Descargar la presentación
La descarga está en progreso. Por favor, espere

1
METABOLISMO MICROBIANO
Es el estudio de las reacciones químicas que se llevan a cabo en las células METABOLISMO CELULAR La energía está involucrada en la mayor parte de las reacciones químicas de la célula microbiana. Cuando los átomos, iones y moléculas se combinan para formar sustancias químicas complejas, requieren un suministro de energía para su formación. Sin la síntesis de sustancias no habría formación de nuevo protoplasma y por ende no habría crecimiento y multiplicación celular. No puede haber vida sin una fuente de energía. María Cecilia Arango Jaramillo

2
Energía para el movimiento, transporte de nutrientes,etc
Catabolismo Productos de desecho Componentes celulares Nutrientes Anabolismo Energía para el desarrollo Fuente de energía

3
FASES DEL METABOLISMO:
ANABOLISMO : Formación o síntesis de compuestos químicos (Biosíntesis) CATABOLISMO : Degradación o descomposición de compuestos En las células microbianas vivas hay mucha actividad. Hay movimiento, transformación de energía, síntesis química de nuevos compuestos, descomposición química y transferencia de materiales. La suma total de estas reacciones en la célula microbiana se llama metabolismo. A las reacciones de síntesis de compuestos, formación de nuevo protoplasma, lo llamamos anabolismo, y a las reacciones de descomposición o degradación, las cuales involucran producción de energía lo llamamos catabolismo. METABOLISMO: ANABOLISMO : Formación o síntesis de compuestos químicos CATABOLISMO : Degradación o descomposición de compuestos
 CATABOLISMO : Degradación o descomposición de compuestos. En las células microbianas vivas hay mucha actividad. Hay movimiento, transformación de energía, síntesis química de nuevos compuestos, descomposición química y transferencia de materiales. La suma total de estas reacciones en la célula microbiana se llama metabolismo. A las reacciones de síntesis de compuestos, formación de nuevo protoplasma, lo llamamos anabolismo, y a las reacciones de descomposición o degradación, las cuales involucran producción de energía lo llamamos catabolismo. METABOLISMO: ANABOLISMO : Formación o síntesis de compuestos químicos. CATABOLISMO : Degradación o descomposición de compuestos.")
4
TRANSPORTADORES DE ENERGÍA
Las células microbianas, al igual que las máquinas, requieren una forma específica de energía. Una máquina de vapor requiere vapor y un motor eléctrico necesita electricidad para poder funcionar. No es sorprendente por lo tanto, que la célula microbiana requiera también una clase especial de energía. La energía que utiliza la célula microbiana es la energía almacenada en los enlaces químicos de los compuestos. A esta energía se le da el nombre de energía química. La energía química almacenada en los alimentos, se transfiere a una molécula transportadora que forma enlaces ricos en energía. Cuando estos enlaces se rompen, liberan la energía acumulada que la célula microbiana puede utilizar.

5
Compuestos ricos en energía : Adenosina trifosfato ( ATP )
Guanosina trifosfato ( GTP ) Acetil fosfato Ácido 1,3-difosfoglicérido Ácido fosfoenolpirúvico ( PEP ) Un compuesto rico en energía resulta análogo a una trampa de ratones. Cuando la trampa se dispone a cazar, tiene una gran energía acumulada en el resorte; cuando se dispara, la energía se libera. En la célula microbiana existen varios compuestos ricos en energía : Adenosina trifosfato ( ATP ) Guanosina trifosfato ( GTP ) Acetil fosfato Ácido 1,3-difosfoglicérido Ácido fosfoenolpirúvico ( PEP ) De los compuestos transportadores de energía, el más importante es el adenosina trifosfato (ATP). La célula microbiana cambia toda la energía química liberada de diferentes compuestos orgánicos a energía química transportada por las moléculas de ATP. Todas las “cuentas energéticas” dentro de la célula microbiana las “paga” el ATP (figura 38).
 Acetil fosfato. Ácido 1,3-difosfoglicérido. Ácido fosfoenolpirúvico ( PEP ) Un compuesto rico en energía resulta análogo a una trampa de ratones. Cuando la trampa se dispone a cazar, tiene una gran energía acumulada en el resorte; cuando se dispara, la energía se libera. En la célula microbiana existen varios compuestos ricos en energía : Adenosina trifosfato ( ATP ) Guanosina trifosfato ( GTP ) Acetil fosfato. Ácido 1,3-difosfoglicérido. Ácido fosfoenolpirúvico ( PEP ) De los compuestos transportadores de energía, el más importante es el adenosina trifosfato (ATP). La célula microbiana cambia toda la energía química liberada de diferentes compuestos orgánicos a energía química transportada por las moléculas de ATP. Todas las cuentas energéticas dentro de la célula microbiana las paga el ATP (figura 38).")
6
El ATP actúa como transportador de energía o como intermediario entre aquellas reacciones que proporcionan energía y las que la consumen. Dentro de las célula microbianas, la energía de los dos enlaces ricos en energía de la molécula de ATP no se pierde como calor, sino que se utiliza para hacer trabajo celular. Cualquier molécula que capte el grupo fosfato del ATP gana energía, se activa y entonces puede reaccionar con otras moléculas de la célula microbiana. El ATP es usado y resintetizado continuamente. Cuando una molécula de ATP libera un grupo fosfato rico en energía, se convierte en difosfato de adenosina (ADP). Cuando el ATP libera sus dos grupos fosfato ricos en energía, se convierte en monofosfato de adenosina (AMP). Para formar nuevamente una molécula de ATP, el ADP debe combinarse con un grupo fosfato y el AMP con dos grupos fosfato. La unión del grupo fosfato con ADP o AMP utiliza energía suministrada por el desdoblamiento de compuestos orgánicos dentro de la célula microbiana. Por ejemplo, la energía liberada por el desdoblamiento de una molécula de glucosa, se ahorra al usarla inmediatamente en la formación de enlaces fosfato ricos en energía entre el ADP y el fosfato. Por lo tanto, el ATP actúa como transportador de energía o como intermediario entre aquellas reacciones de los microorganismos que proporcionan energía y las que la consumen. Glucosa Energía ATP Energía Lípido CO2 + H2 O ADP + P ADP + P Ácido Graso + Glicerol
. Cuando el ATP libera sus dos grupos fosfato ricos en energía, se convierte en monofosfato de adenosina (AMP). Para formar nuevamente una molécula de ATP, el ADP debe combinarse con un grupo fosfato y el AMP con dos grupos fosfato. La unión del grupo fosfato con ADP o AMP utiliza energía suministrada por el desdoblamiento de compuestos orgánicos dentro de la célula microbiana. Por ejemplo, la energía liberada por el desdoblamiento de una molécula de glucosa, se ahorra al usarla inmediatamente en la formación de enlaces fosfato ricos en energía entre el ADP y el fosfato. Por lo tanto, el ATP actúa como transportador de energía o como intermediario entre aquellas reacciones de los microorganismos que proporcionan energía y las que la consumen. Glucosa Energía ATP Energía Lípido. 0 CO2 + H2 O ADP + P ADP + P Ácido Graso + Glicerol.")
7
UTILIZACIÓN DE LA ENERGÍA POR LOS MICROORGANISMOS

8
La célula microbiana utiliza la energía para: El movimiento.
La producción de calor. La electricidad. Biolumniscencia. El movimiento utiliza mucha energía. La producción de calor es otra forma en la cual se utiliza energía. Todas las célula microbianas liberan calor durante sus actividades metabólicas, en un día frío, mucha parte de la energía de los alimentos se usa para mantener esa temperatura. La electricidad es otra forma utilizada por las célula microbianas vivas. La transmisión de los impulsos nerviosos en los animales involucra la alteración del potencial eléctrico de las fibras nerviosas, y la alteración sólo es posible por la utilización de energía, en forma de ATP. Los cambios en el potencial eléctrico también caracterizan la actividad muscular. Los electrocardiogramas son posibles por la medida de los cambios eléçtricos en los latidos del corazón. El electroencefalograma mide las variaciones en los impulsos eléctricos del cerebro y ayuda en el diagnóstico de los desórdenes cerebrales. Algunos animales, como las anguilas, producen descargas eléctricas. Biolumniscencia. La luz es otra forma de energía utilizada por los organismos, por ejemplo, las luciérnagas y los cocuyos utilizan señales luminosas entre los sexos para encontrarse en la oscuridad. Los llamados fuegos fatuos no son más que bacterias luminiscentes que están descomponiendo la madera y las hojas en el bosque.

9
La célula microbiana utiliza la energía química para :
Sintetizar grandes moléculas a partir de otras más pequeñas. Transportar sustancias hacia la célula microbiana y organizarlas en su interior. Sacar las sustancias de desecho de la célula microbiana o para realizar la secreción La célula microbiana utiliza la energía química transportada por el ATP para : Sintetizar grandes moléculas a partir de otras más pequeñas. Transportar sustancias hacia la célula microbiana y organizarlas en su interior. El transporte activo de materiales a través de la membrana plasmática, puede también clasificarse como una utilización de la energía en movimiento. Sacar las sustancias de desecho de la célula microbiana o para realizar la secreción El trabajo mecánico de las célula microbianas. Cuando una ameba se desliza, cuando un paramecio nada, cuando una euglena se estira, cuando un microorganismo se divide, se gasta ATP. El trabajo mecánico de las célula microbianas.

