Descargar la presentación
La descarga está en progreso. Por favor, espere

1
ENZIMAS y CINÉTICA ENZIMÁTICA

2
Cinética experimental
Al derivar las ecuaciones clásicas utilizadas en el estudio experimental de la cinética enzimática se supone que la concentración de la enzima es muy pequeña comparada con la concentración del sustrato Esto es generalmente válido, debido a la gran eficiencia de las enzimas Si se miden las velocidades iniciales, la cantidad de productos formados es despreciable y la disminución de la concentración del sustrato es también despreciable y la velocidad de la reacción es directamente proporcional a la concentración inicial de sustrato Se debe distinguir reacciones con un sustrato de reacciones multisustrato

3
Reacciones con un sustrato Mecanismo de Michaelis Menten
Las enzimas que catalizan reacciones con un sustrato son: hidrolasas, isomerasas y la mayoría de las liasas En un primer paso el sustrato se une a un sitio específico de la enzima, llamado sitio activo, formando el complejo enzima- sustrato de acuerdo al siguiente mecanismo E + S → ES → E + P Que se denomina mecanismo de Michaelis-Menten La unión del sustrato a la enzima se realiza por fuerzas de tipo físico: fuerzas de Van der Waals, interacciones hidrofóbicas e interiónicas El complejo enzima sustrato permite la formación del producto, que se separa de la enzima,posteriormente, regenerándose ésta

4
Ecuación de Michaelis Menten
Como la formación del complejo se produce por interacciones de tipo físico puede suponerse que se llega rápidamente al equilibrio y se define una constante de disociación KS, como Ks= [E] [S] / [ES] Utilizando la aproximación del equilibrio rápido Michaelis y Menten en 1913, llegaron a establecer la ecuación v = Vm [S] / Ks + [S] Vm es la velocidad de la reacción cuando toda la enzima está unida al sustrato ( está saturada Vm = k2 (Eo) @hotmail.com)

5
Planteamiento de Briggs-Haldane
El método del equilibrio rápido supone que la reacción de formación del producto es mucho más lenta que el proceso de formación del complejo, sin embargo esto no siempre se cumple Briggs y Haldane propusieron otra deducción basada en el estado estacionario, obteniendo una ecuación: v = Vm [S] / ( k-1+k2/k1) + [S] Donde k-1+ k2/k1 = Km Actualmente la ecuación de Michaelis Menten se escribe V = Vm [S] / Km + [S]
 + [S] Donde k-1+ k2/k1 = Km. Actualmente la ecuación de Michaelis Menten se escribe. V = Vm [S] / Km + [S]")
6
Representación gráfica de la Ecuación de Michaelis Menten
Las ecuaciones deducidas por MM y BH corresponden a una hipérbole, Ks en la deducción de MM, es sustituida por Km, en la BH Se pueden obtener ecuaciones hiperbólicas similares si se consideran varias reacciones en la etapa de formación del producto, solo que Km será la combinación de más constantes cinéticas La ecuación de Michaelis Menten responde a la cinética de muchísimas enzimas. Se dice entonces que tienen una cinética michaeliana

7
Cinética michaeliana

8
Parámetros de la ecuación de MM
Km es la concentración de sustrato que corresponde la mitad de la velocidad máxima Su valor depende del sistema en estudio ( naturaleza de la enzima y el sustrato ), pH y fuerza iónica Es una medida de la concentración de sustrato necesaria para lograr una catálisis eficaz Se la relaciona con la afinidad de la enzima por un sustrato dado. A mayor Km menor afinidad de la enzima por el sustrato y viceversa, sin embargo esto será estrictamente válido si Km = Ks ,o sea si se cumple el equilibrio rápido
, pH y fuerza iónica. Es una medida de la concentración de sustrato necesaria para lograr una catálisis eficaz. Se la relaciona con la afinidad de la enzima por un sustrato dado. A mayor Km menor afinidad de la enzima por el sustrato y viceversa, sin embargo esto será estrictamente válido si Km = Ks ,o sea si se cumple el equilibrio rápido.")
9
La velocidad de la reacción se puede considerar proporcional a la concentración del complejo ES y v = kcat [ES] Cuando toda la enzima está unida al sustrato se alcanza la velocidad máxima de la reacción Vm, entonces [ES] = Eo y Vm= kcat [Eo] kcat es una medida directa de la producción catalítica del producto en condiciones óptimas ( enzimas saturadas) Se puede considerar como el número de moléculas de sustrato transformadas por una molécula de enzima en la unidad de tiempo
![La velocidad de la reacción se puede considerar proporcional a la concentración del complejo ES y v = kcat [ES]](http://slideplayer.es/slide/10362356/33/images/9/La+velocidad+de+la+reacci%C3%B3n+se+puede+considerar+proporcional+a+la+concentraci%C3%B3n+del+complejo+ES+y+v+%3D+kcat+%5BES%5D.jpg "Cuando toda la enzima está unida al sustrato se alcanza la velocidad máxima de la reacción Vm, entonces [ES] = Eo y. Vm= kcat [Eo] kcat es una medida directa de la producción catalítica del producto en condiciones óptimas ( enzimas saturadas) Se puede considerar como el número de moléculas de sustrato transformadas por una molécula de enzima en la unidad de tiempo.")
10
Especificidad de la enzima
Los valores de Km y Vm(para calcular kcat) son importantes para -determinar la especificidad de una enzima respecto a cierto sustrato - decidir entre un mecanismo rápido y el estado estacionario - conocer el rol de la enzima en el metabolismo Para una enzima que puede trabajar con dos sustratos, como [ES] = [E][S]/Km v = kcat/Km [E][S]tendremos v1/v2 =(kcat/Km)1[E][S]1 / (kcat/Km)2 [E][S]2 y v1/v2 = (kcat/Km)1 / (kcat/Km)2 a concentraciones iguales de sustrato La relación kcat/Km puede considerarse como una medida de la especificidad de la enzima por un sustrato
 son importantes para. -determinar la especificidad de una enzima respecto a cierto sustrato. - decidir entre un mecanismo rápido y el estado estacionario. - conocer el rol de la enzima en el metabolismo. Para una enzima que puede trabajar con dos sustratos, como [ES] = [E][S]/Km v = kcat/Km [E][S]tendremos. v1/v2 =(kcat/Km)1[E][S]1 / (kcat/Km)2 [E][S]2. y v1/v2 = (kcat/Km)1 / (kcat/Km)2 a concentraciones iguales de sustrato. La relación kcat/Km puede considerarse como una medida de la especificidad de la enzima por un sustrato.")
12
Cálculo de Km, kcat y Vm Km y Vm se pueden calcular a partir de las transformaciones lineales de la ecuación de MM La determinación de Vm, y a partir de allí la de Km, utilizando la gráfica hiperbólica no da resultados correctos ya que el valor de Vm se tendría solo a concentración muy grandes Para dicho cálculo se pueden emplear gráficos obtenidos por linealización de la ec de MM A partir de Vm y la concentración empleada de la enzima, se obtiene el valor de kcat = Vm/ [Eo]

13
Representaciones lineales
Se han propuesto varias transformaciones de la ec. De MM que conducen a funciones lineales, las más utilizadas son: Representación de dobles recíprocos o de Lineweaver-Burk Representación de Eadie- Hoftee Representación de Hanes Woolf Representación directa de Eisenthal- Cornish Bowden

14
Representación de Lineweaver- Burk
Se obtiene la ecuación al tomar la inversa de ambos términos en la ec de MM 1/V = 1/Vm + Km/Vm . 1/[S] Al representar 1/V frente a 1/ [S], se obtendrá una recta, si la cinética es michaeliana, La intercepción con el eje de abcisas es 1/Vm y con el eje de ordenads es -1/Km Este tipo de representación es a menudo empleada al estudiar los tipos de inhibición

