Descargar la presentación
La descarga está en progreso. Por favor, espere

1
GENETICA DE POBLACIONES agosto 2013
Luis E. Eguiarte Jaime Gasca Erika Aguirre Valeria Souza Laboratorio de Evolución Molecular y Experimental Instituto de Ecología, UNAM, teléfono página web: evolucionmolecular/index.php/es/ cursos-y-talleres/10-genetica-de-poblaciones

2
GENETICA DE POBLACIONES
Introducción al Curso La genética de poblaciones explora los niveles de variación genética dentro y entre las poblaciones que forman a las especies y explica sus patrones en términos de las fuerzas evolutivas. La genética de poblaciones tiene su origen en artículos y libros de varios investigadores en Inglaterra y Estados Unidos en los años 30 del siglo pasado (Fisher, 1930; Wright, 1931, 1932; Haldane, 1932), y ha demostrado ser la herramienta más poderosa para el estudio de la evolución, haciendo central su estudio no sólo para los biólogos interesados en evolución y ecología, sino para cualquier estudioso serio del material genético y la herencia a todos su niveles. Como señala Michael Lynch en su libro del 2007, “Nada en evolución tiene sentido si no es a la luz de la genética de poblaciones”.
, y ha demostrado ser la herramienta más poderosa para el estudio de la evolución, haciendo central su estudio no sólo para los biólogos interesados en evolución y ecología, sino para cualquier estudioso serio del material genético y la herencia a todos su niveles. Como señala Michael Lynch en su libro del 2007, Nada en evolución tiene sentido si no es a la luz de la genética de poblaciones .")
3
GENETICA DE POBLACIONES
Introducción al Curso II La evolución a su vez es el estudio de los patrones y procesos que han producido el cambio de los organismos en el tiempo, y es el resultado de las fuerzas evolutivas operando sobre la variación genética por mucho tiempo. Este proceso evolutivo ha generado tanto la adaptación (el ajuste de los organismos a su medio ambiente), como la gran diversidad de especies que hay y han existido en el pasado en la tierra (millones de especies). La genética de poblaciones inicialmente consideraba explorar, en un solo gen, el comportamiento de dos alelos que segregan de acuerdo a las leyes de Mendel. Este caso se analiza de manera sencilla, y el Equilibrio de Hardy-Weinberg describe cómo se comporta la variación en ausencia de cualquier fuerza evolutiva, funcionado como modelo nulo de cómo se comporta la variación genética si no opera ninguna fuerza evolutiva. De esta manera, modificando los supuestos del Equilibrio de Hardy-Weinberg se exploran los efectos de las diferentes fuerzas evolutivas sobre la variación genética: flujo génico, deriva génica, mutación y selección natural.
, como la gran diversidad de especies que hay y han existido en el pasado en la tierra (millones de especies). La genética de poblaciones inicialmente consideraba explorar, en un solo gen, el comportamiento de dos alelos que segregan de acuerdo a las leyes de Mendel. Este caso se analiza de manera sencilla, y el Equilibrio de Hardy-Weinberg describe cómo se comporta la variación en ausencia de cualquier fuerza evolutiva, funcionado como modelo nulo de cómo se comporta la variación genética si no opera ninguna fuerza evolutiva. De esta manera, modificando los supuestos del Equilibrio de Hardy-Weinberg se exploran los efectos de las diferentes fuerzas evolutivas sobre la variación genética: flujo génico, deriva génica, mutación y selección natural.")
4
GENETICA DE POBLACIONES
Introducción al Curso III ¿Qué pasa al considerar a modelos más realistas (y por lo tanto más complicados) de la genética? Recientemente, con los avances en los métodos moleculares y análisis de secuencias, la disminución de costos y el incremento en la velocidad de secuenciación y los estudios genómicos, por fin se puede analizar cientos o miles de genes en las poblaciones naturales (Genómica de Poblaciones). Estos datos, junto con las herramientas teóricas recientemente desarrolladas, nos permiten reconstruir finamente la historia evolutiva de las poblaciones y las especies, tanto en términos selectivos y demográficos (i.e., teoría de la coalescencia) como geográficos (i.e., la filogeografía) y avanzar en resolver una serie de problemas ecológicos, taxonómicos y evolutivos.
 de la genética Recientemente, con los avances en los métodos moleculares y análisis de secuencias, la disminución de costos y el incremento en la velocidad de secuenciación y los estudios genómicos, por fin se puede analizar cientos o miles de genes en las poblaciones naturales (Genómica de Poblaciones). Estos datos, junto con las herramientas teóricas recientemente desarrolladas, nos permiten reconstruir finamente la historia evolutiva de las poblaciones y las especies, tanto en términos selectivos y demográficos (i.e., teoría de la coalescencia) como geográficos (i.e., la filogeografía) y avanzar en resolver una serie de problemas ecológicos, taxonómicos y evolutivos.")
5
Objetivo del Curso: En este curso pretendemos que los alumnos entiendan y puedan aplicar los principios de la genética de poblaciones, desde sus bases clásicas hasta los desarrollos más recientes mencionados arriba, revisando rigurosamente tanto la teoría como los patrones empíricos

6
Evaluación y estructura I:
Son 9 temas. A cada tema se le dedicará cuando menos 2 clases. Se espera que todos los alumnos lean y hagan un resumen de cada uno de los capítulos del libro de texto que se van a ver en clase. También deberán hacer y entregar los problemas del libro de texto (cuando menos la mitad de cada capítulo). En este curso NO va a haber seminarios. Además se van hacer otros ejercicios, tareas y talleres de computo para aprender a usar los programas relevantes del estudio empírico de la Genética de poblaciones.
. En este curso NO va a haber seminarios. Además se van hacer otros ejercicios, tareas y talleres de computo para aprender a usar los programas relevantes del estudio empírico de la Genética de poblaciones.")
7
Evaluación y estructura II:
La evaluación se realizará con los siguiente elementos: 1) 3 exámenes, 10% c/u, 30% en total. 2) Trabajo semestral (sobre Genética de Poblaciones!), 20%, 3) Resumenes críticos de los capítulos del libro: 15%, tareas (problemas de cada capítulo). 4) Participación y asistencia a clase: 15% 5) Tareas (incluyendo los Problemas de cada capítulo y de los Talleres de computo): 20%; Los alumnos deben de entregar todos los problemas y presentar todos los exámenes y el trabajo semestral para aprobar el curso.
 3 exámenes, 10% c/u, 30% en total. 2) Trabajo semestral (sobre Genética de Poblaciones!), 20%, 3) Resumenes críticos de los capítulos del libro: 15%, tareas (problemas de cada capítulo). 4) Participación y asistencia a clase: 15% 5) Tareas (incluyendo los Problemas de cada capítulo y de los Talleres de computo): 20%; Los alumnos deben de entregar todos los problemas y presentar todos los exámenes y el trabajo semestral para aprobar el curso.")
8
El trabajo semestral consistirá en un buen análisis de datos relacionados a los temas del curso y se demuestre que se aprendió a usar e interpretar estos datos y análisis. Breve (10 cuartillas) pero de calidad, original y creativo, y en formato de un artículo de revista, y se expondrá de manera breve frente al grupo.
 pero de calidad, original y creativo, y en formato de un artículo de revista, y se expondrá de manera breve frente al grupo.")
9
¿Horario y Lugar: Martes y Jueves de 3:30 PM a 5:30 PM. Aquí, Auditorio Instituto de Ecología, UNAM ¿Requisitos?: Conocimientos básicos de genética y de evolución. 16 semanas, 32 sesiones, 64 horas.

