Descargar la presentación
La descarga está en progreso. Por favor, espere

1
A) Complicaciones: 1. Varios alelos 2.- Genes ligados al sexo
SELECCION NATURAL 2 (avanzada ) Curso Evolución L. Eguiarte et al. A) Complicaciones: 1. Varios alelos 2.- Genes ligados al sexo B) Estimación de la Selección:
 Curso Evolución. L. Eguiarte et al. A) Complicaciones: 1. Varios alelos. 2.- Genes ligados al sexo. B) Estimación de la. Selección:")
2
lx en lo demás es igual que H.-W.

3
equilibrio balanceadora: más difícil!
A) Complicando las suposiciones del modelo: 1.-Alelos múltiples (más de 2 alelos) equilibrio balanceadora: más difícil!
 Complicando las suposiciones del. modelo: 1.-Alelos múltiples (más de 2 alelos) equilibrio balanceadora: más difícil!")
4
Alelos múltiples (más de 2), modelo general
adecuación asociada a i, menos el promedio para que cambie, diferente de 0 si menor que promedio pi decrece

5
adecuación i menos promedio dif. de 0 adecuación promedio n alelos
sumatoria de TODAS las w donde aparece i, por la f. alélica del otro alelo adecuación promedio n alelos

6
el alelo C funciona no tan mal de forma homóciga
Alelos múltiples (más de 2) Casos alelos multiples... el alelo C funciona no tan mal de forma homóciga
 Casos alelos multiples... el alelo C funciona no tan mal de forma homóciga.")
7
recordemos la SN direccional con 2 alelos
SELECCION DIRECCIONAL N-ALELOS recordemos la SN direccional con 2 alelos Selección vs. recesivo Sel. vs. dominante se va perdiendo el alelo desfavorable...

8
sAA = sAa = 0.153 saa= 0 26 años, 2 generaciones por año

9
un caso particular direccional 3 alelos
A1 melánica negra dominante A2 insularia gris recesiva a todos A3 típica, blanca recesiva a A1, dominante a A2

10
típica (clara) 0.79, insularia 0.2, entra melánica 0.01
 0.79, insularia 0.2, entra melánica 0.01")
11
A3claras A1 A2 se pierde rápido típica (clara), se acaba sel. obscuras
melánicas decrece A2 A2 insularias se pierde rápido típica (clara), se acaba sel.
, se acaba sel.")
12
Ventaja del heterócigo en n-alelos
¿en que condiciones hay equilibrio? en el caso de 2 alelos bastaba tener wAA<wAa>waa y que p y q iniciales fueran dif. de 0

13
Ventaja del Heterócigo saa = 2sAA

14
Ventaja del heteróciogo ¿en que condiciones hay equilibrio si dos alelos?
si en ambos la dif. contra el promedio es 0, tenemos el equilibrio

15
2 alelos usando estas w´s w1= 1xq+(1-s1)xp w1=1-p+p-s1p w1=1-s1p
xp w1=1-p+p-s1p w1=1-s1p")
16
2 alelos cont. adecuaciones promedio en el eq. 2 alelos

17
Expansión n-alelos equilibrios
pero más complicado

18
el equilibrio para 3-alelos se calcula de 3 juegos de 3 ecuaciones

19
En el equilibrio de tres alelos x y z
deben ser mayores que 0

20
las frecuencia en el equilibrio para los tres alelos están dadas por la z´s

21
La condición general para el
polimorfismo con 3 alelos: la viabilidad promedio de los heterócigos> viablidad prom. homócigos

22
Ejemplo anemia otros tres casos particulares de eq. ...

23
Ventaja del Heterócigo:

24
Ventaja del Heterócigo: anemia falciforme

25
Ventaja del Heterócigo:

26
Pero la realidad más complicada:
hay tres alelos, el C

29
w un poco más alta (0.897) que con dos alelos (0.878)
frecuencias en el equilibrio w un poco más alta (0.897) que con dos alelos (0.878)
 que con dos. alelos (0.878)")
30
Dinámica: si no hay C o si en muy baja frec.
el equilibrio S y A si no hay C si C en baja freq. inicial, mucho tiempo...

