Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Anabolismo

2
Clasificación de los organismos en función de la forma de nutrición
Dador de electrones Fuente de energía Luz (fotótrofo) Sustrato oxidable (Quimiótrofo) Orgánico Heterótrofo Fotoorganotrofos Bacterias rojas no sulfúreas Quimioorganotrofo Animales, hongos, protrozoos, mayoría de bacterias. Inorgánico Autótrofo Fotolitótrofos Plantas verdes, algas, bacterias rojas y verdes del azufre. Quimiolitótrofo Bacterias del hidrógeno, incoloras del azufre, nitrificantes y férricas.
 Sustrato oxidable (Quimiótrofo) Orgánico. Heterótrofo. Fotoorganotrofos. Bacterias rojas no sulfúreas. Quimioorganotrofo. Animales, hongos, protrozoos, mayoría de bacterias. Inorgánico. Autótrofo. Fotolitótrofos. Plantas verdes, algas, bacterias rojas y verdes del azufre. Quimiolitótrofo. Bacterias del hidrógeno, incoloras del azufre, nitrificantes y férricas.")
3
Fotosíntesis Pigmentos fotosintéticos y fotosistemas
Fase luminosa y fotofosforilación Fase oscura Balance energético Factores que afectan a la fotosíntesis Quimiosíntesis Otras rutas anabólicas

4
Necesidades de los organismos Una fuente de carbono: Heterótrofos
Autótrofos Una fuente de hidrógeno Litótrofos Organotrofos Un aceptor último de hidrógenos Aerobios Anaerobios Una fuente primaria de energía Fotótrofos Quimiótrofos Agua y sales minerales

5
Pigmentos fotosintéticos
Contienen un cromóforo Clorofilas Clorofila a (E. luminosa a E. química) Clorofila b (plantas) Clorofila C (diatomeas y protozoos) Carotenos Xantofilas Ficobilinas En algas rojas (eucariotas ) y algas verde-azuladas (procariotas)
 Clorofila b (plantas) Clorofila C (diatomeas y protozoos) Carotenos. Xantofilas. Ficobilinas. En algas rojas (eucariotas ) y algas verde-azuladas (procariotas)")
6
Clorofila Dos regiones: Anillo de porfirina con Mg Fitol
Los dobles enlaces facilitan el transporte de electrones. La absorción de luz provoca la redistribución electrónica y la pérdida de un electrón hacia otra molécula (un aceptor de electrones)
")
7
La clorofila y los pigmentos accesorios absorben luz de distintas longitudes de onda.

8
No toda esa luz es útil para la planta.
En general la clorofila se especializa en absorber las longitudes de ondas que forman el violeta, el azul y el rojo. El resto es transmitida y reflejada.

10
Cuando una molécula de clorofila absorbe un fotón, pasa a un estado inestable de mayor energía, denominado estado excitado, en el que un electrón periférico se desplaza hacia una posición más externa. Si este electrón pasa a otra molécula (fotooxidación), la energía se habrá transmitido y la molécula de clorofila permanecerá excitada. Para volver a su estado fundamental deberá recibir otro electrón que ocupe el hueco dejado por el primero
, la energía se habrá transmitido y la molécula de clorofila permanecerá excitada. Para volver a su estado fundamental deberá recibir otro electrón que ocupe el hueco dejado por el primero.")
11
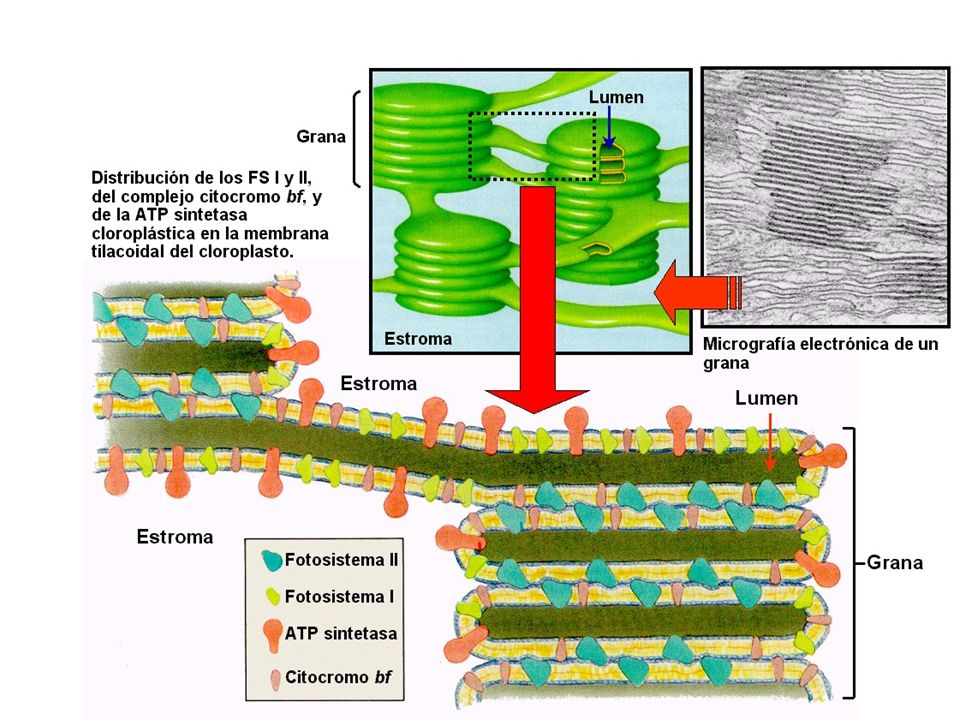
La clorofila esta en el interior de los cloroplastos, en unas membranas especializadas, los tilacoides, junto a otros pigmentos. Los tilacoides, normalmente, presentan un aspecto de sacos o vesículas aplanadas.

