Descargar la presentación
La descarga está en progreso. Por favor, espere

1
TEMA 8 LA MEMBRANA PLASMÁTICA Y LA PARED CELULAR
TEMA 8 LA MEMBRANA PLASMÁTICA Y LA PARED CELULAR. CITOSOL, CITOESQUELETO. SISTEMAS DE MEMBRANA Y ORGÁNULOS. MOTILIDAD CELULAR.

2
1- LA MEMBRANA PLASMÁTICA Y LA PARED CELULAR.
La membrana plasmática es la envoltura (constituida por un complejo molecular) que rodea y limita a las células exteriormente, pero no las aísla del medio que las rodea, pues permite el intercambio de materia y energía. Con el empleo del microscopio electrónico en 1959 Robertson observó que presentaba una estructura trilaminar de unos 75 Å, formada por capas de diferente densidad. Modelo del mosaico fluido de membrana Fue propuesto por Singer Nicolson en Según este modelo las membranas poseen lípidos, proteínas y oligosacáridos que se disponen en una configuración de baja energía libre. Este modelo sostiene que: a) Los lípidos y las proteínas integrales que forman la membrana forman un mosaico molecular. b) Los lípidos y las proteínas pueden desplazarse en el plano de la bicapa. Las membranas son fluidas. c) Las membranas son asimétricas en cuanto a la disposición de sus componentes moleculares.
 que rodea y limita a las células exteriormente, pero no las aísla del medio que las rodea, pues permite el intercambio de materia y energía. Con el empleo del microscopio electrónico en 1959 Robertson observó que presentaba una estructura trilaminar de unos 75 Å, formada por capas de diferente densidad. Modelo del mosaico fluido de membrana. Fue propuesto por Singer Nicolson en Según este modelo las membranas poseen lípidos, proteínas y oligosacáridos que se disponen en una configuración de baja energía libre. Este modelo sostiene que: a) Los lípidos y las proteínas integrales que forman la membrana forman un mosaico molecular. b) Los lípidos y las proteínas pueden desplazarse en el plano de la bicapa. Las membranas son fluidas. c) Las membranas son asimétricas en cuanto a la disposición de sus componentes moleculares.")
3
Modelos de membrana plasmática

4
· Los lípidos forman una doble capa, con los grupos hidrófobos en el centro y los hidrófilos en el exterior (en contacto con la fase acuosa) formando la matriz de la membrana. Las membranas funcionales requieren una matriz lipídica fluida. En las membranas los lípidos pueden hacer movimientos de difusión lateral, rotación y de flexión, y además, es su composición lipídica la que les confiere capacidad de auto-ensamblaje, autosellado, fluidez e impermeabilidad.

5
· Las proteínas son anfipáticas, lo que le permite estar en parte embebidas y en parte sobresalientes de la bicapa lipídica. En función de su mayor o menor afinidad por el agua, se asocian con los lípidos de membrana de diversas formas. Las proteínas pueden clasificarse en: a) Proteínas integrales o intrínsecas (2), sí están muy unidas a los lípidos y sólo pueden extraerse por métodos muy drásticos. b) Proteínas periféricas o extrínsecas (3), si se asocian a la superficie de la bicapa por interacciones no covalentes, estas últimas se encuentran sobre todo en la cara interna de la membrana y suelen corresponder a enzimas. Se ha comprobado que las proteínas se desplazan por la bicapa lipídica y que pueden estar asociadas. Las proteínas también pueden trasladarse por la membrana y hacer los movimientos denominados «flip-flop», esto es, cruzar la membrana de arriba abajo. Según su mayor o menor afinidad por la bicapa lipídica pueden encontrarse: atravesando la membrana lipídica (1) (proteína transmembrana), atravesando sólo parcialmente la membrana, exponiendo uno de sus extremos al medio acuoso, unidas por enlaces no covalentes a otras proteínas de membrana (4) o asociadas covalentemente a cadenas de ácidos grasos.
 Proteínas integrales o intrínsecas (2), sí están muy unidas a los lípidos y sólo pueden extraerse por métodos muy drásticos. b) Proteínas periféricas o extrínsecas (3), si se asocian a la superficie de la bicapa por interacciones no covalentes, estas últimas se encuentran sobre todo en la cara interna de la membrana y suelen corresponder a enzimas. Se ha comprobado que las proteínas se desplazan por la bicapa lipídica y que pueden estar asociadas. Las proteínas también pueden trasladarse por la membrana y hacer los movimientos denominados «flip-flop», esto es, cruzar la membrana de arriba abajo. Según su mayor o menor afinidad por la bicapa lipídica pueden encontrarse: atravesando la membrana lipídica (1) (proteína transmembrana), atravesando sólo parcialmente la membrana, exponiendo uno de sus extremos al medio acuoso, unidas por enlaces no covalentes a otras proteínas de membrana (4) o asociadas covalentemente a cadenas de ácidos grasos.")
6
· Los oligosacáridos (y en menor cantidad lipoproteínas) forman el llamado glicocálix y están asociados con la bicapa. La mayor parte están unidos de forma covalente a los lípidos (glucolípidos) (se estudian en el tema 23) o a las proteínas (glucoproteínas) (se estudian en el tema 24). Se encuentran casi exclusivamente en la cara externa de la membrana contribuyendo a su asimetría. La estructura de este glicocálix es muy variable. En la mayoría de las células forma una delicada capa y suele estar muy desarrollado en células epiteliales.
 (se estudian en el tema 23) o a las proteínas (glucoproteínas) (se estudian en el tema 24). Se encuentran casi exclusivamente en la cara externa de la membrana contribuyendo a su asimetría. La estructura de este glicocálix es muy variable. En la mayoría de las células forma una delicada capa y suele estar muy desarrollado en células epiteliales..")
7
Funciones de la mb. plasmática - Límite físico de la célula - Transporte de sustancias a su través = permeabilidad selectiva A- MOLÉCULAS PEQUENAS A.1- TRANSPORTE PASIVO a) ÓSMOSIS b) DIFUSIÓN SIMPLE: PROTEINAS CANALES c) DIFUSIÓN FACILITADA: PERMEASAS A.2- TRANSPORTE ACTIVO a) BOMBA SODIO-POTASIO b) CANAL DE FUGA DE POTASIO B- MOLÉCULAS GRANDES B.1- ENDOCITOSIS a) PINOCITOSIS b) FAGOCITOSIS B.2- EXOCITOSIS - Mantenimiento del potencial de membrana - Receptora y transmisora de señales = comunicación intercelular, adhesión y unión celular, unión matriz-célula - Soporte de reacciones metabólicas (cadena respiratoria)
 ÓSMOSIS b) DIFUSIÓN SIMPLE: PROTEINAS CANALES c) DIFUSIÓN FACILITADA: PERMEASAS A.2- TRANSPORTE ACTIVO a) BOMBA SODIO-POTASIO b) CANAL DE FUGA DE POTASIO B- MOLÉCULAS GRANDES B.1- ENDOCITOSIS a) PINOCITOSIS b) FAGOCITOSIS B.2- EXOCITOSIS - Mantenimiento del potencial de membrana - Receptora y transmisora de señales = comunicación intercelular, adhesión y unión celular, unión matriz-célula - Soporte de reacciones metabólicas (cadena respiratoria).")
8
A.1- TRANSPORTE PASIVO A) ÓSMOSIS
Ósmosis: cuando se ponen en contacto dos soluciones de distinta concentración separadas por una membrana semipermeable (permite únicamente el paso de moléculas de agua y no de soluto), la tendencia a igualar las concentraciones (hacerlas isotónicas) se manifiesta por el paso de agua desde la más diluida (hipotónica) a la más concentrada (hipertónica). La "presión mecánica" necesaria para contrarrestar el paso de agua se denomina "presión osmótica" y el proceso "ósmosis". La plasmolisis es el proceso de pérdida de agua en células vegetales, teniendo como resultado la contracción del citoplasma alejándose de las paredes celulares. (Se aplica indebidamente a otros tipos de células).