31
C tarda mucho en invadir si bajas freq. inicial
El C puede invadir, pero le toma 5 mil gen. llegar al eq. de 0.1

32
Otra estimación de las w´s:
Burkina Faso

33
Estas adecuaciones son
interesantes... no hay un equilibrio general C siempre invade...

34
la dinámica depende de la freq. de S:
a) S ausente, b) C y S entran al mismo tiempo c) S y A en su equilibrio.
 S ausente, b) C y S entran al mismo tiempo. c) S y A en su equilibrio.")
35
a) a) si no hay S, C aumenta muy rápido a 0.5 en 60 g generaciones 60
 a) si no hay S, C aumenta muy rápido a 0.5 en 60 g generaciones 60")
36
b) b) si S y C se introducen al mismo tiempo ambas aumentan, primero
S más rápido que C, ya que su hetero SA tiene más w

37
c) c) si A y S en su equilibrio, C aumenta muy despacio pC al 0.5
en unas 170 generaciones

38
c común s común si es común S, C es raro

39
dos posibles resultados
Templeton (2006) Topografía de Templeton (2006) dos posibles resultados
 Topografía de. Templeton (2006) dos posibles resultados.")
40
Dos Picos pico adapta- tivo 2 solo C pico adaptativo 1 AS, se pierde C

41
de la “silla de montar” Ampliación si comienzas
es más común, se llega al pico solo C si comienzas aquí, acabas en AS, C muy raro

42
Condiciones incremento tercer alelo
Una z mayor que cero garantiza que entre. Las tres mayor que 0 eq. estable

43
2.- Genes ligados al sexo y organismos haplo-diploides:
A. Complicando las suposiciones del modelo: 2.- Genes ligados al sexo y organismos haplo-diploides:

44
Genes ligados al sexo y Haplo-diploides: recordatorio HW
machos, heredan todo de su mama eq. 2/3 madresAA 1/3 padres A hembras, el promedio de mamás y papás

45
si letal, la freq. de hem. A1A2 1/2 cada gen.
Genes ligados al sexo y Haplo-diploides si letal, la freq. de hem. A1A2 1/2 cada gen.

46
Genes ligados al sexo y Haplo-diploides

47
Cambio q después de la selección:
cada sexo

48
cuatro juegos de adecuaciones…
Genes ligados al sexo y Haplo-diploides vamos a considerar cuatro juegos de adecuaciones…

49
vs. el recesivo

50
1 qf(1-1)=qf x 0
=qf x 0")
51
el cambio en las f. alélicas por sexo diferencias en frecuencias entre sexos

52
vs hembras aa y machos a (más intensa machos)
Genes ligados al sexo y Haplo-diploides; ligado al X, white machos machos vs hembras aa y machos a (más intensa machos)
")
53
condiciones generales de polimorfismo
Genes ligados al sexo y Haplo-diploides: Condiciones de Equilibrio para ligados al sexo: condiciones generales de polimorfismo más reducidas, ya que los machos no pueden tener ventaja heterócigo...

54
un alelo favorecido en machos y
ambos homócigos desfavorables en hembras

55
si hay equilibrio pero los
condiciones polimorfismo estable muy restrictivas (sólo 58% de lo de un locus 2 alelos diploide)
")
56
0<1>0 no conocemos sm (irrelevante) Ejemplo hormiga colonias poligínicas
 Ejemplo hormiga colonias poligínicas")
57
G-6PD (ligado al X) y malaria
RR relative risk (luego) G-6PD (ligado al X) y malaria wBB= wBA-=1 wA-A-<0.972? wB= wA-=1
 G-6PD (ligado al X) y malaria. wBB=0.956 wBA-=1 wA-A-<0.972 wB=0.954 wA-=1.")
58
¿Equilibrio estable?

59
Frecuencia en el Equilibrio
aproximando si s chicas p p

60
Un caso de eq. balanceado 2 sexos
selección opuesta en machos y en hembras equilibrios en dif. p en machos y hembras

61
Hay un polimorfismo balanceado
si fre.alélicas dif. entre sexos, primero oscilaciones y luego se acercan a dif. equilibrios

63
B.-Estimando la Selección: Relative fitness: Adecuación Relativa:
La habilidad de un genotipo, relativa a los otros genotipos, de pasar alelos a la siguiente generación... parece simple, pero tiene varios problemas...

