")
28
Autofagia y heterofagia
Los materiales que puede digerir el lisosoma pueden ser: Materiales u orgánulos obsoletos de la propia célula (autofagia), que son rodeados por una membrana procedente del retículo endoplásmico, formándose un autofagosoma, que se fusionará con un lisosoma, formando un lisosoma secundario. Materiales que llegan del exterior (Heterofagia). Pueden llegar por dos vías diferentes: La endocitosis dependiente de clatrina que llevan los materiales endocitados hacia los endosomas, que se fusionarán con los lisosomas para formar un lisosoma secundario. La fagocitosis que engloba partículas de gran tamaño y dichas partículas forman un fagosoma, que se fusionaran con los lisosomas primarios para formar lisosomas secundarios.
, que son rodeados por una membrana procedente del retículo endoplásmico, formándose un autofagosoma, que se fusionará con un lisosoma, formando un lisosoma secundario. Materiales que llegan del exterior (Heterofagia). Pueden llegar por dos vías diferentes: La endocitosis dependiente de clatrina que llevan los materiales endocitados hacia los endosomas, que se fusionarán con los lisosomas para formar un lisosoma secundario. La fagocitosis que engloba partículas de gran tamaño y dichas partículas forman un fagosoma, que se fusionaran con los lisosomas primarios para formar lisosomas secundarios.")
29
(materiales exterior)
AUTOFAGOSOMA AUTOFAGIA (materiales propios) ENDOCITOSIS ENDOSOMA HETEROFAGIA (materiales exterior) FAGOCITOSIS FAGOSOMA
 ENDOCITOSIS. ENDOSOMA. HETEROFAGIA. (materiales exterior) FAGOCITOSIS. FAGOSOMA.")
30
Endocitosis

31
Función de los lisosomas
Los lisosomas realizan la digestión intracelular de materia orgánica gracias a las hidrolasas ácidas que contienen.

32
Lisosomas especiales (culturilla)
El acrosoma es un lisosoma primario en el que se almacenan enzimas capaces de digerir las membranas foliculares del óvulo, para permitir el paso del espermatozoide y la fecundación. Los granos de aleurona son lisosomas secundarios en donde se almacenan proteínas que, debido a la pérdida de agua, se encuentran en estado cristalino, hasta que al plantarse y absorberse agua se activan las enzimas y "se inicia la digestión de las mismas, con lo que empieza la germinación de la semilla. Cuando una célula muere, la membrana lisosómica se rompe y libera hacia el citoplasma enzimas digestivas, que degradan a la célula en sí. Algunas formas de daño tisular, se relacionan con la existencia de lisosomas "con fugas". Se cree que la artritis reumatoide se debe, en parte, a la lesión de las células del cartílago provocada por enzimas liberadas de los lisosomas.

33
Los peroxisomas Los peroxisomas son vesículas membranosas que contienen enzimas oxidasas. Utilizan el oxígeno molecular para oxidar distintos sustratos orgánicos, produciendo peróxido de hidrógeno (agua oxigenada). El peróxido de hidrógeno es tóxico para la célula por lo que se degrada en el peroxisoma por intervención de una enzima denominada catalasa. Los peroxisomas intervienen en procesos de destoxificación
. El peróxido de hidrógeno es tóxico para la célula por lo que se degrada en el peroxisoma por intervención de una enzima denominada catalasa. Los peroxisomas intervienen en procesos de destoxificación.")
34
Función de los peroxisomas
Destoxificación de ciertas sustancias. Sirven para eliminar el exceso de ácidos grasos, aminoácidos o NADPH. En las células vegetales reciben el nombre de glioxisomas Origen de los peroxisomas Culturilla Se considera que los peroxisomas aparecieron antes que las mitocondrias y que su función era permitir la vida en una atmósfera cada vez más rica en oxígeno, elemento tóxico para los organismos anaerobios. Proceden de la simbiosis con otras células, y su genoma quedó incorporado al genoma celular.