12
Fotosistemas Los pigmentos están estrechamente asociados a proteínas y se alojan en la bicapa lipídica de los tilacoides. Estos complejos proteína-clorofila se encuentran empaquetados formando unidades denominadas fotosistemas. Cada unidad contiene de 200 a 400 moléculas de pigmento que captan la luz y forman el llamado complejo antena. Cuando la energía de la luz se absorbe por uno de los pigmentos de la antena, pasa de una molécula a otra de pigmento del fotosistema hasta que alcanza una forma especial de clorofila a que constituye el centro de reacción del fotosistema.

13
Hay dos tipos de fotosistemas:
Fotosistema I (PS I) Fotosistema II (PS II)
 Fotosistema II (PS II)")
15
En el fotosistema I (PS I) la molécula reactiva de clorofila a se denomina P700 (máximo de absorción a 700 nm). Se localiza, casi exclusivamente, en las lamelas estromales y en la periferia de los grana. Se asocia con la reducción del NADP+

16
El Fotosistema II (PS II) también contiene una molécula de clorofila a reactiva, denominada P680, que absorbe preferentemente a 680 nm. Acepta electrones del agua y por ello se asocia con el desprendimiento del oxígeno. Se localiza, preferentemente, en los grana. Los dos fotosistemas se encuentran espacialmente separados en las membranas tilacoidales. Ambos fotosistemas, al ser excitados por la luz ceden e- a un aceptor primario, quedando ellos oxidados (necesitan recuperar los e- cedidos)
")
17
Relación entre ambos fotosistemas

18
Tiene dos fases Generalidades de la fotosíntesis Fase luminosa
Fase oscura Membrana de los tilacoides NADP+ NADPH Fotofosforilación (ATP) Estroma Fijación del CO2 Obtención de biomoléculas Gasto de ATP y NADPH
 Estroma. Fijación del CO2. Obtención de biomoléculas. Gasto de ATP y NADPH.")
19
Fase luminosa Ocurren en las membranas de los tilacoides:
La clorofila y otras moléculas de pigmento absorben la energía de luz. Aumenta la energía de los electrones en las moléculas de los pigmentos activándolos (nivel de energía más alto). Los electrones regresan a un nivel de energía más bajo al pasar por una cadena de transporte de electrones, en forma muy parecida a lo que ocurre en la respiración celular. En el proceso de liberación de energía de los electrones, se produce ATP que se utiliza en las reacciones de la fase oscura.
. Los electrones regresan a un nivel de energía más bajo al pasar por una cadena de transporte de electrones, en forma muy parecida a lo que ocurre en la respiración celular. En el proceso de liberación de energía de los electrones, se produce ATP que se utiliza en las reacciones de la fase oscura.")
20
Transporte de electrones
Los dos fotosistemas se activan simultáneamente con la luz Los electrones activados van pasando por una cadena de moléculas transportadoras. En el caso del PSI llegan hasta el complejo NADP reductasa En el caso del PSII, llegan hasta el PSI Durante este paso de electrones, se va liberando energía, que se aprovecha para bombear protones (en el complejo de citocromos) al lumen tilacoidal, creando en este espacio un fuerte gradiente de protones (fuerza protomotriz)
 al lumen tilacoidal, creando en este espacio un fuerte gradiente de protones (fuerza protomotriz)")
21
Fdx Feof. PQ Cit H+ H+ PC. Luz H2O 2e- 2H+ NADP+ NADPH 2e- 2e- Fot.I
Fot.II P680 * NADP reductasa Feof. NADP+ NADPH PQ Luz Potencial Redox 2e- Cit 2e- H+ H2O PC. H+ 2e- Fot.I P700 Fot.II P680 2H+ 1/2 O2 2 Fotones de luz 2 Fotones de luz

22
Fotofosforilación no cíclica
Intervienen los dos fotosistemas Se reduce el NADP Se rompe el H2O: se libera O2. Sí se sintetiza ATP

23
Fotofosforilación no cíclica
El ATP se genera de manera similar al proceso de formación en las mitocondrias. H+ Estroma Existen complejos ATP sintetasas semenjantes a los de las mitocondrias. El flujo de H+ por el complejo enzimatico sirve para catalizar la formación de ATP ATP ADP + Pi Membrana tilacoidal Lumen tilacoidal H+ H+ H+ H+ H+

24
Fotofosforilación cíclica
Sólo interviene el fotosistema I. No se reduce el NADP No se rompe el H2O: no se libera O2. Sí se sintetiza ATP. Se activa cuando hay desequilibrio entre ATP y NADPH.