15
La intercepción de la recta con el eje de ordenadas es 1/Vm y la intercepción con el eje de abcisas es – 1/Km

16
Representación de Eadie Hoftee
Multiplicando v por Km + [S] y luego dividiendo el resultado por [S] se llega a la expresión V = Vm – Km V/[S] Ec de Eadie-Hoftee Esta ecuación tiene una sola recíproca y los puntos salen igualmente espaciados La ordenada es Vm La pendiente es - Km

17
Representación de Eadie-Hoftee

18
Representación de Hanes Woolf
Otra representación a menudo empleada es la de Hanes-Woolf, la ecuación correspondiente se obtiene multiplicando por[S] la ec de dobles recíprocas y ordenando [S]/V = Km /Vm /Vm . [S] Se debe graficar [S]/V en ordenadas y [S] en abcisas La ordenada en el origen es Km/Vm y la pendiente es 1/Vm La intercepción de la recta con el eje de ordenadas es igual a - Km

19
Representación directa
Cornish-Bowden y Eisenthal han propuesto otra forma de representar los datos cinéticos Se consideran Km y Vm como variables, x e y Se grafican directamente los valores de v y [S] Los distintos pares de valores de v y S, se unen para dar distintas rectas Las varias rectas trazadas se deben cruzae en un punto cuyas coordenadas son Km y V En realidad , se promedian los valores ya que no todas las rectas se cruzan en el mismo punto

20
Representación directa
La ecuación de Michaelis Menten puede ser reordenada como Vm = V + V/ [S]. Km, donde Vm se toma como la variable y Km como la variable x, así la ordenada en el origen es V y -[S] es el valor de abcisas El punto de intercepción de las diversas rectas que se pueden obtener con distintos valores de v y [S]tiene como coordenadas Km (x) y Vm (y)
 y Vm (y)")
21
Inhibición enzimática
La velocidad de las reacciones catalizadas por enzimas puede disminuir o anularse en presencia de ciertas sustancias denominadas inhibidores El estudio de la inhibicion es importante desde muchos puntos de vista Muchos fármacos actúan como inhibidores, el empleo de ciertos inhibidores ha permitido obtener información sobre el sitio activo de la enzima. Se emplean inhibidores como herbicidas

22
Tipos de inhibición Se distinguen:
Inhibición reversible- La unión del Inhibidor y la enzima es un proceso reversible que está regulado por constantes de equilibrio. La enzima puede inhibirse en mayor o menor grado Inhibición irreversible: una vez que el inhibidor se unió a la enzima ya no se separa, y la enzima queda inutilizada , se vuelve inactiva

23
Tipos de inhibición reversible
Se distinguen cuatro formas de inhibición reversible - Inhibición competitiva -Inhibición acompetitiva -Inhibición mixta -Inhibición no competitiva. Esta última es solo un caso especial de la inhibición mixta Corresponden a la inhibición lineal o simple

24
Inhibición lineal e hiperbólica
En la inhibición reversible el inhibidor forma un complejo dinámico con la enzima que posee propiedades catalíticas diferentes a las de la enzima libre Si la enzima unida al inhibidor, pierde completamente su actividad catalítica cuando está saturada, se tiene la inhibición completa o lineal( llamada así porque Km/Vm y 1/ Vm dependen linealmente de la concentración del inhibidor) Si el complejo enzima-inhibidor, en condiciones de saturación mantiene algo de actividad se denomina inhibición hiperbólica(por la forma que tienen las representaciones de KM/Vm o 1/Vm frente a (I) ) o inhibición parcial
 Si el complejo enzima-inhibidor, en condiciones de saturación mantiene algo de actividad se denomina inhibición hiperbólica(por la forma que tienen las representaciones de KM/Vm o 1/Vm frente a (I) ) o inhibición parcial.")
25
Inhibición competitiva
La Enzima puede unirse al sustrato o al inhibidor pero no a ambos simultáneamente Un inhibidor competitivo es una sustancia que al combinarse con la enzima impide que ésta se una al sustrato En muchos casos un inhibidor competitivo tiene una estructura análoga a la del sustrato,ocupando el sitio activo, por ejemplo la sulfanilamida se asemejan al ácido paraaminobenzoico e inhibe a la enzima que participa en la síntesis del ácido benzoico,vital para las bacterias

26
El inhibidor competitivo puede ser un análogo o derivado del sustrato no metabolizable, un sutrato alternativo o un producto de la reacción Se han estudiado casos de inhibición competitiva en que el inhibidor se une a la enzima en un lugar diferente al sitio activo, pero produce un cambio en la conformación del mismo e impide que el sustrato se una a la enzima El aumento de sustrato produce una competencia con el inhibidor por la enzima y para cierta concentración de sustrato se puede llegar a la misma velocidad máxima

27
Esquema de una inhibición competitiva
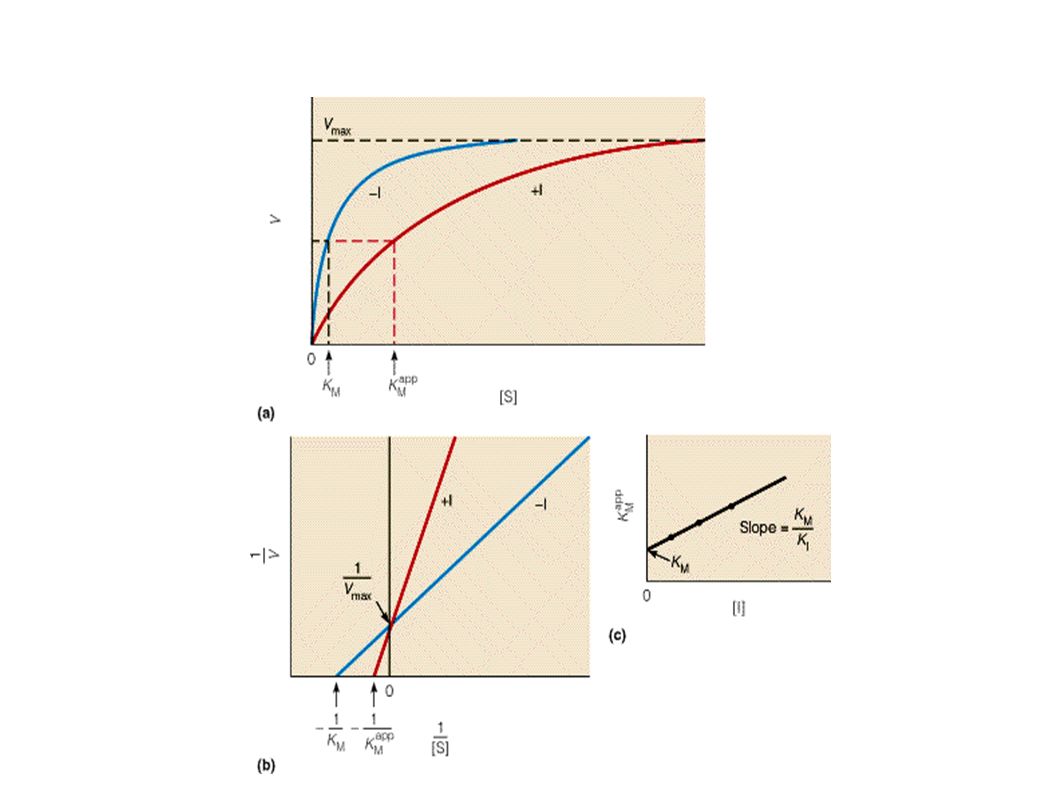
Ks E + S → ES → E + P KEI = [I] [E]/ [EI] + I KEI ↓ EI A partir de este esquema se deduce la ec. V= Vm [S] / (Km + KEI [I]/) + S KEI o KI es la constante de disociación del inhibidor Km + [I]/Ki = Km aparente que es el valor obtenido experimentalmente
 + S. KEI o KI es la constante de disociación del inhibidor. Km + [I]/Ki = Km aparente que es el valor obtenido experimentalmente.")
29
La representación recíproca es
1/V = 1/ Vm + Km(1+[I]/Ki / Vm . 1/ [S] La grafica de LB muestra una familia de rectas que interceptan en el eje de ordenadas. La intercepción muestra que Vm tiene el mismo valor sin ihnibidor o con distintas cantidades de inhibidor, sin embargo se necesita una cantidad mayor de sustrato para obtener la misma velocidad máxima Km aparente por otro lado disminuye a medida que aumenta la concentración de inhibidor Es muy común en muchas reacciones con un sustrato:

30
Inhibición no competitiva
El inhibidor y el sustrato se unen reversiblemente a la enzima pero en sitios diferentes. Se pueden formar 3 tipos de complejos ES, EI y ESI E + S → ES → E + P Ks k Ks = (E) (S)/ (ES) I I KEI = (E)(I) / (EI) IKEI ↓ KESI ↓ KESI = (EI) (S) / (EIS) EI S → ESI KESI
 (S)/ (ES) I I KEI = (E)(I) / (EI) IKEI ↓ KESI ↓ KESI = (EI) (S) / (EIS) EI + S → ESI. KESI.")
31
Se definen Ks = [E] [S]/[ES] = [EI] [S]/ ESI Ki = [E] [I]/ [EI] = [ES][I]/[ESI] Parte de la enzima está unida al inhibidor y al inhibidor y sustrato como EI y EIS, por tanto aunque se añade más sustrato parte del complejo formado ES se une a I, y es inactivo por lo tanto la velocidad máxima disminuye y la disminución es proporcional a la [I] Este tipo de inhibición es rara en reacciones con un sustrato, en cambio se han observado muchos casos en reacciones multisustrato
![Se definen Ks = [E] [S]/[ES] = [EI] [S]/ ESI. Ki = [E] [I]/ [EI] = [ES][I]/[ESI]](http://slideplayer.es/slide/10362356/33/images/31/Se+definen+Ks+%3D+%5BE%5D+%5BS%5D%2F%5BES%5D+%3D+%5BEI%5D+%5BS%5D%2F+ESI.+Ki+%3D+%5BE%5D+%5BI%5D%2F+%5BEI%5D+%3D+%5BES%5D%5BI%5D%2F%5BESI%5D.jpg "Parte de la enzima está unida al inhibidor y al inhibidor y sustrato como EI y EIS, por tanto aunque se añade más sustrato parte del complejo formado ES se une a I, y es inactivo por lo tanto la velocidad máxima disminuye y la disminución es proporcional a la [I] Este tipo de inhibición es rara en reacciones con un sustrato, en cambio se han observado muchos casos en reacciones multisustrato.")
32
La ecuación correspondiente es_
Vm / (1+ [I]/Ki) . [S] V= Km [S] La velocidad máxima disminuye y la Km permanece inalterada Las rectas que corresponden a distintas concentraciones de inhibidor ,se interceptan en el eje de abcisas en un punto
 . [S] V= Km + [S] La velocidad máxima disminuye y la Km permanece inalterada. Las rectas que corresponden a distintas concentraciones de inhibidor ,se interceptan en el eje de abcisas en un punto.")
33
Inhibición acompetitiva
Un inhibidor acompetitivo se une al complejo ES, formando un complejo ternario ESI inactivo La inhibición acompetitiva pura es rara en sistemas de un sustrato, se observa generalmente en reacciones con 2 sustratos E + S → ES → E + P + I ESI

34
De acuerdo al esquema anterior se deduce la ecuación
Vmax/ (1+ [I]/Ki) V = Km/ (1+ [I]/Ki) + [S] Las gráficas de dobles recíprocas obtenidas sin inhibidor o en presencia de distintas concentraciones del inhibidor son rectas paralelas entre si, debido a que Vm y Km disminuyen a medida que [I] aumenta Se han observado ṕocos casos en reacciones con un sustrato ( inhibición de fosfatasa alcalina de ratón con Lfenil alanina) , pero varios ejemplos en reacciones con mas de un sustrato (inhibición de metionina adenosiltranferasa de levadura por S-adenosilmetonina)
 V = Km/ (1+ [I]/Ki) + [S] Las gráficas de dobles recíprocas obtenidas sin inhibidor o en presencia de distintas concentraciones del inhibidor son rectas paralelas entre si, debido a que Vm y Km disminuyen a medida que [I] aumenta. Se han observado ṕocos casos en reacciones con un sustrato ( inhibición de fosfatasa alcalina de ratón con Lfenil alanina) , pero varios ejemplos en reacciones con mas de un sustrato (inhibición de metionina adenosiltranferasa de levadura por S-adenosilmetonina)")
35
Gŕafica para la inhibición acompetitiva

36
Inhibición mixta Se forman tanto complejos binarios ES y EI como ternario ESI Las constantes de disociación de los complejos ternarios y binarios correspondientes al sustrato o al inhibidor son diferentes Se pueden relacionar Ki con K para la descomposición del complejo ternario formado en ES e I, o Ks con K para la disociación del complejo EIS en EI yS K = α Ki o K = α Ks

37
Esquema Ks E + S → ES → E + P I I Ki ↓ ↓ αKi EI + S → ESI αKs kp

38
La ecuación correspondiente a la inhibición mixta clásica es
Vmax /( 1 +[I]/αKi ) [S] V = Ks (1 + [I]/Ki) + (1 +[I] / α Ki) [S] Las líneas que se obtienen con distintas concentraciones de inhibidor se interceptan en un punto que puede estar por encima o por debajo del eje de abcisas El valor de la abcisas que corresponde al punto de intercepción de las rectas es - 1/ α Ki La intercepción de la línea que corresponde a cierta concentración del inhibidor con el eje de abcisas es - 1+[I]/αKi / 1 +[I] /Ki
 [S] V = Ks (1 + [I]/Ki) + (1 +[I] / α Ki) [S] Las líneas que se obtienen con distintas concentraciones de inhibidor se interceptan en un punto que puede estar por encima o por debajo del eje de abcisas. El valor de la abcisas que corresponde al punto de intercepción de las rectas es - 1/ α Ki. La intercepción de la línea que corresponde a cierta concentración del inhibidor con el eje de abcisas es. - 1+[I]/αKi / 1 +[I] /Ki.")
39
Gráficas obtenidas en la inhibición mixta a distintas concentraciones del inhibidor

40
Diferencias entre los distintos tipos de inhibición lineal reversible
Los distintos tipos de inhibición lineal reversible se distinguen desde el punto de vista cinético por las diferencias entre los valores de Km y Km ap ( en presencia del inhibidor), y entre Vmax y Vmax aparente (con inhibidor), es decir por el efecto del inhibidor sobre los parámetros cinéticos, antes que por el tipo de unión, dificil de determinar Vmax Km Inhibición competitiva no varía aumenta Inhibició acompetitiva disminuye disminuye Inhibición no competiva disminuye no varía Inhibición mixta disminuye aumenta
, y entre Vmax y Vmax aparente (con inhibidor), es decir por el efecto del inhibidor sobre los parámetros cinéticos, antes que por el tipo de unión, dificil de determinar. Vmax. Km. Inhibición competitiva no varía aumenta. Inhibició acompetitiva disminuye disminuye. Inhibición no competiva disminuye no varía. Inhibición mixta disminuye aumenta.")
41
Resumen Los casos limitantes de inhibición lineal son: inhibición competitiva(específica) y acompetitiva(catalítica). En aquellos casos en donde el tipo de inhibición no cae en ninguno de los modelos anteriores, se habla de inhibición mixta La inhibición pura no competitiva es un caso de inhibición mixta donde Kei = Kesi La IUBMB recomienda considerar los distintos tipos de inhibición basándose en los efectos sobre Vm/Km y Vm