35
MITOCONDRIAS

36
Mitocondrias Fueron descubiertas por Altman en 1886, que los denominó bioblastos. Las mitocondrias son orgánulos presentes en todas las células eucariotas aerobias (animales y vegetales) que se encargan de la obtención de energía en forma de ATP mediante la respiración celular. El conjunto de mitocondrias de una célula se denomina condrioma. Se observan mal “in vivo” debido a su pequeño tamaño
 que se encargan de la obtención de energía en forma de ATP mediante la respiración celular. El conjunto de mitocondrias de una célula se denomina condrioma. Se observan mal in vivo debido a su pequeño tamaño.")
37
Estructura de las mitocondrias
Las mitocondrias son orgánulos polimorfos, pudiendo variar desde formas esféricas hasta alargadas a modo de bastoncillo. Presentan una doble membrana: una membrana externa lisa una membrana interna con numerosos repliegues internos, denominados crestas mitocondriales. Estas membranas originan dos compartimentos: el espacio intermembraso la matriz mitocondrial

38
Membranas mitocondriales
Membrana mitocondrial externa: Proteínas transmembranosas (porinas) que son proteínas que forman canales por los que pueden pasar moléculas de bajo peso molecular, dándole una gran permeabilidad frente a electrolitos, agua Una gran cantidad de enzimas que intervienen en numerosos procesos metabólicos (síntesis de algunos lípidos o la unión de los ácidos grasos al coenzima A para su ingreso en la mitocondria antes de su degradación)
 que son proteínas que forman canales por los que pueden pasar moléculas de bajo peso molecular, dándole una gran permeabilidad frente a electrolitos, agua. Una gran cantidad de enzimas que intervienen en numerosos procesos metabólicos (síntesis de algunos lípidos o la unión de los ácidos grasos al coenzima A para su ingreso en la mitocondria antes de su degradación)")
39
Membranas mitocondriales
Membrana mitocondrial interna: La membrana interna presenta repliegues denominados crestas mitocondriales que incrementan su superficie y, por tanto, su capacidad metabolizadora. Tiene un contenido en proteínas mayor que cualquier biomembrana de la célula. Su biomembrana posee un alto contenido en cardiolipina (folsfolípido doble (difosfatidilglicerol) que la hace impermeable al paso de partículas con carga). En ella se encuentran las cadenas de transporte electrónico En su cara interna presenta unas partículas que se proyectan hacia la matriz y corresponden a las partículas F1 de los complejos ATP sintasa, cuya fracción F0 está inmersa en la membrana mitocondrial interna. Entre sus lípidos de membrana no aparece el colesterol, por lo que tiene gran fluidez.
 que la hace impermeable al paso de partículas con carga). En ella se encuentran las cadenas de transporte electrónico. En su cara interna presenta unas partículas que se proyectan hacia la matriz y corresponden a las partículas F1 de los complejos ATP sintasa, cuya fracción F0 está inmersa en la membrana mitocondrial interna. Entre sus lípidos de membrana no aparece el colesterol, por lo que tiene gran fluidez.")
40
Las ATP-sintasas o partículas elementales
Están en las crestas mitocondriales, orientadas hacia la matriz

