Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Biología Molecular

2
Estructura La membrana biológica plasmática es una estructura semipermeable que actúa de barrera separadora entre los medios intra y extracelular, la cual permite el mantenimiento de unas condiciones físico- químicas necesarias para el conjunto de reacciones metabólicas que se dan en el interior de los orgánulos celulares. Responde al denominado modelo del mosaico fluido.

3
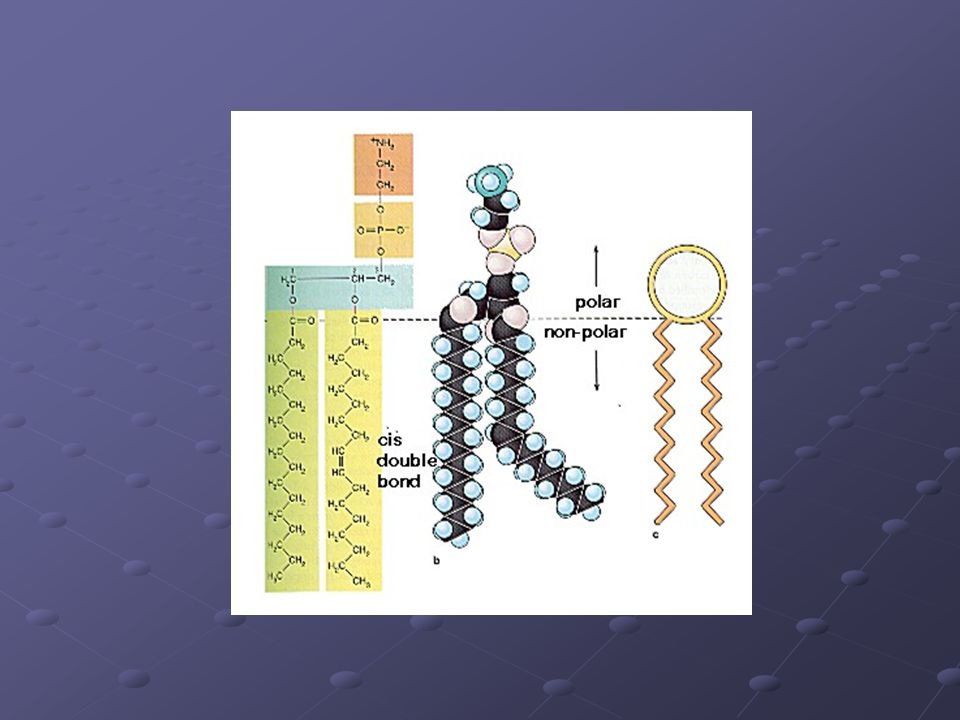
Bicapa lipídica Componente estructural básico de la membrana plasmática. Se basa en una doble capa de moléculas lipídicas: Fosfolípidos y colesterol, fundamentalmente

4
Fosfolípidos Moléculas anfipáticas, esto es, poseen una zona hidrófila polar y una zona hidrofóbica apolar. A consecuencia de esto, las cabezas hidrófilas interaccionan entre sí y con las moléculas de H20, mientras que las cadenas hidrófobas son repelidas por la fase acuosa y se empaquetan hacia el interior de una bicapa. Dicha bicapa lipídica otorga a la membrana celular determinadas propiedades.