10
Transporte de Nutrientes. Difusión simple o pasiva Difusión facilitada
Traslocación en grupo Transporte activo Transporte de Nutrientes.La célula microbiana requiere energía para el transporte de iones o de otros solutos a través de la membrana celular de una concentración baja a otra mayor. Excepto el agua y algunas moléculas solubles en lípidos, son pocos los compuestos que entran a la célula microbiana por difusión simple o pasiva. En este proceso el soluto cruza la membrana gracias al movimiento al azar de las moléculas, sin interactuar con las sustancias de la membrana (Figura 49 A). Otro mecanismo por el cual las sustancias cruzan la membrana semipermeable es la difusión facilitada. En esta, el soluto se combina reversiblemente en con una molécula portadora específica vinculada a la membrana. El complejo soluto-portador va y viene entre la superficie externa e interna de la membrana desprendiendo y ligando alternativamente el soluto (Figura 49 B). La difusión simple y la difusión facilitada no requieren energía metabólica porque no acumulan el soluto contra un gradiente electroquímico u osmótico. Pero otros mecanismos de transporte a través de la membrana como la traslocación en grupo y el transporte activo si requieren energía. En la traslocación en grupo se forma un enlace covalente entre el soluto y una molécula de la membrana, generalmente una enzima. El soluto es modificado químicamente y solo se regenera en el interior de la célula microbiana al ser separado del complejo soluto - transportador con suministro de energía(Figura 49 C). En el transporte activo el soluto se acumula contra un gradiente osmótico o electroquímico mediado por energía metabólica. Se diferencia de la difusión facilitada en que el transporte activo requiere el acoplamiento de un proceso de liberación de energía. Las célula microbianas toman casi todos los solutos, como azúcares, aminoácidos, péptidos, iones y nucleósidos, por transporte activo.
. Otro mecanismo por el cual las sustancias cruzan la membrana semipermeable es la difusión facilitada. En esta, el soluto se combina reversiblemente en con una molécula portadora específica vinculada a la membrana. El complejo soluto-portador va y viene entre la superficie externa e interna de la membrana desprendiendo y ligando alternativamente el soluto (Figura 49 B). La difusión simple y la difusión facilitada no requieren energía metabólica porque no acumulan el soluto contra un gradiente electroquímico u osmótico. Pero otros mecanismos de transporte a través de la membrana como la traslocación en grupo y el transporte activo si requieren energía. En la traslocación en grupo se forma un enlace covalente entre el soluto y una molécula de la membrana, generalmente una enzima. El soluto es modificado químicamente y solo se regenera en el interior de la célula microbiana al ser separado del complejo soluto - transportador con suministro de energía(Figura 49 C). En el transporte activo el soluto se acumula contra un gradiente osmótico o electroquímico mediado por energía metabólica. Se diferencia de la difusión facilitada en que el transporte activo requiere el acoplamiento de un proceso de liberación de energía. Las célula microbianas toman casi todos los solutos, como azúcares, aminoácidos, péptidos, iones y nucleósidos, por transporte activo.")
11
OBTENCIÓN DE LA ENERGÍA CELULAR
La célula microbiana obtiene su energía de dos maneras : Degradando compuestos y liberando su energía Almacenando la energía lumínica del sol mediante el proceso de fotosíntesis. OBTENCIÓN DE LA ENERGÍA CELULAR La célula microbiana obtiene su energía de dos maneras : degradando compuestos y liberando su energía o almacenando la energía lumínica del sol mediante el proceso de fotosíntesis. La energía almacenada en la fotosíntesis puede ser liberada posteriormente, mediante el proceso de respiración.

12
Los procesos por los cuales los microorganismos obtienen su energía son:
FOTOSÍNTESIS QUIMIOSÍNTESIS RESPIRACIÓN Aeróbica Anaeróbica Fermentación

13
FOTOSÍNTESIS

14
Naturaleza de la luz La naturaleza de la luz
La luz blanca se descompone en diferentes colores (color = longitud de onda) cuando pasa por un prisma. La longitud de onda se define como la distancia de pico a pico (o de valle a valle). La energía es inversamente proporcional a la longitud de onda: longitudes de onda larga tienen menor energía que las cortas. La distribución de los colores en el espectro esta determinado por la longitud de onda de cada uno de ellos. La luz visible es una pequeña parte del espectro electromagnético. Cuanto más larga la longitud de onda de la luz visible tanto más rojo el color. Asimismo las longitudes de onda corta están en la zona violeta del espectro. Las longitudes de onda mas largas que las del rojo se denominan infrarrojas, yaquellas mas cortas que el violeta, ultravioletas. La luz tiene una naturaleza dual: se comporta como onda y partícula. Entre las propiedades de la onda luminosa se incluyen la refracción de la onda cuando pasa de un material a otro. El efecto fotoeléctrico demuestra el comportamiento de la luz como partícula. El zinc se carga positivamente cuando es expuesto a luz ultravioleta en razón de que la energía de las partículas luminosas eliminan electrones del zinc. Estos electrones pueden crear una corriente eléctrica. El sodio, potasio y selenio tienen longitudes de onda críticas en el rango de la luz visible. La longitud de onda crítica es la mayor longitud de onda (visible o no) que puede causar un efecto fotoeléctrico. Albert Einstein desarrollo en la teoría de que la luz estaba compuesta de unas partículas denominadas fotones, cuya energía era inversamente proporcional a la longitud de onda de la luz. La Luz por lo tanto tiene propiedades explicables tanto por el modelo ondulatorio como por el corpuscular.
 cuando pasa por un prisma. La longitud de onda se define como la distancia de pico a pico (o de valle a valle). La energía es inversamente proporcional a la longitud de onda: longitudes de onda larga tienen menor energía que las cortas. La distribución de los colores en el espectro esta determinado por la longitud de onda de cada uno de ellos. La luz visible es una pequeña parte del espectro electromagnético. Cuanto más larga la longitud de onda de la luz visible tanto más rojo el color. Asimismo las longitudes de onda corta están en la zona violeta del espectro. Las longitudes de onda mas largas que las del rojo se denominan infrarrojas, yaquellas mas cortas que el violeta, ultravioletas. La luz tiene una naturaleza dual: se comporta como onda y partícula. Entre las propiedades de la onda luminosa se incluyen la refracción de la onda cuando pasa de un material a otro. El efecto fotoeléctrico demuestra el comportamiento de la luz como partícula. El zinc se carga positivamente cuando es expuesto a luz ultravioleta en razón de que la energía de las partículas luminosas eliminan electrones del zinc. Estos electrones pueden crear una corriente eléctrica. El sodio, potasio y selenio tienen longitudes de onda críticas en el rango de la luz visible. La longitud de onda crítica es la mayor longitud de onda (visible o no) que puede causar un efecto fotoeléctrico. Albert Einstein desarrollo en 1905 la teoría de que la luz estaba compuesta de unas partículas denominadas fotones, cuya energía era inversamente proporcional a la longitud de onda de la luz. La Luz por lo tanto tiene propiedades explicables tanto por el modelo ondulatorio como por el corpuscular.")
15
En presencia de luz y clorofila
FOTOSÍNTESIS (CH2O )x + O2 + H2O Carbohidrato CO H2O En presencia de luz y clorofila Los autótrofos : las algas y algunas bacterias, absorben la energía solar y la utilizan para reorganizar los átomos de las moléculas del agua y del bióxido de carbono atmosférico y formar con ellos nuevos compuestos con enlaces químicos ricos energía. El producto de esta reagrupación es oxígeno y compuestos orgánicos que suministran la energía química necesaria para plantas y animales. El proceso por el cual estos organismos usan la luz (energía radiante) para convertir el dióxido de carbono a carbohidratos se llama fotosíntesis. La función de la luz es desdoblar el agua. La fotosíntesis consiste básicamente en la transferencia del hidrógeno del agua al dióxido de carbono (CO2). En este caso el agua es el donador de hidrógeno, y el dióxido de carbono es el aceptor de hidrógeno. Se necesitan cuatro átomos de hidrógeno ( de dos moléculas de agua) por cada molécula de bióxido de carbono. Finalmente, todo el oxígeno que se produce en la fotosíntesis procede del desdoblamiento del agua y no del bióxido ce carbono. El proceso fotosintético se puede resumir en la siguiente ecuación : En presencia de luz y clorofila: CO H2O (CH2O )x + O2 + H2O En esta reacción (CH2O )x representa la unidad fundamental de la estructura de los carbohidratos y no se refiere a un compuesto en particular. Esta ecuación sólo indica los reactivos iniciales y los productos resultantes, aunque en la reacción haya entre ellos muchos pasos intermedios. Note, además, que esta reacción es exactamente el reverso de la reacción de la respiración aerobia. La fotosíntesis es el proceso que convierte la energía lumínica en energía química La fotosíntesis incluye dos tipos de reacciones : reacciones en la luz y reacciones en la oscuridad. Las reacciones de oscuridad pueden o no realizarse en presencia de luz, su nombre se usa para indicar que no necesitan luz para realizarse. Durante el día se realizan las reacciones de luz y, simultáneamente, las de oscuridad. Examinaremos a continuación la fase lumínica y la fase oscura de la fotosíntesis La fotosíntesis es el proceso que convierte la energía lumínica en energía química
x + O2 + H2O. Carbohidrato. CO2 + 2 H2O. En presencia de luz y clorofila. Los autótrofos : las algas y algunas bacterias, absorben la energía solar y la utilizan para reorganizar los átomos de las moléculas del agua y del bióxido de carbono atmosférico y formar con ellos nuevos compuestos con enlaces químicos ricos energía. El producto de esta reagrupación es oxígeno y compuestos orgánicos que suministran la energía química necesaria para plantas y animales. El proceso por el cual estos organismos usan la luz (energía radiante) para convertir el dióxido de carbono a carbohidratos se llama fotosíntesis. La función de la luz es desdoblar el agua. La fotosíntesis consiste básicamente en la transferencia del hidrógeno del agua al dióxido de carbono (CO2). En este caso el agua es el donador de hidrógeno, y el dióxido de carbono es el aceptor de hidrógeno. Se necesitan cuatro átomos de hidrógeno ( de dos moléculas de agua) por cada molécula de bióxido de carbono. Finalmente, todo el oxígeno que se produce en la fotosíntesis procede del desdoblamiento del agua y no del bióxido ce carbono. El proceso fotosintético se puede resumir en la siguiente ecuación : En presencia de luz y clorofila: CO2 + 2 H2O (CH2O )x + O2 + H2O. En esta reacción (CH2O )x representa la unidad fundamental de la estructura de los carbohidratos y no se refiere a un compuesto en particular. Esta ecuación sólo indica los reactivos iniciales y los productos resultantes, aunque en la reacción haya entre ellos muchos pasos intermedios. Note, además, que esta reacción es exactamente el reverso de la reacción de la respiración aerobia. La fotosíntesis es el proceso que convierte la energía lumínica en energía química. La fotosíntesis incluye dos tipos de reacciones : reacciones en la luz y reacciones en la oscuridad. Las reacciones de oscuridad pueden o no realizarse en presencia de luz, su nombre se usa para indicar que no necesitan luz para realizarse. Durante el día se realizan las reacciones de luz y, simultáneamente, las de oscuridad. Examinaremos a continuación la fase lumínica y la fase oscura de la fotosíntesis. La fotosíntesis es el proceso que convierte la energía lumínica en energía química.")
16
Estructura del cloroplasto y de las membranas fotosintéticas.
Los organismos fotosintéticos procariotes y eucariotes poseen sacos aplanados o vesículas llamadas tilacoides, que contienen los pigmentos fotosintéticos Pero solamente los cloroplastos de los eucariotes están rodeados por una doble membrana. ESTRUCTURA DEL CLOROPLASTO Y DE LAS MEMBRANAS FOTOSINTÉTICAS. La unidad estructural de la fotosíntesis es el cloroplasto. Los organismos fotosintéticos procariotes y eucariotes poseen sacos aplanados o vesículas llamadas tilacoides, que contienen los pigmentos fotosintéticos; pero solamente los cloroplastos de los eucariotes están rodeados por una doble membrana. Los tilacoides se disponen como una pila de panquecas, que recibe el nombre de grana. El interior del cloroplasto entre las granas es el estroma proteico, donde se encuentran las enzimas que catalizan la fijación del CO2 . Las mitocondrias constituyen un sistema con dos membranas como los cloroplastos, pero los cloroplastos tienen tres compartimentos: el estroma, el espacio tilacoidal y el espacio entre las membranas. El cloroplasto en su interior tiene un ADN circular y ribosomas

