Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Metabolismo intermediario
M.C. Karla Díaz

2
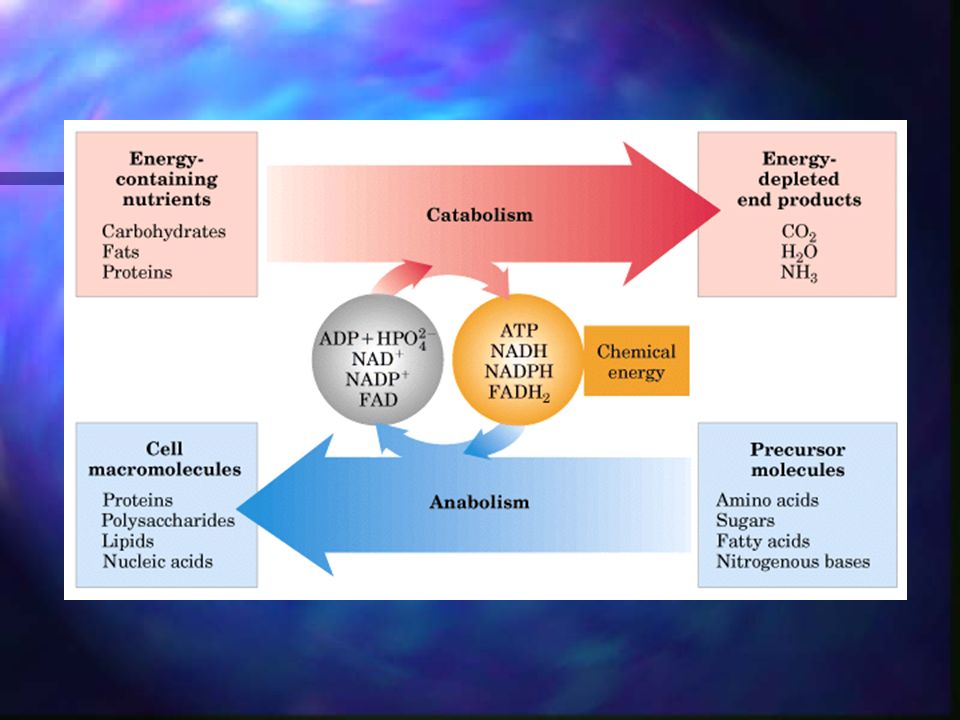
Metabolismo El metabolismo es la suma de todas las transformaciones químicas que se producen en una célula u organismo. Tiene lugar en una serie de reacciones catalizadas enzimáticamente, que constituyen las rutas metabólicas.

3
Metabolismo intermediario
Actividades combinadas de todas las rutas metabólicas que interconvierten precursores, metabolitos, y productos de baja masa molecular.

4
Metabolismo Catabolismo: fase degradadora del metabolismo en la que moléculas nutrientes orgánicas (glúcidos, grasas y proteínas) se convierten en productos más sencillos. Anabolismo: fase en la que precursores sencillos se integran en moléculas mucho más grandes y complejas como los lípidos, polisacáridos, proteínas y ácidos nucléicos.
 se convierten en productos más sencillos. Anabolismo: fase en la que precursores sencillos se integran en moléculas mucho más grandes y complejas como los lípidos, polisacáridos, proteínas y ácidos nucléicos.")
6
Reacciones biológicas de reducción-oxidación
La transferencia de grupos fosfato es una de las características principales del metabolismo. Las reacciones de transferencia electrónica metabólicas son también de importancia crucial.

7
Reacciones biológicas de reducción-oxidación
En estas reacciones interviene la pérdida de electrones por una especie química, que es así oxidada, y la ganancia por otra que es reducida. El flujo de electrones es responsable de todo el trabajo realizado por los organismos vivos.

8
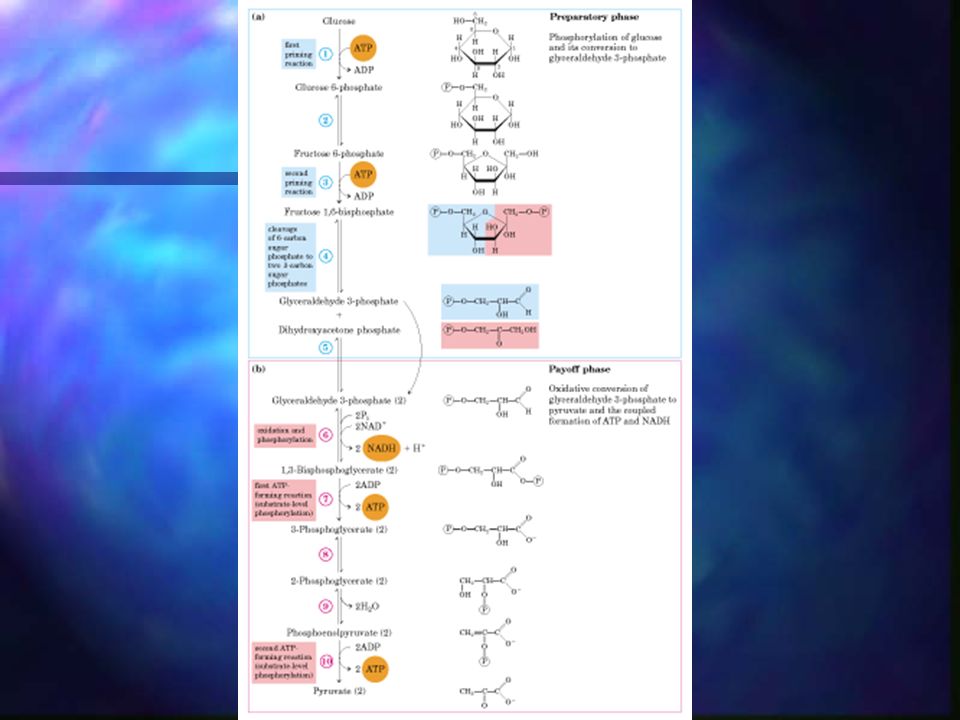
Glucólisis La glucolisis tiene lugar en el citoplasma celular.
Consiste en una serie de diez reacciones, cada una catalizada por una enzima determinada, que permite transformar una molécula de glucosa en dos moléculas de un compuesto de tres carbonos, el ácido pirúvico.