41
CARIDOLIPINA

42
Función de la membrana mitocondrial interna
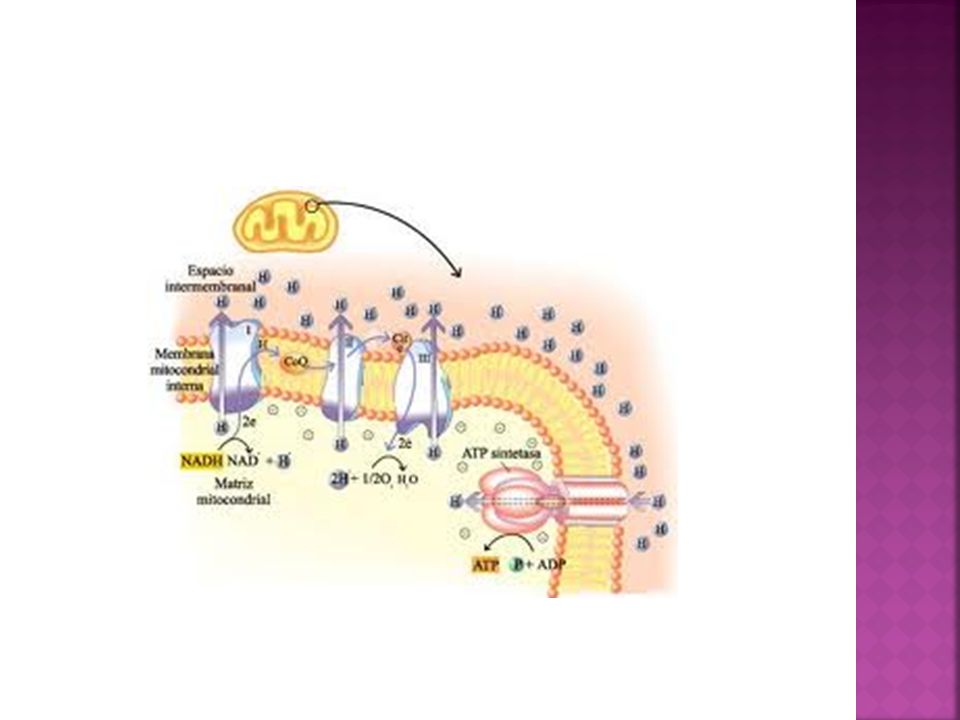
Se llevan a cabo los procesos que utilizan energía liberada de las reacciones de oxidación para sintetizar ATP. Cadena respiratoria: formada por varios complejos enzimáticos, en los que los coenzimas recogen los electrones procedentes de la oxidación hasta el oxígeno que es el aceptor final. Este transporte libera energía que es usada para el bombeo de protones hacia el espacio intermembrana. Fosforilación oxidativa: proceso en el que se sintetiza ATP a partir de ADP+ P, que se produce a nivel de las partículas F0F1

44
Espacio intermembrana o intermebranoso
Se encuentra entre ambas membranas (externa e interna) De contenido similar al del citosol. Tiene enzimas que catalizan la fosforilación oxidativa de otros nucleótidos a partir del ATP (procedente del interior de la mitocondria)
 De contenido similar al del citosol. Tiene enzimas que catalizan la fosforilación oxidativa de otros nucleótidos a partir del ATP (procedente del interior de la mitocondria)")
45
Matriz mitocondrial La matriz mitocondrial es un medio interno, con consistencia de gel, rico en enzimas y en el que se llevan a cabo un gran número de reacciones bioquímicas. Genoma mitocondrial, formado por múltiples copias de ADN en doble hélice cerrada. Contiene información necesaria para la síntesis de algunas proteínas mitocondriales. Ribosomas similares a los de bacterias, denominados mitorribosomas, que son 70S Distintos tipos de ARN Alta concentración de proteínas, entre las cuales se pueden destacar las enzimas responsables de las reacciones del metabolismo oxidativo. Enzimas de la replicación, transcripción y traducción del ADN mitocondrial

46
En la matriz tienen lugar:
Reacciones de oxidación de distintas moléculas para formar ácido acético. Ciclo de krebs (conjunto de reacciones de oxidación del ácido acético (en forma de acetil CoA) procedente de todas las oxidaciones anteriores para formar CO2
 procedente de todas las oxidaciones anteriores para formar CO2.")
48
cloroplastos

49
PLASTOS Con este nombre se denomina genéricamente a un grupo de orgánulos que producen y almacenan productos nutritivos en algas y plantas. Leucoplastos. Acumulan sustancias: Los amiloplastos son plástidos especiales que reservan almidón en los tejidos no fotosintéticos. Cromoplastos son los que poseen pigmentos que les dan color cloroplastos (clorofila de color verde) rodoplastos (ficoeritrina de color rojo)
 rodoplastos (ficoeritrina de color rojo)")
50
Cloroplastos Los cloroplastos son unos orgánulos típicos de las células vegetales que poseen clorofila, por lo que pueden realizar la fotosíntesis, proceso en el que se transforma la energía luminosa en energía química. Descubiertos en 1881 Morfología variada, depende del organismo, pero generalmente ovoides.