17
Excitación de la molécula de clorofila

19
FASES DE LA FOTOSÍNTESIS FASES DE LA FOTOSÍNTESIS
La fotosíntesis es un proceso que ocurre en dos fases. La primera fase es un proceso que depende de la luz (reacciones luminosas), requiere la energía directa de la luz que genera los transportadores que son utilizados en la segunda fase. La fase independiente de la luz (reacciones de oscuridad), se realiza cuando los productos de las reacciones de luz son utilizados para formar enlaces covalentes carbono-carbono (C-C), de los carbohidratos. Las reacciones oscuras pueden realizarse en la oscuridad, con la condición de que la fuente de energía (ATP) y el poder reductor (NADPH) formados en la luz se encuentren presentes. Investigaciones recientes sugieren que varias enzimas del ciclo de Calvin, son activadas por la luz mediante la formación de grupos -SH ; de tal forma que el termino reacción de oscuridad no es del todo correcto. Las reacciones de oscuridad se efectúan en el estroma; mientras que las de luz ocurren en los tilacoides
, requiere la energía directa de la luz que genera los transportadores que son utilizados en la segunda fase. La fase independiente de la luz (reacciones de oscuridad), se realiza cuando los productos de las reacciones de luz son utilizados para formar enlaces covalentes carbono-carbono (C-C), de los carbohidratos. Las reacciones oscuras pueden realizarse en la oscuridad, con la condición de que la fuente de energía (ATP) y el poder reductor (NADPH) formados en la luz se encuentren presentes. Investigaciones recientes sugieren que varias enzimas del ciclo de Calvin, son activadas por la luz mediante la formación de grupos -SH ; de tal forma que el termino reacción de oscuridad no es del todo correcto. Las reacciones de oscuridad se efectúan en el estroma; mientras que las de luz ocurren en los tilacoides.")
20
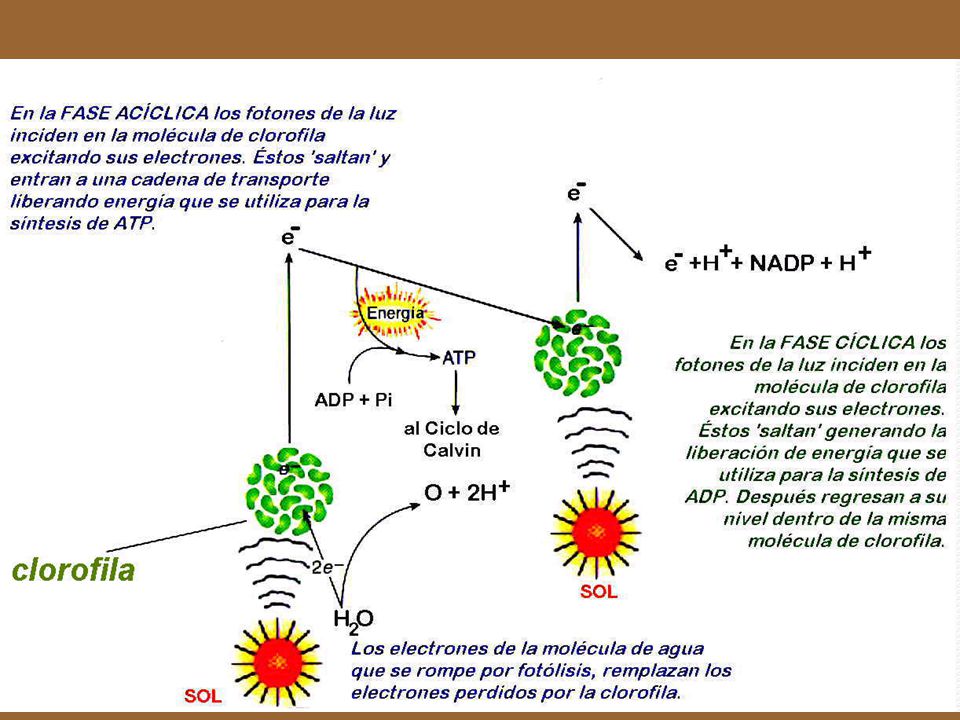
Fase lumínica 12 H2O 18 ATP+ 12 NADPH2 + 6 O2 ADP NADP Clorofila
La fase lumínica de la fotosíntesis es la “fuente de energía” de todo el proceso La clorofila es el pigmento que absorbe la luz. La clorofila está localizada en los cloroplastos de las células microbianas fotosintéticas, como las algas y las bacterias fotosintéticas. Los cloroplastos contienen además enzimas y otros catalizadores necesarios para convertir la energía lumínica en energía química. En efecto, todo el proceso de la fotosíntesis tiene lugar dentro del cloroplasto. Cuando las moléculas absorben energía lumínica, se “excitan” y transfieren a los electrones la energía ganada. Las moléculas de clorofila al excitarse liberan electrones de alta energía. Otras moléculas, los transportadores, recogen estos electrones de alta energía y forman enlaces ricos en energía química. La energía lumínica se convierte en energía química en el momento en que el electrón se transfiere de una molécula de clorofila excitada a la molécula del compuesto transportador. El compuesto transportador que adquiere estos electrones, puede, a su vez, transferirlos a otras moléculas y así liberar su energía química recientemente ganada, la cual será utilizada para formar ATP y NADPH2 ( Nicotinamida-adenina-dinucleótido o niacina-adenina-fosfato). Estas dos sustancias, ATP y NADPH2, provienen de la transformación de la energía luminosa en energía química. Como recordará, el ATP es la forma más común de energía circulante en la célula microbiana. El NADP es un transportador de hidrógeno en las célula microbiana y cuando se combina con él se convierte en NADPH2. Cuando las célula microbianas forman nuevos compuestos orgánicos, las necesidades de átomos de hidrógeno generalmente son suministradas por el NADPH2. Para formar una molécula de glucosa a partir de seis moléculas de CO2 se requieren 18 moléculas de ATP y 12 de NADPH2. De dónde procede el H2 para el NADPH ? Doce moléculas de agua ceden los electrones para la formación de ATP y el hidrógeno para la formación del NADPH2, quedan entonces 12 átomos de oxígeno. Éstos se combinan formando moléculas de oxígeno (O2), que escapan como gas. En resumen, los productos de la fase lumínica son NADPH2 y ATP y un subproducto que no interviene en el resto de las reacciones fotosintéticas, el O2. 12 H2O 18 ATP+ 12 NADPH O2 ADP NADP
. Estas dos sustancias, ATP y NADPH2, provienen de la transformación de la energía luminosa en energía química. Como recordará, el ATP es la forma más común de energía circulante en la célula microbiana. El NADP es un transportador de hidrógeno en las célula microbiana y cuando se combina con él se convierte en NADPH2. Cuando las célula microbianas forman nuevos compuestos orgánicos, las necesidades de átomos de hidrógeno generalmente son suministradas por el NADPH2. Para formar una molécula de glucosa a partir de seis moléculas de CO2 se requieren 18 moléculas de ATP y 12 de NADPH2. De dónde procede el H2 para el NADPH Doce moléculas de agua ceden los electrones para la formación de ATP y el hidrógeno para la formación del NADPH2, quedan entonces 12 átomos de oxígeno. Éstos se combinan formando moléculas de oxígeno (O2), que escapan como gas. En resumen, los productos de la fase lumínica son NADPH2 y ATP y un subproducto que no interviene en el resto de las reacciones fotosintéticas, el O2. 12 H2O. 18 ATP+ 12 NADPH2 + 6 O2. ADP. NADP.")
21
Productos de la fase lumínica y reacciones de la fase oscura
6CO ATP + 12 NADPH2 Enzima C6H12O6—P+ 18 ADP + 17 P inorgánico + 12 NADP Hexosa 1.1.1 Fase oscura Las reacciones de esta fase se llaman Ciclo del carbono, ciclo de Calvin o ciclo del C3. Estas reacciones se llevan a cabo sin necesidad de energía lumínica, pero se utiliza la energía almacenada durante la fase lumínica en el NADPH2 y en el ATP (Figura 41 ). El ciclo del carbono puede resumirse de la siguiente manera : Enzima 6CO ATP + 12 NADPH2 Hexosa—P+ 18 ADP + 17 P inorgánico + 12 NADP C6H12O6
. El ciclo del carbono puede resumirse de la siguiente manera : Enzima. 6CO ATP + 12 NADPH2 Hexosa—P+ 18 ADP + 17 P inorgánico + 12 NADP. C6H12O6.")
22
Ciclo de Calvin o del C3 CO2 H2O H2O ATP NADPH2 Pi Ribulosa 1 fosfato
Ribulosa 1,5 difosfato CO2 H2O Compuesto inestable Fructosa 6-fosfato GLUCOSA Acido 3-difosfoglicérico H2O 3-fosfo gliceraldehído Las principales etapas de la fase oscura se pueden sintetizar así : La ribulosa, una pentosa de cinco carbones ( 5C ), se produce continuamente en la célula microbiana : Actúa como una plantilla en la captación del CO2 y luego se recupera. En el ciclo del carbono se forman compuestos inestables que rápidamente se transforman en otros. En presencia de agua, tres moléculas de CO2 reacciona con 3 moléculas de la ribulosa para formar tres moléculas de un compuesto intermedio inestable de seis carbonos ( 6C ). El compuesto intermedio inestable de seis carbonos se rompe en dos, formando seis moléculas de ácido 3 fosfoglicérico, de 3 carbonos ( 3 C) El ácido 3 fosfoglicérico recibe los H y la energía de la fase lumínica, transportados por el ATP y NADPH2 , y se convierte primero en 6 moléculas de ácido 1,3 difosfoglicérico, que luego se convierten en seis moléculas 3 - fosfogliceraldehído De las seis moléculas 3 - fosfogliceraldehído se utilizan cinco para regenerar el compuesto de cinco carbonos que sirve de plantilla (la ribulosa) para la captación de CO2 y queda una molécula de 3 - fosfogliceraldehído. El ciclo se debe cumplir dos veces para que dos moléculas de 3 - fosfogliceraldehído se conviertan en una molécula de un azúcar sencillo como la fructosa 6-fosfato y esta a su vez se convierta en glucosa, como producto final principal de la fotosíntesis. Algunos de los compuestos intermedios, como el ácido fosfoglicérico pueden seguir otra vía metabólica y ser transformados en ácidos orgánicos, lípidos, aminoácidos y ácidos nucleicos ( Figura 51). La glucosa formada en la fotosíntesis se convierte, por procesos enzimáticos en otras sustancias de los microorganismos : lípidos (grasa, aceites), proteínas (hormonas, enzimas), carbohidratos (glicógeno, celulosa) y vitaminas. Algunos de ellos pueden ser aprovechados más tarde en procesos de respiración aeróbica o anaeróbica para extraerles su energía NADPH2 Pi Acido 1,3-difosfoglicérico ADP +NADP
, se produce continuamente en la célula microbiana : Actúa como una plantilla en la captación del CO2 y luego se recupera. En el ciclo del carbono se forman compuestos inestables que rápidamente se transforman en otros. En presencia de agua, tres moléculas de CO2 reacciona con 3 moléculas de la ribulosa para formar tres moléculas de un compuesto intermedio inestable de seis carbonos ( 6C ). El compuesto intermedio inestable de seis carbonos se rompe en dos, formando seis moléculas de ácido 3 fosfoglicérico, de 3 carbonos ( 3 C) El ácido 3 fosfoglicérico recibe los H y la energía de la fase lumínica, transportados por el ATP y NADPH2 , y se convierte primero en 6 moléculas de ácido 1,3 difosfoglicérico, que luego se convierten en seis moléculas 3 - fosfogliceraldehído. De las seis moléculas 3 - fosfogliceraldehído se utilizan cinco para regenerar el compuesto de cinco carbonos que sirve de plantilla (la ribulosa) para la captación de CO2 y queda una molécula de 3 - fosfogliceraldehído. El ciclo se debe cumplir dos veces para que dos moléculas de 3 - fosfogliceraldehído se conviertan en una molécula de un azúcar sencillo como la fructosa 6-fosfato y esta a su vez se convierta en glucosa, como producto final principal de la fotosíntesis. Algunos de los compuestos intermedios, como el ácido fosfoglicérico pueden seguir otra vía metabólica y ser transformados en ácidos orgánicos, lípidos, aminoácidos y ácidos nucleicos ( Figura 51). La glucosa formada en la fotosíntesis se convierte, por procesos enzimáticos en otras sustancias de los microorganismos : lípidos (grasa, aceites), proteínas (hormonas, enzimas), carbohidratos (glicógeno, celulosa) y vitaminas. Algunos de ellos pueden ser aprovechados más tarde en procesos de respiración aeróbica o anaeróbica para extraerles su energía. NADPH2. Pi. Acido. 1,3-difosfoglicérico. ADP +NADP.")
23
Fotosíntesis: reacciones de luz y oscuridad
La fotosíntesis incluye dos tipos de reacciones : reacciones en la luz y reacciones en la oscuridad. Las reacciones de oscuridad pueden o no realizarse en presencia de luz, su nombre se usa para indicar que no necesitan luz para realizarse. Durante el día se realizan las reacciones de luz y, simultáneamente, las de oscuridad. Examinaremos a continuación la fase lumínica y la fase oscura de la fotosíntesis