6
Propiedades de la membrana
Autoensamblaje espontáneo de las moléculas lipídicas Autosellado de la bicapa

7
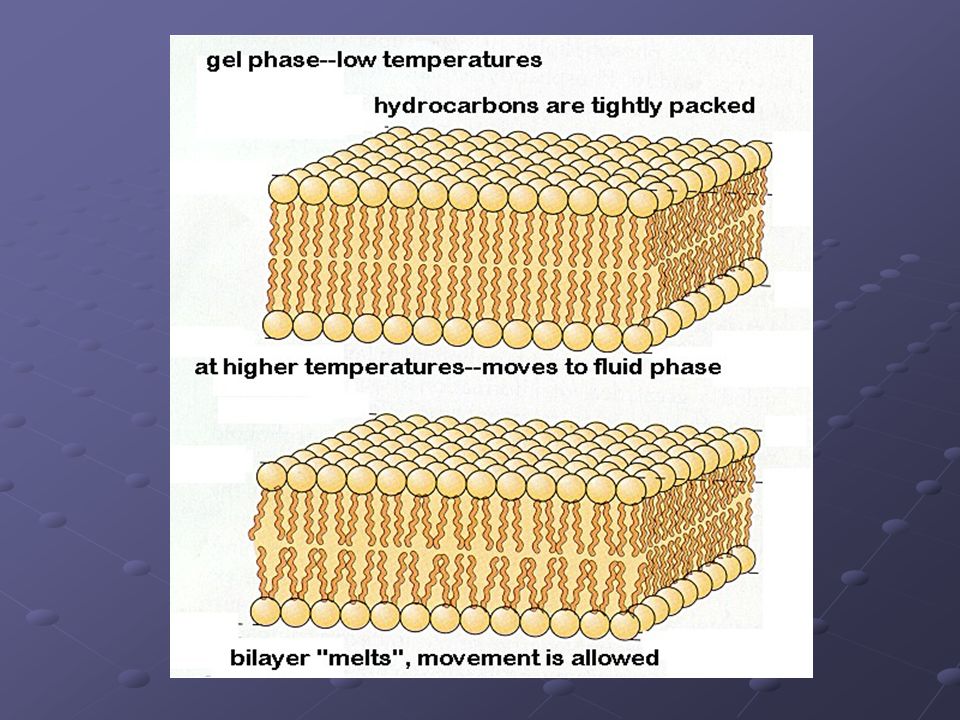
Fluidez. - Causa: Se debe a que las moléculas lipídicas tienen la capacidad de movimientos laterales de difusión dentro de cada monocapa; a pesar de ello, la composición lipídica se mantiene constante (asimetría lipídica) no se dan intercambio de fosfolípidos entre monocapas. - Importancia biológica: La fluidez determina el funcionamiento de la membrana. Los cambios de temperatura en el medio influyen en ella: A menor temperatura, menor fluidez (mayor viscosidad). El descenso de fluidez de la membrana puede detener procesos de transporte y enzimáticos.
 no se dan intercambio de fosfolípidos entre monocapas. - Importancia biológica: La fluidez determina el funcionamiento de la membrana. Los cambios de temperatura en el medio influyen en ella: A menor temperatura, menor fluidez (mayor viscosidad). El descenso de fluidez de la membrana puede detener procesos de transporte y enzimáticos.")
9
Impermeabilidad frente a iones y sustancias hidrosolubles
- Causa: Naturaleza anfipática de la bicapa - Importancia biológica: Regulación del intercambio de sustancias entre el citosol y el medio extracelular

10
Colesterol Las moléculas de colesterol se encuentran intercaladas entre los fosfolípidos, y su función principal es la de regular la fluidez de la bicapa inmovilizando las colas hidrofóbicas próximas a la regiones polares

11
Proteínas de membrana Determinan la función de la membrana biológica. Poseen la propiedad de desplazarse lateralmente a través de ella pero no de invertir su posición (asimetría proteica)
")
12
Clasificación En función de la asociación con lípidos de la membrana se clasifican en: Proteínas integrales Unidas fuertemente a los lípidos de membrana.Se subdividen en: Proteínas de transmembrana. Se hallan inmersas en la bicapa Proteínas periféricas. Exteriores a la bicapa, están unidas mediantes en laces débiles a las cabezas polares lipídicas o a otras proteínas de integrales

13
Glucocálix Superficie externa de la membrana formada por la zona glucídica de glucolípidos y glucoproteínas.

14
Funciones Protección contra daños químicos y/o físicos.
Interviene en fenómenos de reconocimiento celular. Confiere viscosidad a la membrana. Interviene en la comunicación intracelular.

15
Dominios de membrana Regiones de la membrana plasmática que poseen una especialización funcional concreta. Surgen por restricción de movimientos de los componentes de la misma.

16
FUNCIONES DE LA MEMBRANA

17
Funciones de la membrana plasmática
La membrana plasmática controla la entrada y salida de materiales (permeabilidad selectiva) , participa en las interacciones célula - célula y célula – matriz y es un elemento fundamental en la comunicación celular, recibiendo señales externas y transmitiendo dicha informacion al interior celular.
 , participa en las interacciones célula - célula y célula – matriz y es un elemento fundamental en la comunicación celular, recibiendo señales externas y transmitiendo dicha informacion al interior celular.")
18
Permeabilidad selectiva
La impermeabilidad de la membrana, gracias a su naturaleza lipídica, no es absoluta, sino que permite el intercambio de materia y energía con el ambiente externo. Por esto se han desarrollado sistemas de transporte específicos, en los que las proteínas regulan el paso de sustancias hidrófilas, ionizadas o de gran tamaño a través de estas membranas. Este transporte puede ser activo o pasivo. Transporte pasivo No se requiere energía para que la sustancia cruce la membrana plasmática. Las moléculas se mueven desde el lado con mas concentración hasta el lado menos concentrado. Existen dos tipos de transporte pasivo principalmente: difusión simple y difusión facilitada.