10
Referencia y texto básico:
Leer los capítulos y hacer problemas, o pares o nones Hedrick, P.W Genetics of populations. Fourth edition. Jones and Bartlett publishers. Sudbury, Massachusetts. 675 págs.

11
Otras referencias que serán usadas en el curso:
Bell, G Selection. The Mechanism of evolution. Oxford Universiyt press. Oxford, UK., 553 págs. Charelsworth, B. y D. Charelsworth Elements of Evolutionary Genetics. Roberts and Company Publishers. Greenwood Village, Colorado, EUA. 734 págs Eguiarte Luis E., V. Souza y X. Aguirre (Compiladores) Ecología molecular. Semanrant, Conabio, Inst. de Ecología UNAM. D. F., México.574 págs. Libro de texto en español que trata la mayoría de los temas del curso. Felsenstein J Inferring Phylogenies. Sinauer Associates. Sunderland, MA, 664 págs. Gillespie, J.H Population Genetics. A concise guide. Second edition. The John Hopkins Univesity press. Baltimore, 214 págs. Un texto compacto, relativamente económico y actualizado. Hamilton, M Population Genetics. Wiley-Blackwell, Oxford, UK. 424 págs. Hartl D.L. y A. G. Clark Principles of population genetics, 4th edition, Sinauer, Sunderland, Mass. 545 págs. Esta edición es una versión simplificada y ligeramente actualizada de la segunda, pero también recomendamos la segunda por su mayor detalle. Lynch, M The origins of the genome achitecture. Sinauer, Sunderland, Massachusetts. 487 págs.494 págs. Templeton, A.R Population genetics and microevolutionary therory. J. Wiley and sons. N.J., EUA, 705 págs. Wakeley J Coalescent Theory: An Introduction . Roberts & Company Publishers, Colorado, EUA. 432 págs. Texto avanzado sobre la teoría de la coalescencia.
 Ecología molecular. Semanrant, Conabio, Inst. de Ecología UNAM. D. F., México.574 págs. Libro de texto en español que trata la mayoría de los temas del curso. Felsenstein J Inferring Phylogenies. Sinauer Associates. Sunderland, MA, 664 págs. Gillespie, J.H Population Genetics. A concise guide. Second edition. The John Hopkins Univesity press. Baltimore, 214 págs. Un texto compacto, relativamente económico y actualizado. Hamilton, M Population Genetics. Wiley-Blackwell, Oxford, UK. 424 págs. Hartl D.L. y A. G. Clark Principles of population genetics, 4th edition, Sinauer, Sunderland, Mass. 545 págs. Esta edición es una versión simplificada y ligeramente actualizada de la segunda, pero también recomendamos la segunda por su mayor detalle. Lynch, M The origins of the genome achitecture. Sinauer, Sunderland, Massachusetts. 487 págs.494 págs. Templeton, A.R Population genetics and microevolutionary therory. J. Wiley and sons. N.J., EUA, 705 págs. Wakeley J Coalescent Theory: An Introduction . Roberts & Company Publishers, Colorado, EUA. 432 págs. Texto avanzado sobre la teoría de la coalescencia.")
12
PROGRAMA: I. Introducción a la Genética de poblaciones. LEEF y VSS, 3 sesiones. Capítulo 1. a) Objetivos y metas de las genética de poblaciones. b) Historia de la genética de poblaciones y de la evolución molecular. c) Repaso de genética clásica y molecular. II. El estudio de la variación en las poblaciones naturales y la ley del equilibrio de Hardy-Weinberg. LEEF y EAP, 4 sesiones. Capítulo 2. a) Marcadores morfológicos. b) Alozimas/ Isozimas. c) Variación a nivel ADN: métodos moleculares (i.e., RFLPs, PCR, clonación, secuencias, RAPDs, AFLPs, microsatélites, etc.) y patrones. d) La ley del equilibrio de Hardy-Weinberg. e) Complicaciones a Hardy-Weinberg: Diferencias entre sexos, genes ligados al sexo y más de dos alelos. f) El problema de la estimación empírica de las frecuencias alélicas. g) Medidas de variación genética y de distancia genética.
 Historia de la genética de poblaciones y de la evolución molecular. c) Repaso de genética clásica y molecular. II. El estudio de la variación en las poblaciones naturales y la ley del equilibrio de Hardy-Weinberg. LEEF y EAP, 4 sesiones. Capítulo 2. a) Marcadores morfológicos. b) Alozimas/ Isozimas. c) Variación a nivel ADN: métodos moleculares (i.e., RFLPs, PCR, clonación, secuencias, RAPDs, AFLPs, microsatélites, etc.) y patrones. d) La ley del equilibrio de Hardy-Weinberg. e) Complicaciones a Hardy-Weinberg: Diferencias entre sexos, genes ligados al sexo y más de dos alelos. f) El problema de la estimación empírica de las frecuencias alélicas. g) Medidas de variación genética y de distancia genética.")
13
III. La selección natural. LEEF y JGP, 4 sesiones Capíutlo 3
PROGRAMA II: III. La selección natural. LEEF y JGP, 4 sesiones Capíutlo 3 a) Diferentes tipos de selección natural. b) El modelo básico de selección. c) Complicaciones al modelo básico: genes ligados al sexo y alelos múltiples. d) Selección en viabilidad. e) Selección sexual y apareamiento clasificado negativo (negative assortative mating). f) Selección gamética y alelos de incompatibilidad. g) El problema de estimar la intensidad de la selección en el campo. h) Modelos ecológicos, variación espacial y temporal y selección dependiente de la frecuencia.
 Diferentes tipos de selección natural. b) El modelo básico de selección. c) Complicaciones al modelo básico: genes ligados al sexo y alelos múltiples. d) Selección en viabilidad. e) Selección sexual y apareamiento clasificado negativo (negative assortative mating). f) Selección gamética y alelos de incompatibilidad. g) El problema de estimar la intensidad de la selección en el campo. h) Modelos ecológicos, variación espacial y temporal y selección dependiente de la frecuencia.")
14
IV.- La endogamia: JGP y EAP, 3 sesiones. Capítulo 8.
PROGRAMA III: IV.- La endogamia: JGP y EAP, 3 sesiones. Capítulo 8. a) El coeficiente de endogamia y el equilibrio de Hardy-Weinberg. b) Autofertilización total y parcial: teoría y estimaciones. c) Estimación de la endogamia a partir de pedigris. e) La endogamia en las poblaciones naturales. f) La “depresión” por endogamia. e) “Kin selection”. f) Reproducción asexual. V.- La deriva génica, el tamaño efectivo de las poblaciones. LEEF y JGP, 3 sesiones. Capítulo 4 a) ¿Que es la deriva génica? b) Un enfoque de matrices de transición. c) Efecto de fundador y cuellos de botella. d) El tamaño efectivo de las poblaciones, definiciones y métodos ecológicos y genéticos para su estimación. e) Deriva génica y selección natural.
 El coeficiente de endogamia y el equilibrio de Hardy-Weinberg. b) Autofertilización total y parcial: teoría y estimaciones. c) Estimación de la endogamia a partir de pedigris. e) La endogamia en las poblaciones naturales. f) La depresión por endogamia. e) Kin selection . f) Reproducción asexual. V.- La deriva génica, el tamaño efectivo de las poblaciones. LEEF y JGP, 3 sesiones. Capítulo 4. a) ¿Que es la deriva génica b) Un enfoque de matrices de transición. c) Efecto de fundador y cuellos de botella. d) El tamaño efectivo de las poblaciones, definiciones y métodos ecológicos y genéticos para su estimación. e) Deriva génica y selección natural.")
15
PROGRAMA IV: VI. El flujo génico y la estructura de las poblaciones. EAP y LEEF, 3 sesiones, Capítulo 7. a) El modelo contiente-islas de flujo génico. b) El efecto Wahlund. c) Estimaciones directas e indirectas de flujo génico. d) Los estadísticos F de Wright. e) Flujo génico y deriva. f) Flujo génico y selección.
 El modelo contiente-islas de flujo génico. b) El efecto Wahlund. c) Estimaciones directas e indirectas de flujo génico. d) Los estadísticos F de Wright. e) Flujo génico y deriva. f) Flujo génico y selección.")
16
VII. La mutación. VSS y LEEF, 3 sesiones. Capítulo 5
PROGRAMA V: VII. La mutación. VSS y LEEF, 3 sesiones. Capítulo 5 a) Historia natural de la mutación. b) Modelos básicos de mutación. c) Balance selección-mutación. d) Mutación en poblaciones finitas: el modelo de alelos infinitos y el modelo de mutaciones por pasos. e) El problema de la estimación de las tasas de mutación VIII. Modelos de varios genes. LEEF y VSS, 3 sesiones. Capítulo 9 a) El desequilibrio de ligamiento I: teoría básica y métodos de estimación b) El desequilibrio de ligamiento II: relación con las fuerzas evolutivas. c) Selección en varios genes. d) Hitchiking. e) Recombinación, sexualidad, “Muller ratchet”, y selección de fondo.
 Historia natural de la mutación. b) Modelos básicos de mutación. c) Balance selección-mutación. d) Mutación en poblaciones finitas: el modelo de alelos infinitos y el modelo de mutaciones por pasos. e) El problema de la estimación de las tasas de mutación. VIII. Modelos de varios genes. LEEF y VSS, 3 sesiones. Capítulo 9. a) El desequilibrio de ligamiento I: teoría básica y métodos de estimación. b) El desequilibrio de ligamiento II: relación con las fuerzas evolutivas. c) Selección en varios genes. d) Hitchiking. e) Recombinación, sexualidad, Muller ratchet , y selección de fondo.")
17
PROGRAMA VI: IX. Genética de poblaciones molecular. VSS y LEEF, 4 sesiones. Capíutlo 6. a) Estimación de variación genética a nivel molecular. b) El modelo de sitios infinitos. c) La prueba de Tajima y otras pruebas relacionadas. d) La prueba de Ewens-Watterson. e) Pruebas HKA y MK. f) Filogeografía. g) Análisis de paternidad. h) ADN fósil. g) Sesgo en el uso de los codones.
 El modelo de sitios infinitos. c) La prueba de Tajima y otras pruebas relacionadas. d) La prueba de Ewens-Watterson. e) Pruebas HKA y MK. f) Filogeografía. g) Análisis de paternidad. h) ADN fósil. g) Sesgo en el uso de los codones.")
18
Para la próxima semana:
Leer capítulo 1 del Hedrick, 2011. Hacer resumen crítico, en un par de cuartillas. Martes 20 Comenzar a hacer problemas. Pares o nones, para el Jueves 22.