11
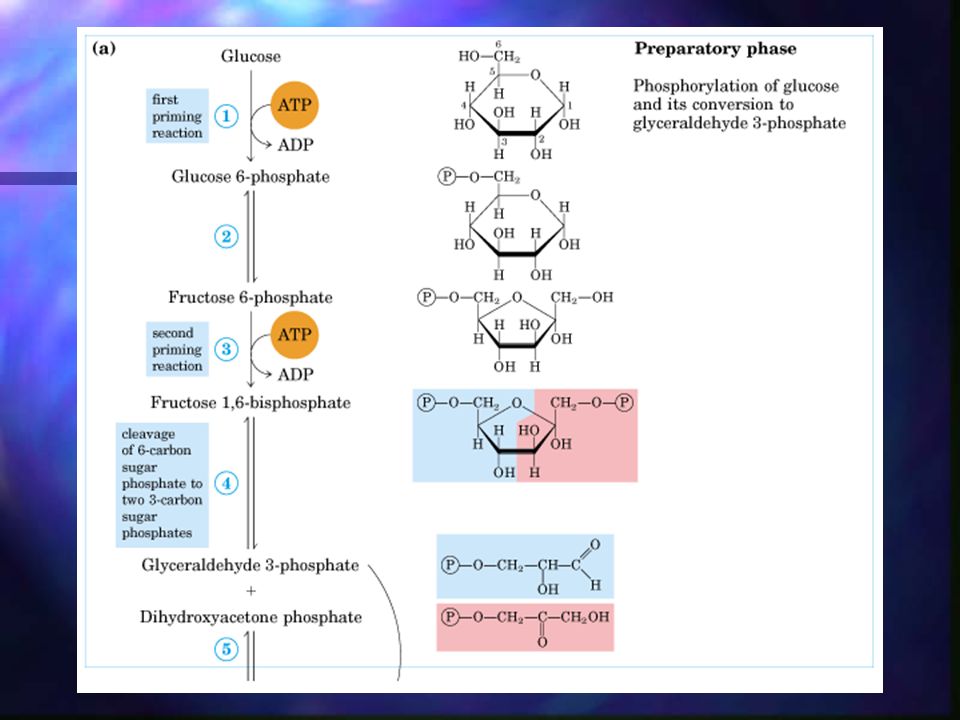
Glucólisis La rotura de la glucosa (6 C) en dos moléculas de piruvato tiene lugar en 10 pasos, de los cuales los primeros 5 son la fase preparatoria. La fase preparatoria consiste en la fosforilación y conversión de la glucosa en gliceraldehído-3-fosfato.

12
Glucólisis – Fase preparatoria
La glucosa es fosforilada en el grupo OH del C6. La D-glucosa-6-fosfato formada se convierte en D-fructosa-6-fosfato. La D-fructosa-6-fosfato es fosforilada en C1, dando D-fructosa-1,6-bifosfato. La D-fructosa-1,6-bifosfato se parte en dos moléculas de 3 C: dihidroxiacetona fosfato y gliceraldehído-3-fosfato (lisis). La dihidroxiacetona se isomeriza a una segunda molécula de gliceraldehído-3-fosfato. *El ATP es el dador de fosfato en ambas fosforilaciones.
. La dihidroxiacetona se isomeriza a una segunda molécula de gliceraldehído-3-fosfato. *El ATP es el dador de fosfato en ambas fosforilaciones.")
14
Glucólisis – Fase de beneficios
6. Cada molécula de gliceraldehído-3-fosfato es oxidada y fosforilada por fosfato inorgánico (no por ATP) formando 1,3-bifosfoglicerato. 7 al 10. Conversión de 2 moléculas de 1,3-bifosfoglicerato en 2 moléculas de piruvato.
 formando 1,3-bifosfoglicerato. 7 al 10. Conversión de 2 moléculas de 1,3-bifosfoglicerato en 2 moléculas de piruvato.")
16
Glucólisis – Balance global
Al utilizar 2 ATP en la fase preparatoria y ganar 4 ATP en la fase de beneficios, se obtiene una ganancia de 2 ATP.