51
Estructura de los cloroplastos
Los cloroplastos son el tipo más común de plastos. Contienen clorofila, un pigmento de color verde del cual hay varios tipos (en las plantas terrestres las clorofilas más comunes son las clorofilas a y b, pero en las algas hay otros tipos). Los cloroplastos también contienen una variedad de pigmentos carotenoides y xantofilas que absorben radiaciones luminosas en zonas del espectro visible donde no absorben las clorofilas (pigmentos fotosintéticos accesorios)
. Los cloroplastos también contienen una variedad de pigmentos carotenoides y xantofilas que absorben radiaciones luminosas en zonas del espectro visible donde no absorben las clorofilas (pigmentos fotosintéticos accesorios)")
52
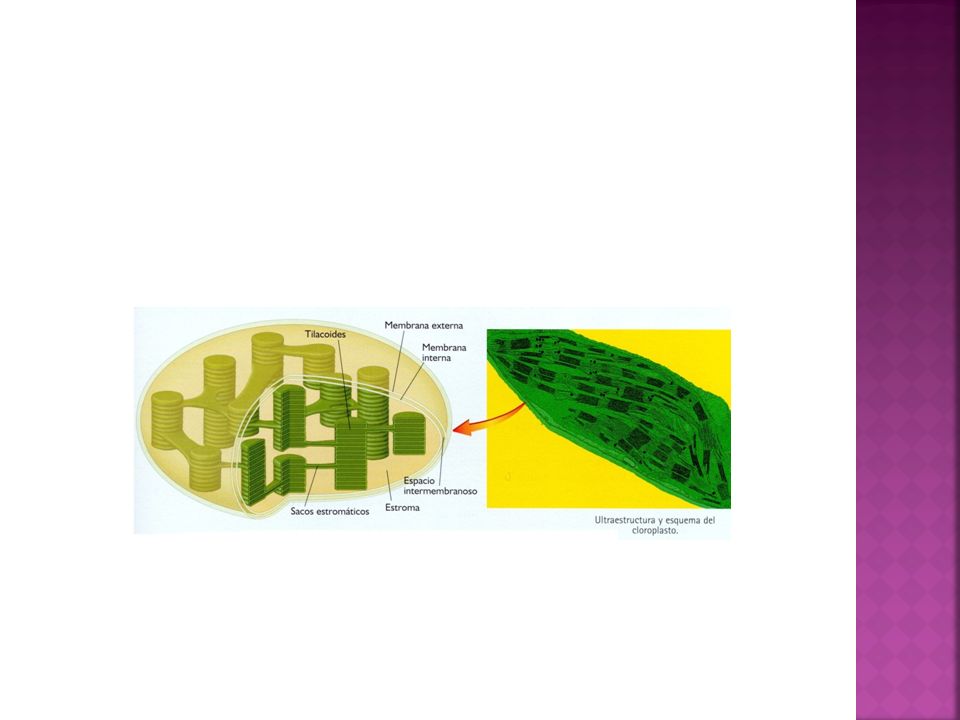
Doble membrana (interna y externa con un espacio intermembranoso).
Estroma ADN circular de doble cadena. Plastorribosomas. Complejos F1 Enzimas (Rubisco) Tilacoides Grana. Sacos apilados Sacos estromáticos Procesos genéticos del cloroplasto. Replicación Transcripción Traducción
 Tilacoides. Grana. Sacos apilados. Sacos estromáticos. Procesos genéticos del cloroplasto. Replicación. Transcripción. Traducción.")
54
Membranas de los cloroplastos
Los cloroplastos están delimitados por dos membranas Membrana plastidial externa: muy permeable. Contiene porinas que permiten el paso de moléculas de pequeño tamaño. Membrana plastidial interna: menos permeable que la externa. Ambas membranas carecen de clorofila y entre sus lípidos, al igual que en las mitocondrias, no está el colesterol. Ambas membranas son continuas y delimitan el cloroplasto. No poseen ni repliegues ni crestas.