19
Genética de poblaciones:
a) Objetivos y metas de las genética de poblaciones. b) Breve historia de la genética de poblaciones y de la evolución molecular. c) Breve repaso de genética clásica y molecular.
 Objetivos y metas de las genética de poblaciones. b) Breve historia de la genética de poblaciones y de la evolución molecular. c) Breve repaso de genética clásica y molecular.")
20
Material genético: archivo del proceso evolutivo...
a partir de este archivo, con la teoría evolutiva, buenos datos y muestreo y análisis estadístico, podremos reconstruir al proceso evolutivo y entender las fuerzas evolutivas que han modelado la diversidad y adaptaciones de los organismos...

21
Variación genética: El primer paso en todo estudio evolutiva
Si no hay variación, no pueden actuar las fuerzas evolutivas Según el tipo de variación, se va a determinar la evolución adaptativa y/o aleatoria de las poblaciones A partir de como se distribuyen dentro y entre las poblaciones, podemos inferir las fuerzas evolutiva y la historia pasada (esta sería “la gran ilusión”, pero a cada vez parece mas seguro que si se puede lograr!!!)
")
22
La EVOLUCION es la parte central, que da estructura y orden a toda la biología moderna.
La única forma de entender a la evolución es a través de la GENETICA DE POBLACIONES Nada en evolución tiene sentido si no es a la luz de la genética de poblaciones. M. Lynch, 2007

23
Debido a los nuevos métodos, se esta acumulando una cantidad impresionante de secuencias y de información genómica, y la mejor forma de analizarla es usando las herramientas de la genética de poblaciones...

24
GENÉTICA DE POBLACIONES:
(= MICROEVOLUCIÓN) *COMO SE COMPORTA LA VARIACIÓN GENÉTICA EN LAS POBLACIONES (DENTRO DE LAS ESPECIES). *ES EL ESTUDIO DE LAS FUERZAS EVOLUTIVAS QUE CAMBIAN A LAS ESPECIES EN EL TIEMPO. ecol. evol Ecología Evolución Genética de poblaciones genética del desarrollo. ev. molecular ecol. molecular Genética
 *COMO SE COMPORTA LA VARIACIÓN GENÉTICA EN LAS POBLACIONES (DENTRO DE LAS ESPECIES). *ES EL ESTUDIO DE LAS FUERZAS EVOLUTIVAS QUE CAMBIAN A LAS ESPECIES EN EL TIEMPO. ecol. evol. Ecología. Evolución. Genética de. poblaciones. genética del desarrollo. ev. molecular. ecol. molecular. Genética.")
25
GENÉTICA DE POBLACIONES:
Aplicaciones: Entender al proceso evolutivo Herramienta ecológica y taxonómica Análisis de enfermedades humanas y otros organismos. Mejoramiento agronómico y de ganado: bases genética cuantitativa Conservación de las especies Bancos de germoplasma y recursos genéticos plantas y animales Fundamentos de la Evolución Molecular, Bioinformática, Genómica, etc.