42
Si Vm/Km disminuye, se dice que hay un componente competitivo
Si el inhibidor no tiene efecto sobre Vm la inhibición es competitiva Si hay el valor aparente de Vm disminuye hay un componente acompetitivo Si el inhibidor no tiene ningún efecto sobre Vm/Km la inhibición es competitiva Si ambos componentes competitivo y acompetitivo están presentes la inhibición es mixta

43
Inhibición irreversible
En algunos casos el inhibidor queda fuertemente unido a la enzima por enlaces covalente En este caso la queda inutilizada y pierde en forma permanente su actividad catalítica Ejemplos de este tipo de inhibición se observa en el efecto tóxico de metales que forman compuestos covalentes con las enzimas

44
Otros casos de inhbición
Inhibidores pseudo irreversibles o fuertemente unidos Inhibidores suicidas Inhibición por producto Inhibición por sustrato Inhibición feedback

45
Enzimas con múltiples sitios catalíticos
Muchas enzimas están constituidas por múltiples cadenas formando oligómeros de 2, cuatro, seis u ocho monómeros. Cada uno de estos monómeros puede tener un sitio activo al que se une el sustrato Estas enzimas pueden desempeñar un papel muy importante en el metabolismo Se distinguen dos grupos_: - Todos los sitios son idénticos e independientes unos de otros : no presentan cooperatividad Si la presencia de una sustrato en un sitio, influye en la ocupación de los otros sitios activos, se dice que existe cooperatividad

46
Enzimas con sitios no cooperativos
Considerando un dímero se pueden formar dos complejos binarios ES y SE, y ESE, ternario,todos con las mismas constantes de disociación Ks y la misma constante catalítica E + S → ES → E + P S S ↓ ↓ E+ P ← SE + S → ESE → SE + P ↓ ES + P

47
Ecuación para un sistema multisustratos no cooperativo
La velocidad dependerá de la concentración de todos los complejos formados V = k(ES) + k(SE) +2 k (ESE) Reemplazando y simplificando se tiene V/ Vmax = S/Ks (1+ S/Ks) / (1 + S/Ks) y simplificando y reordenando, se tendrá V/ Vmax = S/ Ks + S ec idéntica a la de MM Este tipo de enzimas darán cinética michaeliana y por lo tanto por experiencias cinéticas no se pueden diferenciar de las enzimas con un sito activo
 + k(SE) +2 k (ESE) Reemplazando y simplificando se tiene. V/ Vmax = S/Ks (1+ S/Ks) / (1 + S/Ks) 2 y simplificando y reordenando, se tendrá. V/ Vmax = S/ Ks + S ec idéntica a la de MM. Este tipo de enzimas darán cinética michaeliana y por lo tanto por experiencias cinéticas no se pueden diferenciar de las enzimas con un sito activo.")
48
Si se tiene un trímero la ecuación que se obtiene es
V/Vmax = S/Ks (1+ S/Ks)3 / (1 + S/Ks)4 Y simplificando y ordenando también se tendrá la ecuación de MM
3 / (1 + S/Ks)4. Y simplificando y ordenando también se tendrá la ecuación de MM.")
50
Aspartato carbamoiltransferasa, activada por ATP e inhibida por CTP

51
Enzimas que presentan cooperatividad
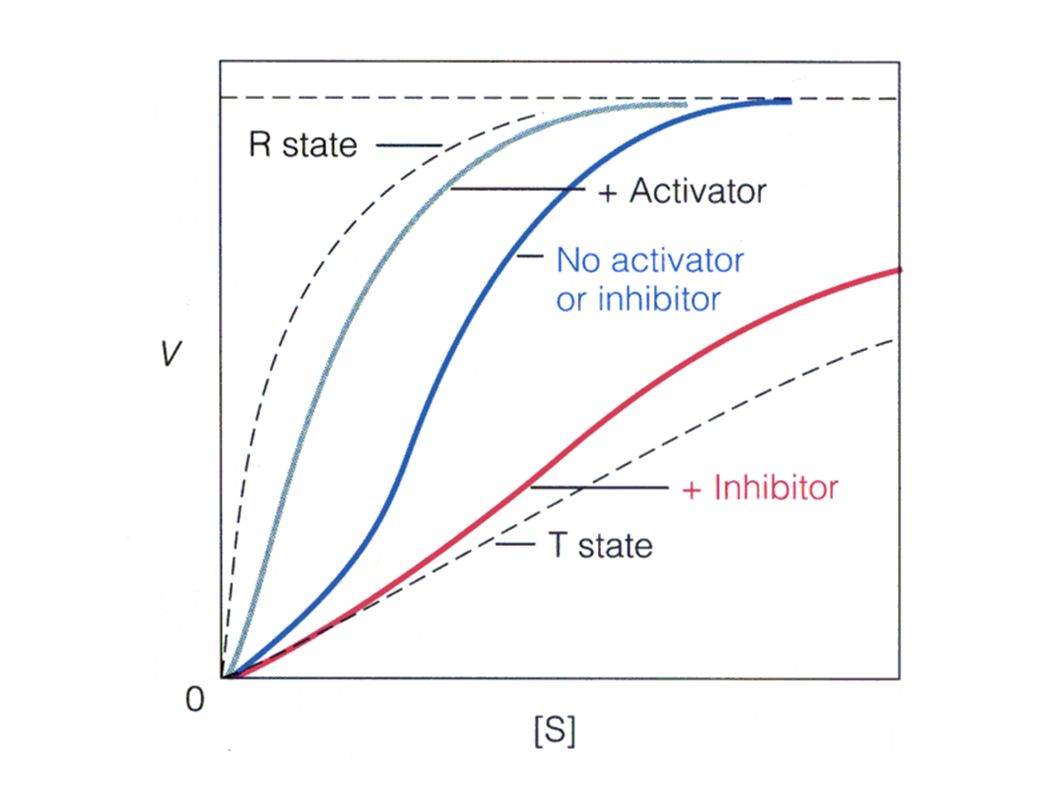
S i la unión del sustrato a un sitio activo produce cambios estructurales o electrónicos en la enzima que alteran la afinidad de la enzima al siguiente sustrato que se une se presenta cooperatividad Las enzimas de este tipo no siguen la cinética michaeliana y se llaman enzimas alostéricas ( otro sitio), generalmente las curvas tienen forma sigmoidea y la cinética se denomina sigmoidal
, generalmente las curvas tienen forma sigmoidea y la cinética se denomina sigmoidal.")
52
Enzimas alostéricas Generalmente la unión de la primera molécula de sustrato favorece la unión de la segunda molécula y ésta de la siguiente y así sucesivamente. Se dice que hay cooperatividad positiva En algunos casos muy raros la unión de un sustrato defavorece la unión del siguiente, teniéndose entonces cooperatividad negativa Cuando la unión del sustrato es la que produce cambios en el comportamiento de la enzima se habla de respuesta homotrópica

53
Activadores e inhibidores de las enzimas alostéricas
Este grupo de enzimas es también sensible a la unión de otras moléculas distintas del sustrato llamadas efectores o moduladores que producen alteraciones en el comportamiento de la enzimas respecto al sustrato, produciendo un aumento de la actividad ( efectores positivos o activadores) o una disminución de la actividad (efectores negativos o inhibidores). En este caso se habla de efectos heterotrópicos.
 o una disminución de la actividad (efectores negativos o inhibidores). En este caso se habla de efectos heterotrópicos.")
54
Diferencia entre la cinética michaeliana y la cinética sigmoidal