, la tendencia a igualar las concentraciones (hacerlas isotónicas) se manifiesta por el paso de agua desde la más diluida (hipotónica) a la más concentrada (hipertónica). La presión mecánica necesaria para contrarrestar el paso de agua se denomina presión osmótica y el proceso ósmosis . La plasmolisis es el proceso de pérdida de agua en células vegetales, teniendo como resultado la contracción del citoplasma alejándose de las paredes celulares. (Se aplica indebidamente a otros tipos de células).")
9
B) DIFUSIÓN SIMPLE Hay solutos que atraviesan la membrana a través de la bicapa lipídica, como por ejemplo gases (como el O y el N), muchas sustancias lipófilas (como algunos fármacos y anestésicos, disolventes, insecticidas, etc.), y moléculas polares, pequeñas y sin carga (agua, urea, etanol, CO2). Se puede realizar también difusión simple a través de canales o proteínas de canal. Intervienen aquí proteínas transmembrana que delimitan en su interior un orificio o canal. Pueden estar siempre abiertas aunque algunas actúan de puerta transitoria y poseen regulados su cierre y su apertura. Hay canales regulados por ligando y regulados por voltaje. Otros canales se abren cuando cambia la concentración iónica de algún ión (estos canales intervienen en la sinapsis, se estudia en el tema 56). Esquema de dos tipos de canales que participan en la difusión simple. A) Canal regulado por ligando: al unirse éste a la proteína se produce la apertura del canal; B) Canal regulado por voltaje: el canal se abre dependiendo de la polaridad de la membrana.
, muchas sustancias lipófilas (como algunos fármacos y anestésicos, disolventes, insecticidas, etc.), y moléculas polares, pequeñas y sin carga (agua, urea, etanol, CO2). Se puede realizar también difusión simple a través de canales o proteínas de canal. Intervienen aquí proteínas transmembrana que delimitan en su interior un orificio o canal. Pueden estar siempre abiertas aunque algunas actúan de puerta transitoria y poseen regulados su cierre y su apertura. Hay canales regulados por ligando y regulados por voltaje. Otros canales se abren cuando cambia la concentración iónica de algún ión (estos canales intervienen en la sinapsis, se estudia en el tema 56). Esquema de dos tipos de canales que participan en la difusión simple. A) Canal regulado por ligando: al unirse éste a la proteína se produce la apertura del canal; B) Canal regulado por voltaje: el canal se abre dependiendo de la polaridad de la membrana.")
10
C) DIFUSIÓN FACILITADA PERMEASA
Difusión facilitada. Se realiza mediante proteínas transportadoras llamadas permeasas, que funcionan mediante un cambio de configuración, o mediante ionóforos, que son pequeñas moléculas no polares que se disuelven en la bicapa lipídica y aumentan su permeabilidad iónica. Actúan protegiendo la carga del ión que transportan, y pueden ser móviles y formadoras de canales. Hay diferencias en el comportamiento de los transportadores. En el caso de la difusión simple la velocidad del transporte es directamente proporcional a la concentración del soluto. En el caso de la difusión facilitada la velocidad sigue una cinética michaeliana, alcanzándose la velocidad máxima cuando el transportador está saturado por el sustrato.

11
A.2- TRANSPORTE ACTIVO A) BOMBA Na - K
Gracias a este tipo de transporte se consigue que las concentraciones extra e intracelulares de diversos iones sean diferentes. Es en contra de gradiente electroquímico y consume energía (se obtiene del desdoblamiento del ATP en ADP y Pi). Un ejemplo es la ATPasa Na+/ K+, o Bomba Na+/ K+, que bombea Na+ hacia el exterior de la célula y K+ hacia el interior; de este modo se consigue que la concentración de Na+ sea mayor en el exterior y ocurre lo contrario con la de K+. Por cada molécula de ATP hidrolizado se transportan 3 Na+ hacia el exterior y 2 K+ hacia el interior.
. Un ejemplo es la ATPasa Na+/ K+, o Bomba Na+/ K+, que bombea Na+ hacia el exterior de la célula y K+ hacia el interior; de este modo se consigue que la concentración de Na+ sea mayor en el exterior y ocurre lo contrario con la de K+. Por cada molécula de ATP hidrolizado se transportan 3 Na+ hacia el exterior y 2 K+ hacia el interior.")
12
B) CANAL DE FUGA DE POTASIO
El canal de fuga del potasio permite la salida de potasio de la célula, a favor de gradiente de concentración, colaborando de este modo a que se acumulen en el exterior los iones con carga +, quedando el interior de la célula cargado negativamente. La salida de cationes K+ se ve frenada por la acumulación de cargas + en el exterior. Esto, unido a que la membrana plasmática es algo permeable al catión Na+, supondría, con el tiempo, la desaparición del potencial de membrana. Para evitarlo, la bomba de sodio-potasio mantiene la diferencia de concentración de estos iones a ambos lados de la membrana. Para que funcione este mecanismo, la célula consume la tercera parte de sus recursos energéticos. En las células nerviosas, en las que se restablecen continuamente los potenciales de membrana, este valor puede alcanzar el 70 % del total.

13
La endocitosis es un mecanismo de transporte de componentes del medio externo de la célula hacia el interior del citoplasma. En este proceso, una región de la membrana se invagina y engloba un pequeño volumen de material extracelular, formando una protuberancia que se desprende hacia el interior de la célula mediante la fisión de membranas. La pequeña vesícula resultante (endosoma) puede desplazarse hacia el interior de la célula, liberando su contenido en otro orgánulo rodeado por una membrana simple (corno por ejemplo un lisosoma) mediante fusión de las dos membranas. De este modo, el endosoma actúa como una extensión intracelular de la membrana plasmática y permite un contacto íntimo entre componentes del medio extracelular y regiones del interior del citoplasma a las que los componentes externos no podrían llegar mediante simple difusión.
 puede desplazarse hacia el interior de la célula, liberando su contenido en otro orgánulo rodeado por una membrana simple (corno por ejemplo un lisosoma) mediante fusión de las dos membranas. De este modo, el endosoma actúa como una extensión intracelular de la membrana plasmática y permite un contacto íntimo entre componentes del medio extracelular y regiones del interior del citoplasma a las que los componentes externos no podrían llegar mediante simple difusión..")
14
La endocitosis también puede incorporar macromoléculas específicas a la célula. En este caso, las macromoléculas se unen a receptores específicos (situados en invaginaciones de la membrana llamadas cavidades revestidas) de la superficie celular. Así es como captan, por ejemplo, el colesterol las células sanguíneas. Dentro de la endocitosis, se habla de fagocitosis si las partículas que entran a la célula son visibles al microscopio óptico, mientras que si se trata de líquido con sustancias disueltas se habla de pinocitosis. .

15
La exocitosis constituye el proceso inverso a la endocitosis, por el que una vesícula del citoplasma se desplaza hacia la superficie interior de la membrana plasmática y se fusiona con ella, liberando el contenido vesicular hacia el exterior de la membrana. Muchas proteínas destinadas a secreción externa se liberan mediante exocitosis después de su empaquetamiento en vesículas secretoras

16
Diferenciaciones de la membrana plasmática
Por diferenciación de la membrana se entiende una región que está especialmente adaptada para una función. Destacan: Microvellosidades, Son prolongaciones citoplasmáticas digitiformes en cuyo interior se localizan filamentos de proteínas y que se localizan en la zona apical de las células. Se encuentran muy desarrolladas en células que necesitan aumentar su superficie, por ejemplo, en las células intestinales. 2. Invaginaciones. El aumento de superficie celular se debe a la existencia de entrantes de la membrana plasmática. Se encuentran en las células de los túbulos renales, en la cara opuesta a la luz del tubo, y se suponen relacionados con la regulación osmótica. 3. Pliegues e interdigitaciones. A veces existen para reforzar la unión entre células. Son muy frecuentes en las células epiteliales. 4. Uniones intercelulares. 5. Otras diferenciaciones. En protozoos ciliados aparecen repliegues en la superficie celular o expansiones a modo de alvéolos. Muchos flagelados y ciliados poseen en el polo apical de algunas células estructuras que se extienden cuando e organismo se irrita, se conocen como tricocistos. Similares son los mucocistos, que expulsan sustancias que paralizan a las presas, o toxicistos, que poseen ciertos ciliados holotricos depredadores.