24
La conversión de CO2 en compuestos orgánicos requiere energía.
Por medio del ciclo del carbono, todas las célula microbianas, autótrofas o no, pueden utilizar el bióxido de carbono. La conversión de CO2 en compuestos orgánicos requiere energía. NO FOTOSINTÉTICOS Obtienen energía para sintetizar compuestos orgánicos del desdoblamiento de otros compuestos orgánicos preexistentes. No hay ganancia en la cantidad total de compuestos orgánicos. Transforman biomasa. FOTOSINTÉTICOS Forman compuestos orgánicos durante la fotosíntesis Utilizan energía procedente de la luz Aumentan la cantidad total de compuestos orgánicos. Sintetizan biomasa. Por medio del ciclo del carbono, todas las célula microbianas pueden utilizar el bióxido de carbono. ¿Cuál es la diferencia entre la forma como los fotosíntéticos y los no fotosintéticos fijan el carbono ? Existe una diferencia importante. La conversión de CO2 en compuestos orgánicos requiere energía. Los no fotosintéticos (animales, protozoos, hongos, etc.) pueden obtener energía para sintetizar los compuestos orgánicos mediante el desdoblamiento de otros compuestos orgánicos preexistentes. Por lo tanto no hay ganancia en la cantidad total de compuestos orgánicos. Pero cuando los fotosintéticos (algas, plantas, bacterias fotosíntéticas) forman compuestos orgánicos durante la fotosíntesis, utilizan energía procedente de la luz y, por consiguiente, pueden aumentar la cantidad total de compuestos orgánicos. A diferencia de la fotosíntesis, los otros procesos de los microorganismos no pueden utilizar la energía solar directamente y sólo pueden hacerlo después de que los organismos fotosintéticos la han convertido en energía química. Por consiguiente, la fotosíntesis se considera como un puente entre la energía del sol y la energía necesaria para la vida en la Tierra.
 pueden obtener energía para sintetizar los compuestos orgánicos mediante el desdoblamiento de otros compuestos orgánicos preexistentes. Por lo tanto no hay ganancia en la cantidad total de compuestos orgánicos. Pero cuando los fotosintéticos (algas, plantas, bacterias fotosíntéticas) forman compuestos orgánicos durante la fotosíntesis, utilizan energía procedente de la luz y, por consiguiente, pueden aumentar la cantidad total de compuestos orgánicos. A diferencia de la fotosíntesis, los otros procesos de los microorganismos no pueden utilizar la energía solar directamente y sólo pueden hacerlo después de que los organismos fotosintéticos la han convertido en energía química. Por consiguiente, la fotosíntesis se considera como un puente entre la energía del sol y la energía necesaria para la vida en la Tierra.")
25
RESPIRACIÓN: DESASIMILACIÓN DE COMPUESTOS ORGÁNICOS Y LIBERACIÓN DE ENERGÍA
El proceso por el cual la célula microbiana libera la energía almacenada en los alimentos se llama respiración celular. En la mayoría de los microorganismos, el oxígeno es necesario para este proceso, por lo cual se le llama respiración aerobia. En las célula microbianas que obtienen su energía sin presencia de oxígeno, el proceso se llama respiración anaerobia.

26
RESPIRACIÓN: Proceso por el cual la célula microbiana libera la energía almacenada en los alimentos.
Este proceso ocurre en las mitocondrias en la mayoría de las células eucariotes o en la membrana celular de las células procariotes

27
Tipos de respiración Respiración aerobia.
Respiración anaerobia (Fermentación).
.")
28
Respiración aeróbica (Rutas de utilización del piruvato por aerobios)
RESPIRACIÓN AEROBIA C6H12O6 + 6 O2 Enzimas 6CO2 +6 H2O+Energía (38 ATP) G = -686 Kcal Respiración aeróbica (Rutas de utilización del piruvato por aerobios) En la respiración aeróbica, la glucosa es la principal fuente primaria de energía de los alimentos. La degradación completa de la molécula de glucosa puede ser resumida por la ecuación: Enzimas C6H12O6 + 6 O2 6 CO2 +6 H2O+Energía (38 ATP) G = -686 Kcal La glucosa puede ser quemada y la energía contenida en ella es liberada con una considerable cantidad de calor. Este proceso llamado combustión y sucede fuera de las célula microbianas vivas. Una célula microbiana se destruiría si la cantidad de energía almacenada en la molécula de glucosa se libera de una vez. En la célula microbiana, la energía se libera gradualmente en etapas y se transforma en energía potencial, almacenada en las moléculas de ATP. La glucosa, tanto en el metabolismo respiratorio como en el fermentativo, se transforma en ácido pirúvico por una de las siguientes rutas: 1. Embden-Meyerhof (eucariotas y procariotas) 2. Pentosa fosfato (eucariotas y procariotas) 3. Entner Doudoroff (sólo en ciertos procariotas como alternativa a la ruta Embden-Meyerhof
 G = -686 Kcal. Respiración aeróbica (Rutas de utilización del piruvato por aerobios) En la respiración aeróbica, la glucosa es la principal fuente primaria de energía de los alimentos. La degradación completa de la molécula de glucosa puede ser resumida por la ecuación: Enzimas. C6H12O6 + 6 O2 6 CO2 +6 H2O+Energía (38 ATP) G = -686 Kcal. La glucosa puede ser quemada y la energía contenida en ella es liberada con una considerable cantidad de calor. Este proceso llamado combustión y sucede fuera de las célula microbianas vivas. Una célula microbiana se destruiría si la cantidad de energía almacenada en la molécula de glucosa se libera de una vez. En la célula microbiana, la energía se libera gradualmente en etapas y se transforma en energía potencial, almacenada en las moléculas de ATP. La glucosa, tanto en el metabolismo respiratorio como en el fermentativo, se transforma en ácido pirúvico por una de las siguientes rutas: 1. Embden-Meyerhof (eucariotas y procariotas) 2. Pentosa fosfato (eucariotas y procariotas) 3. Entner Doudoroff (sólo en ciertos procariotas como alternativa a la ruta Embden-Meyerhof.")
29
Rata de producción de energía por combustión y por respiración celular
Comparación entre la rata de producción de energía por combustión y por respiración celular Por ejemplo, en la respiración aerobia para la conversión de la glucosa hasta sus productos finales, dióxido de carbono y agua, hay alrededor de 20 pasos intermedios, catalizados cada uno por una enzima específica. Para su aprendizaje organizamos los pasos de la respiración aeróbica en tres etapas: la glucólisis, el ciclo de Krebs y la cadena de transporte electrónico acoplada al final.