17
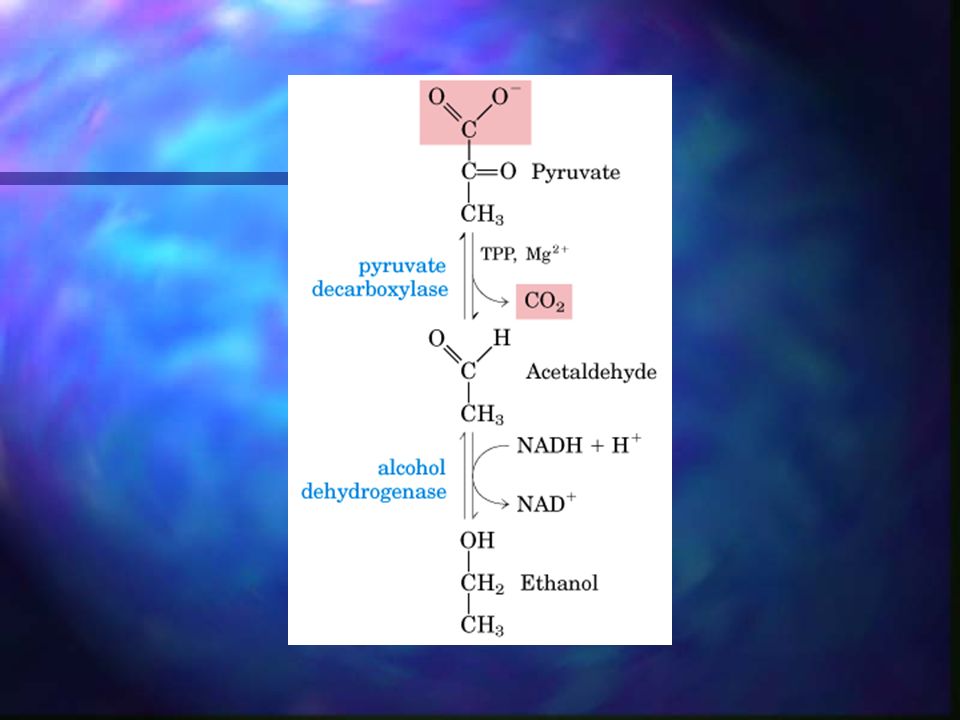
Piruvato El piruvato formado en la glucólisis puede tomar 3 rutas metabólicas alternativas. En organismos o tejidos aeróbicos, el piruvato se oxida, con pérdida de su grupo carboxilo como CO2, dando el grupo acetilo del acetil-CoA. Otra ruta del piruvato es la fermentación del ácido láctico (músculo, anaeróbicamente). La tercera ruta es la fermentación etanólica o alcohólica, que se da en microorganismos.
. La tercera ruta es la fermentación etanólica o alcohólica, que se da en microorganismos.")
21
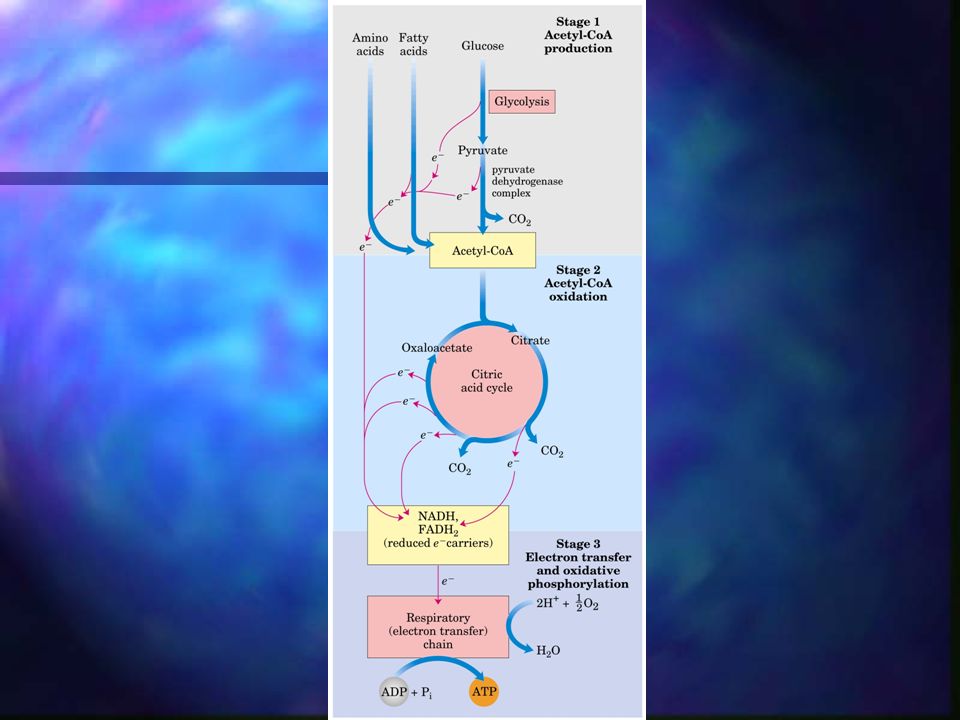
Producción de acetato En organismos aeróbicos, la glucosa y otros azúcares, los ácidos grasos y la mayor parte de los AA son oxidados finalmente a CO2 y H2O a través del ciclo del ácido cítrico. Antes de entrar a este ciclo, los esqueletos carbonados de azúcares y ác. grasos deben sufrir un proceso de degradación para dar lugar al grupo acetilo del acetil-CoA.

23
Producción de acetato El piruvato procedente de la glucosa (glucólisis) se oxida para dar lugar a Acetil-CoA y CO2 a consecuencia de la acción de una agrupación de 3 enzimas: el complejo piruvato deshidrogenasa.
 se oxida para dar lugar a Acetil-CoA y CO2 a consecuencia de la acción de una agrupación de 3 enzimas: el complejo piruvato deshidrogenasa.")
25
Descarboxilación oxidativa
La reacción llevada a cabo por el complejo piruvato deshidrogenasa es la descarboxilación oxidativa. La descarboxilación oxidativa es un proceso de oxidación irreversible en el que el piruvato pierde un grupo carboxilo en forma de CO2, y los dos carbonos restantes se transforman en el grupo acetilo del acetil-CoA.