19
Mecanismos de transporte pasivo
Difusión simple: Atraviesan la membrana las moléculas no polares (liposolubles) como los gases y algunas hormonas esteroideas y tiroideas. También pueden pasar pequeñas moléculas polares como el agua o el etanol. Difusión facilitada: se realiza mediante proteínas transportadoras y proteínas canal. De esta forma las moléculas polares mas grandes pueden atravesar la membrana.
 como los gases y algunas hormonas esteroideas y tiroideas. También pueden pasar pequeñas moléculas polares como el agua o el etanol. Difusión facilitada: se realiza mediante proteínas transportadoras y proteínas canal. De esta forma las moléculas polares mas grandes pueden atravesar la membrana.")
20
Proteínas transportadoras
Son proteínas de transmembrana que se unen específicamente a la molécula que trasportan. Esta unión provoca un cambio en la configuración de la proteína, que hace que la molécula quede libre una vez transportada. La proteína transportadora recupera su forma inicial para seguir actuando en otros procesos. Proteínas canal Son proteínas de transmembrana que forman en su interior un canal acuoso, que permite el paso de iones. Estos canales se abren según un tipo de señal especifico. Dependiendo del tipo de señal encontramos: - Canales iónicos dependientes del ligando: El ligando se une a un receptor en la zona externa de la proteína canal de forma especifica, provocando cambios en su conformación que permiten la apertura del canal, y por tanto la difusión de iones. Proteína canal - Canales iónicos dependientes del voltaje: Se abren en respuesta a los cambios de potencial de membrana, como ocurre en las neuronas, en donde la apertura y cierre de los canales de Na+ y K+ permite la propagación del impulso nervioso. Proteína transportadora

21
Transporte activo En el transporte activo las moléculas atraviesan la membrana en contra de su gradiente de concentración, por lo que consume energía obtenida del ATP. Este proceso se realiza mediante proteínas transportadoras. Ejemplos de sistemas de transporte activo son: - Bomba de Na+/K+ : Consiste en un complejo proteico de transmembrana que, mediante el gasto de un ATP, expulsa de la célula 3 iones Na+ e introduce dos iones K+ , con lo que contribuye a controlar la presión osmótica y el potencial de membrana. - Sistemas de cotransporte: Las proteínas de transmembrana transportan moléculas en contra de su gradiente térmico. Para esto utilizan la energía potencial almacenada en el gradiente iónico del Na+ , que se establece entre un lado y otro de la membrana gracias a la bomba de Na+/K*.

22
Transporte de macromoléculas
El transporte de macromoléculas y partículas a través de la membrana puede ser hacia el interior de la célula (endocitosis) y hacia el exterior (exocitosis). Ambos procesos tienen lugar mediante la formación de vesículas, que son pequeños sacos membranosos que se mueven de un sitio a otro por el citoplasma y ponen en comunicación a unos sistemas de membranas con otros.
 y hacia el exterior (exocitosis). Ambos procesos tienen lugar mediante la formación de vesículas, que son pequeños sacos membranosos que se mueven de un sitio a otro por el citoplasma y ponen en comunicación a unos sistemas de membranas con otros.")
23
Endocitosis Fagosoma Pseudópodos
Las sustancias que van a ser endocitadas son englobadas en invaginaciones de membrana plasmática que acaban cerrándose y forman vesículas intracelulares que contienen el material ingerido. Según el tamaño de las partículas endocitadas podemos distinguir dos tipos de endocitosis: Fagocitosis: El material que se ingiere es muy grande. La célula extiende unas prolongaciones de membrana llamadas pseudópodos, que rodean progresivamente a la partícula hasta formar un fagosoma (vesícula de gran tamaño). Estos materiales acaban digeridos por los lisosomas. Fagosoma Pseudópodos
. Estos materiales acaban digeridos por los lisosomas. Fagosoma. Pseudópodos.")
24
Pinocitosis: El material ingerido es liquido o pequeñas partículas, y queda englobado en vesículas que se forman a partir de depresiones de membrana llamadas pozos recubiertos. Estas regiones se caracterizan por la presencia de un armazón proteico formado por clatrina. A partir de ella se están formando continuamente vesículas de pinocitosis. - En los dos tipos de endocitosis participan proteínas especiales denominadas receptores de endocitosis, que se acumulan en las regiones de las membranas que realizan estos procesos y reconocen específicamente los materiales que van a ser ingeridos, uniéndose a ellos. De este modo las células incorporan por endocitosis únicamente las sustancias que les interesan. material vesícula