30
La glucólisis, ruta metabólica común a todos los organismos
LA GLUCÓLISIS (GLICÓLISIS) La degradación de la glucosa puede tener lugar por glucólisis, una de las rutas más importantes que utiliza la célula microbiana para producir energía. La glucólisis no requiere presencia de oxígeno y puede tener lugar tanto en célula microbianas aerobias como anaerobias. En ella la fructosa 1,6 difosfato que se forma a partir de la glucosa, se escinde en dos unidades de tres átomos de carbono (dihidroxiacetona fosfato y gliceraldehído 3-fosfato), que son oxidadas a su vez a ácido pirúvico. En el paso de oxidación del gliceraldehído 3-fosfato se pierden un par de electrones (dos átomos de hidrógeno). En ausencia de oxígeno, este par de electrones puede utilizarse para reducir el ácido pirúvico a ácido láctico o etanol. En presencia de oxígeno, entran en la cadena de transporte electrónico. Por cada molécula de glucosa metabolizada se consumen dos moléculas de ATP y se producen cuatro. El balance, por tanto, es de dos moléculas de ATP producidas en la glucólisis. La reacción general se puede esquematizar de la siguiente forma: C6H12O6 + 2NAD +2ADP+2 P 2 CH3—CO—COOH+2NADH2+ 2ATP Glucosa Nicotinamida Fosfato Ácido pirúvico adenina inorgánico dinucleótido Terminada la glucólisis la reacción puede seguir en dos vías : La respiración aeróbicao la respiración anaeróbica (fermentación)
 La degradación de la glucosa puede tener lugar por glucólisis, una de las rutas más importantes que utiliza la célula microbiana para producir energía. La glucólisis no requiere presencia de oxígeno y puede tener lugar tanto en célula microbianas aerobias como anaerobias. En ella la fructosa 1,6 difosfato que se forma a partir de la glucosa, se escinde en dos unidades de tres átomos de carbono (dihidroxiacetona fosfato y gliceraldehído 3-fosfato), que son oxidadas a su vez a ácido pirúvico. En el paso de oxidación del gliceraldehído 3-fosfato se pierden un par de electrones (dos átomos de hidrógeno). En ausencia de oxígeno, este par de electrones puede utilizarse para reducir el ácido pirúvico a ácido láctico o etanol. En presencia de oxígeno, entran en la cadena de transporte electrónico. Por cada molécula de glucosa metabolizada se consumen dos moléculas de ATP y se producen cuatro. El balance, por tanto, es de dos moléculas de ATP producidas en la glucólisis. La reacción general se puede esquematizar de la siguiente forma: C6H12O6 + 2NAD +2ADP+2 P 2 CH3—CO—COOH+2NADH2+ 2ATP. Glucosa Nicotinamida Fosfato Ácido pirúvico. adenina inorgánico. dinucleótido. Terminada la glucólisis la reacción puede seguir en dos vías : La respiración aeróbicao la respiración anaeróbica (fermentación)")
31
La glucólisis es una vía común para la producción de energía puesto que éste es un proceso activo, tanto cuando hay presencia de oxígeno ( Glucólisis aeróbica), como cuando no hay oxigeno disponible en la célula (glucólisis anaeróbica). Esta característica de la glucólisis es crucial porque es la que permite que las células sobrevivan y relizen trabajo, durante un determinado período de tiempo, en condiciones anaeróbicas o en situaciones " anóxicas. La glucólisis también es una vía metabólica para la oxidación de otros monosacáridos como la fructosa y la galactosa. Para los propósitos de este curso, lo más importante no es aprenderse los nombres de todos los intermediarios,ni de todas las enzimas, en las diferentes etapas de la glucólisis, sino comprender la estrategia general de este sub-proceso en relación con el proceso general de la respiración celula y reconocer los principales intermediarios del proceso para tener una visión general del mismo. La glucólisis es un proceso es "exergónico”( libera energía), porque es a la vez oxidativo y catabólico: 1. Oxidativo porque mediante el se le "quitan" electrones a la molécula de glucosa . En términos del metabolismo energético, los electrones que se le "extraen" a una molécula son "energía potencial" . produce energía en forma de ATP 2. Catabólico porque se "divide" la molécula de glucosa ( C6 ) en dos móleculas que contienen exactamente la mitad de átomos de carbono cada una (C3 ). Produce "energía potencial" ( ¡ojo!, este concepto, aquí no es el mismo utilizado por la física) y se trata de la energía que transportan los "portadores de electrones como el NADH , coenzima que participa activamente en este proceso. Cuando se completa la vía anaeróbica de la glucólisis, el NADH que se produce en las etapas intermedias del proceso, se reinvierte en la etapa final , por lo tanto la producción de NADH neto está condicionada a si el proceso de producción de energía es aeróbico o anaeróbico.
, porque es a la vez oxidativo y catabólico: 1. Oxidativo porque mediante el se le quitan electrones a la molécula de glucosa . En términos del metabolismo energético, los electrones que se le extraen a una molécula son energía potencial . produce energía en forma de ATP. 2. Catabólico porque se divide la molécula de glucosa ( C6 ) en dos móleculas que contienen exactamente la mitad de átomos de carbono cada una (C3 ). Produce energía potencial ( ¡ojo!, este concepto, aquí no es el mismo utilizado por la física) y se trata de la energía que transportan los portadores de electrones como el NADH , coenzima que participa activamente en este proceso. Cuando se completa la vía anaeróbica de la glucólisis, el NADH que se produce en las etapas intermedias del proceso, se reinvierte en la etapa final , por lo tanto la producción de NADH neto está condicionada a si el proceso de producción de energía es aeróbico o anaeróbico.")
32
A partir la glicólisis pueden darse la respiración aerobia o la anaerobia.
En la glucólisis, tal como indica la figura, la fructosa 1,6 difosfato que se forma a partir de la glucosa, se escinde en dos unidades de tres átomos de carbono (dihidroxiacetona fosfato y gliceraldehído 3-fosfato), que son oxidadas a su vez a ácido pirúvico. En el paso de oxidación del gliceraldehído 3-fosfato se pierden un par de electrones (dos átomos de hidrógeno). En ausencia de oxígeno, este par de electrones puede utilizarse para reducir el ácido pirúvico a ácido láctico o etanol. En presencia de oxígeno, este par de electrones puede entrar en la cadena de transporte electrónico como veremos más adelante. Por cada molécula de glucosa metabolizada se consumen dos moléculas de ATP y se producen cuatro. El balance, por tanto, es de dos moléculas de ATP producidas en la metabolización de una glucosa por glucólisis. La reacción general se puede esquematizar de la siguiente forma: C6H12O NAD + 2ADP P2CH3—CO—COOH + 2NADH ATP Glucosa Nicotinamida Fosfato Ácido pirúvico adenina inorgánico dinucleótido Cuando termina la glucólisis la reacción puede seguir en dos vías : La respiración aeróbica, en presencia de oxígeno, o la respiración anaeróbica (fermentación), en ausencia de oxígeno La glicólisis es una ruta metabólica común a todos los organismos, por eso a partir de ella pueden darse la respiración aerobia o la anaerobia.
, que son oxidadas a su vez a ácido pirúvico. En el paso de oxidación del gliceraldehído 3-fosfato se pierden un par de electrones (dos átomos de hidrógeno). En ausencia de oxígeno, este par de electrones puede utilizarse para reducir el ácido pirúvico a ácido láctico o etanol. En presencia de oxígeno, este par de electrones puede entrar en la cadena de transporte electrónico como veremos más adelante. Por cada molécula de glucosa metabolizada se consumen dos moléculas de ATP y se producen cuatro. El balance, por tanto, es de dos moléculas de ATP producidas en la metabolización de una glucosa por glucólisis. La reacción general se puede esquematizar de la siguiente forma: C6H12O6 + 2NAD + 2ADP + 2 P2CH3—CO—COOH + 2NADH2 + 2ATP. Glucosa Nicotinamida Fosfato Ácido pirúvico. adenina inorgánico. dinucleótido. Cuando termina la glucólisis la reacción puede seguir en dos vías : La respiración aeróbica, en presencia de oxígeno, o la respiración anaeróbica (fermentación), en ausencia de oxígeno. La glicólisis es una ruta metabólica común a todos los organismos, por eso a partir de ella pueden darse la respiración aerobia o la anaerobia.")
33
Ciclo de Krebs o Ciclo del ácido tricarboxílico (ATC)
La etapa del ciclo de Krebs comienza con el último producto de la glucólisis : el ácido pirúvico. Como en el paso anterior la molécula de glucosa se escinde en dos de gliceraldehido fosfato, al final se producen 2 moléculas de ácido pirúvico. O sea que por cada molécula de glucosa se repite dos veces el ciclo de del ácido tricarboxílico. El ciclo del ácido tricarboxílico (ATC) es una secuencia de reacciones que genera energía en forma de ATP y de moléculas de coenzimas reducidas (Nicotinamida-adenina-dinucleótido, Nadh2 Y Flavina adenina dinucleótido, FADH2). El ciclo empieza cuando el ácido pirúvico es parcialmente degradado, al perder una molécula de bióxido de carbono y dos hidrógenos y luego se combina con la coenzima A para formar acetil CoA.
 es una secuencia de reacciones que genera energía en forma de ATP y de moléculas de coenzimas reducidas (Nicotinamida-adenina-dinucleótido, Nadh2 Y Flavina adenina dinucleótido, FADH2). El ciclo empieza cuando el ácido pirúvico es parcialmente degradado, al perder una molécula de bióxido de carbono y dos hidrógenos y luego se combina con la coenzima A para formar acetil CoA.")
34
El ATC, También desempeña otras funciones
El ATC, También desempeña otras funciones. Muchos intermediarios del ciclo son también precursores de la biosíntesis de aminoácidos, purinas, pirimidinas, etc. Por lo tanto, el ciclo ATC es un ciclo anfibólico, lo cual significa que no funciona solamente en el catabolismo (degradación) sino también en reacciones anabólicas (síntesis). El ATC, También desempeña otras funciones. Muchos intermediarios del ciclo son también precursores de la biosíntesis de aminoácidos, purinas, pirimidinas, etc. Por lo tanto, el ciclo ATC es un ciclo anfibólico, lo cual significa que no funciona solamente en el catabolismo (degradación) sino también en reacciones anabólicas (síntesis).
 sino también en reacciones anabólicas (síntesis). El ATC, También desempeña otras funciones. Muchos intermediarios del ciclo son también precursores de la biosíntesis de aminoácidos, purinas, pirimidinas, etc. Por lo tanto, el ciclo ATC es un ciclo anfibólico, lo cual significa que no funciona solamente en el catabolismo (degradación) sino también en reacciones anabólicas (síntesis).")
35
2CO2 + CoA + 3NADH2 + FADH2 + ATP
El conjunto de reacciones del ciclo ATC se puede resumir en la siguiente forma: Acetil-CoA + 3H2O + 3NAD+ + FAD+ + ADP + Pi 2CO2 + CoA + 3NADH2 + FADH2 + ATP El conjunto de reacciones del ciclo ATC se puede resumir en la siguiente forma: Acetil-CoA + 3h2O + 3NAD+ + FAD+ + ADP + Pi 2CO2 + CoA + 3NADH2 + FADH2 + ATP Como la degradación glucolítica de una molécula de glucosa da lugar a dos de Acetil CoA, que pueden entrar en este ciclo, el resumen total será el doble del indicado en esta reacción, por molécula de glucosa degradada. Una molécula de glucosa da lugar a dos de acetil- CoA, que pueden entrar en este ciclo El total será el doble del indicado en esta reacción