55
Papel regulador de las enzimas alostéricas
Las enzimas alostéricas tienen las siguientes características a)Presentan gran sensibilidad a las variaciones de concentración del sustrato, pueden pasar de una actividad muy baja a concentraciones pequeñas a gran actividad con concentraciones mayores. En una enzima de cinética michaeliana para aumentar la velocidad de 10 % Vm a 90 % Vm , la concentración debe aumentar 81 veces, y en una enzima alostérica, mucho menos, para 2 sitios activos es necesario que aumente solo 9 veces b) pueden ser reguladas por otras moléculas (efectores) que modifican la actividad de la enzima según las necesidades metabólicas
Presentan gran sensibilidad a las variaciones de concentración del sustrato, pueden pasar de una actividad muy baja a concentraciones pequeñas a gran actividad con concentraciones mayores. En una enzima de cinética michaeliana para aumentar la velocidad de 10 % Vm a 90 % Vm , la concentración debe aumentar 81 veces, y en una enzima alostérica, mucho menos, para 2 sitios activos es necesario que aumente solo 9 veces. b) pueden ser reguladas por otras moléculas (efectores) que modifican la actividad de la enzima según las necesidades metabólicas.")
56
Tratamiento de las enzimas alostéricas
Para estudiar las enzimas alostéricas se pueden emplear varios métodos - la ecuación de Hill - la ecuación de Adair - ecuaciones derivadas del modelo de Monod- Wyman-Changeaux (MWCh) - ecuaciones derivadas del modelo de Koshlan- Nemethy-Filmer (KNF)
 - ecuaciones derivadas del modelo de Koshlan- Nemethy-Filmer (KNF)")
57
V/Vmax= S/Ks + S2/ a Ks 2 /1 + 2S/Ks + S2 /aKs
Considerando una enzima alostérica con 2 sitios activos se puede llegar a la ecuación V/Vmax= S/Ks + S2/ a Ks 2 /1 + 2S/Ks + S2 /aKs Para una enzima con 4 sitios de unión se tendría S/Ks + 3S2/aKs2 + 3 S3/a2bKs3 + S4/a3b2cKs4 v/Vmax= 1+4 S/Ks + 6S2/aKs2 + 4 S3/a2bKs3 + S4/a3b2cKs4 a, b, c etc. son los llamados factores de interrelacción que relacionan las constantes para los distintos complejos que se pueden formar con Ks O sea son los coeficientes por los cuales se multiplican las constantes intrínsecas de los distintos complejos para dar la constante de la disociación de ese complejo; si son valores pequeños la cooperatividad es fuerte y predominan las formas E y ES4

58
Ecuación de Hill La ecuación de Hill se puede escribir a partir de la función de saturación Y Y = (S) h / K + (S) Esta ecuación corresponde a un sistema que presenta gran cooperatividad, es decir si el S se une a la proteína inmediatamente se unirán los siguientes y el equilibrio se puede considerar entre E y Esn y K= (E) (S) n/(ES) O también expresando en función a la velocidad
 (S) n/(ES) O también expresando en función a la velocidad.")
59
Si la cooperatividad es muy fuerte y los coeficientes de interacción son muy pequeños la ecuación queda reducida a v/Vmax = [S] h / K' + [S]h Ecuación de Hill Donde K´ = a3 b2c Ks4 para un tretrámero, es decir es una constante que relaciona la Ks con los factores de interacción h es un coeficiente que se determina experimentalmente,que no tiene un significado físico, es>1 si hay cooperatividad positiva y<1 si la cooperatividad es -

60
La ecuación de Hill se puede linealizar , escribiendola en la forma
V / Vmax – v = [S]h /K´ Llevando a logaritmos Log v/ Vmax-v = hlog [S] - log K´ La pendiente es el valor de h, coeficiente de Hill cuyo valor está relacionado con el número de sitios de unión

61
Ecuación de Adair Adair propuso una descripción más real de la unión de un ligando a una proteína, para un tetrámero consideró 4 equilibrios y cuatro constantes de disociación. Considerando una Enzima con 4 sitios de unión E'+ S = ES K1 = (ES)/(E)(S) ES + S = ES2 K2 = (ES2) / (ES) (S) ES2 + S = ES3 K3 = (ES3) /(ES2)(S) ES3 + S = ES4 K4= (ES4)/ (ES3) (S) K1,K2,K3, y K4 son las macroscópicas de disociación, que pueden relacionarse con las constantes microscópicas o intrínsecas
/(E)(S) ES + S = ES2 K2 = (ES2) / (ES) (S) ES2 + S = ES3 K3 = (ES3) /(ES2)(S) ES3 + S = ES4 K4= (ES4)/ (ES3) (S) K1,K2,K3, y K4 son las macroscópicas de disociación, que pueden relacionarse con las constantes microscópicas o intrínsecas.")
62
Fracción de saturación
La fracción de saturación de sitios activos puede expresarse en función de la concentración del ligando libre,(S) y, en el caso de una enzima con 4 sitios activos, se llega a (S)/K1+2(S)2/K1K2+ 3(S)3/K1K2K3+4(S)4/K1K2K3K4 Y= 4[1+(S)/K1+(S)2/K1K2+(S)3/K1K2K3+(S)4/K1K2K3K4 ] La relación entre las constantes de disociación y las constantes intrínsecas K'1K'2K'3 y K'4 son: K1 =1/4K K2= 2/3K'2 K3 = 3/2 K'3 K4= 4K'4 Si K'1>K'2>K'3>K'4 hay cooperatividad positiva K'4>K'3>K'2>K'1 hay cooperatividad negativa
 y, en el caso de una enzima con 4 sitios activos, se llega a. (S)/K1+2(S)2/K1K2+ 3(S)3/K1K2K3+4(S)4/K1K2K3K4. Y= [1+(S)/K1+(S)2/K1K2+(S)3/K1K2K3+(S)4/K1K2K3K4 ] La relación entre las constantes de disociación y las constantes intrínsecas K 1K 2K 3 y K 4 son: K1 =1/4K K2= 2/3K 2 K3 = 3/2 K 3 K4= 4K 4. Si K 1>K 2>K 3>K 4 hay cooperatividad positiva K 4>K 3>K 2>K 1 hay cooperatividad negativa.")
63
Constantes macroscópicas y microscópicas
Las constantes macroscópicas Ki se refieren a la disociación de un complejo ESi sin tener en cuenta la posición del sustrato S + ESi → ES i+1 Las constantes intrínsecas o microscópicas se refieren a la disociación de un complejo definido es decir teniendo en cuenta la posición del Por ejemplo si consideramos la concentración de todos los compejos que tienen 2 moléculas de sustrato[ ES2] y la concentración de cada uno de los posibles complejos con distinta posición de los 2 sustratos [ ES2] total = 6[ ES2] para cada posición

64
Relaciones entre K y K' En un tetrámero
K1 = ¼ K' K2 = 2/3 K2 K3 = 3/2K'3 y K4 = 4K' 4 Y así la ecuación anterior,considerando una enzima, se transforma en: v/Vm= S/K'1 + 3S2/K'1K'2+ 3S3/K'1K'2K'3+S4/K'1K'2K'3K'4 1 + 4S/K'1+6S2/K'1K'2+4S3/K'1K'2K'3+S4/K'1K'2K'3K'4 Donde Vm = 4 kcat. Cuando la cooperatividad es muy alta el termino de S4/ es mucho mayor que los otros y la ecuación puede simplificarse a v/Vm = S4 / K' + S4 ecuación de Hill

65
Factores o coeficientes de interacción.
En muchos casos la ecuación anterior se expresa en función a los factores de interacción a,b,c etc, siendo a= K'2/K' b = K'3/K'2 K'3=abK'1 c= K'4/cK'3 K'4= abcK'1 y la ecuación anterior queda S/Ks + 3S2/aKs2 + 3 S3/a2bKs3 + S4/a3b2cKs4 v/Vmax= 1+4 S/Ks + 6S2/aKs2 + 4 S3/a2bKs3 + S4/a3b2cKs4