25
Exocitosis Endocitosis Exocitosis
Es el proceso contrario a la endocitosis. Mediante este proceso se secretan los materiales necesarios para renovar la membrana plasmática y los componentes de la matriz extracelular. También se vierten al exterior hormonas, neurotransmisores, enzimas digestivos… Endocitosis Exocitosis

26
UNIONES CELULARES Las uniones celulares son regiones especializadas de la membrana plasmática en las que se concentran proteínas de transmembrana especiales, mediante las cuales se establecen conexiones entre dos células o entre una célula y la matriz extracelular. Aparecen en todos los tejidos pero son especialmente importantes en los epitelios. Según su forma, las uniones celulares pueden ser: Zónulas: Son uniones que rodean totalmente a célula Máculas: Son uniones puntuales de forma redondeada Según su función, las uniones celulares pueden ser: Uniones ocluyentes, comunicantes o de anclaje.

27
Uniones ocluyentes: son uniones que se dan en las células que separan medios de composición muy diferente. Cierran el espacio intercelular impidiendo el paso de moléculas entre ellas. Medio externo de composición diferente Células epiteliales Medio interno Uniones de anclaje: son uniones frecuentes en tejidos sometidos a estrés mecánico como la piel, aumentan la resistencia de las células frente a tensiones mecánicas fuertes que acabarían rompiendo una sola célula. Existen tres tipos:

28
Zonulas adherens: Que conectan los filamentos de actina del citoesqueleto entre células vecinas.
Maculas adherens: Llamadas desmosomas. Anclan filamentos intermedios del citoesqueleto de células adyacentes. Hemidesmosomas: Anclan filamentos intermedios del citoesqueleto de una célula a la matriz extracelular. Uniones comunicantes: se denominan uniones gap. Son puntos de comunicación directa entre los citoplasmas de dos células, a través de los cuales intercambian iones y pequeñas moléculas.

29
Comunicación celular Las células son capaces de responder a los estímulos externos. Esta capacidad se denomina irritabilidad o excitabilidad. Estos estímulos se transmiten mediante moléculas de señalización producidas por una molécula señalizadora. Y son recogidos por receptores específicos en la célula diana, la cual convierte la señal extracelular en una intercelular mediante los sistemas de transducción de señales.

30
Moléculas de señalización
Según el tipo de comunicación en el que participen se clasifican en tres grupos: hormonas (comunicación endocrina) mediadores químicos locales (comunicación paracrina) y neurotransmisores (comunicación sináptica).
 mediadores químicos locales (comunicación paracrina) y neurotransmisores (comunicación sináptica).")
31
Sistemas de transducción de señales
Receptores Son proteínas de transmembrana que se encuentran distribuidas por la membrana plasmática. Tienen dos regiones funcionales distintas. Una región extracelular, que funciona como cerradura reconociendo específicamente la molécula señal determinada. Otra región citosólica, funciona de transmisor y es la responsable de la transducción de la señal. Los receptores se encuentran en un estado inactivo hasta que llega la molécula de señalización correspondiente. Sistemas de transducción de señales Los sistemas de transducción de señales se encargan de transformar las señales extracelulares en señales intracelulares. Primero una molécula señal llega a la célula diana y activa un receptor que transforma esta señal extracelular en una intracelular, llamada segundo mensajero. El Ion Ca+2 y el AMPc son los mas usados. Este segundo mensajero generado en gran cantidad se introduce hacia el interior celular y actúa sobre enzimas o factores intracelulares, que a su vez actúan sobre otros, iniciando una cadena de acontecimientos que transmiten la señal al interior celular. Finalmente estos factores actúan sobre las proteínas diana que serán las responsables de la respuesta de la célula. Una vez producida la respuesta la célula se encarga de degradar o aislar el primer y segundo mensajero, volviendo a la normalidad en solamente unos segundos