36
Fosforilación oxidativa: formación directa de ATP a partir de ADP y Pi
Fosforilación oxidativa es la formación directa de ATP a partir de ADP y Pi, el proceso de síntesis tiene lugar en la interfase membrana interna y la matriz mitocondrial y es catalizado por una enzima denominada ATPasa. La ATPasa es una enzima compleja, que obtiene la energía necesaria para sintetizar cada molécula de ATP, a partir de un " gradiente de protones" entre el espacio intermembranal y la matriz de la mitocondria. La ATPasa utiliza energía electroquímica para trabajar. No utiliza calor, ni directamente la energía liberada por una reacción acoplada como suele ocurrir con las reacciones de biosíntesis; en su lugar aprovecha un "voltaje" o "delta" de potencial eléctrico que se crea por la diferencia entre la concentración de protones (H+) entre el lado externo y el lado interno dela membrana interna de la mitocondria: una verdadera pila voltáica biológica.
 entre el lado externo y el lado interno dela membrana interna de la mitocondria: una verdadera pila voltáica biológica.")
37
El flujo de electrones ocurre expontáneamente, puesto que la cadena de intermediarios que los "transportan" en la membrana, constituyen una "cascada" de potencial electrico. ¿Que quiere decir esto?. Las leyes del cosmos, hasta ahora estudiadas (las clásicas), nos demuestran un principio universal: los sistemas tienden a adquirir los estados energéticos mas bajos posibles. Eso explica porqué un cuerpo cae en la tierra, porqué los gases tienden a escapar de los recipientes que los contienen y a ocupar todo el volume disponible; porqué el agua de los rios corre hacia el mar; porqué un cuerpo transmite calor a otro, y en nuestro estudio particular, porqué existen las corrientes eléctricas: Las corrientes eléctricas se desatan debido a que, si se establece un puente entre dos conjuntos de cargas eléctricas cuyos estados energéticos son diferentes, el sistema conformado con el puente tiende a adquirir su menor estado energético posible; ésto lo logra cuando la concentración de cargas electricas en todo el sistema es homogénea. Para lograr esa homogeneidad, el exceso de cargas en un sitio se desplaza hasta otros sitios donde haya menor cantidad de cargas. Dicho desplazamiento de cargas desde un sitio a otro es lo que conocemos como "corriente eléctrica". Otro principio de la termodinámica, asociado al anterior es que cuando un sistema pasa del desequilibrio al equilibrio, dicho cambio es posible en tiempos finitos o cortos, solo si el sistema le entrega energía al entorno (aunque no lo vamos a discutir aquí, este principio se conoce como la segunda ley de la termodinámica y tiene que ver con el concepto de orden y desorden del universo). Esta "entrega" o liberación de energía al entorno depende de las características del puente que se ha establecido entre los dos sistemas en desequilibrio. En el caso de sistemas de cargas eléctricas el puente se conoce como "sistema de conductores y resistencias" o "cascada de potencial". Lo que estamos planteando es que las leyes de la termodinámica nos dicen varias cosas acerca de la corriente electrica: 1. Que ésta se presenta porque existe un sistema en desequilibrio que está tratando de llegar al equilibrio. 2. Que el sistema llega al equilibrio en un tiempo y en una serie de etapas finitas sólo si libera parte de su energía al entorno. 3. Las dos anteriores conclusiones llevan a una tercera, y es que un sistema en desequilibrio, con su correspondiente "cascada de potencial" o "puente", es una fuente de energía. Estos principios, entre otros, son los fundamentales para los sistemas de generación de energía. de hecho el término "cascada" se ha tomado del fenómeno natural de una caida de agua, de la cual es posible obtener energía para convertirla en energía rotacional , la cual se convierte en energía elétrica mediante un dispositivo magnético. En el caso de la cadena de transporte electrónico de la mitocondria celular , la cascada de potencial se forma entre las coenzimas NADH y FADH2 y la molécula de oxigeno. El sistema de conductores y resistencias es el grupo de enzimas y coenzimas oxidoreductasas que se listó en la tabla anterior La ATPasa utiliza el potencial eléctrico que se crea por la diferencia entre la concentración de protones (H+) entre el lado externo y el lado interno de la membrana interna de la mitocondria: una verdadera pila voltáica biológica
. Esta entrega o liberación de energía al entorno depende de las características del puente que se ha establecido entre los dos sistemas en desequilibrio. En el caso de sistemas de cargas eléctricas el puente se conoce como sistema de conductores y resistencias o cascada de potencial . Lo que estamos planteando es que las leyes de la termodinámica nos dicen varias cosas acerca de la corriente electrica: 1. Que ésta se presenta porque existe un sistema en desequilibrio que está tratando de llegar al equilibrio. 2. Que el sistema llega al equilibrio en un tiempo y en una serie de etapas finitas sólo si libera parte de su energía al entorno. 3. Las dos anteriores conclusiones llevan a una tercera, y es que un sistema en desequilibrio, con su correspondiente cascada de potencial o puente , es una fuente de energía. Estos principios, entre otros, son los fundamentales para los sistemas de generación de energía. de hecho el término cascada se ha tomado del fenómeno natural de una caida de agua, de la cual es posible obtener energía para convertirla en energía rotacional , la cual se convierte en energía elétrica mediante un dispositivo magnético. En el caso de la cadena de transporte electrónico de la mitocondria celular , la cascada de potencial se forma entre las coenzimas NADH y FADH2 y la molécula de oxigeno. El sistema de conductores y resistencias es el grupo de enzimas y coenzimas oxidoreductasas que se listó en la tabla anterior. La ATPasa utiliza el potencial eléctrico que se crea por la diferencia entre la concentración de protones (H+) entre el lado externo y el lado interno de la membrana interna de la mitocondria: una verdadera pila voltáica biológica.")
38
La siguiente curva representa La caida de potencial eléctrico desde el NAD hasta el oxígeno,receptor final de los electrones de la cadena respiratoria. ( Eo representa el potencial eléctrico en Voltios ; también se expresa la energía equivalente al potencial eléctrico en Kilocalorías).
..")
39
Los electrones son eliminados de los transportadores de energía por medio de la reducción de algún aceptor terminal de electrones como: el oxígeno (en la respiración aeróbica) nitrógeno, sulfato o dióxido de carbono (en la respiración anaeróbica). En la respiración los electrones son transferidos de manera secuencial a través de una serie de proteínas transportadoras adosadas a la membrana celular. Esta es la cadena de transporte de electrones.
 nitrógeno, sulfato o dióxido de carbono (en la respiración anaeróbica). En la respiración los electrones son transferidos de manera secuencial a través de una serie de proteínas transportadoras adosadas a la membrana celular. Esta es la cadena de transporte de electrones.")
40
CADENA DE TRANSPORTE ELECTRÓNICO :
NADH2 NAD+ Sustancia reducida Sustancia oxidada ATP FADH2 FAD FMN FMNH2 CoQ CoQH2 H+ Flavoproteína ADP + P Coenzima Q Citocromo b- Citocromo c1- Citocromo c- Citocromo a-/a3 Fe+2 Fe+3 O2 H2O Figura 45. Cadena de transporte electrónico CADENA DE TRANSPORTE ELECTRÓNICO : Secuencia de reacciones de oxidación-reducción para la generación de ATP. La cadena se acopla al ciclo de Krebs para convertir la energía liberada en él, en ATP--> fosforilación oxidativa. Capta electrones a partir de compuestos reducidos y los transfire al aceptor final, el oxígeno, con la consiguiente formación de agua. La cadena de transporte electrónico La cadena de transporte electrónico, también conocida como sistema de citocromos o cadena respiratoria, es una secuencia de reacciones de oxidación-reducción para la generación de ATP. La cadena se acopla al ciclo de Krebs para convertir la energía liberada en él, en ATP. Esta secuencia de reacciones funciona captando electrones a partir de compuestos reducidos (NADH2, FADH2, FMNH2, flavina mononucleótido reducido) y transfiriéndolos al oxígeno con la consiguiente formación de agua. El oxígeno es necesario como aceptor final de los electrones, por lo tanto las reacciones deben ser aerobias. En varios de los pasos intermedios se libera suficiente energía para sintetizar ATP a partir de ADP y fosfato inorgánico. Esta síntesis de ATP se denomina fosforilación oxidativa, debido a la formación de enlaces altamente energéticos en los que interviene el fosfato. En la figura se puede observar que los átomos de hidrógeno (electrones más protones, H+) que pierden las sustancias que se oxidan son transferidos por las deshidrogenasas que contienen NAD (nicotinamida-adenina-dinucleótido) o NADP (fosfato de nicotinamida-adenin-dinucleótido) y citocromos que contienen hierro, al oxígeno molecular con la siguiente formación de agua. Nótese así mismo, que en tres pasos específicos de la cadena, tiene lugar síntesis de ATP.
 y transfiriéndolos al oxígeno con la consiguiente formación de agua. El oxígeno es necesario como aceptor final de los electrones, por lo tanto las reacciones deben ser aerobias. En varios de los pasos intermedios se libera suficiente energía para sintetizar ATP a partir de ADP y fosfato inorgánico. Esta síntesis de ATP se denomina fosforilación oxidativa, debido a la formación de enlaces altamente energéticos en los que interviene el fosfato. En la figura se puede observar que los átomos de hidrógeno (electrones más protones, H+) que pierden las sustancias que se oxidan son transferidos por las deshidrogenasas que contienen NAD (nicotinamida-adenina-dinucleótido) o NADP (fosfato de nicotinamida-adenin-dinucleótido) y citocromos que contienen hierro, al oxígeno molecular con la siguiente formación de agua. Nótese así mismo, que en tres pasos específicos de la cadena, tiene lugar síntesis de ATP.")
41
Rendimiento total en ATP por molécula de glucosa
Rendimiento energético de la respiración aerobia El rendimiento energético de la degradación de una molécula de glucosa en condiciones aeróbicas, se produce cuando los electrones de las moléculas de coenzimas reducidas alimentan la cadena de transporte electrónico. Los electrones se transfieren escalonadamente desde las coenzimas transportadoras al oxígeno molecualr, que se acoplan a la formación de ATP por fosforilación oxidativa. Por cada molécula de glucosa degradada se producen 12 coenzimas reducidas: dos de FADH2 (una por cada vuelta del ciclo ATC) y 10 de NADH2 (dos de la glucólisis, dos del paso de la glucólisis al ciclo ATC, es decir, de ácido pirúvico a acetil CoA y seis de las dos vueltas del ciclo ATC). Como cada NADH2 genera tres moléculas de ATP y cada FADH2 dos moléculas de ATP aparecerán 34 moléculas de ATP por oxidación de estas coenzimas en la cadena respiratoria
 y 10 de NADH2 (dos de la glucólisis, dos del paso de la glucólisis al ciclo ATC, es decir, de ácido pirúvico a acetil CoA y seis de las dos vueltas del ciclo ATC). Como cada NADH2 genera tres moléculas de ATP y cada FADH2 dos moléculas de ATP aparecerán 34 moléculas de ATP por oxidación de estas coenzimas en la cadena respiratoria.")
42
C6H12O6 + 6O2 6CO2 + 6H2O + 38 ATP (= 686 Kcal)
El rendimiento total en ATP por respiración aeróbica a partir de una molécula de glucosa La oxidación completa de la glucosa, vía glucólisis, ciclo ATC y cadena respiratoria, se resume en la reacción siguiente: C6H12O O2 6CO H2O ATP (= 686 Kcal) El rendimiento total en ATP por respiración aeróbica a partir de una molécula de glucosa es de 38 moléculas: 34 por oxidación de coenzimas, dos en la glucólisis y dos por reacciones del ciclo ATC que originan dos moléculas de GTP. Este rendimiento total en ATP por molécula de glucosa se esquematiza en la figura 46. La oxidación completa de la glucosa, vía glucólisis, ciclo ATC y cadena respiratoria, se resume en la reacción siguiente: C6H12O O2 6CO H2O ATP (= 686 Kcal) Glucosa Bióxido de Carbono
 El rendimiento total en ATP por respiración aeróbica a partir de una molécula de glucosa es de 38 moléculas: 34 por oxidación de coenzimas, dos en la glucólisis y dos por reacciones del ciclo ATC que originan dos moléculas de GTP. Este rendimiento total en ATP por molécula de glucosa se esquematiza en la figura 46. La oxidación completa de la glucosa, vía glucólisis, ciclo ATC y cadena respiratoria, se resume en la reacción siguiente: C6H12O6 + 6O2 6CO2 + 6H2O + 38 ATP (= 686 Kcal) Glucosa Bióxido. de Carbono.")
44
RESPIRACIONES ANAEROBIAS
El oxígeno gaseoso no interviene. El aceptor de electrones es un compuesto distinto al oxígeno. Cuando el aceptor es un compuesto como orgánico se denomina fermentación Cuando es inorgánico respiración anaerobia