26
Descarboxilación oxidativa
La deshidrogenación y descarboxilación combinadas del piruvato hasta acetil-CoA son resultado de la acción de 3 enzimas y 5 coenzimas. Enzimas: piruvato deshidrogenasa (E1), dihidrolipoil transacetilasa (E2) y dihidrolipoil deshidrogenasa (E3). Coenzimas: Pirofosfato de tiamina (TPP), dinucleótido de flavina y adenina (FAD), coenzima a (CoA), dinucleótido de nicotinamida y adenina (NAD) y el lipoato.
, dihidrolipoil transacetilasa (E2) y dihidrolipoil deshidrogenasa (E3). Coenzimas: Pirofosfato de tiamina (TPP), dinucleótido de flavina y adenina (FAD), coenzima a (CoA), dinucleótido de nicotinamida y adenina (NAD) y el lipoato.")
27
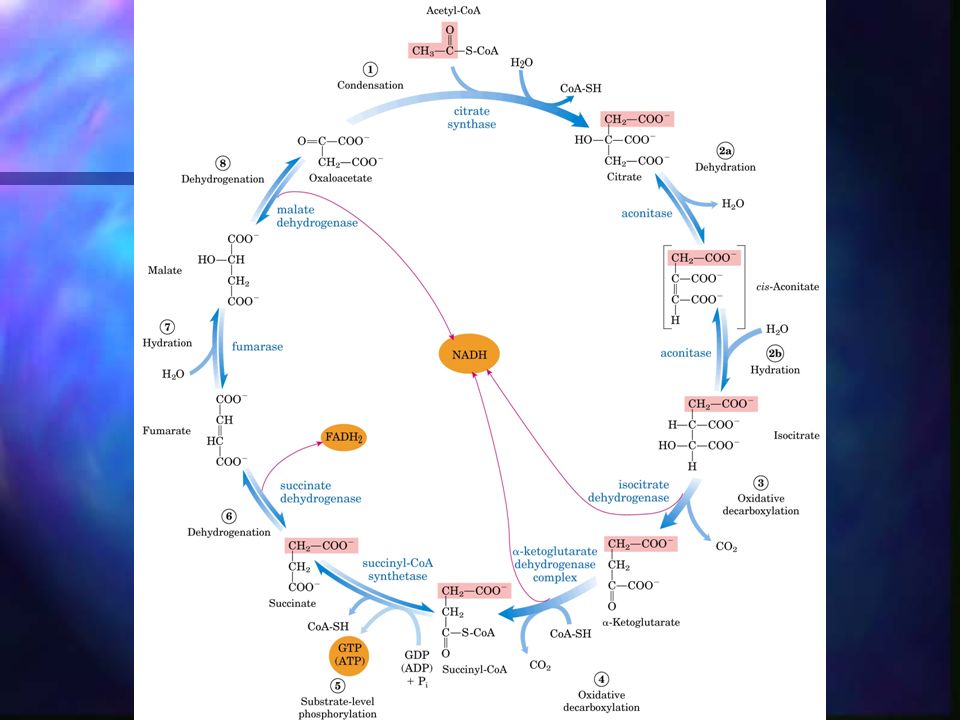
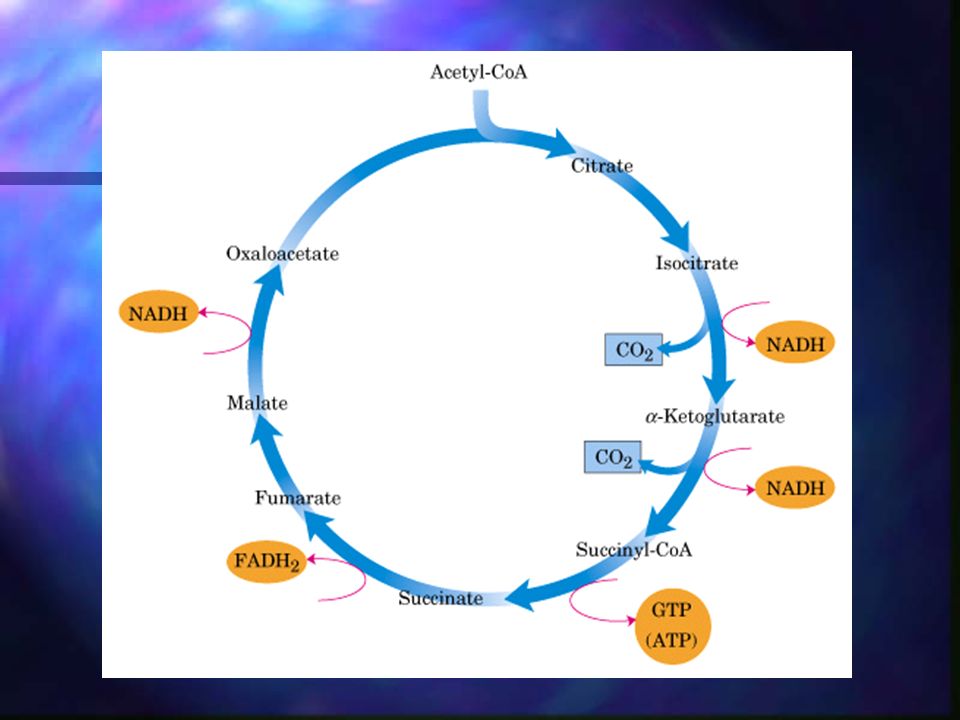
Ciclo de Krebs Ciclo de los ácidos tricarboxílicos, se realiza en la mitocondria. A diferencia de la glicólisis, en el ciclo de Krebs las reacciones son cíclicas y consta de 8 pasos. En cada vuelta del ciclo entra un grupo acetilo (2 C) en forma de Acetil-CoA y la salida de dos moléculas de CO2. Cuatro de los pasos de este proceso son oxidaciones en las que la energía de oxidación se conserva, con gran eficiencia, mediante la formación de cofactores reducidos (NADH y FADH2).
 en forma de Acetil-CoA y la salida de dos moléculas de CO2. Cuatro de los pasos de este proceso son oxidaciones en las que la energía de oxidación se conserva, con gran eficiencia, mediante la formación de cofactores reducidos (NADH y FADH2).")
29
Ciclo de Krebs – Paso 1 Formación de citrato: el acetil-CoA se condensa con oxalacetato para formar el citrato. Catalizada por la citrato sintasa. El carbono metílico del acetilo se une al grupo carbonílico del oxalacetato.

30
Ciclo de Krebs – Paso 2 2. Formación de isocitrato vía cis-aconitato: La enzima aconitasa (aconitato hidratasa) cataliza la conversión de citrato en isocitrato, a través de la formación intermedia del ácido tricarboxílico cis-aconitato.
 cataliza la conversión de citrato en isocitrato, a través de la formación intermedia del ácido tricarboxílico cis-aconitato.")
31
Aconitasa La aconitasa tiene un centro de hierro-azufre que actúa en la fijación del sustrato en el centro activo como en la catálisis de adición o eliminación de H2O.

32
Ciclo de Krebs – Paso 3 3. Oxidación del isocitrato a -cetoglutarato y CO2: La isocitrato deshidrogenasa cataliza la descarboxilación oxidativa del isocitrato dando lugar a la formación de -cetoglutarato.

33
Ciclo de Krebs – Paso 4 4. Oxidación del -cetoglutarato a succinil-CoA y CO2: Descarboxilación oxidativa que ocurre por acción del complejo -cetoglutarato deshidrogenasa. El NAD+ actúa como aceptor de electrones.

34
Ciclo de Krebs – Paso 5 5. Conversión de succinil CoA en succinato: El succinil-CoA tiene una energía libre negativa de la hidrólisis de su enlace tioéster, que se utiliza para sintetizar de ATP o GTP y dar lugar al succinato. Catalizada por succinil-CoA sintetasa o succínico tioquinasa.

36
Ciclo de Krebs – Paso 6 6. Oxidación del succinato a fumarato: Esta reacción sucede por acción de la flavoproteína succinato deshidrogenasa.

37
El malonato, un análogo del succinato, es un fuerte inhibidor competitivo de la succinato deshidrogenasa y puede bloquear el ciclo del ácido cítrico.

38
Ciclo de Krebs – Paso 7 7. Hidratación del fumarato y producción de L-malato: Catalizada por la fumarasa (fumarato hidratasa).
.")
39
Ciclo de Krebs – Paso 8 8. Oxidación del L-malato a oxalacetato: Última reacción del ciclo, catalizada por la L-malato deshidrogenasa, dependiente de NAD.

40
Ciclo de Krebs - Rendimiento
Cada vuelta del ciclo produce: 3 NADH 1 FADH2 1 GTP o ATP. Se liberan 2 CO2 mediante descarboxilación oxidativa.