32
Activación del enzima que sintetiza el segundo mensajero

33
Síntesis de ATP en la mitocondria
El gradiente de protones se produce como resultado de la entrada de NADH (producido en las reacciones de oxido-reducción ) a la cadena transportadora de electrones . Los protones se acumulan en el espacio intermembrana hasta un gradiente de concentración tal que pueden ser utilizados para producir ATP.
 a la cadena transportadora de electrones . Los protones se acumulan en el espacio intermembrana hasta un gradiente de concentración tal que pueden ser utilizados para producir ATP.")
34
Los Protones (indicados por +) entran nuevamente en la matriz mitocondrial a través de los canales que forma el complejo enzimático de la ATP sintetasa. Esta entrada se acopla a la síntesis de ATP a partir de ADP y Fosfato (Pi)
.")
35
Cadena de transporte de electrones
El esquema en la parte superior muestra una mitocondria. En la animación, se observa como los iones H+ se acumulan en el compartimiento mitocondrial externo (espacio intermembrana). En la imagen inferior, se esquematiza lo que le sucede el hidrógeno cedido por el NADH a la cadena de transporte: los electrones son transferidos a lo largo de las proteinas de la cadena, y el protón al espacio intermembrana,donde genera un gradiente. Los protones re-entran pasando por el complejo ATP-sintetasas, generando ATP. Puntos claves: Los protones son transferidos a través de la membrana, desde la matriz al espacio intermembrana, como resultado del transporte de electrones que se originan cuando el NADH cede un hidrogeno. La continuada producción de esos protones crea un gradiente de protones. La ATP sintetasa es un gran complejo proteico con canales para protones que permiten la re-entrada de los mismos. La síntesis de ATPse produce como resultado de la corriente de protones fluyendo a través de la membrana: ADP + Pi ---> ATP
. En la imagen inferior, se esquematiza lo que le sucede el hidrógeno cedido por el NADH a la cadena de transporte: los electrones son transferidos a lo largo de las proteinas de la cadena, y el protón al espacio intermembrana,donde genera un gradiente. Los protones re-entran pasando por el complejo ATP-sintetasas, generando ATP. Puntos claves: Los protones son transferidos a través de la membrana, desde la matriz al espacio intermembrana, como resultado del transporte de electrones que se originan cuando el NADH cede un hidrogeno. La continuada producción de esos protones crea un gradiente de protones. La ATP sintetasa es un gran complejo proteico con canales para protones que permiten la re-entrada de los mismos. La síntesis de ATPse produce como resultado de la corriente de protones fluyendo a través de la membrana: ADP + Pi ---> ATP.")
36
Los protones son translocados a traves de la membrana, desde la matriz hasta el espacio intermembrana. Los electrones son transportados a lo largo de la membrana, por medio de una serie de proteinas transportadoras. ("carriers"). El oxígeno es el aceptor terminal del electrón, combinándose con electrones e iones H+para producir agua A medida que el NADH proporciona mas H+ y electrones a la Cadena Transportadora de electrones, se incrementa el gradiente de protones, con los H+ que se mantienen externamente a la membrana interna de la mitocondria, y los OH- dentro.
. El oxígeno es el aceptor terminal del electrón, combinándose con electrones e iones H+para producir agua. A medida que el NADH proporciona mas H+ y electrones a la Cadena Transportadora de electrones, se incrementa el gradiente de protones, con los H+ que se mantienen externamente a la membrana interna de la mitocondria, y los OH- dentro.")
37
Teoría Molecular del efecto de los anestésicos

38
Teoría molecular del efecto de los anestésicos
“El efecto es consecuencia de la acción”. “Una acción puede dar múltiples efectos”. “Una droga puede tener múltiples acciones”.

39
Teoría molecular del efecto de los anestésicos
Mecanismo de acción. Proceso íntimo celular que explica la acción del medicamento. Cualquier acción metabólica o fisiológica lo ocasiona. Selectividad. Mayor respuesta o afinidad de una droga por determinada célula o tejido donde se produce un efecto mayor.