17
FUNCIÓN DE UNIÓN INTERCELULAR
Uniones intercelulares. Se localizan en las superficies laterales de las células manteniéndolas unidas y permitiendo el contacto entre ellas. Con respecto a la distancia entre las membranas de las células que se unen, se clasifican en: a) Occludens, íntima o estrecha si no existe ninguna distancia entre membranas. b) Adherens si esa distancia es de 20 a 25 nm. e) De acoplamiento, nexo o hendidura si la distancia intercelular es de 2 nm. En este tipo de uniones existen zonas en las que las membranas de ambas células están separadas,
 Occludens, íntima o estrecha si no existe ninguna distancia entre membranas. b) Adherens si esa distancia es de 20 a 25 nm. e) De acoplamiento, nexo o hendidura si la distancia intercelular es de 2 nm. En este tipo de uniones existen zonas en las que las membranas de ambas células están separadas,")
18
PROPIEDADES DE LA MEMBRANA.
FLUIDEZ: depende de lo lipidos y es importante pues algunos procesos de transporte y actividades enzimáticas depende de la viscosidad de la membrana. Hay bacterias, levaduras y otros microorganismos cuyas temperaturas varian con las de su entorno controlan la composición de los AG de su membrana para mantener su fluidez relativamente constante. Las células eucariotas mantienen su fluidez con ayuda del colesterol que da rigidez pero impide la cristalizacion de los AG ASIMETRIA: Se basa en la composición de los PL, así las moléculas con colina se encentran en la mitad exterior de la bicapa, mientras que las que presentan un grupo amino se hallan en la mitad interior. Los glucolípidos muestran esta propiedad claramente pues la parte lipidica siempre se halla hacia el exterior constituyendo el glucocalix

19
La pared celular posee varias capas que, de fuera hacia dentro de la célula, son:. En la mayoría de los tejidos vegetales la lamela o lámina media es compartida por las células que se encuentran adyacentes. Está constituida por pectatos (polímeros de ácido galacturónico) y proteínas. La pared primaria es más gruesa y al microscopio electrónico se observan las macrofibrillas formadas por haces de microfibrillas adosadas longitudinalmente. La pared secundaria sólo está presente en algunos tipos celulares y suele ser mucho más gruesa que la pared primaria. Además de celulosa suele contener otras sustancias como hemicelulosa, proteínas, pectatos, lignina, cutina, suberina, sales minerales, etc, que, en general, tienden a darle mayor consistencia. Comprende tres subcapas que se denominan S1, S2 y S3 (de fuera hacia dentro). En la S1 las microfibrillas se disponen helicoidalmente, en la S2 forman una hélice vertical y en la S3, que es la capa más delgada e incluso puede faltar, se disponen como en la S1.
 y proteínas. La pared primaria es más gruesa y al microscopio electrónico se observan las macrofibrillas formadas por haces de microfibrillas adosadas longitudinalmente. La pared secundaria sólo está presente en algunos tipos celulares y suele ser mucho más gruesa que la pared primaria. Además de celulosa suele contener otras sustancias como hemicelulosa, proteínas, pectatos, lignina, cutina, suberina, sales minerales, etc, que, en general, tienden a darle mayor consistencia. Comprende tres subcapas que se denominan S1, S2 y S3 (de fuera hacia dentro). En la S1 las microfibrillas se disponen helicoidalmente, en la S2 forman una hélice vertical y en la S3, que es la capa más delgada e incluso puede faltar, se disponen como en la S1..")
20
Entre las funciones de la pared celular destacan principalmente las de dar forma y soporte mecánico a las células, a modo de exoesqueleto y ayudar en el mantenimiento del equilibrio hidrostático entre la célula y el medio que la rodea (manteniendo la presión hidrostática o presión de turgencia) y que es origen del movimiento de las células vegetales (apertura y cierre estomático, etc.). A pesar de su resistencia y grosor, la pared es permeable, tanto al agua como a las sustancias disueltas en ella, debido a la existencia de una serie de conductos que la atraviesan, como son los plasmodesmos, (canales atravesados por prolongaciones del RE), los campo de poros primarios (depresión en la pared primaria atravesadas por muchos plasmodesmos) y las punteaduras (adelgazamientos de la pared celular).
, los campo de poros primarios (depresión en la pared primaria atravesadas por muchos plasmodesmos) y las punteaduras (adelgazamientos de la pared celular).")
21
Parede celular de fungos
Quitina Polisacárido estructural en hongos y artrópodos Funciónes: - Protección frente ea la presión osmótica célular, turgencia (Hongos) - Sostén . Protección (Hongos y artrópodos) Polímero lineal de N-acetilglucosamina con enlaces b(1->4)
 - Sostén . Protección (Hongos y artrópodos) Polímero lineal de N-acetilglucosamina con enlaces b(1->4)")
22
Parede celular de bacterias

23
2- CITOSOL Y CITOESQUELETO.
El citoplasma es la parte de la célula comprendida entre la membrana y el núcleo. Está constituido por el citosol y los orgánulos. El citosol, hialoplasma o citoplasma fundamental, es la fracción soluble del citoplasma, el medio en el que se encuentran inmersos todos los orgánulos de la célula. Consiste en un medio acuoso (85% de agua) en el que aparecen disueltas gran cantidad de moléculas (aminoácidos, enzimas, proteínas, sales minerales, iones,...), constituyendo una disolución coloidal. Es un medio dinámico que puede pasar del estado de sol (fluido) al de gel (viscoso). Este cambio le permite crear corrientes internas que se conocen como ciclosis. Funciones del citosol 1. Es el medio en el que se mueven los orgánulos y el núcleo. 2. Algunas células, gracias a su capacidad de pasar de sol a gel, pueden emitir prolongaciones citoplasmáticas (pseudópodos) para trasladarse o incorporar sustancias. 3. Es el medio en donde se realizan muchos procesos metabólicos, como la síntesis de glucógeno y de pentosas, la glucólisis, las fermentaciones, el metabolismo de aminoácidos, etc.
 en el que aparecen disueltas gran cantidad de moléculas (aminoácidos, enzimas, proteínas, sales minerales, iones,...), constituyendo una disolución coloidal. Es un medio dinámico que puede pasar del estado de sol (fluido) al de gel (viscoso). Este cambio le permite crear corrientes internas que se conocen como ciclosis. Funciones del citosol. 1. Es el medio en el que se mueven los orgánulos y el núcleo. 2. Algunas células, gracias a su capacidad de pasar de sol a gel, pueden emitir prolongaciones citoplasmáticas (pseudópodos) para trasladarse o incorporar sustancias. 3. Es el medio en donde se realizan muchos procesos metabólicos, como la síntesis de glucógeno y de pentosas, la glucólisis, las fermentaciones, el metabolismo de aminoácidos, etc.")
24
Las estructuras filamentosas que constituyen el citoesqueleto pueden ser microtúbulos, microfilamentos y filamentos intermedios. · Microfilamentos o filamentos de actina. Son filamentos del citoesqueleto formados a partir de una proteína globular denominmada actina. Ésta forma polímeros de alrededor de 5 a 6 nm de diámetro. Están presentes en las células animales mostrando una organización de haces paralelos en dominios subcorticales y citoplasmáticos de la célula; también están presentes en las células vegetales. Los haces de filamentos de actina se encuentran en los fibroblastos formando las denominadas fibras de estrés. Los monómeros de forma globular (G-actina) se polimerizan en un proceso dependiente de ATP (análogo a la polimerización de microtúbulos dependiente de GTP), para formar el polímero de F-actina, que consta de dos filamentos centrales enrollados helicoidalmente en la estructura del microfilamento. Los microfilamentos son estructuras altamente dinámicas, cuya polimerización está regulada por proteínas de una familia conocida como proteínas de unión a actina (ABPs). SIGUE
 se polimerizan en un proceso dependiente de ATP (análogo a la polimerización de microtúbulos dependiente de GTP), para formar el polímero de F-actina, que consta de dos filamentos centrales enrollados helicoidalmente en la estructura del microfilamento. Los microfilamentos son estructuras altamente dinámicas, cuya polimerización está regulada por proteínas de una familia conocida como proteínas de unión a actina (ABPs). SIGUE.")