55
Espacio intermembranoso
Se localiza entre las dos membranas plastidiales (externa e interna). Composición semejante al citosol.
. Composición semejante al citosol.")
56
Estroma Es análogo a la matriz mitocondrial, contiene: Enzimas encargadas de producir glucosa a partir de dióxido de carbono y agua (la más importante es la Rubisco) Ribosomas semejantes a los de bacterias ARN ADN circular y bicatenario Los enzimas para la transcripción, traducción y replicación del ADN Inclusiones de granos de almidón y lípidos
 Ribosomas semejantes a los de bacterias. ARN. ADN circular y bicatenario. Los enzimas para la transcripción, traducción y replicación del ADN. Inclusiones de granos de almidón y lípidos.")
57
Membrana tilacoidal. Tilacoides Existe un tercer sistema de membranas, formado por sacos planos, de forma discoidal, interconectados unos con otros, llamados tilacoides. Estas membranas forman un tercer compartimiento, cuyo interior se denomina espacio tilacoidal o espacio intratilacoidal. Los sáculos se agrupan formando pilas (grana). Los grana se comunican por los tilacoides intergrana La membrana tilacoidal es rica en clorofila e interviene en la formación de ATP (ATP sintasas-).
. Los grana se comunican por los tilacoides intergrana. La membrana tilacoidal es rica en clorofila e interviene en la formación de ATP (ATP sintasas-).")
59
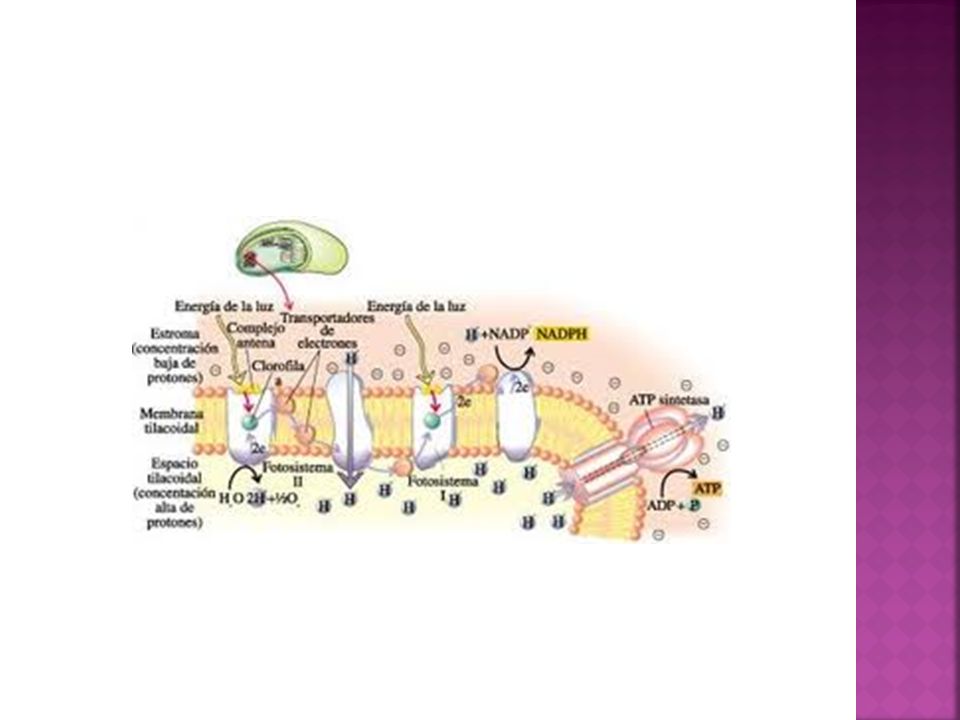
Fisiología del cloroplasto
En la membrana tilacoidal: Se llevan a cabo todas las reacciones de la fotosíntesis que son dependientes de la luz. Se comienza con la captación de la energía solar por los pigmentos fotosintéticos y la fotolísis del agua (ruptura de la molécula de agua por acción de la luz). De la ruptura del agua se generan electrones que pasaran a un transporte electrónico (que servirá para reducir el coenzima NADP+ a NADPH) y se desprenderá oxígeno molecular. El transporte de los electrones a través de la cadena de transporte genera un gradiente de protones (desde el estroma hacia el espacio tilacoidal) que se utiliza para la síntesis de ATP en los complejos ATPsintasa de la membrana tilacoidal. Este proceso se denomina fotofosforilación o fosforilación fotosintética.
. De la ruptura del agua se generan electrones que pasaran a un transporte electrónico (que servirá para reducir el coenzima NADP+ a NADPH) y se desprenderá oxígeno molecular. El transporte de los electrones a través de la cadena de transporte genera un gradiente de protones (desde el estroma hacia el espacio tilacoidal) que se utiliza para la síntesis de ATP en los complejos ATPsintasa de la membrana tilacoidal. Este proceso se denomina fotofosforilación o fosforilación fotosintética.")
61
Fisiología del cloroplasto
En el estroma: En el estroma se localiza la ribulosa-1,5 difosfato carboxilasa y otras enzimas implicadas en la fijación del CO2 en el ciclo de Calvin. Tienen lugar aquellas reacciones no dependientes de la luz. En ellas el ATP y el NADPH se utilizan para reducir el dióxido de carbono, iones nitrato o sulfato y elaborar moléculas orgánicas.