42
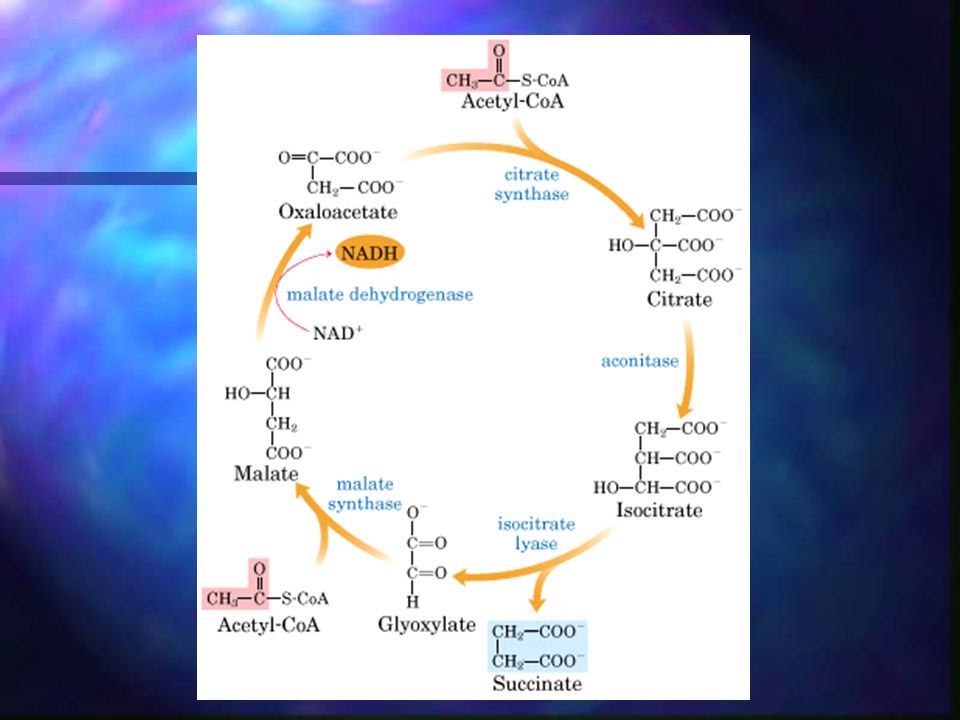
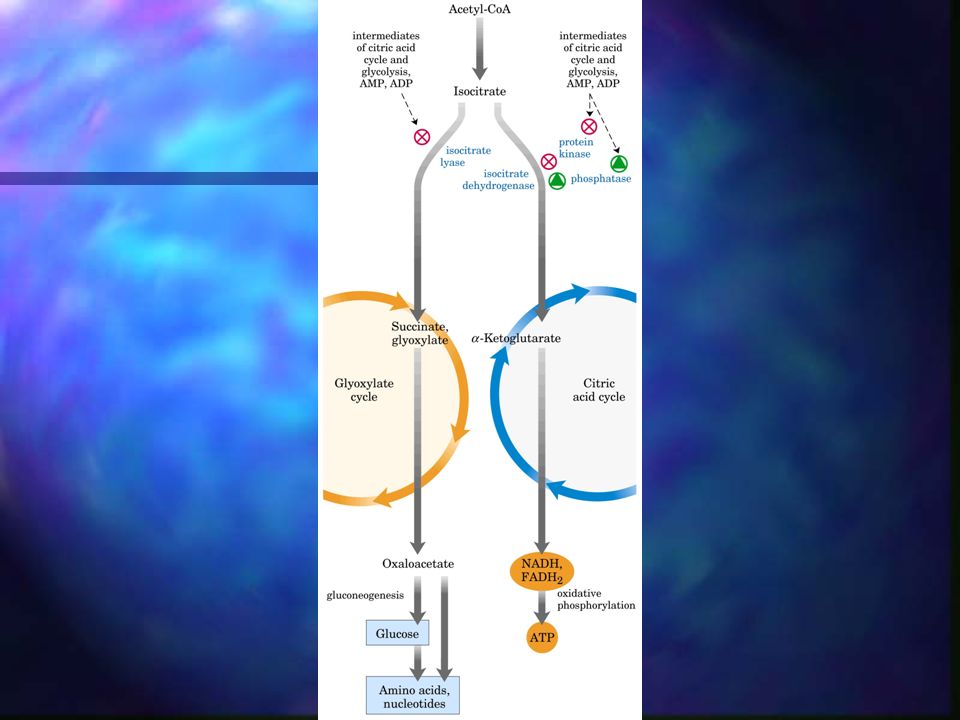
Ciclo del glioxilato Variación del ciclo de Krebs.
El acetil-CoA se condensa con el oxalacetato para formar citrato exactamente igual que en el ciclo de Krebs. La degradación del isocitrato no se produce a través de la isocitrato deshidrogenasa sino mediante una rotura catalizada por la isocitrato liasa, dando lugar a succinato y glioxilato. El glioxilato se condensa con el acetil-CoA para dar lugar a malato, catalizada por la malato sintasa. El malato se oxida a oxalacetato, para iniciar nuevamente el ciclo.