25
Fase oscura: Ciclo de Calvin
El proceso de conoce también como vía C3 Supone la reducción del carbono del CO2 para formar glucosa.. Se produce tanto haya luz o no. Se necesitan 3 moléculas de CO2 para formar cada fosfogliceraldehido La rubisco capta CO2 . Luego la Rubisco carboxila al RuBP y genera ac. Fosfoglicérico (PGA). Con el consumo de ATP y NADPH el PGA se transforma en fosfogliceraldehido. El resto sigue en el ciclo para regenerar Ribulosa bifosfato.
. Con el consumo de ATP y NADPH el PGA se transforma en fosfogliceraldehido. El resto sigue en el ciclo para regenerar Ribulosa bifosfato.")
26
Animación sobre el ciclo de calvin
Animación sobre la fotosíntesis

27
RUBISCO RuBisCO es la forma abreviada con que se designa a la ribulosa-1,5-bisfosfato carboxilasa oxigenasa. Cataliza dos procesos opuestos: La fijación del CO2 (carboxilasa). La fotorrespiración, en la que actúa como oxigenasa del mismo sustrato. Es un enzima compuesto de 8 subunidades. Sólo fija 3 moléculas de CO2 por segundo (muy lenta comparada con otras enzimas). Por esto es tan abundante en los cloroplastos y puede alcanzar el 15% de sus proteínas, y de hecho, es la proteína más abundante en la biosfera.
. La fotorrespiración, en la que actúa como oxigenasa del mismo sustrato. Es un enzima compuesto de 8 subunidades. Sólo fija 3 moléculas de CO2 por segundo (muy lenta comparada con otras enzimas). Por esto es tan abundante en los cloroplastos y puede alcanzar el 15% de sus proteínas, y de hecho, es la proteína más abundante en la biosfera.")
28
Balance energético Para formar una molécula de glucosa (6C) se necesitan fijar 6 CO2 y gastar 18 ATP y 12 NADPH (formados previamente en la fase luminosa)
 se necesitan fijar 6 CO2 y gastar 18 ATP y 12 NADPH (formados previamente en la fase luminosa)")
29
Fotorrespiración Es la actuación de la Rubisco como oxigenasa.
Se produce cuando la [CO2] es baja y la de [O2] alta. Produce la rotura de la ribulosa (por oxigenación) en dos moléculas de 2 y 3 carbonos. Finalmente se desprende CO2 Este proceso ocurre durante el día, captura O2 y desprende CO2 , pero no hay fosforilación oxidativa Es un proceso donde la energía se pierde, y no se produce ni ATP ni NADH En la fotorrespiración, después de varios pasos que implican a los cloroplastos, peroxisomas y mitocondrias, hay liberación de CO2 y la formación de algunos aminoácidos
 en dos moléculas de 2 y 3 carbonos. Finalmente se desprende CO2. Este proceso ocurre durante el día, captura O2 y desprende CO2 , pero no hay fosforilación oxidativa. Es un proceso donde la energía se pierde, y no se produce ni ATP ni NADH. En la fotorrespiración, después de varios pasos que implican a los cloroplastos, peroxisomas y mitocondrias, hay liberación de CO2 y la formación de algunos aminoácidos.")
30
Tomado de http://www.euita.upv.es

31
Las condiciones que conducen a la fotorrespiración son bastante comunes.
El CO2 no siempre se encuentra disponible para las células fotosintéticas de la planta. Entra en la hoja por los estomas, que se abren y se cierran, dependiendo, entre otros factores de la cantidad de agua. Cuando la planta está sometida a unas condiciones calurosas y secas, debe cerrar sus estomas para evitar la pérdida de agua. Esto provoca también una disminución del CO2 y permite que el oxígeno producido en la fotosíntesis se acumule. También, cuando las plantas crecen muy juntas y el aire está muy calmado, el intercambio de gases entre el aire que rodea la hoja y la atmósfera global puede ser muy reducido. En estas condiciones, el aire cercano a las hojas de la planta activa tendrá concentraciones de CO2 demasiado pequeñas para sus actividades fotosintéticas. Incluso si los estomas están abiertos, el gradiente de concentración entre el exterior de la hoja y el interior será tan poco importante, que muy poco CO2 se podrá difundir hacia la hoja. La combinación de concentraciones bajas de CO2 y altas concentraciones de oxígeno conduce a la fotorrespiración.