62
Funciones de los cloroplastos
Realización de la fotosíntesis, en dos fases: Fase luminosa de la fotosíntesis Fase oscura de la fotosíntesis Síntesis de ATP mediante la quimioósmosis. Se origina un gradiente químico de H+ cuya energía es utilizada por las ATP-sintasas para la formación de ATP.

63
Pared celular

64
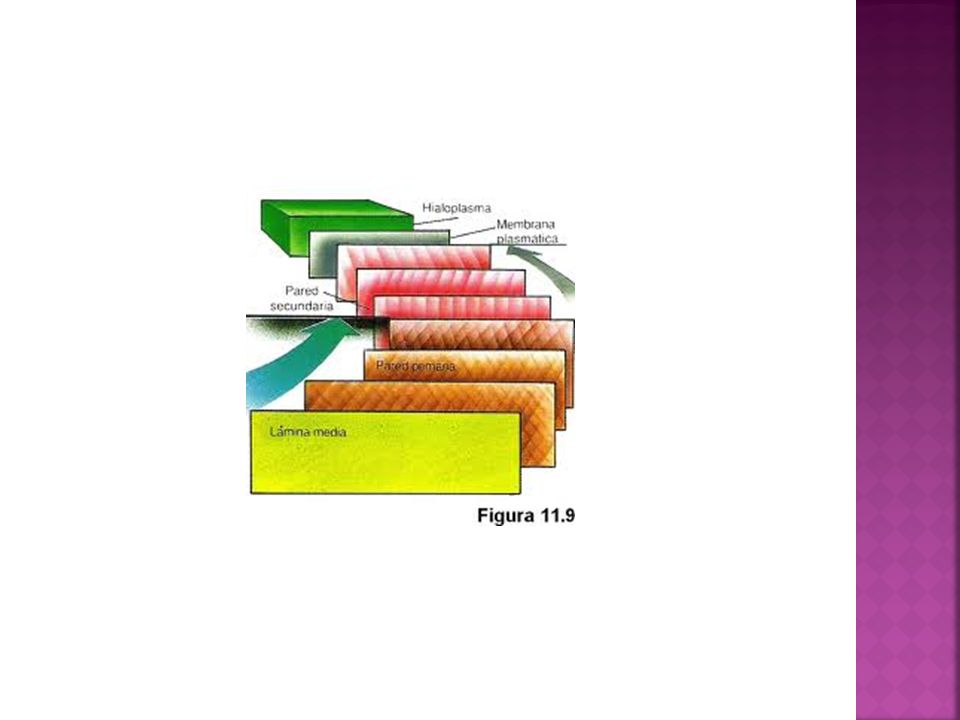
Tipo de matriz extracelular presente en células vegetales.
Pared celular Tipo de matriz extracelular presente en células vegetales. Compuesta fundamentalmente por celulosa Puede presentar tres capas, que se disponen externamente a la membrana plasmática lámina media (más externa) pared primaria pared secundaria (más interna)
 pared primaria. pared secundaria (más interna)")
65
Composición Células eucariotas Hongos Quitina Plantas Celulosa Hemicelulosa Pectina