40
Teoría molecular del efecto de los anestésicos
Factores que modifican la acción farmacológica de las drogas. Factores intrínsecos del medicamento. Comprenden todas las características físicas y químicas que el medicamento debe de poseer en base al principio activo en lugar y concentración adecuada para su absorción. Factores intrínsecos (fisiológicos) del paciente. Edad Peso Sexo Embarazo Biorritmo Factores farmacológicos. Dosis Absorción y excreción de las drogas Tolerancia
 del paciente. Edad Peso Sexo. Embarazo. Biorritmo. Factores farmacológicos. Dosis. Absorción y excreción de las drogas. Tolerancia.")
41
Teoría molecular del efecto de los anestésicos
Mecanismos de acción de los fármacos. El mecanismo de acción de un fármaco puede considerarse a los cuatro niveles diferentes siguientes: Sistemas corporales. Componentes tisulares. Células constituyentes. Moléculas.

42
Teoría molecular del efecto de los anestésicos
Dianas moleculares de los fármacos. Interacción fármaco – diana molecular Diana = componente macromolecular Componente lipídico o proteoliídico de la membrana celular Ácidos nucleicos Dianas moleculares de los fármacos: Receptores. Enzimas. Moléculas portadoras (cotransportadores o antitransportadores). Canales iónicos (accionados por ligando o accionados por voltaje). Dianas peculiares (iones metálicos, proteínas del surfactante, contenidos gastrointestinales). Ácidos nucleicos. Tres conceptos fundamentales: Afinidad es la tendencia a unirse a receptores. Eficacia es la relación entre la ocupación de receptores y la capacidad para iniciar una respuesta en los niveles molecular, tisular y sistémico. Actividad intrínseca es la capacidad de un sólo complejo fármaco - receptor para evocar una respuesta.
. Canales iónicos (accionados por ligando o accionados por voltaje). Dianas peculiares (iones metálicos, proteínas del surfactante, contenidos gastrointestinales). Ácidos nucleicos. Tres conceptos fundamentales: Afinidad es la tendencia a unirse a receptores. Eficacia es la relación entre la ocupación de receptores y la capacidad para iniciar una respuesta en los niveles molecular, tisular y sistémico. Actividad intrínseca es la capacidad de un sólo complejo fármaco - receptor para evocar una respuesta.")
43
Teoría molecular del efecto de los anestésicos
Agonismo. Es la producción de una respuesta molecular y celular a una interacción entre un fármaco (agonista) y un receptor que activa al receptor. La actividad intrínseca de un agonista completo se define como igual a 1. Agonismo parcial. Tiene lugar cuando un fármaco interacciona con un receptor y produce en promedio una respuesta molecular menor que 1. La actividad intrínseca molecular media está comprendida entre 0 y1. Antagonismo. Se produce cuando un fármaco interacciona con un receptor para inhibir la acción de un agonista. La actividad intrínseca molecular es 0. Agonismo inverso. Tiene lugar cuando un fármaco interacciona con un receptor para reducir su nivel de reposo de actividad molecular. La actividad intrínseca molecular es igual a -1. Agonismo inverso parcial. Tiene lugar cuando un fármaco interacciona con un receptor para reducir su nivel de reposo de actividad molecular. La actividad intrínseca molecular está comprendida entre 0 y -1.
 y un receptor que activa al receptor. La actividad intrínseca de un agonista completo se define como igual a 1. Agonismo parcial. Tiene lugar cuando un fármaco interacciona con un receptor y produce en promedio una respuesta molecular menor que 1. La actividad intrínseca molecular media está comprendida entre 0 y1. Antagonismo. Se produce cuando un fármaco interacciona con un receptor para inhibir la acción de un agonista. La actividad intrínseca molecular es 0. Agonismo inverso. Tiene lugar cuando un fármaco interacciona con un receptor para reducir su nivel de reposo de actividad molecular. La actividad intrínseca molecular es igual a -1. Agonismo inverso parcial. Tiene lugar cuando un fármaco interacciona con un receptor para reducir su nivel de reposo de actividad molecular. La actividad intrínseca molecular está comprendida entre 0 y -1.")
44
Teoría molecular del efecto de los anestésicos