64
Fitness: Adecuación, w AA 100 80 0.8 1 Aa 200 160 0.8 1
Parámetro que mide la intensidad de la selección Ad. absoluta May. minuscu. Genotipo N. inicial N.final lx (=W) w relativa AA Aa aa La w es la adecuación relativa, que se obtiene dividiendo la w entre una de las W´s (a veces la de heterócigo, a veces la más alta...).
 w relativa. AA Aa aa La w es la adecuación. relativa, que se obtiene dividiendo. la w entre una de las W´s (a veces la de heterócigo, a veces la más alta...).")
65
Caso I: Selección vs. recesivo w A1A1 1 A1A2 1 A2A2 1-s
w11= w12 > w22 Selección direccional contra el recesivo w A1A1 1 A1A2 1 A2A2 1-s donde s es el coeficiente de selección (s= 1-w) s=0: no te “ve” la SN (tienes el mejor genotipo) s=1: mata a todos! (letal)
 s=0: no te ve la SN (tienes el mejor genotipo) s=1: mata a todos! (letal)")
66
La estimación de la selección es central
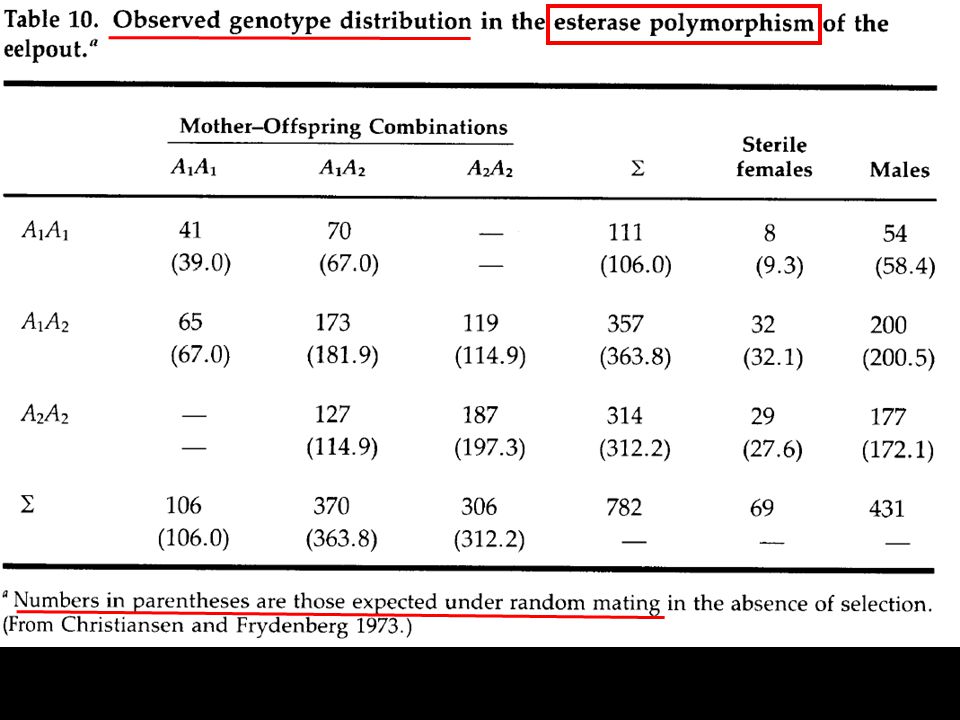
para la ecología evolutiva, pero... hay muchos problemas para le estimación, relacionados tanto al tamaño de muestra, como estadísticos... 1. Método de Christiansen y Frydenberg 1973 (eelpout fish, Zoarces viviparus): analizar los diferentes eventos a lo largo del ciclo de vida “con chi-cuadradas”
: analizar los diferentes. eventos a lo largo del ciclo de vida. con chi-cuadradas")
67
Christiansen y Frydenberg 1973 (Zoarces viviparus):
:")
68
Números observados Números esperados

70
hipótesis explícitas:
Christiansen y Frydenberg (Zoarces) hipótesis explícitas: i) mitad de hijos de madres heterócigas, deben de ser heterócigos
 hipótesis explícitas: i) mitad de. hijos de madres. heterócigas, deben de ser. heterócigos.")
71
ii)apareamiento al azar, iii) sexual male selection
La segunda hipótesis es que los gametos se unen al azar ii)apareamiento al azar, iii) sexual male selection iv) female sexual selection, v) zygotic selection
apareamiento al azar, iii) sexual male selection. iv) female sexual selection, v) zygotic selection.")
72
NS

73
un año bien un homócigo, otro
año el otro homócigo

74
B. Estimando la Selección: 2. Selección en Viabilidad:
cociente de frecuencias genotípicas ¿cómo cambian las frecuencias genotípicas de antes a después de la selección?