32
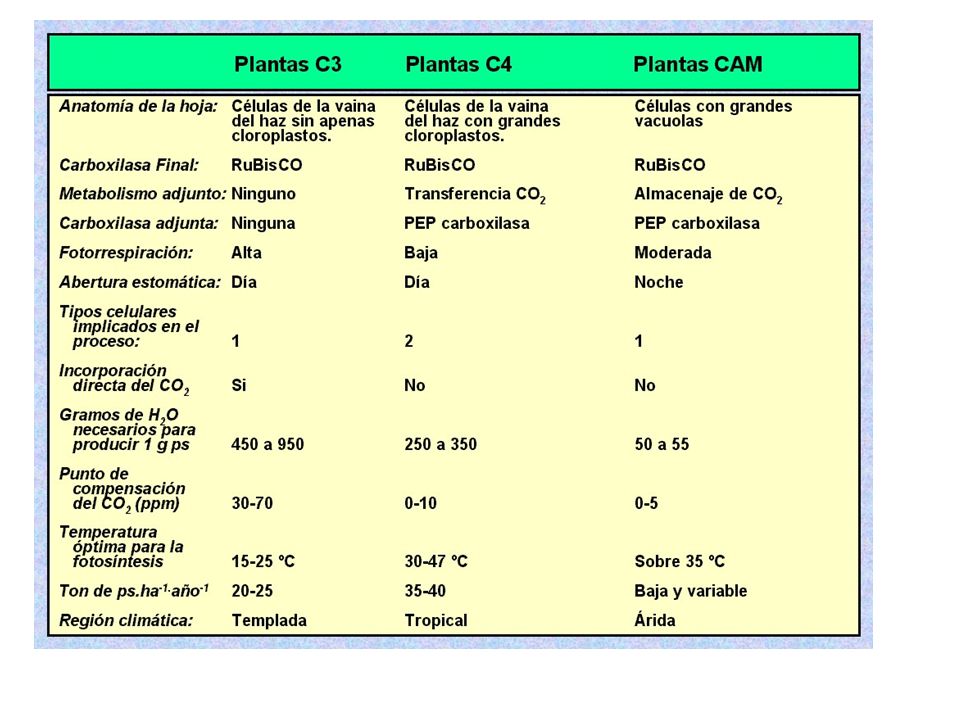
Las plantas C4 Las plantas C4 presentan una anatomía foliar peculiar, conocida como anatomía de tipo Kranz o en corona. En el corte transversal de estas hojas se observan dos tipos de células fotosintéticas: unas grandes, que rodean a los haces conductores (a modo de “corona”) formando una vaina Otras que ocupan el mesófilo, menores y dispuestas por lo general más o menos radialmente alrededor de la vaina.
 formando una vaina. Otras que ocupan el mesófilo, menores y dispuestas por lo general más o menos radialmente alrededor de la vaina.")
33
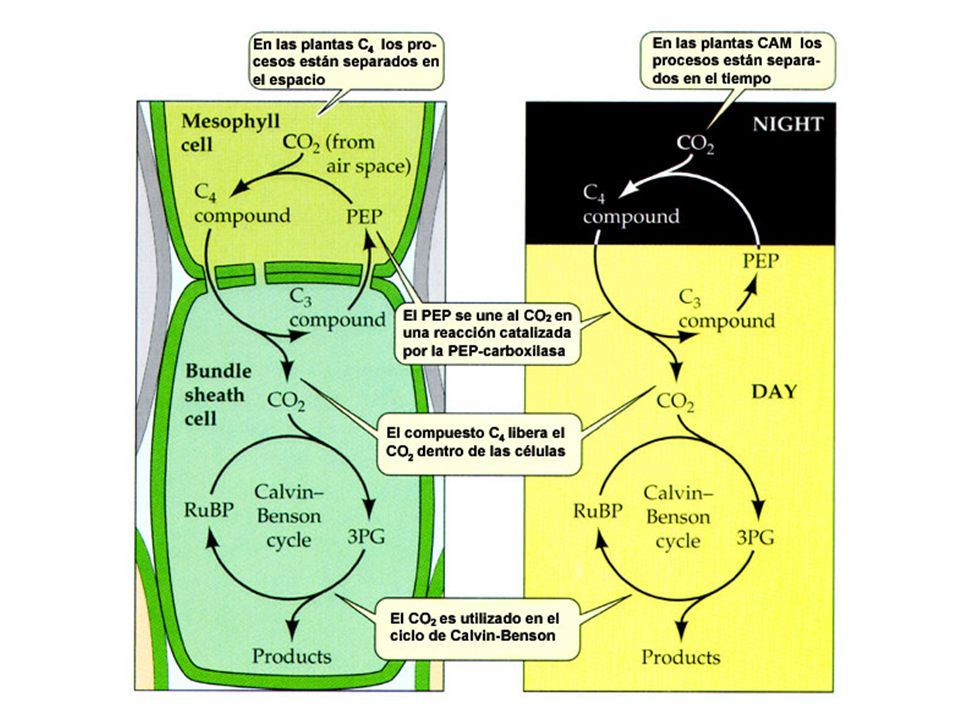
Vía C4 Es un proceso de fijación del CO2 atmosférico, captado en las células del mesófilo, pero en vez de ir al ciclo de Calvin, el CO2 reacciona con el PEP (Fosfoenolpiruvato). El producto final entre el PEP y el CO2 es el ácido oxalacético, que luego se convierte en malato. El malato es llevado a las células de la vaina, en donde es descarboxilado, produciendo el CO2 necesario para el ciclo de Calvin, además de ácido pirúvico. Este último es enviado nuevamente al mesófilo en donde es transformado por medio de ATP en fosfoenolpiruvato (PEP), para quedar nuevamente disponible para el ciclo.
. El producto final entre el PEP y el CO2 es el ácido oxalacético, que luego se convierte en malato. El malato es llevado a las células de la vaina, en donde es descarboxilado, produciendo el CO2 necesario para el ciclo de Calvin, además de ácido pirúvico. Este último es enviado nuevamente al mesófilo en donde es transformado por medio de ATP en fosfoenolpiruvato (PEP), para quedar nuevamente disponible para el ciclo.")
34
Ventajas de las Plantas C4
Las ventajas radican en el hecho de que al tener la Rubisco situada en las células de la vaina, se le impide reaccionar con O2 en situaciones en las cuales la concentración de CO2 sea muy baja, por lo cual el CO2 perdido a través de la fotorrespiración se reduce considerablemente. Incluso las moléculas de CO2 expulsadas por la fotorrespiración son reutilizadas a través del PEP, que las captura en el mesófilo para ingresar en el ciclo de Calvin. Las plantas que usan esta vía para la fijación del carbono son denominadas C4, entre las cuales se distinguen el maíz, la caña de azúcar, la invasora Cynodon dactylon (Bermuda grass), el sorgo y el amaranto. La fijación de CO2 por este sistema tiene un mayor coste energético que en las plantas de vía C3, pero se compensa por la casí nula fotorrespiración que presentan. Son especialmente eficientes en condiciones de altas temperaturas y baja humedad relativa (climas tropicales y sutropicales)
, el sorgo y el amaranto. La fijación de CO2 por este sistema tiene un mayor coste energético que en las plantas de vía C3, pero se compensa por la casí nula fotorrespiración que presentan. Son especialmente eficientes en condiciones de altas temperaturas y baja humedad relativa (climas tropicales y sutropicales)")