45
La respiración sin oxígeno, está restringida en gran parte a los saprófogos (bacterias, levaduras, mohos, protozoos).
.")
46
Fermentación

47
Alcohol etílico Dióxido de carbono Energía
GLUCOSA Glucólisis 2C3H4O3 (ácido pirúvico) H 2C2H5OH CO ATP Alcohol etílico Dióxido de carbono Energía Respiración anaerobia o fermentación Recuerde que la glucólisis es la primera etapa de la degradación de la glucosa, común a aerobios y anaerobios y que no requiere oxígeno. La glucosa es fraccionada durante la glucólisis en dos moléculas de ácido pirúvico, con la formación de ATP e hidrógeno. El hidrógeno liberado no puede quedar libre, debe haber un aceptor de ese hidrógeno. En ausencia de oxígeno, para actuar como aceptor final, el ácido pirúvico sirve a sí mismo como aceptor. El alcohol etílico se forma de la unión de los cuatro átomos de hidrógeno producidos en la glucólisis con el ácido pirúvico como sigue: 2C3H4O H 2C2H5OH CO ATP ácido Alcohol Dióxido Energía pirúvico etílico de carbono Como el ciclo de Krebs no está acoplado a estas reacciones, la energía producida por una molécula de glucosa es menor que la producida en la respiración aerobia (alrededor de un 5%). La energía restante queda en los enlaces de la molécula de alcohol. Muchos microorganismos como las levaduras y las bacterias realizan numerosos tipos de fermentaciones para producir energía. Por ejemplo, Streptococcus lactis, bacteria responsable de la acidificación de la leche, desasimila glucosa a ácido láctico como producto de fermentación. En ausencia de oxígeno, para actuar como aceptor final, el ácido pirúvico sirve a sí mismo como aceptor.
 + 4H. 2C2H5OH + 2CO2 +2 ATP. Alcohol etílico Dióxido de carbono Energía. Respiración anaerobia o fermentación. Recuerde que la glucólisis es la primera etapa de la degradación de la glucosa, común a aerobios y anaerobios y que no requiere oxígeno. La glucosa es fraccionada durante la glucólisis en dos moléculas de ácido pirúvico, con la formación de ATP e hidrógeno. El hidrógeno liberado no puede quedar libre, debe haber un aceptor de ese hidrógeno. En ausencia de oxígeno, para actuar como aceptor final, el ácido pirúvico sirve a sí mismo como aceptor. El alcohol etílico se forma de la unión de los cuatro átomos de hidrógeno producidos en la glucólisis con el ácido pirúvico como sigue: 2C3H4O3 + 4H 2C2H5OH + 2CO2 +2 ATP. ácido Alcohol Dióxido Energía. pirúvico etílico de. carbono. Como el ciclo de Krebs no está acoplado a estas reacciones, la energía producida por una molécula de glucosa es menor que la producida en la respiración aerobia (alrededor de un 5%). La energía restante queda en los enlaces de la molécula de alcohol. Muchos microorganismos como las levaduras y las bacterias realizan numerosos tipos de fermentaciones para producir energía. Por ejemplo, Streptococcus lactis, bacteria responsable de la acidificación de la leche, desasimila glucosa a ácido láctico como producto de fermentación. En ausencia de oxígeno, para actuar como aceptor final, el ácido pirúvico sirve a sí mismo como aceptor.")
48
H +2NAD+ + 2 NADH + 2 H+ COOH Ácido pirúvico CH3 O C 2 Ácido láctico
La formación de ácido pirúvico es común a todos los organismos, pero el tipo de fermentación resulta de la forma como éstos utilizan el ácido pirúvico para formar varios productos. Por lo tanto, todos los microorganismos fermentadores no metabolizan el mismo sustrato exactamente de la misma forma. La respiración anaerobia no se da únicamente en microorganismos, en algunas formas de vida superiores, incluyendo el hombre, también se da. La respiración anaerobia no es suficiente para sostener la vida por un período de tiempo largo, pero es un valioso suplemento a la respiración aerobia. Usted nunca podría subir dos tramos de escalera hasta un tercer piso si dependiera únicamente del suplemento de oxígeno que llegara con suficiente rapidez a sus músculos para permitir la respiración aeróbica únicamente. Debería hacer muchas paradas a lo largo de la escalera para proveerse del oxígeno necesario. En realidad, somos más aptos para realizar movimientos porque las célula microbianas musculares pueden obtener energía a través de la respiración anaerobia. Los jugadores de fútbol no podrían realizar el intenso ejercicio de un partido si no fuera por la respiración anaerobia. COOH Ácido pirúvico CH3 O C + 2 NADH H+ 2 H Ácido láctico CH + 2 NAD+ Cuando el aceptor de electrones es un ácido orgánico se le llama fermentación, cuando el aceptor es una sustancia inorgánica como NO2, NO3, SO4, CO3 y fumarato

49
Diferentes rutas de fermentación
GLUCOSA Ácido pirúvico Ácido acético + Ácido fórmico Ácido succínico Acetona Acetil CoA Ácido fórmico Alcohol etílico CO2 H2

50
Respiración anaerobia

51
En algunas bacterias, al final de la cadena de transporte electrónico, puede existir un aceptor diferente del oxígeno (respiración anaerobia). Los aceptores y sus respectivos productos reducidos (A à AH2) son: NO3-- à N2 SO42- à SH2 fumarato à succinato CO2 à CH4 Fe3+ à Fe2+ Con estos aceptores se obtiene menos energía que con el oxígeno, porque la pareja O2/H2O es más oxidante que las otras.