46
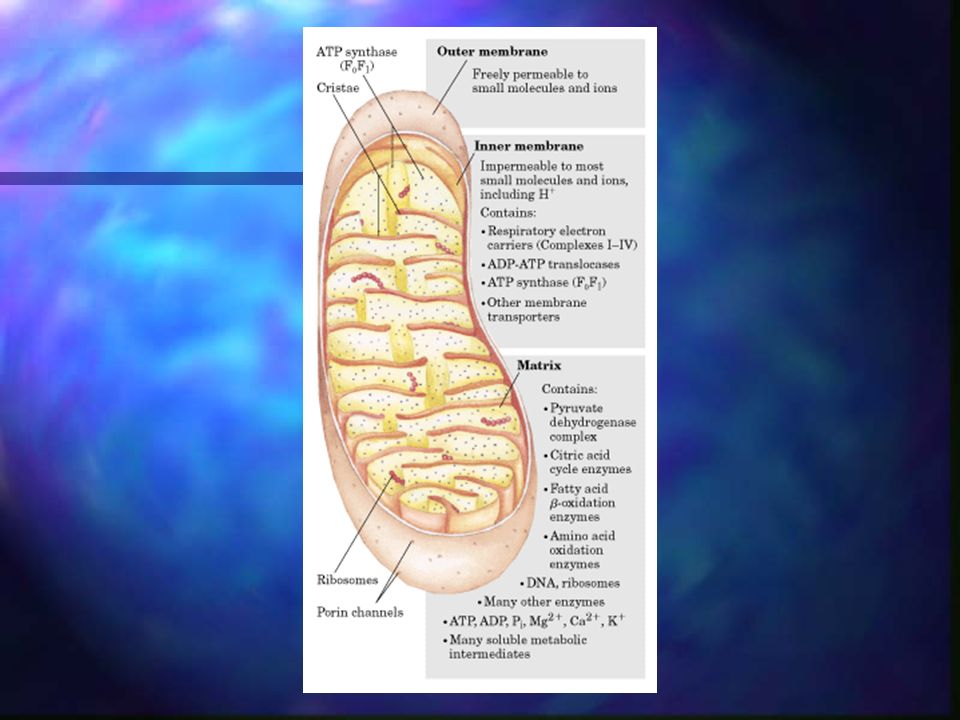
Fosforilación oxidativa
La fosforilación oxidativa o cadena de transporte de electrones es la transferencia de electrones de los equivalentes reducidos NADH, NADPH, FADH, obtenidos en la glucólisis y en el ciclo de Krebs hasta el oxígeno molecular, acoplado con la síntesis de ATP. Este proceso metabólico está formado por un conjunto de enzimas complejas que catalizan varias reacciones de óxido-reducción, donde el oxígeno es el aceptor final de electrones y donde se forma finalmente agua. Es donde se sintetiza la mayor cantidad de ATP, y se lleva a cabo en las membranas biológicas (membrana mitocondrial).
.")
48
Fosforilación oxidativa
La mayor parte de los electrones que entran a la cadena respiratoria mitocondrial provienen de la acción de deshidrogenasas que captan electrones de las reacciones oxidativas, canalizándolos en forma de pares electrónicos. Estas deshidrogenasas utilizan NAD o NADP, FMN o FAD como aceptores electrónicos.

49
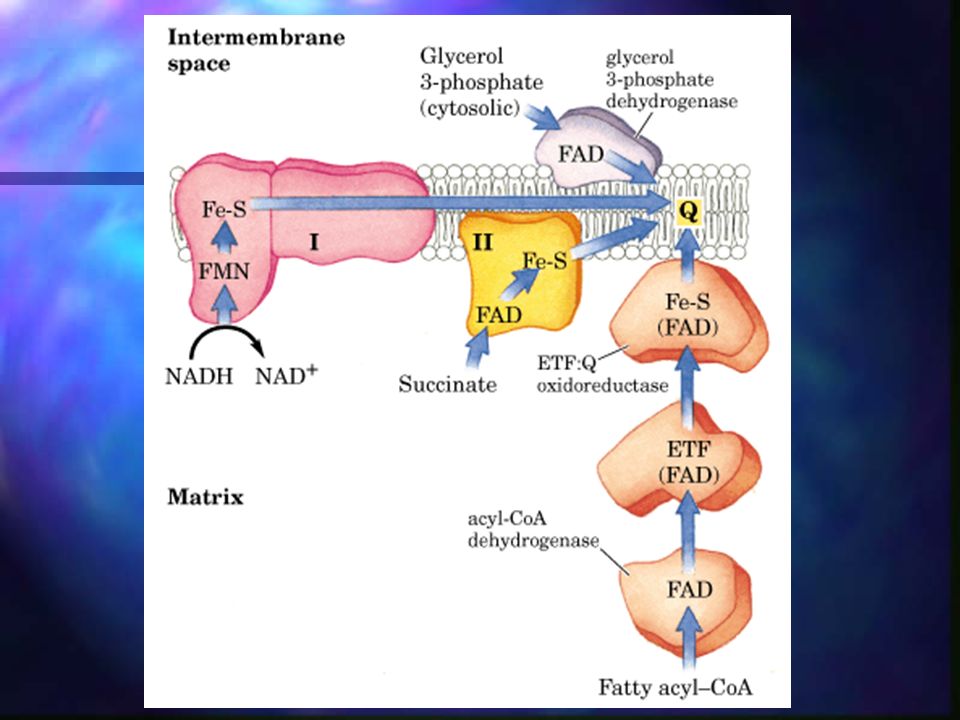
Fosforilación oxidativa
La cadena respiratoria mitocondrial posee una serie de transportadores electrónicos, la mayoría proteínas integrales de membrana con grupos prostéticos capaces de aceptar y donar uno o dos electrones. Cada componente de la cadena puede aceptar electrones del transportador precedente y transferirlos al siguiente en una secuencia específica.

52
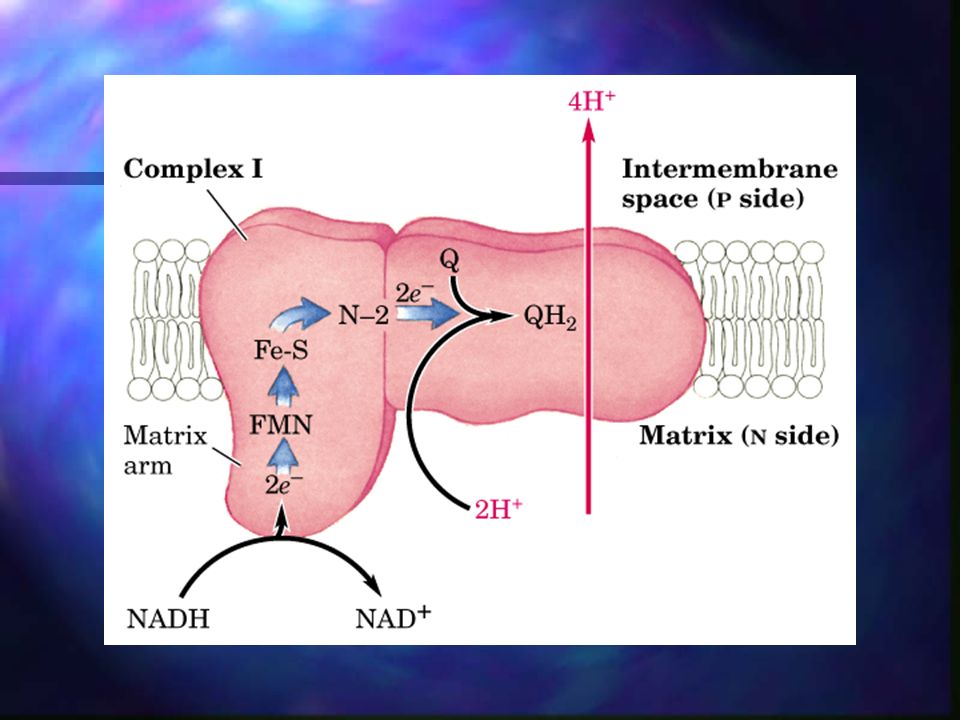
Fosforilación oxidativa – Complejo I
NADH a Ubiquinona. Complejo de flavoproteínas incrustado en la membrana mitocondrial interna. La reacción global catalizada es: NADH + H+ + UQ NAD+ + UQH2 La ubiquinona oxidada acepta un ion hidruro (2 electrones y 1 protón) desde el NADH y un protón desde el agua disolvente de la matriz.
 desde el NADH y un protón desde el agua disolvente de la matriz.")
54
Fosforilación oxidativa – Complejo II
Succinato a ubiquinona. Succinato deshidrogenasa, única enzima del ciclo de Krebs ligada a la membrana. Los electrones pasan desde el succinato al FAD y a continuación a la ubiquinona a través de los centros Fe-S.