25
Los filamentos de actina poseen gran importancia en todos los procesos de desplazamiento y adhesión celular (emisión de pseudópodos). También juegan un papel importantísimo en la división celular, pues forman el anillo de contracción que permite el estrangulamiento celular durante la citocinesis. Otras funciones importantes son su intervención en la deformación de la membrana plasmática formando vesículas de endocitosis y exocitosis, su participación junto con la miosina en la contracción muscular así como su acción de soporte en estructuras como las microvellosidades. · Microtúbulos. Son filamentos del citoesqueleto que se caracterizan por estar construidos a partir de tubulina, una proteína dimérica (13 subunidades alfa y beta por vuelta) que se auto-ensambla en un proceso dependiente de GTP. Los microtúbulos tienen un diámetro de 25 nm y se originan en los centros organizadores de microtúbulos (principalmente en los centrosomas), adoptando una organización radial en las células interfásicas. Forman también parte del huso mitótico de todas las células eucariotas; se localizan en forma de haces en el axón neuronal, y también están presentes en el aparato locomotor de cilios y flagelos. Dan rigidez mecánica a las células y sirven de canales para el transporte intracelular (melenina). Los microtúbulos son estructuras altamente dinámicas, estabilizadas por un grupo de proteínas denominadas proteínas asociadas a microtúbulos (MAPs).En una célula se produce un recambio continuo de la red de microtúbulos. Este comportamiento, llamado inestabilidad dinámica, juega un papel muy importante en el posicionamiento de los microtúbulos en la célula. · Filamentos intermedios. Son estructuras del citoesqueleto de alrededor de 10 nm de diámetro, formados por un conjunto de proteínas específicas para cada tipo celular. Algunos ejemplos son los tonofilamentos (formados por queratina y confluyen en los desmosomas, por ejemplo en células epidérmicas), neurofilamentos (se encuentran en células nerviosas), filamentos de desmina (en células musculares), etc.
 que se auto-ensambla en un proceso dependiente de GTP. Los microtúbulos tienen un diámetro de 25 nm y se originan en los centros organizadores de microtúbulos (principalmente en los centrosomas), adoptando una organización radial en las células interfásicas. Forman también parte del huso mitótico de todas las células eucariotas; se localizan en forma de haces en el axón neuronal, y también están presentes en el aparato locomotor de cilios y flagelos. Dan rigidez mecánica a las células y sirven de canales para el transporte intracelular (melenina). Los microtúbulos son estructuras altamente dinámicas, estabilizadas por un grupo de proteínas denominadas proteínas asociadas a microtúbulos (MAPs).En una célula se produce un recambio continuo de la red de microtúbulos. Este comportamiento, llamado inestabilidad dinámica, juega un papel muy importante en el posicionamiento de los microtúbulos en la célula. · Filamentos intermedios. Son estructuras del citoesqueleto de alrededor de 10 nm de diámetro, formados por un conjunto de proteínas específicas para cada tipo celular. Algunos ejemplos son los tonofilamentos (formados por queratina y confluyen en los desmosomas, por ejemplo en células epidérmicas), neurofilamentos (se encuentran en células nerviosas), filamentos de desmina (en células musculares), etc.")
26
Centrosoma : En las células eucariotas, el centrosoma se localiza en una zona cercana al núcleo y está rodeado muchas veces por los dictiosomas del Aparato de Golgi. Está formado por: a) Un par de centriolos o diplosoma. se sitúan ocupando el centro del centrosoma y se disponen perpendicularmente entre sí. En la división celular, cada uno de ellos da origen por duplicación a su pareja. De este modo se obtienen dos diplosomas, uno para cada célula hija. Las células de los vegetales superiores carecen de centríolos. b) El material pericentriolar o esfera atractiva, formado por un material de aspecto amorfo, que rodea al diplosoma. c) Las fibras del áster, formadas por microtúbulos que crecen y se organizan en forma de radios a partir del material pericentriolar. Durante la división celular, los microtúbulos del áster dan origen a los microtúbulos del huso acromático. El material pericentriolar y las fibras del áster, aparecen incluso cuando no hay centríolos. Función del centrosoma: La función de los centríolos está relacionada con la organización de los microtúbulos. Sin embargo basta con el material pericentriolar para realizar esa función, como se demuestra en la formación del huso acromático en las células vegetales que carecen de centriolos.
 Un par de centriolos o diplosoma. se sitúan ocupando el centro del centrosoma y se disponen perpendicularmente entre sí. En la división celular, cada uno de ellos da origen por duplicación a su pareja. De este modo se obtienen dos diplosomas, uno para cada célula hija. Las células de los vegetales superiores carecen de centríolos. b) El material pericentriolar o esfera atractiva, formado por un material de aspecto amorfo, que rodea al diplosoma. c) Las fibras del áster, formadas por microtúbulos que crecen y se organizan en forma de radios a partir del material pericentriolar. Durante la división celular, los microtúbulos del áster dan origen a los microtúbulos del huso acromático. El material pericentriolar y las fibras del áster, aparecen incluso cuando no hay centríolos. Función del centrosoma: La función de los centríolos está relacionada con la organización de los microtúbulos. Sin embargo basta con el material pericentriolar para realizar esa función, como se demuestra en la formación del huso acromático en las células vegetales que carecen de centriolos.")
27
CILIOS Y FLAGELOS Son prolongaciones móviles localizadas en la superficie de muchas células. Cilios y flagelos son estructuralmente semejantes y se diferencian en el tamaño y en el número Poseen dos partes: 1. Tallo o axonema: es una evaginación digitiforme de la membrana plasmática en cuyo interior existen 20 microtúbulos con disposición 9+1 (9 pares periféricos, los dobletes, y uno central, los singletes) encontrándose estos últimos rodeados por una vaina. 2. Cuerpo basal: es la zona por la que el cilio o flagelo está sujeto al citoplasma cortical y es perpendicular a la superficie celular. La estructura del cuerpo basal es idéntica a la de los centriolos y está formado por un corto cilindro de 3 tripletes de microtúbulos (A, B y C), sin microtúbulos centrales. Funciona como centro organizador de microtúbulos, desde donde crece el cilio o flagelo. En la zona de transición entre los cuerpos basales y el eje del cilio/flagelo acaban los túbulos C y comienzan los singletes: los túbulos A y B del cuerpo basal se continúan en los 9 dobletes del axonema.
 encontrándose estos últimos rodeados por una vaina. 2. Cuerpo basal: es la zona por la que el cilio o flagelo está sujeto al citoplasma cortical y es perpendicular a la superficie celular. La estructura del cuerpo basal es idéntica a la de los centriolos y está formado por un corto cilindro de 3 tripletes de microtúbulos (A, B y C), sin microtúbulos centrales. Funciona como centro organizador de microtúbulos, desde donde crece el cilio o flagelo. En la zona de transición entre los cuerpos basales y el eje del cilio/flagelo acaban los túbulos C y comienzan los singletes: los túbulos A y B del cuerpo basal se continúan en los 9 dobletes del axonema.")
28
Los cilios y flagelos bacterianos son distintos a los de las células eucarióticas, pues son estructuras extracelulares que no están rodeadas por la membrana plasmática. Lo forman un filamento en espiral de flagelina (proteína globular) y se encuentran anclados a la bacteria por una base formada por varios compuestos. El factor que permite el delizamiento de los microtúbulos es la dineína. En un cilio o flagelo en reposo los brazos de dineína de un doblete no lle-gan a tocar al doblete vecino, pero establecen contacto cuan-do se les proporciona ATP; es la actividad ATP-asa de la dineína la que aporta la energía necesaria para el movimiento.