26
Empírico: análisis de las frecuencias alélicas en
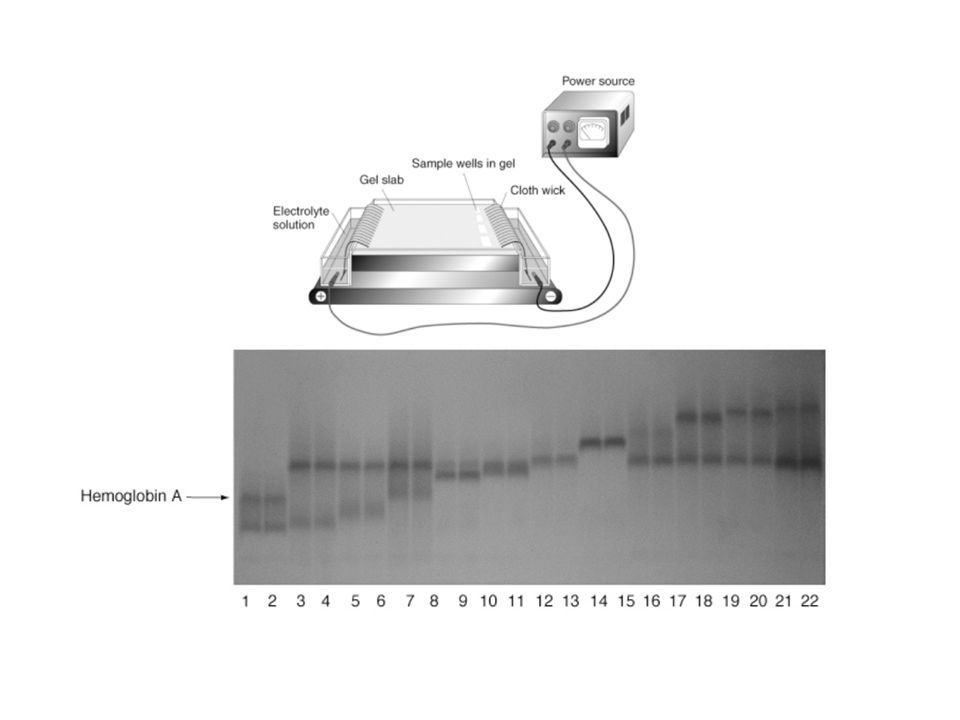
GENÉTICA DE POBLACIONES: Enfoques: Teórico Experimental Empírico Empírico: análisis de las frecuencias alélicas en las poblaciones. Variación genética dentro y entre las poblaciones y en el tiempo. Correlaciones con patrones ambientales. Sugiere problemas e ideas interesantes Datos: morfológicos, grupos sanguíneos, cromosomas (inversiones), alozimas, secuencias ADN, otros moleculares...
, alozimas, secuencias ADN, otros moleculares...")
27
Experimental: apoya a hipótesis generadas de datos empíricos:
Teórico Experimental Empírico Experimental: apoya a hipótesis generadas de datos empíricos: transplantes recíprocos (efectos ambientales, adaptación local) Generaciones múltiples: drosofilas, bacterias, virus, Arabidopsis
 Generaciones múltiples: drosofilas, bacterias, virus, Arabidopsis.")
28
Experimental Empírico Teórico
Teórico: Genera modelos adecuados a los datos empíricos y experimentales. Da el marco de referencia para generalizar y entender datos, en términos de las fuerza evolutivas y parámetros relevantes.

29
Breve historia de la genética de poblaciones y de la evolución molecular.
La teoría de la evolución trata de explicar dos características fundamentales de la vida en la tierra: la diversidad en el número de especies y la adaptación.

30
En general podemos resumir las ideas originales de Charles Darwin en tres grandes puntos:
1) Los organismos actuales son resultado de un proceso evolutivo, y todos están relacionados entre si en un gran árbol filogenético. 2) La evolución es gradual. 3) La evolución es resultado del proceso de selección natural. Los dos primeros puntos fueron rápidamente aceptados por la comunidad científica, pero el tercero fue mucho más controversial.
 Los organismos actuales son resultado de un proceso evolutivo, y todos están relacionados entre si en un gran árbol filogenético. 2) La evolución es gradual. 3) La evolución es resultado del proceso. de selección natural. Los dos primeros puntos fueron rápidamente. aceptados por la comunidad científica, pero el tercero fue mucho más controversial.")
31
Proceso de selección natural según Darwin y Wallace en 4 pasos:
a) Existe variación en las poblaciones naturales, b) Parte de esta variación tiene que ver con que el funcionamiento los organismos y es heredable, c) Dado que los recursos son limitados, se genera competencia entre los individuos de cada especie, y d) Como consecuencia automática de los tres puntos anteriores, tenemos un proceso de selección natural, donde los organismos van cambiando y de esta forma produce le proceso de la adaptación. El proceso es Ecológico (Poblacional)...
 Existe variación en las poblaciones naturales, b) Parte de esta variación tiene que ver con que el funcionamiento los organismos y es heredable, c) Dado que los recursos son limitados, se genera competencia entre los individuos de cada especie, y. d) Como consecuencia automática de los tres puntos anteriores, tenemos un proceso de selección natural, donde los organismos van cambiando y de esta forma produce le proceso de la adaptación. El proceso es Ecológico (Poblacional)...")
32
Para Darwin entonces, proceso evolutivo es Ecológico (POBLACIONAL)...
Este proceso gradual de selección natural eventualmente genera divergencia (por competencia) entre las poblaciones y así se puede explicar tanto la gran diversidad actual como la gran adaptación de las especies.
 entre las poblaciones. y así se puede explicar. tanto la gran diversidad actual como la. gran adaptación de las especies.")
33
Darwin: ESTAN LAS IDEAS DE POBLACIONES, PERO FALTA LA GENETICA Y EL DESARROLLO CUANTITATIVO!
El principal problema del modelo Darwiniano original era la falta de conocimiento de la herencia. Generalmente se pensaba en modelos estilo “mezcla de sangre”, aunque Darwin eventualmente propone un modelo totalmente hipotético de las gémulas para evitar que se perdiera la variación, y no se descartaba la posibilidad de la herencia de los caracteres adquiridos (herencia “lamarquiana”).
.")
34
tenía la clave de la herencia:
Gregor Mendel ( ) tenía la clave de la herencia: diploides caracteres discretos dominantes/ recesivos AA Aa aa
 tenía la clave de la herencia: diploides. caracteres discretos. dominantes/ recesivos. AA Aa aa.")
35
Los Darwinianos originales eran Naturalistas como Darwin, y aceptaban la idea de cambio, de evolución, e interpretaban, usualmente sin experimentos ni análisis estadísticos, que las características de los seres vivos eran resultado de la selección natural y por lo tanto adaptaciones Ernst Haeckel: filogenias “imaginarias”

36
En 1900 se redescubre la genética y las ciencias experimentales
comienzan a adquirir un papel fundamental dentro de la biología. Los genetistas desde el principio dudaron sobre la relevancia de la selección natural, y propusieron que la evolución sucedía gracias a grandes mutaciones que generaban nuevos tipos de organismos y atacan la forma de hacer ciencia de los naturalistas (que se convierten en Ecólogos y Taxónomos), tensión que sigue hasta nuestros días. .
, tensión que sigue hasta nuestros días. .")
37
Hugo de Vries redescubridor de las leyes de Mendel con Oenothera lamarckiana .