56
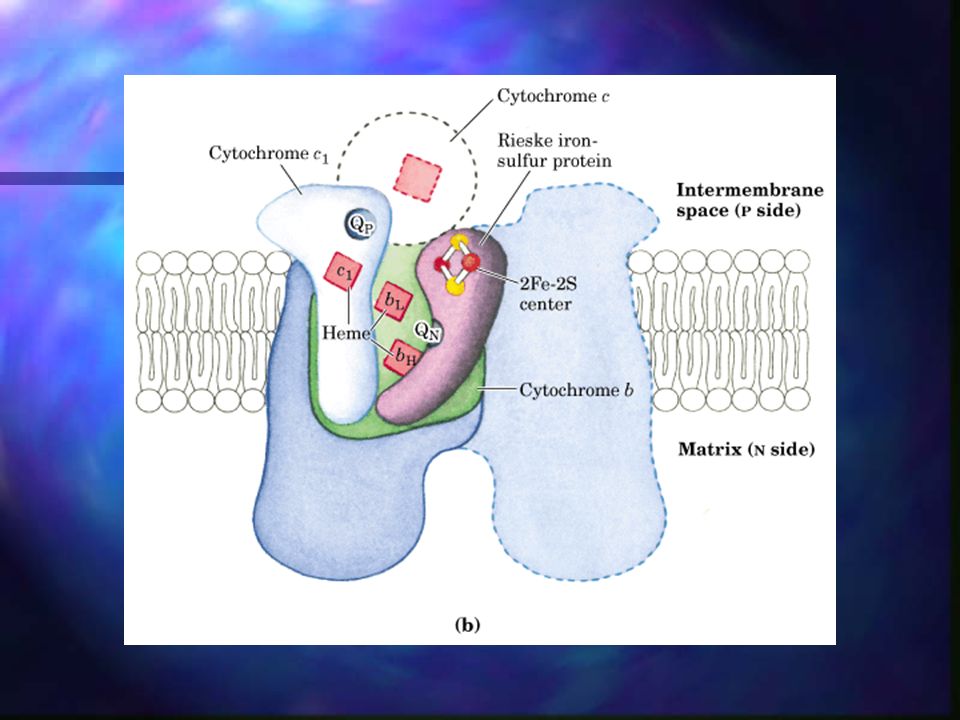
Fosforilación Oxidativa – Complejo III
Ubiquinona a citocromo C. Contiene los citocromos b562 y b566 citocromo c1, una proteína ferrosulfurada y al menos otras seis subunidades protéicas. Funciona como una bomba de protones. La ruta del flujo de electrones en este complejo es complicada, pero el efecto neto de la transferencia es sencillo: UQH2 se oxida a UQ y se reduce el citocromo c.

58
Fosforilación oxidativa – Complejo IV
Reducción del O2. Complejo citocromo oxidasa, contiene citocromos a y a3, formados por dos grupos hemo, también contiene dos iones cobre de importancia crucial para la transferencia de electrones al O2. Funciona como bomba de protones.