29
Son orgánulos globulares (sin membrana) que se originan en el nucleolo del núcleo celular. Pueden encontrarse dispersos por el hialoplasma, unidos a la superficie externa de la envoltura nuclear, unidos a la membrana del RER o formando acúmulos en forma de rosario unidos a un filamento de ARNm (llamados entonces polisomas o polirribosomas). También pueden aparecer en el interior de mitocondrias y cloroplastos, aunque en este caso se trata de ribosomas semejantes a los de procariotas. Están compuestos por agua (80%), proteínas (10%) y ARNr (10%), que se distribuyen en dos subunidades globulares. En eucariotas la mayor de ellas es de 65 S (unidades Svedberg) y la menor de 40S, mientras que en procariotas las subunidades son de 50S y 30S, respectivamente. Su función es la síntesis de proteínas, que tiene lugar cuando un ribosoma establece contacto con una cadena de ARNm y traduce la secuencia de nucleótidos a una secuencia de aminoácidos (tema 24).
, proteínas (10%) y ARNr (10%), que se distribuyen en dos subunidades globulares. En eucariotas la mayor de ellas es de 65 S (unidades Svedberg) y la menor de 40S, mientras que en procariotas las subunidades son de 50S y 30S, respectivamente. Su función es la síntesis de proteínas, que tiene lugar cuando un ribosoma establece contacto con una cadena de ARNm y traduce la secuencia de nucleótidos a una secuencia de aminoácidos (tema 24).")
30
Inclusiones Son enclaves que carecen de membrana y en los que se acumulan sustancias que difícilmente se dispersan por el citoplasma. Las sustancias pueden ser de naturaleza muy diversa. Algunos ejemplos son los siguientes: 1. En células vegetales: gotas de grasa, aceites esenciales, látex, drusas (cristales de oxalato cálcico), etc. 2. En células animales: glucógeno, lípidos (adipocitos), pigmentos (melanina, hemosiderina), etc.
, etc. 2. En células animales: glucógeno, lípidos (adipocitos), pigmentos (melanina, hemosiderina), etc.")
31
3- SISTEMAS DE MEMBRANA Y ORGÁNULOS.
Los orgánulos no se distribuyen al azar en la célula, esto es así porque sus interiores se comunican intensamente entre si y con el exterior celular, vía vesículas de transporte, esto se pude explicar teniendo en cuenta como han evolucionado.

32
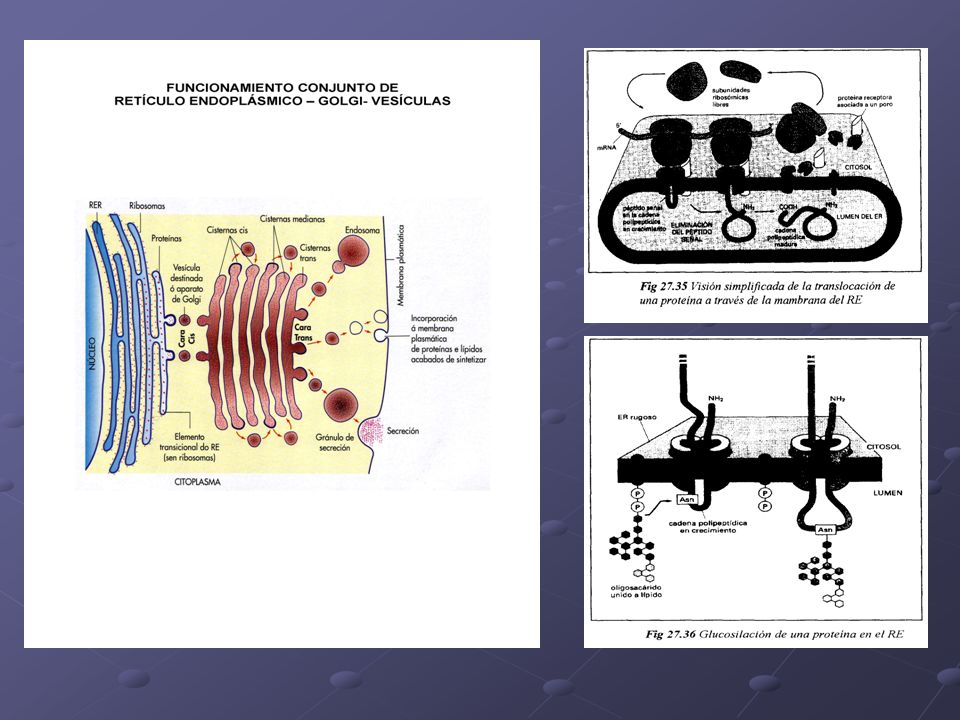
Es un orgánulo membranoso que ocupa alrededor del 10 % del volumen celular y está constituido por una amplia red de conductos y cavidades cerradas, cuya forma (láminas aplanadas, vesículas globulares o tubos de aspecto sinuoso) y extensión varía con la actividad de la célula. En las células nerviosas, los sáculos del retículo aparecen formando acúmulos conocidos como grumos de Nissl. La membrana del RE es más delgada que la membrana plasmática, pero su estructura es análoga con la diferencia que en el retículo la proporción de lípidos es menor y la de proteínas mayor. La porción del retículo endoplasmático que hace frontera entre el núcleo y el hialoplasma, constituye la envoltura nuclear, Existen dos tipos de retículo endoplasmático: el, con ribosomas adosados en el lado de la membrana que da al hialoplasma, y el sin ribosomas. · Retículo endoplasmático rugoso (RER):. Se continúa con la envoltura nuclear. Sus funciones principales son la síntesis y el transporte de proteínas. Si estas proteínas forman parte de los productos de secreción de la célula son transferidas y acumuladas en el interior de las cisternas (por ejemplo, los anticuerpos). Si, su destino es formar parte de las membranas celulares, son transferidas a la membrana del RE. · Retículo endoplasmático liso (REL): Está en continuidad con el RER, del cual se forma como si se tratara de evaginaciones. Su función es sintetizar, transportar y almacenar sustancias. Interviene en la síntesis de fosfolipidos y de colesterol, en las reacciones de glicosilación y en las de detoxificación (eliminación de sustancias tóxicas transformándolas en otras solubles que pueden abandonar la célula y ser excretadas).
:. Se continúa con la envoltura nuclear. Sus funciones principales son la síntesis y el transporte de proteínas. Si estas proteínas forman parte de los productos de secreción de la célula son transferidas y acumuladas en el interior de las cisternas (por ejemplo, los anticuerpos). Si, su destino es formar parte de las membranas celulares, son transferidas a la membrana del RE. · Retículo endoplasmático liso (REL): Está en continuidad con el RER, del cual se forma como si se tratara de evaginaciones. Su función es sintetizar, transportar y almacenar sustancias. Interviene en la síntesis de fosfolipidos y de colesterol, en las reacciones de glicosilación y en las de detoxificación (eliminación de sustancias tóxicas transformándolas en otras solubles que pueden abandonar la célula y ser excretadas).")
33
Es un orgánulo membranoso formado por un conjunto de sáculos discoidales y aplanados, ligeramente dilatados en sus extremos y que se agrupan en pilas de 5 a 10 unidades que se denominan dictiosomas. Las cavidades están delimitadas por una membrana unitaria y llenas de fluido. Los distintos dictiosomas se encuentran conectados entre sí. Suele encontrarse rodeando al núcleo o al centrosoma. Es una parte especializada del retículo endoplasmático, ya que se forma a sus expensas, y sus funciones son complementarias. La cara cóncava de los sáculos (cara Cis o de formación), que es más fina y similar a la del retículo, está rodeada por cisternas del RE que, por gemación, desprenden vesículas. Estas vesículas se denominan vesículas de transición y están cargadas de productos almacenados en el retículo endoplasmático. Varias se fusionan formando los sáculos del dictiosoma, que van siendo desplazados a medida que se van formando los nuevos. Al llegar a la cara convexa del dictiosorna (cara Trans, que presenta membranas gruesas y similares a las de la membrana plasmática) se fragmentan en vesículas de secreción. El aparato de Colgi interviene en la secreción y en la acumulación de sustancias glicoxiladas (unión de glúcidos a proteínas de membrana, anticuerpos, etc.), en la producción y renovación de la membrana plasmática, en el transporte de moléculas a través del citoplasma mediante vesículas (como pueden ser la celulosa y los pectatos que forman la pared celular), y en la formación de lisosomas. A veces, puede tener funciones concretas, como por ejemplo, la formación del acrosoma de los espermatozoides
, que es más fina y similar a la del retículo, está rodeada por cisternas del RE que, por gemación, desprenden vesículas. Estas vesículas se denominan vesículas de transición y están cargadas de productos almacenados en el retículo endoplasmático. Varias se fusionan formando los sáculos del dictiosoma, que van siendo desplazados a medida que se van formando los nuevos. Al llegar a la cara convexa del dictiosorna (cara Trans, que presenta membranas gruesas y similares a las de la membrana plasmática) se fragmentan en vesículas de secreción. El aparato de Colgi interviene en la secreción y en la acumulación de sustancias glicoxiladas (unión de glúcidos a proteínas de membrana, anticuerpos, etc.), en la producción y renovación de la membrana plasmática, en el transporte de moléculas a través del citoplasma mediante vesículas (como pueden ser la celulosa y los pectatos que forman la pared celular), y en la formación de lisosomas. A veces, puede tener funciones concretas, como por ejemplo, la formación del acrosoma de los espermatozoides.")