38
La biología evolutiva moderna tiene
cuando menos 2 orígenes “sintéticos”. 1) La síntesis original entre las ideas de Mendel y Darwin que elaboraron de manera más o menos independiente, y sin ninguna colaboración relevante con otros científicos Ronald A. Fisher, Sewall Wright y J.B.S. Haldane: origen de la genética de poblaciones (teórica).
 La síntesis original entre las ideas de Mendel y Darwin que elaboraron de manera más o menos independiente, y sin ninguna colaboración relevante con otros científicos. Ronald A. Fisher, Sewall Wright y J.B.S. Haldane: origen de la genética de poblaciones (teórica).")
39
Inicia la genética de poblaciones empírica y experimental.
La biología evolutiva moderna tiene cuando menos 2 orígenes “sintéticos”. 2) Posteriormente viene la segunda fase, en la que T. Dobzhansky. Inicia la genética de poblaciones empírica y experimental. Esta etapa llamaron (él, Mayr, Simpson, Stebbins, Huxley) la “síntesis moderna”: básicamente, popularizar, ampliar y adaptar las ideas de Fisher Wright y Haldane al estudio de diferentes grupos de organismos y subdisciplinas dentro de la biología.
 Posteriormente viene la segunda fase, en la que T. Dobzhansky. Inicia la genética de poblaciones empírica. y experimental. Esta etapa llamaron (él, Mayr, Simpson, Stebbins, Huxley) la síntesis moderna : básicamente, popularizar, ampliar y adaptar las ideas de Fisher Wright y Haldane al estudio de diferentes grupos de organismos y subdisciplinas. dentro de la biología.")
40
RA Fisher (1922-1930) JBS Haldane (1924-1932) S Wright (1931- 1932)
Fundadores de la Genética de Poblaciones Teórica: RA Fisher ( ) JBS Haldane ( ) S Wright ( ) desarrollan de manera impresionante e independiente la TEORIA, el “paradigma” y desarrollan los modelos y conceptos fundamentales ... pero casi sin datos hasta
 JBS Haldane ( ) S Wright ( ) desarrollan de manera impresionante e independiente la TEORIA, el paradigma y desarrollan los modelos y conceptos fundamentales ... pero casi sin datos. hasta")
41
Genética de Poblaciones Clásica:
Trata del comportamiento de genes mendelianos en un contexto poblacional. El modelo básico es el de un locus con dos alelos: A y a

42
Las frecuencias alélicas p y q varían dentro y entre poblaciones
p frecuencia alelo A q frecuencia del alelo a . ESPECIE poblaciones Aa AA aa

43
Frecuencias genotípicas esperadas
Homócigos AA = pxp= p2 Heterócigos Aa = pq +qp= 2pq Homógcigos aa= q x q=q2 . Aa AA aa

44
Evolución: Cambio de las frecuencias alélicas en el tiempo
(T. Dobzhansky, 1941) Equilibro de Hardy-Weinberg: En una población ideal (grande, de reproducción sexual, diploide, y de apareamiento aleatorio entre individuos), las frecuencia alélicas p y q no cambian N individuos Tiempo 0 Frecuencia de A= p0 N individuos Tiempo 1 Frecuencia de A= p1 N individuos Tiempo t Frecuencia de A = pt
 Equilibro de Hardy-Weinberg: En una población ideal (grande, de reproducción sexual, diploide, y de apareamiento aleatorio entre individuos), las frecuencia alélicas p y q no cambian. N individuos. Tiempo 0. Frecuencia de A= p0. N individuos. Tiempo 1. Frecuencia de A= p1. N individuos. Tiempo t. Frecuencia de A = pt.")
45
Selección natural (Natural selection),
Las fuerzas evolutivas violan los supuestos del equilibrio de Hardy-Weinberg y por lo tanto cambian las frecuencias alélicas Deriva génica (Genetic drift), Selección natural (Natural selection), Migración o flujo génico (Migration), Mutación (Mutation), Otros procesos evolutivos no cambian las frecuencias alélicas, pero son importantes: Sistemas reproductivos /endogamia (Mating systems/inbreeding) Recombinación (Recombination)
, Selección natural (Natural selection), Migración o flujo génico (Migration), Mutación (Mutation), Otros procesos evolutivos no cambian las frecuencias. alélicas, pero son importantes: Sistemas reproductivos /endogamia. (Mating systems/inbreeding) Recombinación (Recombination)")
46
Ronald A. Fisher ( ): The Genetical Theory of Natural Selection (1930) La Selección Natural fuerza más importante
: The Genetical Theory of Natural Selection (1930) La Selección Natural fuerza más importante.")
47
Ronald A. Fisher ( ): Interés fundamental en evolución: realizar una síntesis entre las ideas de Darwin y las de Mendel, con una perspectiva estadística. Obra: Cinco libros sobre métodos estadísticos para biólogos y agrónomos, dos sobre evolución y genética, alrededor de 300 artículos científicos sobre problemas estadísticos, genéticos evolutivo y otros, y cientos de reseñas de libros.

48
. En 1928 inició la escritura de su obra seminal, que apareció en 1930: “The genetical theory of natural selection”. Describe como se comporta la variación genética en poblaciones naturales y como opera sobre esta variación las fuerzas evolutivas, con especial énfasis en la selección natural. Analiza el proceso de la adaptación y para extrapolar el futuro de las poblaciones humanas, con una perspectiva eugenista.

49
. Una importante influencia para escribir el libro fueron sus discusiones con Julian Huxley en los 20s, El libro de Fisher es el primero de la síntesis original, y presenta las ideas fundamentales para una síntesis entre Darwinismo y Mendelismo.

50
Para Fisher, la fuerza evolutiva fundamental es la selección natural:
consideraba que la mutación y la deriva génica eran despreciable, ya que para él las poblaciones tenían tamaños poblacionales casi infinitos, en los cuales puede actuar de manera eficiente la selección aunque sea muy débil. Carta a Sewall Wright, 13 de agosto de 1929: “Para los propósitos relevantes, considero que N (el tamaño efectivo de las poblaciones) generalmente debe de ser el total de la población en el planeta, enumerada a al madurez sexual, y al mínimo anual o de otras fluctuaciones periódicas. Para las aves, el doble del número de nidos sería razonable”.
 generalmente debe de ser el total de la población en el planeta, enumerada a al madurez sexual, y al mínimo anual o de otras fluctuaciones periódicas. Para las aves, el doble del número de nidos sería razonable .")
51
Fisher: Gradualismo: las mutaciones que es más probables que sean favorables son las más chicas posibles, ya que si fueran más grandes, seguramente bajarían el funcionamiento del organismo (metáfora del microscopio), oponiéndose así a las ideas de evolución por mutación, que eran las dominantes. “Teorema Fundamental de la Selección Natural”: Niveles de variación en las poblaciones. En los modelos de mezcla de sangre, la variación se perdería en cada generación, el consideraba que las poblaciones eran ricas en variación genética, y que la: “tasa de incremento de un organismo en cualquier tiempo es igual a su varianza genética en la adecuación en ese momento”, o sea que a mayor variación genética no-neutra, el cambio evolutivo es mayor. Otras contribuciones a la ecología: el valor reproductivo, la tasa de adaptación en especies sexuales vs. asexuales, modelos de selección sexual.
, oponiéndose así a las ideas de evolución por mutación, que eran las dominantes. Teorema Fundamental de la Selección Natural : Niveles de variación en las poblaciones. En los modelos de mezcla de sangre, la variación se perdería en cada generación, el consideraba que las poblaciones eran ricas en variación genética, y que la: tasa de incremento de un organismo en cualquier tiempo es igual a su varianza genética en la adecuación en ese momento , o sea que a mayor variación genética no-neutra, el cambio evolutivo es mayor. Otras contribuciones a la ecología: el valor reproductivo, la tasa de adaptación en especies sexuales vs. asexuales, modelos de selección sexual.")
52
J.B.S. Haldane (1882-1964) The Causes of Evolution (1932)
Un punto de vista intermedio: no todo es selección, no todo es deriva génica