60
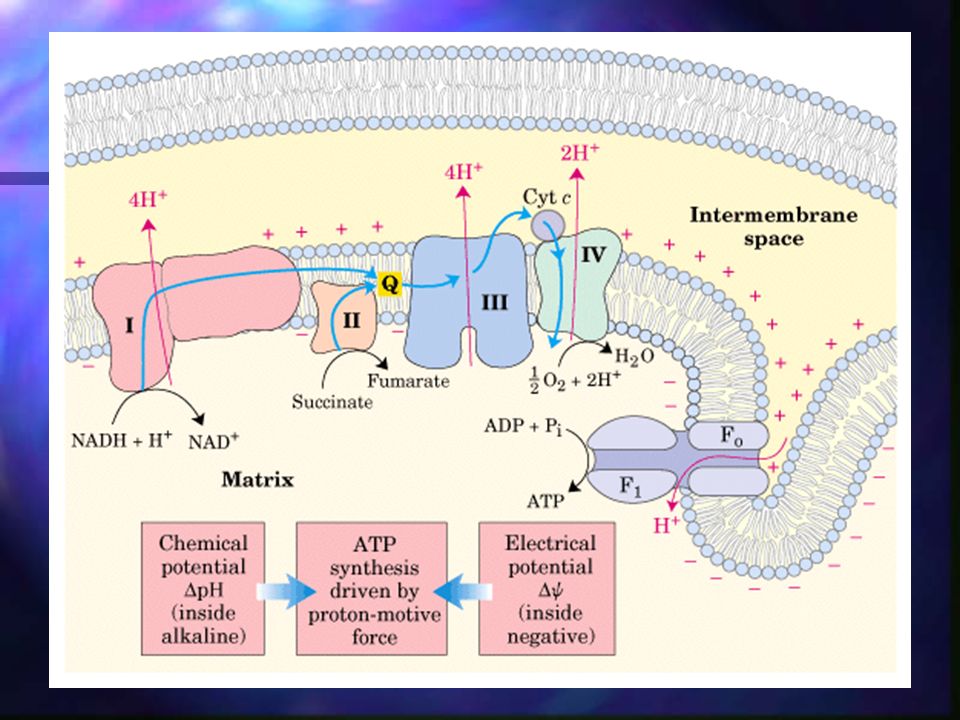
Fosforilación oxidativa - Resumen
Los electrones llegan a la UQ vía complejos I y II. UQH2 actúa de transportador móvil de electrones y protones, y pasa electrones al complejo III, el cual los pasa al citocromo c (móvil). El complejo IV transfiere electrones desde el citocromo c reducido al O2. El flujo electrónico a través de los complejos I, III y IV va acompañado de flujo de protones desde la matriz al espacio intermembrana.
. El complejo IV transfiere electrones desde el citocromo c reducido al O2. El flujo electrónico a través de los complejos I, III y IV va acompañado de flujo de protones desde la matriz al espacio intermembrana.")
62
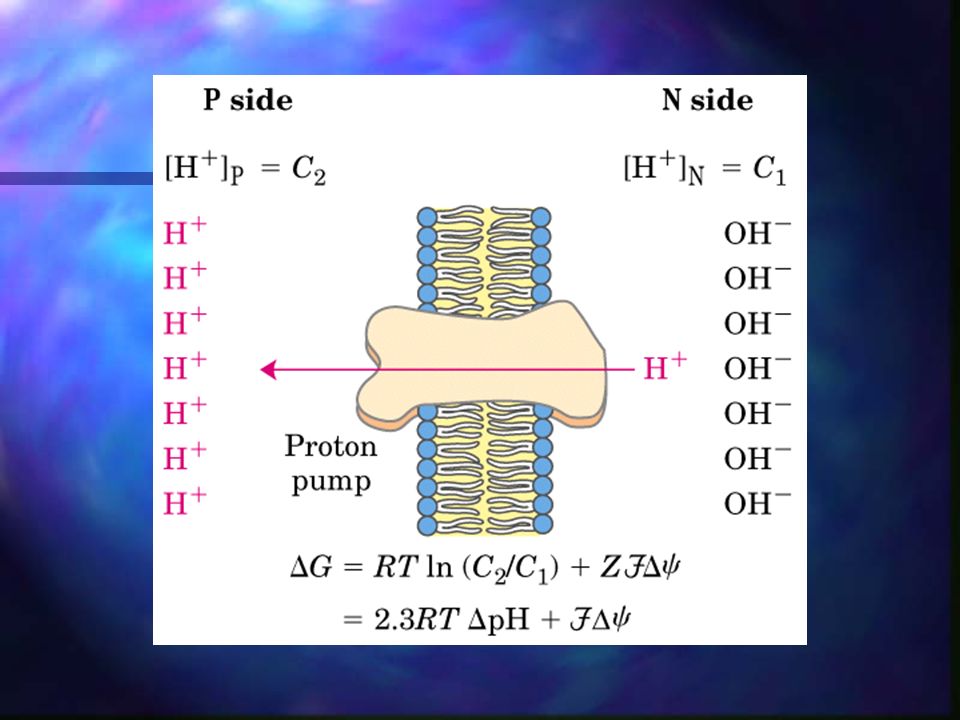
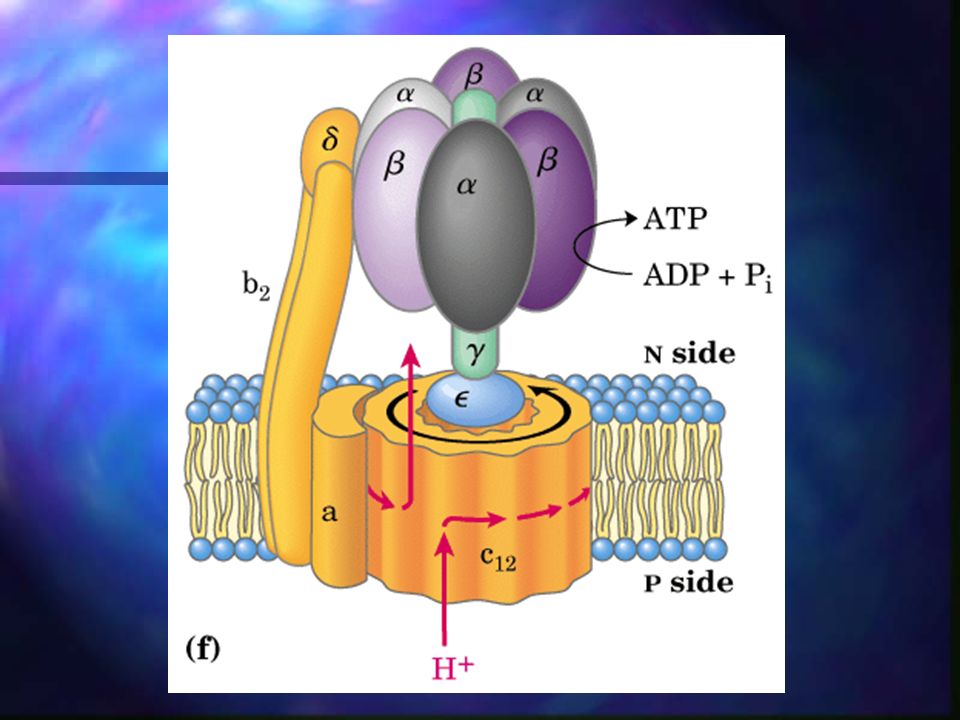
Fosforilación oxidativa – Síntesis de ATP
Debido a que los protones se han bombeado al espacio intermembranoso de la mitocondria en contra de gradiente, ahora pueden fluir nuevamente dentro de la matriz mitocondrial y mediante la vía ATP-sintetasa, se genera ATP en el proceso.

66
RENDIMIENTO TOTAL DE ATP POR MOLÉCULA DE GLUCOSA
PROCESO PRODUCTO DIRECTO ATP FINAL Glucólisis 2 NADH (Citosólico) 4 o 6 2 ATP 2 Oxidación del piruvato (2 por glucosa) 2 NADH (Matriz mitocondrial) 6 Oxidación del acetil-CoA (matriz mitocondrial) 6 NADH (Matriz mitocondrial) 18 2 FADH2 4 2 ATP o 2 GTP RENDIMIENTO TOTAL 36 o 38
 4 o 6. 2 ATP. 2. Oxidación del piruvato. (2 por glucosa) 2 NADH (Matriz mitocondrial) 6. Oxidación del acetil-CoA. (matriz mitocondrial) 6 NADH (Matriz mitocondrial) FADH ATP o 2 GTP. RENDIMIENTO TOTAL. 36 o 38.")
Presentaciones similares