35
Plantas CAM El Metabolismo Ácido de las Crassulaceae (CAM) es un tipo de metabolismo que se da en plantas y que se descubrió en la familia de las Crassulaceae. El nombre de metabolismo ácido hace referencia a la acumulación de ácidos orgánicos durante la noche por las plantas que poseen este mecanismo de fijación de carbono. Esta vía metabólica es semejante a la vía C4, sin embargo en la vía CAM la separación de las dos carboxilaciones no es espacial, como ocurre en las plantas C4, sino temporal.
 es un tipo de metabolismo que se da en plantas y que se descubrió en la familia de las Crassulaceae. El nombre de metabolismo ácido hace referencia a la acumulación de ácidos orgánicos durante la noche por las plantas que poseen este mecanismo de fijación de carbono. Esta vía metabólica es semejante a la vía C4, sin embargo en la vía CAM la separación de las dos carboxilaciones no es espacial, como ocurre en las plantas C4, sino temporal.")
36
NOCHE Las plantas CAM tienen dos carboxilaciones separadas temporalmente: La primera se da en la noche cuando tienen los estomas abiertos. El CO2 atmosférico se incorpora al fosfoenolpiruvato (PEP) que se transforma en oxalacetato y este en malato, que se almacena durante la noche. DIA Con la luz, los estomas se cierran (pérdida de agua) impidiendo la adquisición de CO2. El ácido málico sale de la vacuola y se descarboxila liberando el CO2, que va al ciclo de Calvin y ácido pirúvico el cual es devuelto al ciclo produciendo nuevamente PEP.
 que se transforma en oxalacetato y este en malato, que se almacena durante la noche. DIA. Con la luz, los estomas se cierran (pérdida de agua) impidiendo la adquisición de CO2. El ácido málico sale de la vacuola y se descarboxila liberando el CO2, que va al ciclo de Calvin y ácido pirúvico el cual es devuelto al ciclo produciendo nuevamente PEP.")
37
Ventajas de las Plantas CAM
La concentración elevada en el interior de CO2 evita la fotorrespiración. Las plantas CAM suelen ser crasas y relegadas a ambientes secos. Su rendimiento total fotosintético es bajo (ya que la absorción de dióxido de carbono está limitado a la cantidad de ácido málico que se puede almacenar en la vacuola) por lo que son malas competidoras con las plantas C3 o C4. El mecanismo CAM le permite a la planta maximizar la eficiencia en el uso de agua, por lo tanto, las CAM tienen una ventaja competitiva en ambientes con poco agua.
 por lo que son malas competidoras con las plantas C3 o C4. El mecanismo CAM le permite a la planta maximizar la eficiencia en el uso de agua, por lo tanto, las CAM tienen una ventaja competitiva en ambientes con poco agua.")
43
Factores que influyen en la fotosíntesis
Concentración de CO2 ambiental Concentración de O2 ambiental Humedad Temperatura Intensidad luminosa Tipo de luz