53
1921 Cambridge, Readership in Biochemistry, en el Trinity College, para luego inicia en 1924 una serie de 5 artículos titulados: “A mathematical theory of natural and artificial selection”: análisis de las fuerzas evolutivas y desarrollo de la Genética de Poblaciones. En el primer artículo presentó el análisis inicial del de selección natural direccional: el melanismo industrial en Biston betularia. Estos cinco artículos fueron la base conceptual de su libro “The causes of evolution”, publicado en 1932.

54
JBS Haldane, 1957: costo de la selección: para que un alelo sea completamente reemplazado de una población , se requiere que se elimina un número fijo de organismos, entre de 10N a 30N, independientemente de la velocidad a la que se realice el proceso (o sea de los coeficientes de selección). Desarrolla con claridad las ideas de la genética de poblaciones y de la evolución moderna. Revisa las evidencias genéticas. paleontológicas y taxonómicas. Da estimaciones y métodos para estimar los parámetros relevantes como los coeficientes de selección y las tasas de mutación.
. Desarrolla con claridad las ideas de la genética de poblaciones y de la evolución moderna. Revisa las evidencias genéticas. paleontológicas y taxonómicas. Da estimaciones y métodos para estimar los parámetros relevantes como los coeficientes de selección y las tasas de mutación.")
55
Shifting balance (Topografía adaptativa)
Sewall Wright ( ) Evolution in Mendelian populations (1931) Papel fundamental de la Deriva Génica Shifting balance (Topografía adaptativa)
 Evolution in Mendelian populations (1931) Papel fundamental de la Deriva Génica. Shifting balance (Topografía adaptativa)")
56
Contribuciones evolutivas iniciales:
1)Un extenso artículo Genetics, publicado en 1931: Evolution in medelian populations, 2)Un trabajo breve publicado en las memorias del VII congreso Internacional de genética en Ithaca.
Un extenso artículo Genetics, publicado en 1931: Evolution in medelian populations, 2)Un trabajo breve publicado en las memorias del VII congreso Internacional de genética en Ithaca.")
57
Posteriormente publicó gran cantidad de estudios sobre evolución, varios en colaboración con Dobzhansky, tanto sobre drosofilas como sobre plantas (Oentohera organenesis, Linahthus parryae). En 1950 se retiró de Chicago a la Universidad de Winsconsin en Madison, en donde escribió su obra magna: un tratado en 4 tomos publicados entre 1968 y 1978: Evolution and the genetics of populations y donde continuo activo hasta su muerte en 1988.

58
La principales características de sus ideas sobre evolución es darle un papel muy fuerte al azar, actuando como la deriva génica y endogamia, y su interacción con las otra fuerzas evolutivas dentro de la topografía adaptativa, para genera la evolución por “shifting balances”. W x z SN DG

59
Evolution in mendelian populations. 1931. Genetics 16: 97-159.
Trata del equilibrio de Hardy-Weinberg, y analiza los efectos de la mutación y la migración y la selección básica en las frecuencias alélicas, discutiendo brevemente lo que sucede cuando hay más de dos alelos. Luego introduce los efectos de lo que hoy llamamos deriva génica (Random variation in gene frequencies ) y la importancia del número de individuos en las poblaciones.
 y la importancia del número de. individuos en las poblaciones.")
60
The roles of mutation, inbreeding, crosbreeding, and selection in evolution Proceeding of the Sixth International Congress of Genetics 1: Analiza el papel de cada una de estas fuerza dentro de la topografía adaptativa. Básicamente, la idea es que la selección, al eliminar a los peores genotipos, eleva a las poblaciones en la topografía adaptativa, mientras que la deriva génica, al generar combinaciones aleatorias, bajaría a las poblaciones de los valles adaptativos y les permitiría explorar esta topografía adaptativa. Por último, da ejemplos posible de la evolución por deriva génica y endogamia, en particular los caracoles terrestres de Hawaii de Gulick.

61
La motivación original de Fisher, Wright y Haldane era demostrar que las ideas de Darwin y Wallace eran compatibles con la Genética Mendeliana y analizar a las selección natural de manera cuantitativa como la sobrevivencia y reproducción diferencial

62
La Selección Natural según Darwin y Wallace
Existe variación en las poblaciones naturales Esta variación incluye caracteres relevante para la sobrevivencia y la reproducción Parte de esta variación es heredable. d) Si se cumplen los puntos anteriores, tenemos automáticamente un proceso de Selección Natural que adapta a los organismos
 Si se cumplen los puntos anteriores, tenemos automáticamente un proceso. de Selección Natural. que adapta a los organismos.")
63
fitness (adecuación)= W.
Darwin & Wallace had no idea on the basis of heredity... Fisher, Haldane & Wright show that Natural Selection could be understood considering differences in the fitness (“efficiency”) of the different genotypes: one locus, two alleles: AA, Aa, aa fitness (adecuación)= W. We can predict well the behavior of the alleles in populations in cases with Mendelian inheritance and clear-cut patterns in W.
 of the different genotypes: one locus, two alleles: AA, Aa, aa. fitness (adecuación)= W. We can predict well the behavior of the alleles in populations in cases with Mendelian inheritance and clear-cut patterns in W.")
64
Biston betularia: color, 1 locus, 2 alelos

65
bosque ciudad

66
Selección a favor del recesivo (polillas claras)
")
67
Diferentes tipos de Selección Natural
Selection Viability Fecundity Selection Sexual selection Gametic Selection Frequency-dependant Denso-dependent

68
Modes o kinds of selection:
1) Stabilizing or Balancing (one locus two alleles) Eliminates both tails in the distribution t 2) Directional (Biston betularia) o puryfing (if it elliminates the products of mutation /genetic load) Eliminates one of the tails. t
 Stabilizing or Balancing (one locus two alleles) Eliminates both tails in the distribution. t. 2) Directional (Biston betularia) o puryfing (if it elliminates the products of mutation /genetic load) Eliminates one of the tails. t.")
69
3) Disruptive selection
Eliminates the modal categories, the tails higher W o latter we will see with more care each Mode of selection...