75
viabilidad promedio fre. genotípica despúes de la selección

76
cociente para eliminar v promedio

77
Estimando la Selección: Selección en Viabilidad: despejamos las “v”
la w como la viabilidad se estima de la f. genotípicas antes y después de la selección

78
Genotipo N. inicial N.final lx (=W) w relativa
AA Aa aa La w es la adecuación relativa, que se obtiene dividiendo la w entre una de las W´s (a veces la de heterócigo, a veces la más alta...). En realidad sólo formalizamos lo que vimos al principio
. En realidad. sólo formalizamos. lo que vimos al. principio.")
79
homócigo después homócigo antes =(P´11/P11)/ P´12/P12 lx homócigo
estandarizado entre la lx heterócigo, que son 1

80
Ejemplo vx Cebada Mike Clegg (Horedum vulgare)
")
81
P´ P (estandarizado a A1 A1) errores std grandes!
 errores std grandes!")
82
se van co-adaptando los
genes, se eliminan los heterócigos y selección tal en otros genes

83
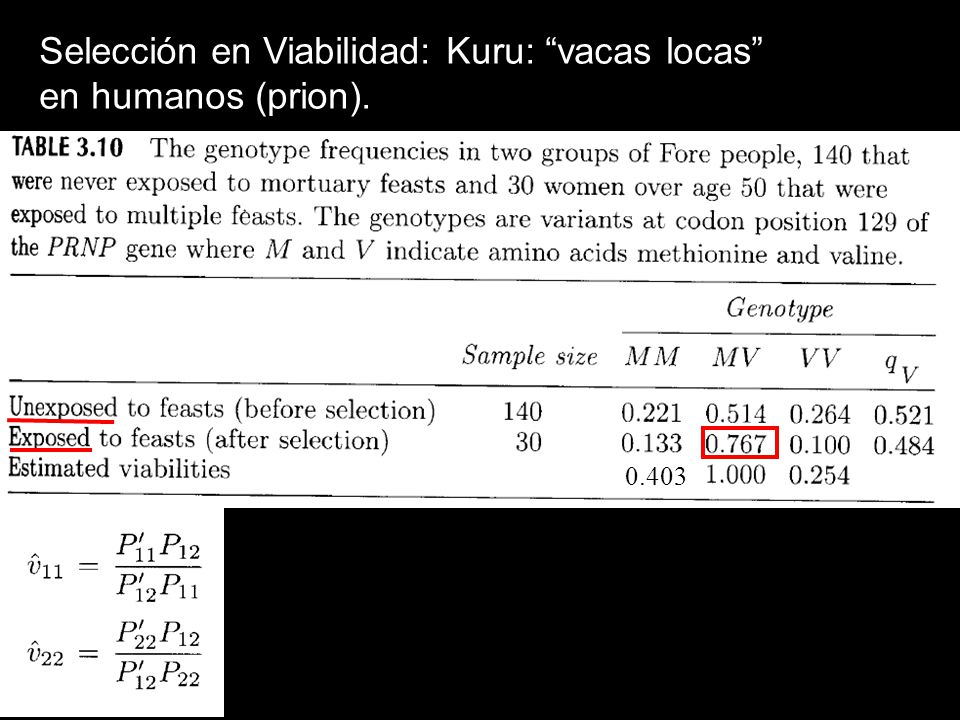
Selección en Viabilidad: Ejemplo Kuru: “vacas locas” en humanos (prion).
.")
84
Ejemplo Kuru:

85
Selección en Viabilidad: Kuru: “vacas locas” en humanos (prion).
0.403
86
Selección en Viabilidad: Kuru: “vacas locas” en humanos (prion).
0.403