Canales iónicos accionados por voltaje y por receptor. Los canales iónicos desempeñan un importante papel como dianas moleculares de los fármacos (y en muchos casos, como componentes de transducción). Los canales iónicos son proteínas que se extienden a través de la membrana (proteínas “transmembrana”, según el uso) y permiten el paso selectivo de iones específicos cuando el canal se abre. El paso de iones tiene lugar cuando la estructura molecular del canal lo permite. La estructura molecular define por tanto el estado de éste que puede ser: · Canal en reposo (es decir, cerrado, pero susceptible de abrirse en respuesta a un estímulo). · Estado activado (abierto). · Estado inactivado (es decir, cerrado, pero no susceptible de abrirse en respuesta a un estímulo).
. Los canales iónicos son proteínas que se extienden a través de la membrana (proteínas transmembrana , según el uso) y permiten el paso selectivo de iones específicos cuando el canal se abre. El paso de iones tiene lugar cuando la estructura molecular del canal lo permite. La estructura molecular define por tanto el estado de éste que puede ser: · Canal en reposo (es decir, cerrado, pero susceptible de abrirse en respuesta a un estímulo). · Estado activado (abierto). · Estado inactivado (es decir, cerrado, pero no susceptible de abrirse en respuesta a un estímulo).")
45
Teoría molecular del efecto de los anestésicos
Interacciones farmacológicas con canales de Na+. Los canales de sodio de neuronas, músculo cardíaco y músculo esquelético difieren ligeramente en estructura y composición de proteínas. Los fármacos que dificultan la apertura del canal de sodio durante la despolarización de la membrana suelen denominarse bloqueantes de canales de Na+ y, en cierta medida, discriminan entre los diferentes subtipos. Por ejemplo, la tetrodotoxina (una toxina que se encuentra en el pez globo, en algunas salamandras y en un tipo de pulpo) puede bloquear los canales de sodio en las neuronas y el músculo esquelético a concentraciones tan bajas como 10 nM, pero la concentración necesaria para bloquear los canales de sodio del músculo cardíaco es 100 veces mayor. Los anestésicos locales y los fármacos antiarrítmicos de la clase I bloquean los canales de sodio: · Anestésicos locales como la lidocaína y la bupivacaína pueden tener alguna selectividad relativa para la forma neuronal del canal de sodio, pero esta selectividad no es muy notable. Las pruebas actuales indican que la mayoría de los anestésicos locales interaccionan con la localización de reconocimiento del ligando sobre la superficie intracelular del canal y que el fármaco tiene que acceder al espacio intracelular para alcanzar su lugar de acción. Este mecanismo difiere del de la tetrodotoxina, molécula altamente cargada que accede a su lugar de reconocimiento de ligando cerca de la superficie extracelular del canal. · Actualmente se cree que los fármacos antiarrítmicos de la clase I, empleados para tratar ciertas formas de arritmia cardíaca, interaccionan principalmente con un lugar de reconocimiento de ligando de localización intracelular. Parecen dividirse en tres clases (clases Ia, Ib y Ic), según la forma en que su actividad depende del estado del canal iónico y de la cinética aparente de unión y disociación (llamada desunión en este contexto) con el canal en sus tres estados.
 puede bloquear los canales de sodio en las neuronas y el músculo esquelético a concentraciones tan bajas como 10 nM, pero la concentración necesaria para bloquear los canales de sodio del músculo cardíaco es 100 veces mayor. Los anestésicos locales y los fármacos antiarrítmicos de la clase I bloquean los canales de sodio: · Anestésicos locales como la lidocaína y la bupivacaína pueden tener alguna selectividad relativa para la forma neuronal del canal de sodio, pero esta selectividad no es muy notable. Las pruebas actuales indican que la mayoría de los anestésicos locales interaccionan con la localización de reconocimiento del ligando sobre la superficie intracelular del canal y que el fármaco tiene que acceder al espacio intracelular para alcanzar su lugar de acción. Este mecanismo difiere del de la tetrodotoxina, molécula altamente cargada que accede a su lugar de reconocimiento de ligando cerca de la superficie extracelular del canal. · Actualmente se cree que los fármacos antiarrítmicos de la clase I, empleados para tratar ciertas formas de arritmia cardíaca, interaccionan principalmente con un lugar de reconocimiento de ligando de localización intracelular. Parecen dividirse en tres clases (clases Ia, Ib y Ic), según la forma en que su actividad depende del estado del canal iónico y de la cinética aparente de unión y disociación (llamada desunión en este contexto) con el canal en sus tres estados.")
46
Teoría molecular del efecto de los anestésicos