66
Formada fundamentalmente por proteínas y pectinas.
Estructura Lámina media. Primera en formarse entre dos células que acaban de dividirse y permanecen unidas. En algunas zonas de comunicación entre células vecinas no aparece esta lámina (plasmodesmos) Formada fundamentalmente por proteínas y pectinas.
 Formada fundamentalmente por proteínas y pectinas.")
67
Pared primaria: Más gruesa que la lámina media
Formada por celulosa en una matriz de hemicelulosa y pectina. Se deposita durante el crecimiento entre la membrana plasmática y la lámina media.

68
Pared secundaria: Solo aparece en algunos tipos celulares (tejidos esqueléticos como el colénquima y esclerénquima y tejidos conductores como el xilema y floema) Más gruesa que la primaria Más interna en contacto con la membrana celular. Abundante celulosa y matriz con menos hemicelulosa. Puede impregnarse de diferentes sustancias como suberina, cutina, lignina y algunas sales minerales.
 Más gruesa que la primaria. Más interna en contacto con la membrana celular. Abundante celulosa y matriz con menos hemicelulosa. Puede impregnarse de diferentes sustancias como suberina, cutina, lignina y algunas sales minerales.")
69
Función de la pared celular
Da forma y rigidez a la célula e impide su ruptura. La célula vegetal contiene en su citoplasma una elevada concentración de moléculas que, debido a la presión osmótica, origina una corriente de agua hacia el interior celular que acabaría por hincharla y romperla si no fuera por la pared. Es responsable de que la planta se mantenga erguida. Protección frente abrasión mecánica, ataque de insectos, microorganismos patógenos. Participa en la comunicación entre las células, a través de plasmodesmos y punteaduras. Orienta el crecimiento de las células.