66
Modelo de Monod-Wyman-Changeaux (modelo concertado)
Se propuso este modelo para explicar el comportamiento de las enzimas alostéricas En el oligómero hay al menos un eje de simetría La conformación de una unidad depende de la interacción con otra unidad El oligómero puede existir en 2 estados conformacionales R (relajado) y T (tenso), que presentan distinta afinidad para un ligando Ambas formas existen en un equilibrio R / T El sustrato se une a la forma R del protómero La forma R es catalíticamente activa y la forma T inactiva
 y T (tenso), que presentan distinta afinidad para un ligando. Ambas formas existen en un equilibrio R / T. El sustrato se une a la forma R del protómero. La forma R es catalíticamente activa y la forma T inactiva.")
67
Los efectores o moduladores se unen preferentemente a una de las formas T o R, por ej, si un efector se une a la forma T el equilibrio se desplaza de R hacia T, disminuye la cantidad de R y se forma menos ER Si el efector (activador) se une a la forma R, el equilibrio se desplaza de T a R y aumen ta la posibilidad de formrse más complejos ES Los valores de KR KT y L son los mismos para todos los complejos que se pueden formar ESn Se emplean 3 parámetro L, α y c L = [To]/[Ro] α = (S)/KR c= KR/KT [E]total = [Ro] + [To] + [RS] [RS2] considerando una enzima con dos sitios de unión V= kcat [RS] + 2 kcat [RS2] v/Vm = [S]/ KR ( 1 + [S]/ KR))n-1 / L +(1 + [S]/ KR ) n
 se une a la forma R, el equilibrio se desplaza de T a R y aumen ta la posibilidad de formrse más complejos ES. Los valores de KR KT y L son los mismos para todos los complejos que se pueden formar ESn. Se emplean 3 parámetro L, α y c. L = [To]/[Ro] α = (S)/KR c= KR/KT. [E]total = [Ro] + [To] + [RS] + [RS2] considerando una enzima con dos sitios de unión. V= kcat [RS] + 2 kcat [RS2] v/Vm = [S]/ KR ( 1 + [S]/ KR))n-1 / L +(1 + [S]/ KR ) n.")
68
Ecuaciones Se suelen evaluar dos cantidades Y y R
La fracción de saturación es: Lcα(1+cα)n-1 + α (1+α) n- Y = α=[S] libre / KR L (1+cα)n + (1+α)n (1 + α) n R = L(1+cα) n + ( 1+ α) n La cooperatividad es mayor cuanto mayor es L y menor es c
n-1 + α (1+α) n- Y = α=[S] libre / KR. L (1+cα)n + (1+α)n. (1 + α) n. R = L(1+cα) n + ( 1+ α) n. La cooperatividad es mayor cuanto mayor es L y menor es c.")
69
Enzimas K y V De acuerdo a este modelo la cooperatividad en la unión de ligandos se debe a que la proteina esta preferentemente en uno de los estados (por ej T) y el ligando se une a la forma R, al unirse y formar RS el equilibrio se desplaza de T a R, Al aplicar el modelo a las enzimas es más conveniente utilizar Vmax y Km Las dos formas de la enzima tendrán distintas afinidades por las formas R y T (Enzimas K)o distinta actividad catalítica (Enzimas V)
 y el ligando se une a la forma R, al unirse y formar RS el equilibrio se desplaza de T a R, Al aplicar el modelo a las enzimas es más conveniente utilizar Vmax y Km. Las dos formas de la enzima tendrán distintas afinidades por las formas R y T (Enzimas K)o distinta actividad catalítica (Enzimas V)")
70
Efectores o moduladores de la teoría M-W-Ch
Los efectores o moduladores de las enzimas alostéricas actúan sobre el equilibrio R↔T En las enzimas K,la unión del efector modifica la afinidad de la enzima por el sustrato variando Km Ej aspartato cabamoiltransferasa activada por ATP e inhibida por CTP En las enzimas V , el sustrato tiene la misma afinidad por las dos formas sin embargo ambas difieren en su actividad catalítica, el efector desplaza el equilibrio R↔T , variando Vm Ej. fosforilasa b, activada por AMP

71
Modelo de Koshland-Nemethy-Filmer
En ausencia de ligandos la enzima está en una forma conformacional determinada Al unirse un sustrato experimenta un cambio Los cambios conformacionales son secuenciales, las formas van cambiando a medida que los ligandos se van uniendo Las interacciones entre las distintas subunidades pueden ser positivas o negativas La expresión para la fracción de saturación se puede obtener a partir de los equilibrios entre las distintas formas, y sus constantes de disociación

72
Esquema para un tetrámero

73
Cada constante de disociación se considera que tiene 3 componentes
- Una constante de disociación para la unión de la subunidad más favorecida - una constante de equilibrio para el cambio de conformacion □↔○ - constantes de interacción que depende del grado de estabilidad de los complejos entre las subunidades en relación a la estabilidad del complejo solo Las ecuaciones derivadas son más complejas que las correspondientes al modelo MWCh

74
Comparación de ambos modelos
Ambos modelos el concertado de MWCh y el secuencial de KNF puede ser considerados como los extremos de un modelo más general que considera todas las conformaciones posibles y de estados de unión al ligando ○○↔ ○□ ↔□□ ■ proteína unida al ligando ↕ ↕ ↕ Este modelo da lugar a ○●↔ ○■ ↔□■ ecuaciones extraordinariamente ↕ ↕ complejas ●●↔ ●■ ↔ ■■ ■

75
En algunos casos la enzima se comporta de acuerdo a uno de los dos modelos y en otros no
Se pueden establecer 3 diferencias entre ambos modelos -el modelo concertado no explica la cooperatividad negativa -La observación de cambios conformacionales se pueden monitorear en algunos casos y determinar R e Y la gráfica de R vs Y es lineal en el modelo secuencial y curva en el concertado -demostración de reacciones de idomerización

76
Las interacciones alostéricas permiten un alto grado de regularización del metabolismo
Métodos para estudiar enzimas reguladoras Tipo de información método de estudio En los estudios cinéticos Se realizan estudios de actividad se observa cooperaatividad Se observan las gráficas si hay o no linealidad Se observa cooperatividad Se emplean métodos como equilibrio en los estudios de ligando? de diálisis, de ultracentrifugación Las moléculas reguladoras Prueba sobre efectos de los se unen a sitios diferentes alS? Moduladores en las curvas cinéticas Consiste la enzima en varias El número de subunidades puede subunidades? Determinarse por M, simetría, y otros

77
Mecanismos de enzimas con dos sustratos

78
Enzimas que actúan en reacciones con dos sustratos diferentes
Muchas enzimas trabajan con dos sustratos diferentes, las deshidrogenasas por ejemplo se unen al sustrato que se oxida y al NAD+ aceptor de los protones y electrones En la mayoría de los casos, se puede estudiar este tipo de reacciones teniendo en cuenta la cinética de un sustrato, cuando se mantiene constante la concentración de uno de los sustratos y se varía la concentración del otro

79
Se distinguen los siguientes tipos de mecanismos
Cuando se distingue un compuesto ternario Mecanismo secuencial ordenado Mecanismo secuencial aleatorio Cuando no se distingue un compuesto ternario Mecanismo ping-pong Mecanismo Theorell-Chance

80
Mecanismo secuencial ordenado
Los sustratos se unen a la enzima en un orden determinado, y los productos también siguiendo cierto orden A B P Q ↓ ↓ ↑ ↑ E EA EAB → EPQ EQ E Este tipo de mecanismo se observa en las deshidrogenasas que actúan con NAD*, donde la coenzima se fija en primer lugar y luego el sustrato que se oxida. La unión del dinucleótido produce una modificación de la enzima favoreciendo la unión del sustrato Así también se libera primero el producto y luego el NADH