44
Concentración de CO2 ambiental
La concentración de CO2 en la atmósfera no es optima para la fotosíntesis, en la practica agrícola se utiliza una adición artificial de CO2 gaseoso, bajo condiciones de iluminación constante, para aumentar la tasa fotosintética y con esta el rendimiento en la producción de materias biológicas. Por encima de un determinado valor (el óptimo), el rendimiento fotosintético se estabiliza.
, el rendimiento fotosintético se estabiliza.")
45
Concentración de O2 ambiental
En similares situaciones de intensidad luminosa, las plantas sometidas a una menor concentración de O2 tienen un rendimiento fotosintético más alto (evitan la fotorrespiración) 0.5% de O2 Asimilación CO2 20% de O2 Intensidad de luz
 0.5% de O2. Asimilación CO2. 20% de O2. Intensidad de luz.")
46
Rendimiento fotosintético
Humedad Asimilación CO2 Humedad Si disminuye la humedad, se cierran los estomas, no entra CO2 y disminuye el rendimento. Si aumenta la humedad, se abren los estomas y aumenta el rendimiento Humedad Apertura estomas Entrada de CO2 Rendimiento fotosintético

47
Temperatura La temperatura optima coincide con el optimo de los enzimas encargados de la fotosíntesis. A partir de ese valor, el rendimiento disminuye Asimilación CO2 Temperatura El rendimiento óptimo depende del tipo de planta

48
Intensidad luminosa En general, a mayor intensidad luminosa, mayor actividad fotosintética. Pero, cada especie está adaptada a unos niveles de iluminación óptima, de intensidad variable. Si se superan esos niveles, se llega a la saturación lumínica e, incluso, podrían deteriorarse los pigmentos fotosintéticos. El exceso de luz puede provocar fotoinhibición

49
Tipo de luz La clorofila a y la clorofila b absorben energía lumínica en la región azul y roja del espectro Los carotenos y xantofilas, en la azul Las ficocianinas, en la naranja Las ficoeritrinas, en la verde.

50
Membranas lamelares de bacterias púrpura halófilas
Fotosíntesis anoxigénica Proceso exclusivo de bacterias. No se genera oxígeno Las bacterias no poseen cloroplastos, realizan la fotosíntesis en los mesosomas Solo tienen un fotosistema. El transporte de electrones es cíclico. Los pigmentos fotosintéticos son bacterioclorofila y carotenoides. Solo se produce ATP. El poder reductor cuando es necesario se obtiene por un transporte inverso de electrones (consumo de energía) y es en forma de NADH, no de NADPH El dador de electrones no es H2O sino otros compuestos inorgánicos reducidos, por lo que no liberan O2 Membranas lamelares de bacterias púrpura halófilas
 y es en forma de NADH, no de NADPH. El dador de electrones no es H2O sino otros compuestos inorgánicos reducidos, por lo que no liberan O2. Membranas lamelares de bacterias púrpura halófilas.")
51
Fotofosforilación cíclica durante la fotosíntesis
anoxigénica

52
Microorganismos Fotosintéticos
Producción de O2 Fuente de H Fuente de C Clasificación Algas oxigénica H2O CO2 Fotoautótrofos Cianobacterias Bacterias verdes del S Chlorobiaceae anoxigénica H2, H2S, S2O32- CO2, acetato, butirato Principalmente fotoautótrofos Bacterias púrpura del S Chromatiaceae Bacterias púrpura no del S H2, compuestos orgánicos CO2, compuestos orgánicos Principalmente fotoorganotrofos, pueden ser quimioorganotrofos en oscuridad y sin O2 Bacterias verdes deslizantes Chloroflexaceae Heliobacterias Compuestos orgánicos Fotoorganotrofos