70
Formación de la pared celular
Comienza con la formación de una delgada lámina (media) entre dos células que acaban de dividirse. Las proteínas, hemicelulosa y pectina son segregadas por el Aparato de Golgi, así como suberina, lignina. La celulosa es sintetizada por la cara externa de la membrana plasmática, gracias a la participación de una enzima denominada celulosa sintasa. Esta enzima es una proteína integral que elabora la celulosa a partir de la glucosa y une las moléculas de celulosa para formar microfibrillas. La direccionalidad de las microfibrillas de celulosa depende de hacia donde se muevan la celulosa sintasa, que a su vez está dirigida por los microtúbulos situados en la cara citosólica de la membrana.
 entre dos células que acaban de dividirse. Las proteínas, hemicelulosa y pectina son segregadas por el Aparato de Golgi, así como suberina, lignina. La celulosa es sintetizada por la cara externa de la membrana plasmática, gracias a la participación de una enzima denominada celulosa sintasa. Esta enzima es una proteína integral que elabora la celulosa a partir de la glucosa y une las moléculas de celulosa para formar microfibrillas. La direccionalidad de las microfibrillas de celulosa depende de hacia donde se muevan la celulosa sintasa, que a su vez está dirigida por los microtúbulos situados en la cara citosólica de la membrana.")
72
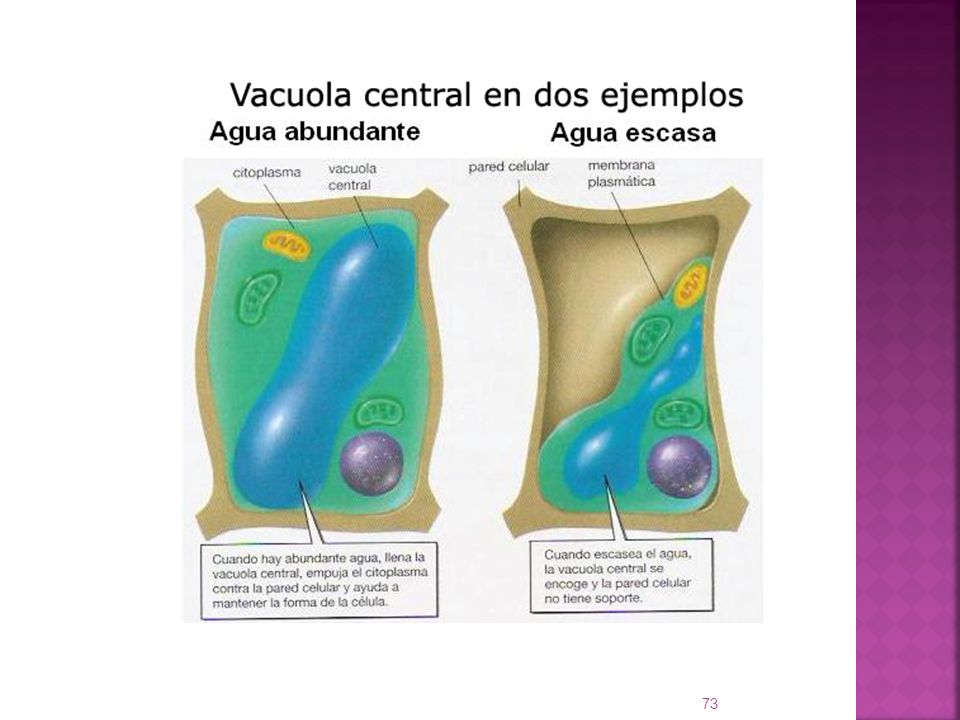
VACUOLAS Las vacuolas son vesículas constituidas por una membrana plasmática, y cuyo interior es predominantemente acuoso. Estructura de las vacuolas Las vacuolas de las células animales, suelen ser pequeñas, y se denominan vesículas. Las vacuolas de las células vegetales suelen ser muy grandes. Suele haber una o dos en cada célula. La membrana recibe el nombre de tonoplasto. A medida que la célula vegetal joven madura, las vacuolas crecen, llegando a ocupar en ocasiones hasta un 90%, de la célula vegetal madura. El conjunto de vacuolas de una célula vegetal recibe el nombre de vacuoma.

76
El tonoplasto presenta distintos sistemas de transporte activo.
Bomba de iones hacia la vacuola, favoreciendo la entrada de agua por ósmosis. El bombeo de protones hacia el interior de la vacuola, hace descender el pH (digestión intracelular)
")
77
Funciones de las vacuolas
Almacenamiento de nutrientes (azúcares, aminoácidos, proteínas, polisacáridos, iones, sustancias de desecho). Regulación homeostática, permite la adecuación a cambios ambientales. Digestión intracelular, que se produce en las vacuolas, por acumulo de enzimas hidrolíticas que proceden del aparato de Golgi. Acumulan sustancias tóxicas, que son liberadas cuando la célula es atacada. Acumulación de pigmentos que proporcionan color como atracción para la polinización entomófila.
. Regulación homeostática, permite la adecuación a cambios ambientales. Digestión intracelular, que se produce en las vacuolas, por acumulo de enzimas hidrolíticas que proceden del aparato de Golgi. Acumulan sustancias tóxicas, que son liberadas cuando la célula es atacada. Acumulación de pigmentos que proporcionan color como atracción para la polinización entomófila.")
78
Los glioxisomas Los glioxisomas son una clase de peroxisomas que sólo se encuentran en las células de los vegetales Su nombre deriva de que poseen las enzimas responsables del ciclo del ácido glioxílico, una variante del ciclo de Krebs, que permite sintetizar glúcidos a partir de lípidos. Esto resulta esencial para las semillas en germinación, ya que les permite, .a partir de sus reservas lipídicas, sintetizar glucosa, única molécula que admite el embrión, hasta que el nuevo vegetal pueda extender sus hojas y realizar la fotosíntesis. Peroxisomas de las hojas están relacionados con la fotorrespiración. Proceso por el cual la célula consume oxígeno y expulsa CO2. se piensa que evita la fotooxidación de la membrana tilacoidal.

Presentaciones similares