81
Mecanismo aleatorio o al azar
Los sustratos pueden unirse en cualquier orden y la liberación de los productos también se hace en forma aleatoria A → EA → B P \/ → Q E EAB →EPQ B → EB → A Q → P

82
Mecanismo de Theorell- Chance
Es un caso de mecanismo ordenado en que el complejo ternario no se acumula,y no se observa en las condiciones en que se realiza la reacción Este tipo de mecanismo se observa en la alcohol deshidrogenasa de hígado de caballo A B P E ↓ EA ↓ EQ --↑----E Q Se libera primeramente acetaldehido y luego el NADH

83
Mecanismo ping-pong o de doble desplazamiento
En este mecanismo no se forma compuesto ternario La enzima se une primero a un sustrato, se forma el complejo binario y se libera un producto y la enzima queda modificada en forma covalente, esta forma se une luego al segundo sustrato, se forma el complejo correspondiente y se libera otro producto, la enzima vuelve a la forma inicial E + A → EA → E´ + P E´ + B → EB → E + Q Este mecanismo se observa en muchas enzimas, sobre todo en las que participan en la transferencia de grupos

84
Empleando el esquema de Cleland A P B Q ↓ _________↑ ↓ _ ↑
↓ _________↑ ↓ _ ↑ E EA E'P E'Q E'B EQ E Este tipo de mecanismo es muy comun en muchas enzimascomo transaminasas y flavoenzimas por ejemplo la aspartato transaminasa que pqrticipa en la reacción Glutamato+Epiridoxal –2oxoglutarato +Epiridoxamina Oxaloacetato + Epiridoxamina –aspartato + Epiridoxal

85
Las gráficas que se obtienen cuando se mantiene la concentración de un sustrato constante y se varía el otro,y se toma 1/v vs. 1/[S] son rectas paralelas entre sí Este tipo de mecanismo se observa también en varias enzimas de transferencia de fosfatos, como la fosfoglicerilmutasa Se produce primero una reacción donde un grupo fosforilo se transfiere a la enzima dando un derivado fosforilenzima, el grupo fosfato es transferido luego a un segundo sustrato y la enzima se separa en su forma inicial
![Las gráficas que se obtienen cuando se mantiene la concentración de un sustrato constante y se varía el otro,y se toma 1/v vs. 1/[S] son rectas paralelas entre sí](http://slideplayer.es/slide/10362356/33/images/85/Las+gr%C3%A1ficas+que+se+obtienen+cuando+se+mantiene+la+concentraci%C3%B3n+de+un+sustrato+constante+y+se+var%C3%ADa+el+otro%2Cy+se+toma+1%2Fv+vs.+1%2F%5BS%5D+son+rectas+paralelas+entre+s%C3%AD.jpg "Este tipo de mecanismo se observa también en varias enzimas de transferencia de fosfatos, como la fosfoglicerilmutasa. Se produce primero una reacción donde un grupo fosforilo se transfiere a la enzima dando un derivado fosforilenzima, el grupo fosfato es transferido luego a un segundo sustrato y la enzima se separa en su forma inicial.")
86
En el mecanismo Ping-pong el grupo G es transferido dos veces del 1er sustrato a la enzima y de esta al 2do sustrato, se denomina por eso de doble desplazamiento Los mecanismos con formación de compuesto ternaario donde el grupo G se transfierre una sola vez se denomina de desplazamiento simple Esta división fue introducida por Koshland y se refiere más a aspectos químicos que cinéticos

87
Ecuaciones en cinética de dos sustratos
Para obtener las ecuaciones de velocidad en la cinética de dos sustratos se puede aplicar el método de King y Altman,considerando diversos mecanismos posibles o no, sin embargo esto da lugar a múltiples ecuaciones y es más útil emplear otros caminos Por ej para el mecanismo ordenado la ecuación hallada contiene 13 coeficientes, cuando solo puede haber 8 constantes de velocidad

88
Recomendaciones de la IUBMB
Las constantes consideradas pueden reducirse a 3 tipos: Velocidades limitantes (Vm) Constates de Michaelis (Km) Constantes de inhibición (Ki) Las Km respecto a A o B corresponde a la Km en la cinética de un sustrato Las Ki respecto a A o B corresponde a Ki cuando A (oB) se consideran como producto inhibidor de la reación inversa Si se conoce la (E) puede calcularse las constantes catalíticas y calcularse las constantes de especificidad
 Constates de Michaelis (Km) Constantes de inhibición (Ki) Las Km respecto a A o B corresponde a la Km en la cinética de un sustrato. Las Ki respecto a A o B corresponde a Ki cuando A (oB) se consideran como producto inhibidor de la reación inversa. Si se conoce la (E) puede calcularse las constantes catalíticas y calcularse las constantes de especificidad.")
89
Ecuaciones Para el mecanismo al azar, suponiendo el equilibrio rápido se puede llegar a V+ (A)(B) V-(A) (B) v= KiA KmB KiQ KmP 1+(A)/KiA+(B)/KiB+(P)/KiP+(Q)/KiQ+(A)(B)/KiAKMQ+(P)(Q)//KipKmq Sin embargo si se toman las velocidades iniciales donde las concentraciones iniciales de productos son despreciables y multipliando por KiAKmB la ecuación se reduce a V(A)(B) v= KiAKmB + KmB (A) + KmA(B) + (A)(B)
/KiA+(B)/KiB+(P)/KiP+(Q)/KiQ+(A)(B)/KiAKMQ+(P)(Q)//KipKmq. Sin embargo si se toman las velocidades iniciales donde las concentraciones iniciales de productos son despreciables y multipliando por KiAKmB la ecuación se reduce a. V(A)(B) v= KiAKmB + KmB (A) + KmA(B) + (A)(B)")
90
Vm o V es la velocidad límite cuando la enzima está saturada con ambos sustratos
Si la concentración de uno de los sustratos es mucho mayor que la del otro , por ej B, la ecuación se reduce a v = V(A)/ KmA +(A) Asi se puede definir KmA como el valor de la constante límite de Michaelis para A cuando B está en una concentración saturante De igual modo KmB es el valor de Km para B cuando A está en concentración saturante
/ KmA +(A) Asi se puede definir KmA como el valor de la constante límite de Michaelis para A cuando B está en una concentración saturante. De igual modo KmB es el valor de Km para B cuando A está en concentración saturante.")
91
Parámetros aparentes Para estudiar la cinética de las reacciones con dos sustratos se debe repetir varias veces la experiencia, manteniendo en cada la concentración de un sustrato constante, por ej B y variando el otro La ecuación tomará la forma v= V(B)/KmB(B) (A) / KiAKmB+ KmB(B)/KmB+(B) donde será constantes V(B)/ KmB(B) = V ap y KiAKmB +KmA(B)/ KmB + (B) = Kmap Los parámetros Vmap, Kmap y Vmap/Kmap son función de la concentración de B
/KmB(B) (A) / KiAKmB+ KmB(B)/KmB+(B) donde será constantes V(B)/ KmB(B) = V ap. y KiAKmB +KmA(B)/ KmB + (B) = Kmap. Los parámetros Vmap, Kmap y Vmap/Kmap son función de la concentración de B.")
92
Gráficas primarias y secundarias en los mecanismo con complejo ternario
Los datos obtenidos experimentalmente se pueden llevar a cualquiera de los gráficos lineales L-B H-W o E- H,tales gráficas se denominan primarias En la representación de L-B se obtienen rectan que se interceptan en el 2do cuadrante en un punto cuya abcisa es -1/KiA y la ordenada 1/V(1-KmA/KiA) Esta gráfica da directamente el valor de KiA, los otros parámetros se obtienen de gráficas secundarias Vap vs o Vap/Kmap vs (B) corresponden a hipérbolas y la grafica de dobles inversan dan rectas Representando 1/Vap vs 1/(B) se obtiene una recta cuya pendiente es KmB/V y en el eje de abcisas -1/KmB Kmap/(B) vs 1/(B) da como pendiente KiAKmB/V
 Esta gráfica da directamente el valor de KiA, los otros parámetros se obtienen de gráficas secundarias. Vap vs o Vap/Kmap vs (B) corresponden a hipérbolas y la grafica de dobles inversan dan rectas. Representando 1/Vap vs 1/(B) se obtiene una recta cuya pendiente es KmB/V y en el eje de abcisas -1/KmB. Kmap/(B) vs 1/(B) da como pendiente KiAKmB/V.")
93
Gráficas en el mecanismo pingpong
En ausencia de productos la velocidad inicial es v= V (A) (B) / KmB (A) + KmA (B) + (A)(B) Si se toman valores constantes de B y se varía A se tiene Vap = V(B) / KmB + (B) Kap = KmA (B) / KmB + (B) y Vap/Kmap = V/ KmA, valor que no depende de (B) Lás gráficas de 1/v vs 1/ (A) son rectas paralelas entre sí Las gráficas secundarias 1/Vap vs (B), tiene una pendiente igual a KmB/V y la intercepción en ordenadas es 1/V Kmap vs (B) tiene pendiente 0
 (B) / KmB (A) + KmA (B) + (A)(B) Si se toman valores constantes de B y se varía A se tiene. Vap = V(B) / KmB + (B) Kap = KmA (B) / KmB + (B) y Vap/Kmap = V/ KmA, valor que no depende de (B) Lás gráficas de 1/v vs 1/ (A) son rectas paralelas entre sí. Las gráficas secundarias 1/Vap vs (B), tiene una pendiente igual a KmB/V y la intercepción en ordenadas es 1/V. Kmap vs (B) tiene pendiente 0.")
94
Retroinhibición o inhibición feed-back
En muchas vías metabólicas importantes se observan casos de la llamada inhibición feedback o retroinhibición, cuando un producto final de una vía metabólica se encuentra en exceso , actúa como inhibidor de una enzima que participa en una reacción al comienzo de la vía, disminuyendo así la velocidad de la reacción dada y por lo tanto disminuyendo también el producto considerado A → B → C → D E E E3 El exceso de D actúa como inhibidor de la enzima E1