53
Importancia biológica de la fotosíntesis
Conversión de materia inorgánica en orgánica. Base de las cadenas tróficas. Conversión de la energía luminosa en energía química Fijación de CO2 (actualmente combate el incremento de efecto invernadero) Proporciona O2 y en su momento fue el proceso responsable del cambio de la atmósfera primitiva anaerobia y reductora a la actual. De la fotosíntesis depende también la energía almacenada en combustibles fósiles como carbón, petróleo y gas natural. El equilibrio necesario entre seres autótrofos y heterótrofos no sería posible sin la fotosíntesis. O2
 Proporciona O2 y en su momento fue el proceso responsable del cambio de la atmósfera primitiva anaerobia y reductora a la actual. De la fotosíntesis depende también la energía almacenada en combustibles fósiles como carbón, petróleo y gas natural. El equilibrio necesario entre seres autótrofos y heterótrofos no sería posible sin la fotosíntesis. O2.")
54
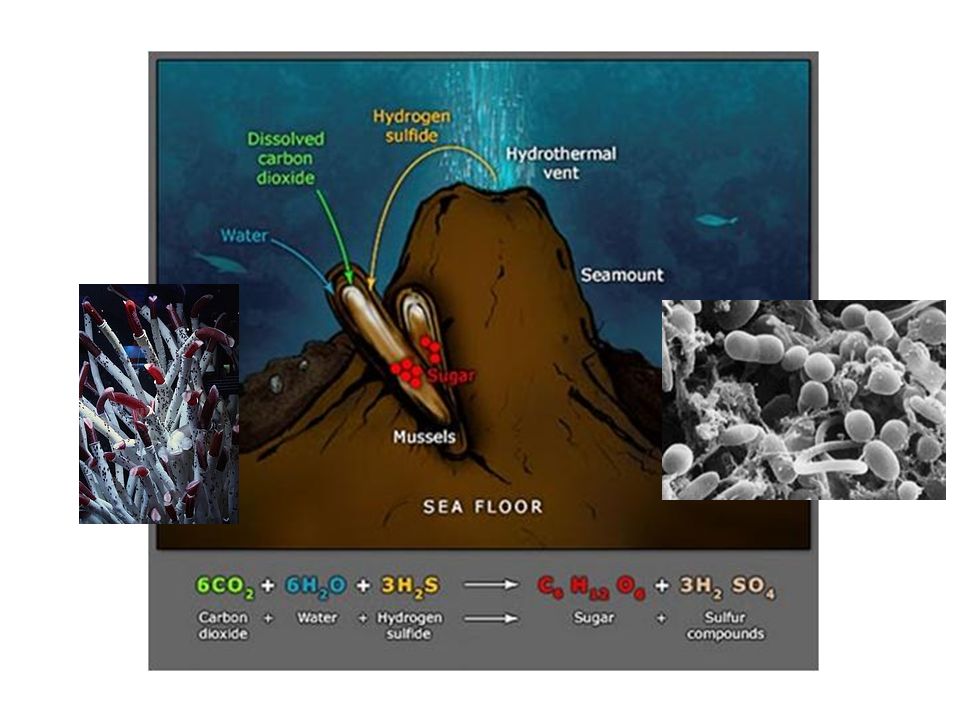
Quimiosíntesis Bacterias del nitrógeno Bacterias del azufre Tipos
Fuente de carbono CO2 ambiental Nutrición autótrofa no fotosintética La energía procede de reacciones de oxidación de sustancias inorgánicas Exclusiva de bacterias Tipos Bacterias del azufre Bacterias del hierro Bacterias del hidrógeno

55
Los organismos quimiosintéticos presentan una serie de características comunes:
Son procariotas autótrofas. Solamente algunas bacterias poseen metabolismo quimiosintético. Viven de una fuente inorgánica: agua, sales, O2, CO2 y compuestos inorgánicos de cuya oxidación obtienen energía. Obtienen la energía de una reacción química específica. Solamente crecen con compuestos específicos de origen inorgánico, o producidos por la actividad de otros organismos (descomposición, excreción). Son aerobios. Utilizan el oxígeno como último aceptor de electrones. Sintetizan materia orgánica por medio del ciclo de Calvin.
. Son aerobios. Utilizan el oxígeno como último aceptor de electrones. Sintetizan materia orgánica por medio del ciclo de Calvin.")
56
Las bacterias quimiolitotrófas tienen un papel crucial en el reciclado del N , C, y S en todo el planeta, puesto que convierten gases y sales sin utilidad para plantas y animales en compuestos orgánicos a su disposición: la conservación de la biosfera depende de su virtuosismo metabólico. Es la cima culminante de la evolución metabólica, “ viven de aire, sales y agua y de una fuente inorgánica de Energía” Este tipo de nutrición autótrofa se da en grupos de bacterias que les permite independizarse del Sol como fuente universal de E. En los ecosistemas marinos, en las zonas afóticas el nivel de productores lo constituyen bacterias quimiosintéticas.

58
Reacciones exergónicas
Fases de la quimiosíntesis Oxidación del sustrato reducido y obtención de la energía: Se obtienen los coenzimas reducidos (NADH+ H+) y el ATP gracias a la energía desprendida en la reacción de oxidación. Fijación del CO2 : Se produce la síntesis de materia orgánica por medio del ciclo de Calvin CO2 y H2O Compuesto reducido Ciclo de Calvin Reacciones exergónicas NADH+H+ ATP Compuesto oxidado Fase I Materia orgánica Fase II
 y el ATP gracias a la energía desprendida en la reacción de oxidación. Fijación del CO2 : Se produce la síntesis de materia orgánica por medio del ciclo de Calvin. CO2 y H2O. Compuesto reducido. Ciclo de Calvin. Reacciones exergónicas. NADH+H+ ATP. Compuesto oxidado. Fase I. Materia orgánica. Fase II.")
59
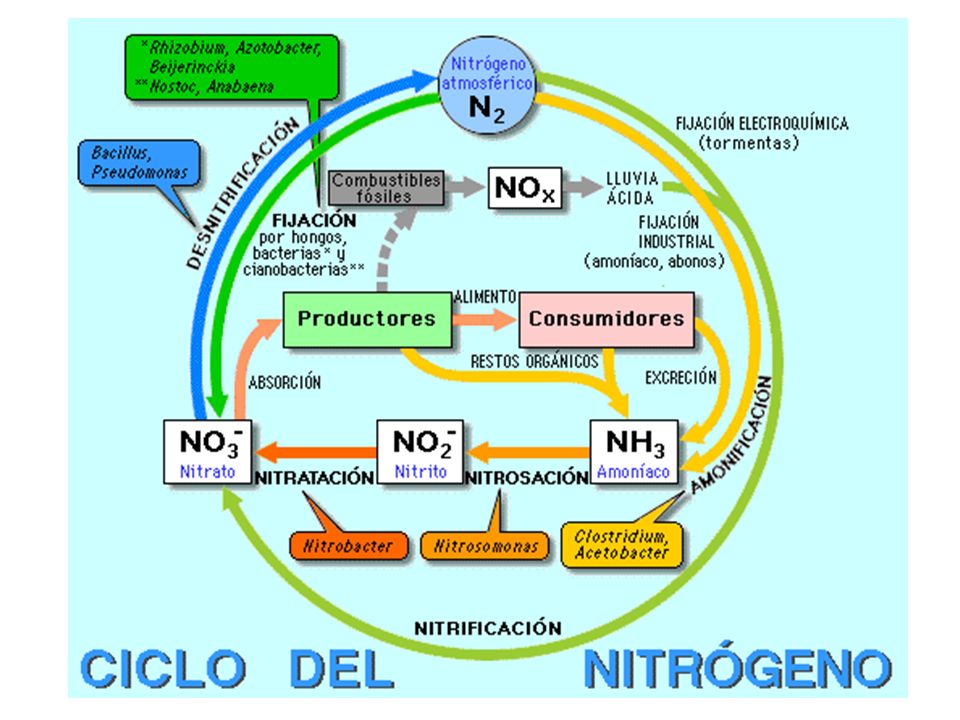
Bacterias del nitrógeno
Presentes en suelos y aguas. Oxidan amoniaco a nitritos y otras especies, los nitritos a nitratos Nitrosomonas 2 NH O2 2 NO2- + 4H+ + 2 H2O Nitrobacter 2 NO2- + O2 2 NO3- Ambos tipos de bacterias se complementan y contribuyen a cerrar el ciclo del nitrógeno