Interacciones farmacológicas con los canales de Ca2+. Al menos cuatro tipo de canal de calcio en la membrana plasmática permiten selectivamente la entrada de iones calcio en las células. Estos canales de calcio se encuentran en muchos tipos diferentes de tejidos. El mejor caracterizado y más importante desde el punto de vista clínico es el canal de calcio de tipo L (del inglés large, grande) que se abre durante la despolarización y después se inactiva (más despacio que el canal de sodio), mediante accionamiento dependiente del voltaje. Es el canal de calcio predominante en el músculo cardíaco y el músculo liso y es bloqueado por diversos fármacos importantes en clínica. Hay tres clases comunes de antagonista de Ca2+ de tipo L clínicamente importantes: 1. Los derivados de la benzotiacepina (por ejemplo el diltiazem). 2. Las fenetilalquilaminas (por ejemplo el verapamilo). 3. Las 1,4 dihidropiridinas (por ejemplo nifedipino, amlodipino). Otros tipos de canales de calcio, a saber N, P y T pueden ser bloqueados selectivamente por diversos compuestos, en especial por péptidos obtenidos a partir de ciertos venenos de moluscos. Pueden surgir fármacos originales con eficacia farmacoterapéutica en seres humanos a partir del bloqueo selectivo de esos canales.
 que se abre durante la despolarización y después se inactiva (más despacio que el canal de sodio), mediante accionamiento dependiente del voltaje. Es el canal de calcio predominante en el músculo cardíaco y el músculo liso y es bloqueado por diversos fármacos importantes en clínica. Hay tres clases comunes de antagonista de Ca2+ de tipo L clínicamente importantes: 1. Los derivados de la benzotiacepina (por ejemplo el diltiazem). 2. Las fenetilalquilaminas (por ejemplo el verapamilo). 3. Las 1,4 dihidropiridinas (por ejemplo nifedipino, amlodipino). Otros tipos de canales de calcio, a saber N, P y T pueden ser bloqueados selectivamente por diversos compuestos, en especial por péptidos obtenidos a partir de ciertos venenos de moluscos. Pueden surgir fármacos originales con eficacia farmacoterapéutica en seres humanos a partir del bloqueo selectivo de esos canales.")
47
Teoría molecular del efecto de los anestésicos
Interacciones farmacológicas con los canales de K+. La apertura con acción selectiva para el potasio conduce a la generación de corrientes dirigidas hacia fuera (hiperpolarizantes). Hay muchos tipo de canales de potasio y constituyen un grupo muy heterogéneo, en lo que se refiere a su dependencia de voltaje y tiempo y a su accionamiento por ligando. Hay más de diez tipos diferentes, cuya expresión varía según el tipo de tejido; a su vez, algunos tejidos expresan numerosos tipos.
. Hay muchos tipo de canales de potasio y constituyen un grupo muy heterogéneo, en lo que se refiere a su dependencia de voltaje y tiempo y a su accionamiento por ligando. Hay más de diez tipos diferentes, cuya expresión varía según el tipo de tejido; a su vez, algunos tejidos expresan numerosos tipos.")
48
Teoría molecular del efecto de los anestésicos
Otros canales iónicos accionados por voltaje. Aunque la mayoría de la bibliografía científica sobre canales iónicos se ha concentrado en los canales de cationes (sodio, calcio y potasio), recientemente se ha hecho más evidente que existen canales accionados por voltaje para aniones, por ejemplo el Cl-. Los canales de cloruro se encuentran tanto en el sistema nervioso central como en el periférico. Algunos no son selectivos para un sólo ión, como por ejemplo el canal que permite el flujo iónico de los iones sodio y calcio en el corazón. Es así que podemos indicar que los receptores se pueden reunir en cuatro grandes superfamilias, a saber: 1. Canal accionado por receptor. 2. Receptores ligados a proteínas G. 3. Receptores que son enzimas. 4. Receptores ligados al ADN.
, recientemente se ha hecho más evidente que existen canales accionados por voltaje para aniones, por ejemplo el Cl-. Los canales de cloruro se encuentran tanto en el sistema nervioso central como en el periférico. Algunos no son selectivos para un sólo ión, como por ejemplo el canal que permite el flujo iónico de los iones sodio y calcio en el corazón. Es así que podemos indicar que los receptores se pueden reunir en cuatro grandes superfamilias, a saber: 1. Canal accionado por receptor. 2. Receptores ligados a proteínas G. 3. Receptores que son enzimas. 4. Receptores ligados al ADN.")
49
…………………………………………………

Presentaciones similares












.>")

>")


.>")