70
Wright: Genetic drift Random changes in the Allelic frequencies, due to the finite sizes of populations... Stronger in smaller populations!

71
Genetic Drift Ne =9 The smaller the Ne, the changes are more
Allelic frequency p 1 0.5 Ne =9 Genetic Drift The smaller the Ne, the changes are more violent and the populations diverge and genetic variation is lost at a faster rate. 10 Generations 20 1 Ne =50 Allelic frequency p 0.5 10 20 Generations

72
Large original population, N
Genetic Drift S. Wright basic model Time 0 Frequency de A1 = p0 Small Population (Size n) 1 9 Time 1 2 8 3 7 4 5 6 Increases the variance in the allelic frequencies among populations (= genetic differentiation) Reduces the genetic diversity within each population, Increase inbreeding, due to the small size of the populations 1.0 p0 Frequency de A1 0.5 0.0 Time
 Time Increases the variance in the allelic frequencies among populations. (= genetic differentiation) Reduces the genetic diversity within each population, Increase inbreeding, due to the small size of the populations p0. Frequency de A Time.")
73
Genetic Drift H= expected heterocigosity, a measure of genetic variation Ht = ( 1- 1/2 Ne) t Ho if Ne =1, the genetic variation decreases by 1/2H in one generation if Ne = infinite, Ht=Ho; no changes with enough time and Ne finite... Ht= 0: there is always some decrease in genetic variation due to drift! (but low if Ne is large)
")
74
Genetic Drift: Differentiation among populations
Vqt = poqo [1-(1-1/2 Ne ) t ] if Ne infinite, Vqt =0, no divergence among the populations if t is very large, Vqt = poqo. The differentiation is faster/ larger if the populations are small (“less than 100 individuals”)
 t ] if Ne infinite, Vqt =0, no divergence among the populations. if t is very large, Vqt = poqo. The differentiation is faster/ larger if the populations are small. ( less than 100 individuals )")
75
Effective population size Ne:
The size of an ideal population that would experience the same effects due to genetic drift than the actual population studied. Generally assumed to be similar to the number of reproductive adults, but it is more complex....

76
m= migration rate, Genetic Flow: probability that a randomly
In population genetics it is considered a synonym of migration. The incorporation of genes into the gene pool of one population from one or more other populations (Futuyma, 1986). m= migration rate, probability that a randomly chosen gene in any subpopulation comes from a migrant.
. m= migration rate, probability that a randomly. chosen gene in any subpopulation comes from. a migrant.")
77
estructura geográfica de las especies.
Modelos de la estructura geográfica de las especies. Violaciones al modelo básico, ayudan a entender el papel de la geografía... a Island model of migration. b Stepping-stone model. c Isolation by distance model. d Metapopulation model. In nature, not only do individuals move between populations, but also individual populations come and go over time (t) with the founding and extinction of entire populations being an important component of population structure
 with the. founding and extinction of entire populations being an important. component of population structure.")
78
A single effective migrant per generation is enough to inhibit
Effective population size Ne is usually studied at the same time that the gene flow: Ne m = number of migrants per generation. If Ne m is large (larger than 1) the different populations behave as a single unit: no divergence due to drift. If Ne m is low (smaller than 1): each population evolves independently by genetic drift. A single effective migrant per generation is enough to inhibit any genetic divergence among population
 the different populations behave as a single unit: no divergence due to drift. If Ne m is low (smaller than 1): each population evolves independently by genetic drift. A single effective migrant per. generation is enough to inhibit. any genetic divergence among. population.")
79
Different indirect methods for the estimate of Nem
A partir de la distribución de las frecuencias alelicas se pueden hacer inferencias con los datos y modelos clásicos: Different indirect methods for the estimate of Nem from genetic variation (island model): a) From Fst = 1/ (4 Ne m +1) b) From the private alleles method of Slatkin (1985): Average frequency of alleles found in only one population log10[p(1)] = a log 10 Ne m + b a and b are constants derived by simulated data Logic: with high gene flow, the rare alleles should be very rare c) Isolation by distance analysis (genetic differentiation as a function of the geographic distance), using estimates of Ne m for pairs of populations (M), following Slatkin (1993)
: a) From Fst = 1/ (4 Ne m +1) b) From the private alleles method of Slatkin (1985): Average frequency of alleles found in only one population. log10[p(1)] = a log 10 Ne m + b. a and b are constants derived by simulated data. Logic: with high gene flow, the rare alleles should be very rare. c) Isolation by distance analysis (genetic differentiation as a function of the geographic distance), using estimates of Ne m for pairs of populations (M), following Slatkin (1993)")
80
Theodosius Dobzhansky (1900-1975):
GENETICA DE POBLACIONES EMPIRICA y EXPERIMENTAL: Theodosius Dobzhansky ( ): 10 años más joven que Fisher, Wright y Haldane, Nació en Meriov, cerca de Kiev, en Ucrania. En 1927 Dobzhansky llegó a Estados Unidos, para realizar una estancia de investigación con Thomas H. Morgan, padre de la genética con drosofilas
: 10 años más joven que Fisher, Wright y Haldane, Nació en Meriov, cerca de Kiev, en Ucrania. En 1927 Dobzhansky llegó a Estados Unidos, para realizar una estancia de investigación con Thomas H. Morgan, padre de la genética con drosofilas.")
81
(Genetics of Natural Populations; 1937)
Dobzhansky decidió utilizar lo aprendido en el laboratorio de Morgan para desarrollar un proyecto de investigación sobre la Genética de Poblaciones en D. pseudoobscura: (Genetics of Natural Populations; 1937) Cromosomas gigantes/ Inversiones y cruzas para revelar variación genética proyecto que inicialmente diseño junto con Stutervant, con el que luego se peleó, y en el que siempre quiso involucrar de manera más activa a Wright.
 Cromosomas gigantes/ Inversiones y cruzas para revelar variación genética. proyecto que inicialmente diseño junto con Stutervant, con el que luego se peleó, y en el que siempre quiso involucrar de manera más activa a Wright.")
82
la estructura genética de las poblaciones”:
De sus experimentos y datos de drosofilas, propone el “modelo balanceado de la estructura genética de las poblaciones”: poblaciones e individuos ricos en variación genética: cada individuo es rico en loci heterócigos, mantenidos por selección balanceadora (ventaja del heterócigo)
")
83
la estructura genética de las poblaciones”:
“Modelo balanceado de la estructura genética de las poblaciones”: individuos ricos en variación genética: cada individuo es rico en loci heterócigos, mantenidos por selección balanceadora (ventaja del heterócigo)
")
84
“modelo clásico de las estructura genética de las poblaciones”:
Lo contrasta con el “modelo clásico de las estructura genética de las poblaciones”: las poblaciones, casi sin variación genética, la mayor parte de las mutaciones deletéreas, que son constantemente eliminada por la selección natural purificadora... Se lo atribuye a H.J. Muller, genetista que descubrió el efecto de los rayos X en generar mutaciones, etc.