61
Bacterias del azufre H2S + 2 O2 SO42- + 2 H+
Bacterias y tiobacterias sulfurosas. Oxidan compuestos de azufre (S, H2S, S2O32-) hasta ácido sulfúrico, aumentando la acidez del suelo. H2S + 2 O2 SO H+ S2O32- + H2O SO H+
 hasta ácido sulfúrico, aumentando la acidez del suelo. H2S + 2 O2 SO H+ S2O32- + H2O SO H+")
62
Bacterias del Hierro 4 Fe 2+ 4 H+ + O2 4 Fe 3+ + 2 H2O
Bacterias que oxidan compuestos de hierro ferroso a férrico Abundantes en las aguas de minas. 4 Fe 2+ 4 H+ + O2 4 Fe H2O

63
Bacterias del Hidrógeno
Bacterias que oxidan hidrógeno. Pueden utilizar materia orgánica como fuente de carbono además del CO2 (autótrofos facultativos) Formación de ATP y poder reductor en bacterias del H2
 Formación de ATP y poder reductor en bacterias del H2.")
64
Síntesis de aminoácidos
Los aminoácidos son necesarios para la formación de proteínas Sólo los autótrofos son capaces de sintetizar todos los aminoácidos El resto de organismos pueden sintetizar algunos y el resto (aminoácidos esenciales) los tienen que tomar en la dieta. El nitrógeno necesario procede de la fijación de N2 atmosférico (bacterias fijadoras de nitrogeno) o de la asimilación de nitritos, nitratos o amoniaco por las plantas. El esqueleto de carbono de los aminoácidos sintetizados procede del glutámico o de otros intermediarios metabólicos. Nitratos Amoniaco Acido glutámico Resto de aminoácidos Desaminaciones Transaminaciones Reducción Incorporación de α-cetoglutárico
 los tienen que tomar en la dieta. El nitrógeno necesario procede de la fijación de N2 atmosférico (bacterias fijadoras de nitrogeno) o de la asimilación de nitritos, nitratos o amoniaco por las plantas. El esqueleto de carbono de los aminoácidos sintetizados procede del glutámico o de otros intermediarios metabólicos. Nitratos. Amoniaco. Acido glutámico. Resto de aminoácidos. Desaminaciones. Transaminaciones. Reducción. Incorporación de α-cetoglutárico.")
65
Metabolitos del ciclo de Krebs
Gluconeogénesis Ruta anabólica de síntesis de glucosa a partir de precursores no glucídicos. Ácido láctico Aminoácidos Metabolitos del ciclo de Krebs Tiene lugar en el hígado y riñones Participan rutas de la glucólisis mas otras enzimas específicas de la ruta Ácido Láctico gluconeogénesis Aminoácidos GLUCOSA Metabolitos del ciclo de Krebs

66
Gluconeogénesis Se gasta más ATP en producir glucosa a partir de lactato del que obtenemos en su degradación: En la glucólisis obtenemos 2 ATP En la gluconeogénesis se gastan 4 ATP y 2 GTP Es una ruta ventajosa por que evita la acumulación de ácido láctico en los músculos cuando hay poco oxígeno

Presentaciones similares













: Autótrofos: A partir de la materia inorgánica y del CO2 de la atmósfera.>")



: Autótrofos: A partir de la materia inorgánica y del CO2 de la atmósfera.>")