95
En muchos casos una vía metabólica se difurca, y a partir de un metabolito se pueden formar dos o más productos por caminos diferente

96
En el esquema anterior los productos G y J pueden inhibir a la primera enzima de la vía E1 o actuar solamente sobre la enzima donde se inicia la bifurcación correspondiente E4 o E7 respectivamente En el primer caso disminuiran todos los metabolitos desde B en adelante, en el segundo caso, disminuira la vía correspondiente, si afecta a E4, E,F y G, o si afecta a E7 , H I y J

97
Inhibición por 2 inhibidores
Se suelen distinguir los distintos tipos de inhibición Competitiva Cooperativa Concertada Aditiva Acumulativa Secuencial

98
Inhibición cooperativa
Dos inhibidores diferentes X e I se unen a la enzima a distintos sitios de unión La unión de cualquiera de los inhibidores X o I,puede actuar solo, impidiendo la unión del sustrato X solo o I solo, tienen efecto inhibitorio, pero la acción conjunta tienen un efecto mayor que la que tendría X solo o I solo auna concentración equivalente a la concentración total de ambos Se pueden formar EXI, EXIS A nivel de saturación cualquiera de los inhibidores puede hacer que la velocidad llegue a cero Ejemplo Fosforibosilaminasintetasa

99
Esquema de retroinhibición
F→G →H →I E1 E2 E3 E4 E9 S →B →C →D →N E5 J →K → L →X E6 E E8 E E E12

100
Inhibición concertada
La inhibición se produce solamente con la acción simultánea de los dos inhibidores Ni X ni I tienen ningún efecto solos (por ejemplo EX yEI tienen la misma afinidad por el sustrato pero EXS y EIS son catalíticamente activos, pero EXI no puede unirse al sustrato y por los tanto no forma producto La unión de X o I a la enzima aumenta la afinidad por el otro inhibidor Ejemplo: algunas aspartil quinasa

101
Imhibición acumulativa
Cada producto final es un inhibidor parcial., I solo ni X solo pueden hace que la velocidad de la reacción se anule. Esto implica que EX e Ei se unen a S pero no tan bien como S, o que EXS o EIS no son tan activos como ES Ejemplo: gñutamina sintetasa

102
Inhibición aditiva Hay dos enzimas diferentes que son inhibidas, cada una sensible a un retroinhibidor o dos sitios catalíticos en la enzima de modo que ambos pueden ser ocupados simultáneamente por ambos inhibidores Ejemplos_ algunas aspartilquinasas

103
Inhibición secuencial
I inhibe a una enzima de la ruta metabólica y X inhibe a otra enzima que actúa más adelante Cuando ambos inhibidores X e I tienen concentraciones altas, los dos procesos se inhiben y aumenta la cantidad de un metabolito N puede inhibir a otra enzima

104
Enzimas interconvertibles
El control de la actividad enzimática se puede realizar por medio de modificaciones covalentes reversibles que permiten que la enzima pase de un estado más activo a otro menos activo, o vicecersa, de acuerdo a las necesidades metabolicas. Las enzimas que pueden experimentar este tipo de transformación se llaman enzimas interconvertibles Las modificaciones covalentes se realizan por acción de otras enzimas que se denominan enzimas convertidoras

105
Fosforilación Uno de los ejemplos más estudiados es el de la fosforilasa, que catalisa la fosforilación del glicógeno Esta enzima puede existir en dos formas: ATP ADP Fosforilasa b Fosforilasa a E-CH2- OH E-CH2-O-PO3 PO H2O La reacción de b a a requiere ATP y Mg, esta catalizada por la fosforilasa quinasa La reacción de a hacia b, libera fosfato y está catalizada por la fosforilasa fosfatasa

106
Regulación por fosforilación
Existen numerosas enzimas y proteínas cuya actividad está regulada por mecanismos de fosforilación-desfosforilación. La forma fosforilada puede se la más activa o no , dependiendo de la proteína Se han observado otros mecanismos de regulación covalente en proteínas, la adenilación,ribosilación, metilación, acetilación, miristilación, pero en enzimas además de las fosforilación que es el mecanismo más común se conocen ejemplos de adenilación ( en glutamino sintetasa de E.coli) y ribosilación en varias enzimas ( nitrogenasa, RNA polimerasa y otras)
 y ribosilación en varias enzimas ( nitrogenasa, RNA polimerasa y otras)")
107
La modificación covalente se produce en residuos de serina, treonina , tirosina y otros
Las enzimas que participan en la fosforilación son las proteína quinasas, y las que catalizan las desfosforilaciones son las proteínas fosfatasas La adenilación de la glutamino sintasa se produce en un resto de tirosina, y la forma adenilada muestra poca actividad La nitrogenasa de Rodospirillum rubrum es regulada por la ribosilación de una arginina, y la forma ribosilada es menos activa

108
Características de la regulación covalente
La conversión de una forma de la enzima a la otra está regulada por la acción de otra enzima, lo que produce un cambio rápido en la cantidad de enzima activa presente y una gran amplificación de una señal inicial La modificación reversible permite una respuesta mucho mejor controlada a los diferentes requerimientos del metabolismo Las enzimas convertidoras a su vez generalmente responden a una señal hormonal Todo este proceso permite una cascada de amplificación como respuesta a las necesidades metabólicas

Presentaciones similares














![1 KM 1 1 Vo= Vmax [S] + Vmax Y = m x + b = m = b.](/2/161459/big_thumb.jpg "1 KM 1 1 Vo= Vmax [S] + Vmax Y = m x + b = m = b.>")