35
Las vacuolas son orgánulos rodeados por una membrana, llamada tonoplasto, y están mucho más desarrolladas en las células vegetales que en la animales. En el caso de las células vegetales, pueden ocupar hasta el 50% del volumen celular y, en algunos casos, llegar a superar el 90%. En las células meristemáticas son abundantes y de pequeño tamaño. En las más diferenciadas pueden fusionarse, originando vacuolas que ocupan gran parte del citoplasma. Al conjunto de vacuolas de una célula se le llama vacuoma. Se forman en células jóvenes, por fusión de vesículas derivadas del RE y del aparato de Golgi. Están relacionadas estructural y funcionalmente con los lisosomas y contienen gran cantidad de enzimas hidrolíticos, por lo que pueden intervenir en los procesos de nutrición (vacuolas digestivas) Pueden, además, almacenar sustancias de reserva (almidón, grasas, proteínas,...), pigmentos, productos de deshecho resultantes del metabolismo, esencias, alcaloides, resinas, glucósidos (muchos son venenos), cristales de sustancias minerales, etc. Además, participan en la regulación de la presión osmótica de la célula y en el crecimiento (las células vegetales crecen, entre otras cosas, por acumulación de agua en las vacuolas). Un tipo especial de vacuolas son las vacuolas pulsátiles, que poseen una salida al exterior por la que expulsan el exceso de agua mediante pulsaciones (típicas de protozoos que viven en ambientes hipotónicos).
 Pueden, además, almacenar sustancias de reserva (almidón, grasas, proteínas,...), pigmentos, productos de deshecho resultantes del metabolismo, esencias, alcaloides, resinas, glucósidos (muchos son venenos), cristales de sustancias minerales, etc. Además, participan en la regulación de la presión osmótica de la célula y en el crecimiento (las células vegetales crecen, entre otras cosas, por acumulación de agua en las vacuolas). Un tipo especial de vacuolas son las vacuolas pulsátiles, que poseen una salida al exterior por la que expulsan el exceso de agua mediante pulsaciones (típicas de protozoos que viven en ambientes hipotónicos).")
36
Son unos orgánulos globulares que se encuentran rodeados por una membrana unitaria, en cuyo interior se alojan enzimas hidrolíticos o digestivos. Un lisosoma puede llegar a tener hasta 50 enzimas distintos en estado inactivo. Se cree que la membrana de los lisosomas no es atacada por los enzimas que contiene debido a que interiormente presenta una gruesa capa de glucoproteínas, que protegería al lisosoma y, por ello, también a la célula, de la autodigestión. Están presentes en todas las células eucariotas, excepto en los glóbulos rojos de mamíferos. Se originan a partir del aparato de Golgi o del retículo endoplasmático liso. Se habla de lisosomas primarios si sólo contienen enzimas hidroliticos y su aspecto es homogéneo, mientras que se llaman lisosomas secundarios si además de los enzimas, contienen sustancias en proceso de digestión; poseen entonces un contorno irregular, mayor tamaño y contenido heterogéneo. La digestión que llevan a cabo puede ser de dos tipos: 1. Extracelular. En ella los lisosomas vierten su contenido al exterior de la célula por exocitosis. 2. Intracelular. Los sustratos a digerir pueden ser: a) externos (heterofagia) es una función de nutrición o de defensa frente a infecciones (ej.: los leucocitos fagocitan bacterias), de limpieza (ej. los macrófagos fagocitan restos), de reabsorción (ej.: renal), de destrucción de sustancias (ej.: los hepatocitos), etc. Los sustratos son capturados por endocitosis y se forma una vesícula que se fusiona con un lisosoma primario, originando una vacuola digestiva.
 externos (heterofagia) es una función de nutrición o de defensa frente a infecciones (ej.: los leucocitos fagocitan bacterias), de limpieza (ej. los macrófagos fagocitan restos), de reabsorción (ej.: renal), de destrucción de sustancias (ej.: los hepatocitos), etc. Los sustratos son capturados por endocitosis y se forma una vesícula que se fusiona con un lisosoma primario, originando una vacuola digestiva.")
37
Según su función se habla de:
b) internos (autofagia) el sustrato es un constituyente celular (porciones de otros orgánulos, vacuolas,...). Sirve para destruir zonas dañadas o innecesarias de las células. A estos lisosomas se les llama autolisosomas. Esta función desempeña un papel importante en la vida de las células, ya que destruye zonas dañadas o innecesarias, interviene en los procesos de desarrollo y asegura la nutrición en condiciones desfavorables. Según su función se habla de: · Fagolisosomas (vacuolas digestivas o heterolisosomas). Se trata de orgánulos formados a partir de la unión de un lisosoma primario con una vacuola fagocítica, que contienen partículas alimenticias de naturaleza muy variada según el tipo de célula. Son abundantes en las amebas, que experimentan procesos de fagocitosis, los cuales constituyen el mecanismo fundamental de nutrición en estos protozoos, y son esenciales en las células implicadas en la defensa del organismo, como los macrófagos (leucocitos especializados en la fagocitosis de microorganismos patógenos). · Autofagolisosomas. En este caso, los lisosomas se fusionan con vacuolas autofagocíticas para eliminar restos celulares, corno orgánulos dañados por sustancias tóxicas o que deben ser reciclados. Son importantes en los procesos de autofagia que se producen en la formación de los tejidos de sostén en los vegetales y en la metamorfosis de los insectos, durante la que se eliminan algunos elementos larvales. Cuerpos multivesiculares. Son lisosomas que contienen en su interior numerosas vesículas. Puede tratarse de vesículas autofágicas o de endocitosis en el interior de lisosomas primarios, o de varios lisosomas primarios reunidos en una membrana común. Además de la función digestiva, los lisosomas también pueden participar en procesos de almacenamiento de sustancias de reserva. En las semillas existe un tipo especial de lisosomas secundarios, los granos de aleurona, que almacenan sustancias de reserva. En ellos no tiene lugar la digestión celular hasta que no llega el momento de la germinación; llegado éste, los tejidos de la semilla, que habían perdido gran cantidad de agua, se rehidratan, activándose las enzimas hidrolíticas, e iniciándose la digestión intracelular cuyos productos serán utilizados por el embrión en desarrollo.
 internos (autofagia) el sustrato es un constituyente celular (porciones de otros orgánulos, vacuolas,...). Sirve para destruir zonas dañadas o innecesarias de las células. A estos lisosomas se les llama autolisosomas. Esta función desempeña un papel importante en la vida de las células, ya que destruye zonas dañadas o innecesarias, interviene en los procesos de desarrollo y asegura la nutrición en condiciones desfavorables. Según su función se habla de: · Fagolisosomas (vacuolas digestivas o heterolisosomas). Se trata de orgánulos formados a partir de la unión de un lisosoma primario con una vacuola fagocítica, que contienen partículas alimenticias de naturaleza muy variada según el tipo de célula. Son abundantes en las amebas, que experimentan procesos de fagocitosis, los cuales constituyen el mecanismo fundamental de nutrición en estos protozoos, y son esenciales en las células implicadas en la defensa del organismo, como los macrófagos (leucocitos especializados en la fagocitosis de microorganismos patógenos). · Autofagolisosomas. En este caso, los lisosomas se fusionan con vacuolas autofagocíticas para eliminar restos celulares, corno orgánulos dañados por sustancias tóxicas o que deben ser reciclados. Son importantes en los procesos de autofagia que se producen en la formación de los tejidos de sostén en los vegetales y en la metamorfosis de los insectos, durante la que se eliminan algunos elementos larvales. Cuerpos multivesiculares. Son lisosomas que contienen en su interior numerosas vesículas. Puede tratarse de vesículas autofágicas o de endocitosis en el interior de lisosomas primarios, o de varios lisosomas primarios reunidos en una membrana común. Además de la función digestiva, los lisosomas también pueden participar en procesos de almacenamiento de sustancias de reserva. En las semillas existe un tipo especial de lisosomas secundarios, los granos de aleurona, que almacenan sustancias de reserva. En ellos no tiene lugar la digestión celular hasta que no llega el momento de la germinación; llegado éste, los tejidos de la semilla, que habían perdido gran cantidad de agua, se rehidratan, activándose las enzimas hidrolíticas, e iniciándose la digestión intracelular cuyos productos serán utilizados por el embrión en desarrollo.")