52
El uso de nitratos, sulfatos y CO2 como aceptores finales de electrones y no como material a incorporar al metabolismo plástico se denomina metabolismo desasimilativo. El producto reducido se excreta al ambiente de la bacteria.

53
Tipos de quimiolitotrofos
La capacidad de obtener energía por fosforilación oxidativa a partir de donadores inorgánicos de electrones sólo se presenta en ciertos grupos de procariotas.

54
bacterias de hidrógeno (H2) bacterias del hierro (Fe2+)
Cada grupo fisiológico de quimiolitotrofos usa un tipo de donador inorgánico: bacterias de hidrógeno (H2) bacterias del hierro (Fe2+) bacterias del azufre (S2-, S0). bacterias nitrificantes, con dos subtipos diferentes: las oxidadoras de amoniaco (llamadas nitrosas) y las oxidadoras del nitrito (llamadas nítricas).
 bacterias del hierro (Fe2+) bacterias del azufre (S2-, S0). bacterias nitrificantes, con dos subtipos diferentes: las oxidadoras de amoniaco (llamadas nitrosas) y las oxidadoras del nitrito (llamadas nítricas).")
55
El uso desasimilativo de nitrato se llama desnitrificación, y ocurre por medio de una serie de fases donde el N va cambiando su estado de oxidación: NO3-- à NO2- (nitrito) à NO (óxido nítrico) à N2O (óx. nitroso) à N2 (dinitrógeno)
 à NO (óxido nítrico) à N2O (óx. nitroso) à N2 (dinitrógeno)")
56
Sólo las bacterias sulfatorredutoras usan el sulfato como aceptor de electrones
Por una ruta especial en la que el sulfato primero tiene que activarse con ATP formando la adenosina-fosfo-sulfato ( APS). La mayoría son quimiorganotrofas, pero algunas quimiolitotrofas pueden usar H2 donador de electrones.
. La mayoría son quimiorganotrofas, pero algunas quimiolitotrofas pueden usar H2 donador de electrones.")
57
Actúan en estas condiciones como quimiolitotrofos:
Las arqueobacterias metanogénicas son los únicos seres vivos capaces de obtener energía acoplando la oxidación del hidrógeno molecular con el uso de CO2 como aceptor de electrones Actúan en estas condiciones como quimiolitotrofos: 4H2 + CO2 à CH4 + 2H2O

58
QUIMIOSÍNTESIS

59
LAS BACTERIAS QUIMIOSINTÉTICAS:
No necesitan nutrientes orgánicos porque utilizan bióxido de carbono para producir sus nutrientes Obtienen la energía de la oxidación de compuestos inorgánicos como hidrógeno molecular, amoníaco, nitrito, tiosulfato, etc. Los electrones resultantes entran en la cadena respiratoria con producción de ATP. QUIMIOSÍNTESIS Algunos bacterias no necesitan nutrientes orgánicos porque, al igual que las plantas, utilizan el bióxido de carbono para producir sus nutrientes, pero a diferencia de los fotosíntéticos obtienen la energía de la oxidación de compuestos inorgánicos como hidrógeno molecular, amoníaco, nitrito, tiosulfato, etc. A este proceso se le llama quimiosíntesis. A los organismos que utilizan este mecanismo de obtención de energía se les llama quimiosintéticos. Los electrones resultantes de estas oxidaciones entran en la cadena respiratoria con producción de ATP. Por ejemplo, para obtener energía, la bacteria Nitrobacter oxida nitritos a nitratos y Thiobacillus obtiene su energía de la oxidación del azufre elemental y otros compuestos de azufre inorgánicos. Las bacterias quimiosintéticas produce menos ATP que aquellas que obtienen su energía de la degradación de compuestos orgánicos en la respiración aerobia.

60
Catabolismo de los lípidos
Glicerina Ácido graso Lípido 3 Ácidos grasos

61
Todos los compuestos de esta reacción entran a la vía glucolítica.
Catabolismo de lípidos La degradación de lípidos o grasas comienza con la hidrólisis de triglicéridos a glicerina y ácidos grasos, por la acción de enzimas denominadas liasas: Glicerina +ATP --> ADP + Glicerina 3P-->NAD+Glicerina 3P-->Fosfato f de hidroacetona + NADH2 Los ácidos grasos se oxidan por la eliminación sucesiva de fragmentos de dos átomos de carbono en forma de acetil CoA. Este acetil CoA formado puede entrar en el ciclo ATC y los átomos de hidrógeno con sus electrones, en la cadena de transporte electrónico, dando lugar a fosforilación oxidativa. Hay mayor rendimiento energético por gramo de grasa que por gramo de carbohidrato o de proteína : 1 gramo de lípido 9.3 Kcal 1 gramo de carbohidrato 3.6 Kcal 1 gramo de proteína 4.0 Kcal Todos los compuestos de esta reacción entran a la vía glucolítica.

62
Oxidación de los ácidos grasos

63
CATABOLISMO DE PROTEÍNAS
Las proteínas son demasiado grandes para atravesar las membranas Los microorganismos excretan proteasas que hidrolizan las proteínas exógenas a péptidos. Proteasas Peptidasas Proteínas----- Péptidos Aminoácidos Catabolismo de proteínas Las proteínas exógenas, como las ingeridas durante la alimentación, pueden ser degradadas utilizando sus productos como fuentes de carbono, nitrógeno y energía. Como las moléculas de las proteínas son demasiado grandes para atravesar las membranas de los microorganismos, los organismos excretan proteasas, que hidrolizan las proteínas exógenas a péptidos. El proceso sería el siguiente: Proteasas Peptidasas Proteínas ----- Péptidos Aminoácidos Cuando se produce la degradación de los aminoácidos, sus esqueletos carbonados se transforman en compuestos que pueden entrar en el ciclo ATC para sufrir una mayor oxidación. Esta entrada en el ciclo ATC puede ser vía acetil CoA, ácido cetoglutárico, ácido succínico, ácido fumárico o ácido oxaloacético Los esqueletos carbonados de los aminoácidos entran en el ciclo ATC para sufrir una mayor oxidación vía acetil CoA, ácido cetoglutárico, ácido succínico, ácido fumárico o ácido oxaloacético

64
RUTAS DE OTROS COMPUESTOS EN LA RESPIRACIÓN AERÓBICA
Otros aminoácidos de más de 3C Acetil CoA 2C Ácido pirúvico Aminoácidos Alcohol Ácidos Grasos Ácido Láctico Glicerol Azúcares complejos Almidón Gliceraldehído 3 P Glucosa 6C Ciclo de los ácidos tricarboxílicos RUTAS DE OTROS COMPUESTOS EN LA RESPIRACIÓN AERÓBICA Rutas del catabolismo o degradación de compuestos en la respiración aeróbica La glucosa es la fuente de energía más importante para la mayor parte de las célula microbianas. Sin embargo, otras sustancias como lípidos y proteínas, pueden utilizarse como fuentes alternativas (Figura 48). Todas estas reacciones son mediadas por enzimas. Existe una regla general que rige su utilización: se han de convertir de la forma más rápida y eficiente en intermediarios de las rutas glicolítica y del ciclo ATC, de manera para llevar a cabo su degradación completa, el número adicional de enzimas sea el mínimo posible. Esta es la razón por la que las rutas glicolítica y el ciclo ATC constituyen el esqueleto alrededor del cual se componen el resto de las vías degradativas
. Todas estas reacciones son mediadas por enzimas. Existe una regla general que rige su utilización: se han de convertir de la forma más rápida y eficiente en intermediarios de las rutas glicolítica y del ciclo ATC, de manera para llevar a cabo su degradación completa, el número adicional de enzimas sea el mínimo posible. Esta es la razón por la que las rutas glicolítica y el ciclo ATC constituyen el esqueleto alrededor del cual se componen el resto de las vías degradativas.")
65
BIOSÍNTESIS (Anabolismo)
El proceso más complejo y que mas energía requiere es la biosíntesis de constituyentes de los microorganismos, o macromóleculas que hacen parte de la estructura celular. En la célula microbiana, continuamente se utilizan las moléculas sencillas, producto de la degradación de sustancias complejas, para resintetizar otros compuestos complejos que le permita formar nuevas estructuras, reproducirse, formar enzimas, etc. Como la formación de nuevas moléculas implica la formación de enlaces químicos, la célula microbiana necesita utilizar energía.

66
Unidades estructurales
Intermediarios de bajo peso molecular Unidades estructurales Macromoléculas (Alto peso molecular) Asociaciones supra - moleculares Organelas Acetato, Malonato Ácidos grasos, Glicerina Lípidos Fosfopiruvato, Malato Azúcares sencillos Poli - sacáridos Cetoácidos Aminoácidos Proteínas Ribosa carbamil fosfato Mono - nucleóticos Ácidos Nucléicos Complejos enzimáticos, Ribosomas, Sistemas contráctiles Núcleo, mitocondria, cloroplasto, etc.
 Asociaciones. supra - moleculares. Organelas. Acetato, Malonato. Ácidos grasos, Glicerina. Lípidos. Fosfopiruvato, Malato. Azúcares sencillos. Poli - sacáridos. Cetoácidos. Aminoácidos. Proteínas. Ribosa. carbamil fosfato. Mono - nucleóticos. Ácidos Nucléicos. Complejos enzimáticos, Ribosomas, Sistemas contráctiles. Núcleo, mitocondria, cloroplasto, etc.")
67
RELACIÓN ENTRE LAS TRANSFORMACIONES CATABÓLICAS Y ANABÓLICAS

68
FASE I FASE II FASE III CO2 Lípidos Polisacáridos Proteínas
Aminoácidos Hexosas Pentosas Acidos grasos Glicerina Gliceraldehido 3-fosfato Fosfoenolpirúvato Á.Pirúvico CO2 Succinil CoA Acido Fumárico Acido Málico Acido Cítrico Acido - cetoglutárico Acido Isocítrico Acido Succínico A. Oxaloacético Acetil CoA FASE III FASE II FASE I Ciclo de los ácidos tricarboxílicos

Presentaciones similares















: Autótrofos: A partir de la materia inorgánica y del CO2 de la atmósfera.>")