85
“modelo clásico de las estructura genética de las poblaciones”:
las poblaciones, casi sin variación genética, la mayor parte de las mutaciones deletéreas, que son constantemente eliminada por la selección natural purificadora... H.J. Muller

86
Dobzhansky: Los resultados de su programa de investigación en drosofila ambiguos: no necesariamente apoyaban al modelo balanceado, ni la SN como fuerza evolutiva dominante... 1966: Richard Lewontin (su alumno) Hubby y Harris: aloenzimas (isoenzimas), geles de almidón: sugieren MUCHA variación genética a nivel molecular ¿mantenida por selección balanceadora?
 Hubby y Harris: aloenzimas (isoenzimas), geles de almidón: sugieren MUCHA. variación. genética. a nivel molecular. ¿mantenida por. selección. balanceadora")
88
Levels of Genetic Variation in Natural Populations
Lewontin y Hubby (1966): average H= 0.12 & P 30% in 18 loci in D. pseudoobscura... similar high levels in most organisms!!! Superficially seem to support the Balanced Model of T. Dobzhansky: neodarwinians happy!!!
: average H= 0.12 & P 30% in 18 loci in D. pseudoobscura... similar high levels in most organisms!!! Superficially seem to support the Balanced Model of T. Dobzhansky: neodarwinians. happy!!!")
91
Expected Heterozygosity Hs Proportion of Polymorphic Loci P
= microsatellite

92
Según Motoo Kimura (1924-1994): NO!!!!
Si hay mucha variación, pero es nula para la selección natural (no la ve, y si hay, es purificadora) Balance entre la Deriva Génica y la Mutación (que hacen que se pierda o se fije la variación genética) Fundamentos Evolución Molecular!! . Kimura es alumno, indirecto, de Wright y de Muller...
 Balance entre la Deriva Génica y la. Mutación (que hacen que se pierda o se fije la variación. genética) Fundamentos Evolución. Molecular!! . Kimura es alumno, indirecto, de Wright y de Muller...")
93
Kimura two observations
But the data were not conclusive (could support any of the models)! 12 years latter, Kimura Genetic Club Meeting Fukuoka, November 1967 (Nature, 1968): Kimura two observations The rate of evolution in proteins is almost constant and high. (Zuckerkandl E. y L. Pauling (1962, 1965) Molecular Clock). b) High levels of genetic variation in proteins (Lewontin y Hubby (1966)) (too high to be maintained only by balancing selection)
! 12 years latter, Kimura Genetic Club Meeting Fukuoka, November 1967 (Nature, 1968): Kimura two observations. The rate of evolution in proteins is almost constant and high. (Zuckerkandl E. y L. Pauling (1962, 1965) Molecular Clock). b) High levels of genetic variation in proteins. (Lewontin y Hubby (1966)) (too high to be. maintained only by balancing selection)")
94
Segregational Load. KIMURA: Genetic Variation.
If all the variation is maintained by Balancing Selection: If there are 2000 polymorphic genes (reasonable, considering a total genome of 13,000 genes and a polymorphism of 30%), and each gene has only and advantage of s=0.01 The total fraction of eliminated individuals should be , each adult should leave a progeny of 22,000 to maintain the population (at constant size). I.e., we should be several times dead to maintained the observed genetic variation (if all due to balancing selection).
, and each gene has only and advantage of s=0.01. The total fraction of eliminated individuals should be , each adult should leave a progeny of 22,000. to maintain the population (at constant size). I.e., we should be several times dead to maintained the observed genetic variation (if all due to balancing selection).")
95
A simple solution: What if the molecular evolution is controlled by a balance between mutation and drift? I.e., extend Wright´s ideas to the Molecular World (no only Mendel´s)
")
96
W K

97
Crítica: But Kimura neutraL theory does not work for all molecular data. It should be considered as the NULL MODEL, that describe the evolutionary behavior in terms of population genetics when there is no selection Is the corner stone of the study of Molecular Evolution But the really interesting things happen when it is not followed by our data! Heterogeneity in substitution rates among lineages, in time and among In general we have less genetic variation than expected by IAM There are ample evidences and signals in the sequence that indicates different kinds do selection: purifying, balancing and directional!

98
In species /population with a large Ne we have far less genetic variation than the predicted by Infinite Alleles Model o sea... Kimura no es todo! Predicted Observed!!

99
Estas son las ideas básicas del
curso... ¡Vamos a explorarlas con más cuidado en el resto de las clases!

100
GENETICA DE POBLACIONES
Pero en los años 60s, la revolución de la genética molecular alcanza a ésta área de la biología. Paralelamente, por un lado se comienzan a aplicar los métodos de electroforesis en proteínas, que revelan una riqueza extraordinaria de variación genética. Por el otro lado se comienzan a acumular los primeros datos de evolución molecular, en la forma de secuencias de aminoácidos para algunas proteínas. Los datos de electroforesis de proteínas se podían interpretar usando la extraordinaria herramienta analítica derivadas de la teoría de Fisher, Wright y Haldane, y permitieron analizar la estructura genética para muchos genes en gran cantidad de individuos para cualquier especie de organismo.

101
Sin embargo, pronto resultó claro que ésta información y la obtenida en los estudios de evolución molecular no se ajustaban a los modelos o paradigmas de evolución propuesta hasta ese momento, y obligó a M. Kimura en 1968 a desarrollar la “Teoría neutra de la evolución molecular”, que propone que esta variación genética se comporta, básicamente, de manera independiente de la adaptación (era selectivamente “neutra”), y que la variación es generada por la mutación que se pierde o conserva simplemente por procesos aleatorios conocidos como deriva génica y permitía interpretar a los datos de evolución molecular desde una perspectiva de genética de poblaciones.
, y que la variación es generada por la mutación que se pierde o conserva simplemente por procesos aleatorios conocidos como deriva génica y permitía interpretar a los datos de evolución molecular desde una perspectiva de genética de poblaciones..")
102
Aunque las propuestas de Kimura fueron inicialmente muy mal recibidas por la comunidad científica, eventualmente se ha reconocido que la mayor parte del material genético se comporta de esta manera. Esto es una ventaja, ya que simplifica los análisis e implica que el material genético funciona como un archivo, un registro viviente del pasado evolutivo. Así, a partir de los años 60 hemos visto el desarrollo de nuevas herramientas moleculares que permiten analizar directamente la variación genética a nivel ADN y el desarrollo de nuevos análisis basados en el comportamiento más fino del material genético en términos de la genética de poblaciones.

103
Con el desarrollo del PCR, de protocolos moleculares sencillos y económicos y de los secuenciadores automáticos, se ha logrado abatir los costos y de esta forma se puede tener gran cantidad de información genética detallada para cualquier organismo. Al mismo tiempo, recientemente se han desarrollado métodos genómicos de secuenciación masiva y la teoría de la coalescencia, que representan una nueva revolución dentro de la genética de poblaciones.

104
VI. El flujo génico y la estructura de las poblaciones.
VII. La mutación. VIII. Modelos de varios genes IX. Genética de poblaciones molecular.

Presentaciones similares