38
Son orgánulos muy parecidos a los lisosomas, pero se diferencian en que no contienen hidrolasas ácidas; contienen "enzimas oxidativos", de las cuales las más abundantes son: la catalasa, que es una enzima oxidativa y las oxidasas que contiene FAD como coenzimas. Son responsables de dos funciones básicas: formación de agua oxigenada y descomposición de la misma: catalasa H2O H2O O2 En las semillas en germinación, existe un tipo especial de peroxisomas: los glioxisomas (ciclo del glioxilato), que transforman los ácidos grasos de la semilla en azúcares para el desarrollo del embrión. La b-oxidación en mitocondrias (eucariotas) y peroxisomas (eucariotas,vegetales y levaduras solo aqui) Las teorías más recientes sobre la evolución de las células eucariotas propone un origen endosimbiótico . Su función en las células ancestrales habría consistido en la detoxificación de los compuestos tóxicos generados por la creciente concentración de oxígeno en la atmósfera. Posteriormente surgirían las mitocondrias, cuya mayor eficacia para metabolizar este elemento y su capacidad para generar ATP por medio de la fosforilación oxidativa harían obsoletas varias de sus funciones y sólo se mantendrían aquellas que todavía resultaran útiles para la célula.
, que transforman los ácidos grasos de la semilla en azúcares para el desarrollo del embrión. La b-oxidación en mitocondrias (eucariotas) y peroxisomas (eucariotas,vegetales y levaduras solo aqui) Las teorías más recientes sobre la evolución de las células eucariotas propone un origen endosimbiótico . Su función en las células ancestrales habría consistido en la detoxificación de los compuestos tóxicos generados por la creciente concentración de oxígeno en la atmósfera. Posteriormente surgirían las mitocondrias, cuya mayor eficacia para metabolizar este elemento y su capacidad para generar ATP por medio de la fosforilación oxidativa harían obsoletas varias de sus funciones y sólo se mantendrían aquellas que todavía resultaran útiles para la célula.")
39
Se encuentran en todas las células eucariotas
Se encuentran en todas las células eucariotas. Son unos orgánulos ovoideos limitados por una doble membrana; la membrana mitocondrial externa es lisa y la separa del hialoplasma, mientras que la membrana mitocondnal interna forma invaginaciones o pliegues llamados crestas mitocondriales. Entre ambas membranas existe un espacio intermembrana. El espacio que queda delimitado por la membrana interna es la matriz. Suelen aparecer en elevado número y a su conjunto se le conoce como condrioma. · La matriz mitocondrial está ocupada por un líquido acuoso, que incluye: 1. Moléculas de ADN doble y circular, con información para sintetizar muchas de las proteínas mitocondriales. 2. Ribosomas 70S (mitorribosomas), tanto libres como adosados a la membrana interna. 3. Enzimas para la transcripción, replicación y traducción del ADN (síntesis proteica en los mitorribosomas) y enzimas que intervienen en la oxidación de moléculas en el ciclo de Krebs. 4. Iones Ca+2 y fosfato, ATP, ADP y Coenzima A. 5. En ocasiones, en la matriz pueden haber gránulos que no están rodeados por membrana, sino que están constituidos por laminillas dispuestas de forma des ordenada.
, tanto libres como adosados a la membrana interna. 3. Enzimas para la transcripción, replicación y traducción del ADN (síntesis proteica en los mitorribosomas) y enzimas que intervienen en la oxidación de moléculas en el ciclo de Krebs. 4. Iones Ca+2 y fosfato, ATP, ADP y Coenzima A. 5. En ocasiones, en la matriz pueden haber gránulos que no están rodeados por membrana, sino que están constituidos por laminillas dispuestas de forma des ordenada.")
40
· La membrana mitocondrial extrena se asemeja a otras membranas celulares, en particular a las del retículo endoplásmico. Entre sus proteínas destacan las formadoras canales acuosos, que la hacen muy permeable, y enzimas que activan los ácidos grasos para ser oxidados en la matriz. · La membrana mitocondrial intrena posee una gran superficie gracias a las crestas mitocondriales. Es más rica en proteínas (80%) que otras membranas celulares, pobre en lípidos (20%) y no presenta colesterol. Entre sus proteínas se pueden distinguir: 1. Proteínas de la cadena de transporte electrónico. 2. La ATP-sintetasa, un complejo enzimático que cataliza la síntesis de ATP. Está formada por tres partes: el factor F1, que es la parte catalítica del complejo, el factor F0, que funciona de pedúnculo del primero, y una base hidrófoba que integra el conjunto en la membrana. 3. Proteínas transportadoras, que permiten el paso de determinados iones y moléculas, pues la membrana interna es bastante selectiva. · El espacio intermembrana presenta una composición similar a la del hialoplasma debido a la alta permeabilidad de la membrana externa. Las enzimas que posee le permiten transferir la energía del ATP fabricado en la mitocondria a otros nucleótidos, como por ejemplo el AMP. La principal función de las mitocondrias es la de obtener energía metabólica en forma de ATP como resultado de los procesos de la respiración celular (ciclo de Krebs y transporte electrónico). Otra función es su intervención en la síntesis de ácidos grasos y algunas proteínas, así como en la b-oxidación de los primeros. Además, la mitocondrias pueden funcionar como almacenes de lípidos, prótidos, Fe2+, Na+, K+, etc.
 que otras membranas celulares, pobre en lípidos (20%) y no presenta colesterol. Entre sus proteínas se pueden distinguir: 1. Proteínas de la cadena de transporte electrónico. 2. La ATP-sintetasa, un complejo enzimático que cataliza la síntesis de ATP. Está formada por tres partes: el factor F1, que es la parte catalítica del complejo, el factor F0, que funciona de pedúnculo del primero, y una base hidrófoba que integra el conjunto en la membrana. 3. Proteínas transportadoras, que permiten el paso de determinados iones y moléculas, pues la membrana interna es bastante selectiva. · El espacio intermembrana presenta una composición similar a la del hialoplasma debido a la alta permeabilidad de la membrana externa. Las enzimas que posee le permiten transferir la energía del ATP fabricado en la mitocondria a otros nucleótidos, como por ejemplo el AMP. La principal función de las mitocondrias es la de obtener energía metabólica en forma de ATP como resultado de los procesos de la respiración celular (ciclo de Krebs y transporte electrónico). Otra función es su intervención en la síntesis de ácidos grasos y algunas proteínas, así como en la b-oxidación de los primeros. Además, la mitocondrias pueden funcionar como almacenes de lípidos, prótidos, Fe2+, Na+, K+, etc.")
41
Son una familia de orgánulos propios de células vegetales y que se caracterizan por tener un origen común, información genética propia y doble membrana. Los tipos de plastos son: 1. Cromoplastos: dan color amarillo, anaranjado o rojo a las flores y frutos. Acumulan carotenoides. 2. Leucoplastos: se encuentran en las zonas no verdes del vegetal, son de color blanco y entre ellos destacan los arniloplastos (acumulan almidón). 3. Etioplastos: se forman cuando una célula vegetal crece en oscuridad. Poseen un pigmento amarillo, la protoclorofila. Se convierten en cloroplastos si son expuestos a la luz. 4. Cloroplastos: poseen color debido a la presencia de clorofila.
. 3. Etioplastos: se forman cuando una célula vegetal crece en oscuridad. Poseen un pigmento amarillo, la protoclorofila. Se convierten en cloroplastos si son expuestos a la luz. 4. Cloroplastos: poseen color debido a la presencia de clorofila.")
42
CLOROPLASTO Poseen forma lenticular. Se encuentran rodeados por dos membranas, la membrana plastidial externa y la membrana plastidial interna, entre las que existe un espacio intermembrana. La membrana interna encierra un gran espacio central, el estroma, en el que se baña un tercer tipo de membrana, la membrana tilacoidal La membrana tilacoidal se organiza formando unas vesículas discoidales y aplanadas que se superponen formando pilas y que reciben el nombre de grana. En algunos vegetales, como en el maíz, los cloroplastos no poseen grana. · Las membranas interna y externa presentan alrededor de un 60% de lípidos y un 40% de proteínas y carecen de clorofila y de colesterol. La membrana externa es mucho más permeable que la interna. · El estroma es un fluido que contiene moléculas de ADN plastidial doble y circular (con información para la síntesis de ARN y proteínas propias del cloroplasto), ribosomas 70S (plastorribosomas, que llegan a representar el 50% de los ribosomas de la célula vegetal) y enzimas (que intervienen en los procesos de replicación del ADN plastidial y en reacciones de reducción del CO2, nitratos y sulfatos a materia orgánica). · La membrana de los tilacoides está compuesta por un 38% de lípido, un 50% de proteínas y un 12% de pigmentos. Los pigmentos corresponden el 2% a carotenoides (carotenos y xantofilas, de color amarillo o anaranjado) y el 10% a clorofilas a y b. En ciertas algas existen pigmentos accesorios (ficoeritnna o ficocianina). Entre las proteínas se distinguen: 1. Proteínas asociadas a los pigmentos, formando grandes complejos integrados en la membrana: son los fotosistemas I y II, que poseen una función muy importante en la primera etapa de la fotosíntesis (ver tema 28). 2. Partículas F o factores de acoplamiento, con forma de esferas pedunculadas que sobresalen por la cara de los tilacoides que están en contacto con el estroma del plasto. Contiene la enzima ATP-sintetasa, responsable de formar el ATP que se produce en la fase luminosa de la fotosíntesis. 3. Proteínas transportadoras de electrones. En los cloroplastos tiene lugar la síntesis de gran número de proteínas, aunque la función más importante de estos orgánulos es la de llevar a cabo las reacciones de la fotosíntesis (tema 28).
, ribosomas 70S (plastorribosomas, que llegan a representar el 50% de los ribosomas de la célula vegetal) y enzimas (que intervienen en los procesos de replicación del ADN plastidial y en reacciones de reducción del CO2, nitratos y sulfatos a materia orgánica). · La membrana de los tilacoides está compuesta por un 38% de lípido, un 50% de proteínas y un 12% de pigmentos. Los pigmentos corresponden el 2% a carotenoides (carotenos y xantofilas, de color amarillo o anaranjado) y el 10% a clorofilas a y b. En ciertas algas existen pigmentos accesorios (ficoeritnna o ficocianina). Entre las proteínas se distinguen: 1. Proteínas asociadas a los pigmentos, formando grandes complejos integrados en la membrana: son los fotosistemas I y II, que poseen una función muy importante en la primera etapa de la fotosíntesis (ver tema 28). 2. Partículas F o factores de acoplamiento, con forma de esferas pedunculadas que sobresalen por la cara de los tilacoides que están en contacto con el estroma del plasto. Contiene la enzima ATP-sintetasa, responsable de formar el ATP que se produce en la fase luminosa de la fotosíntesis. 3. Proteínas transportadoras de electrones. En los cloroplastos tiene lugar la síntesis de gran número de proteínas, aunque la función más importante de estos orgánulos es la de llevar a cabo las reacciones de la fotosíntesis (tema 28).")
43
4- MOTILIDAD CELULAR. La motilidad celular implica la transformación de energía química ATP, en energía mecánica. Podemos agrupar estas reacciones motrices en dos grupos: 1- Desplazamiento de toda la célula, locomoción · Movimientos ameboides. Se producen en células libres, sin membrana rígida, como amebas, leucocitos, etc. En ellas, el ectoplasma y el endoplasma están en estado de gel y de sol, respectivamente. La formación de un seudópodo se origina mediante un desplazamiento del endoplasma en el sentido de la emisión. Luego éste se extiende lateralmente y adquiere la consistencia de gel. Al mismo tiempo, el gel posterior pasa a sol y fluye hacia adelante. · Movimientos vibrátiles. Se dan en células que presentan cilios o flagelos, como los espermatozoides, protozoos ciliados, etc. El movimiento de un cilio consiste en la vibración a modo de golpe de remo, volviendo al estado inicial al curvarse desde la base al extremo. Según la hipótesis del deslizamiento, el cilio se curva al deslizarse un doblete sobre el otro. La energía necesaria para el deslizamiento la suministra el ATP en presencia de Ca2+ y Mg2+. El movimiento de un flagelo puede ser semejante al ciliar o de tipo ondulatorio. Cuando las vibraciones de una hilera de cilios, denominada cinetia, ocurren de forma que cada cilio comienza su vibración poco después del anterior y un poco antes que el posterior, se dice que el ritmo es metacronal. Cuando todos los cilios vibran al mismo tiempo, posee un ritmo isocronal. 2- Movimientos de algunas partes de la célula: Tropismos y tactismos. Hay estímulos que producen una reacción de la célula, la cual puede consistir en movimientos de desplazamiento respecto al estímulo denominados taxias o tactismos. Si no hay desplazamiento, sino únicamente la orientación de la célula, se habla de tropismo. Si el movimiento se realiza hacia el estímulo, se denomina tactismo positivo y si se efectúa alejándose del estímulo, tactismo negativo. Cuando el estímulo es mecánico, se habla de tigmotactismo; si es la fuerza de la gravedad geotactismo; si es la temperatura, termotactismo; si es la luz, fototactismo y si es una sustancia química, se habla de quimiotactismo.

44
· Movimientos de ciclosis
· Movimientos de ciclosis. Son corrientes endocelulares del hialoplasma que arrastran a los orgánulos. Se dan en las células dotadas de una estructura esquelética que mantiene su forma. Así ocurre en las células vegetales, que poseen una membrana esquelética y una gran vacuola central. El hialoplasma más próximo a la membrana plasmática (ectoplasma) está en estado de gel, y el más interno (endoplasma) es más fluido. El gel cortical presenta numerosos microtúbulos paralelos a la membrana plasmática que sirven, probablemente, para orientar las corrientes del hialoplasma. En el ectoplasma se sitúan los complejos proteicos contráctiles que originan el movimiento del endoplasma. · Movimientos contráctiles. Consisten en movimientos del hialoplasma en una dirección fija, lo cual provoca un acortamiento de la célula. Así, por ejemplo, en las células musculares, el movimiento de contracción se debe a la presencia de moléculas proteicas fibrosas en el hialoplasma. Estas moléculas constituyen los miofilamentos que están agrupados en miofibrillas, las cuales pueden contraerse debido al desplazamiento de unos miofilamentos respecto de otros.
 está en estado de gel, y el más interno (endoplasma) es más fluido. El gel cortical presenta numerosos microtúbulos paralelos a la membrana plasmática que sirven, probablemente, para orientar las corrientes del hialoplasma. En el ectoplasma se sitúan los complejos proteicos contráctiles que originan el movimiento del endoplasma. · Movimientos contráctiles. Consisten en movimientos del hialoplasma en una dirección fija, lo cual provoca un acortamiento de la célula. Así, por ejemplo, en las células musculares, el movimiento de contracción se debe a la presencia de moléculas proteicas fibrosas en el hialoplasma. Estas moléculas constituyen los miofilamentos que están agrupados en miofibrillas, las cuales pueden contraerse debido al desplazamiento de unos miofilamentos respecto de otros.")
Presentaciones similares