87
Selección en Viabilidad: Kuru: “vacas locas” en humanos (prion).
estimación final, corregir ya que sólo en mujeres SMM = 0.358, Svv=0.373, los más altos en humanos eqqv=0.483

88
B. Estimando la Selección:
3. Estimación a partir de desviaciones a HW en las frecuencias genotípicas

89
menos H de los esperados más H de los esperados

90
Estimación a partir de desviaciones a H-W: ineficiente
F = 1 – P´12/2p´q´ para detectar selección fuerte, s=0.2 n de más de ochocientos... débil más

91
4. Desviación de las expectativas Mendelianas
Estimando la Selección: 4. Desviación de las expectativas Mendelianas haciendo cruzas controladas, la forma más elegante A1A1 x A1 A2, se espera iguales números de N11 y de N12

92
Desviación de las expectativas Mendeliana: Biston betularia
no es una cruza, pero las proporciones y la lógica análogas

93
V11= 39/ 58+1=0.661 viabilidad claras fondo obscuro
V22= 24/32+1= viabilidad obs. fondo claro S11= 0.339 S22= 0.273

94
4.- Desviación de las expectativas Mendeliana:
MHC Salmón: ventaja de los heterócigos 2 métodos (2.Selección en Viabilidad y 4 Exp. mendel) cruzas y vs. patógenos
 cruzas y vs. patógenos.")
95
/604= 0.495 /473= 0.518 /604= 0.505 /473= 0.482

96
Virus IHNV: ventaja heterócigos

97
s vs. los homócigos = 0.088 ventaja en el MHC a favor de los heterócigos

98
HLA pocos alelos en Nativos de Sudamérica
HLA (= MHC en humanos) selección balanceadora HLA pocos alelos en Nativos de Sudamérica se puede analizar la segregación en la progenie
 selección balanceadora. HLA pocos alelos en Nativos de Sudamérica. se puede analizar la segregación en la progenie.")
99
Claro ejemplo de ventaja del heterócigo:
exceso de heterócigos en la progenie Selección balanceadora s ca. 0.5, selección MUY intensa

100
otras estimaciones de s muy altas! ventaja de heterócigo! s=1-v 1:1
3:1 s=1-v otras estimaciones de s muy altas! ventaja de heterócigo!

101
5. Riesgo Relativo RR estudios médicos epidemiológicos
Estimando la Selección: 5. Riesgo Relativo RR estudios médicos epidemiológicos

102
Riesgo de los que tiene un genotipo, de enfermarse
frecuencia del genotipo en enfermos frecuencia del genotipo en el control RR<1 genotipo aumenta resistencia RR=1 no hace diferencia RR>1 genotipo aumenta susceptibilidad

103
RR<1 gen aumenta resist.
RR=1 no hace diferencia RR>1 gen aumente sucep

104
m = tasa de mortandad de los enfermos, indep. del genotipo
Riesgo Relativo, w resistentes, RR<1 m = tasa de mortandad de los enfermos, indep. del genotipo

105
Seguimos con w resistentes, RR<1 s=m(1-RR) = 0.5 (1-0) = 0.5
para obtener la w hay que re-escalar, w= 1/(1-s) si RR=0, un genotipo totalmente resistente m =0.5, la tasa de mortalidad de los infectados s=m(1-RR) = 0.5 (1-0) = 0.5 w resistente = 1/(1-s)= 1/0.5=2
 si RR=0, un genotipo totalmente resistente. m =0.5, la tasa de mortalidad de los infectados. s=m(1-RR) = 0.5 (1-0) = 0.5. w resistente = 1/(1-s)= 1/0.5=2.")
106
ahora el caso del susceptible, RR>1
para los genotipos susceptibles si 1/RR=0, son completamente suceptibles mortalidad m= 0.5 s= 0.5(1-0) s=0.5 w= 1-s=0.5
 s=0.5 w= 1-s=0.5.")
107
ejemplo anemia falciforme,
Riesgo relativo: ejemplo anemia falciforme, Burkina Faso de Modiano

108
RR<1 gen aumenta resist.
RR=1 no hace diferencia RR>1 gen aumenta suceptibilidad

109
s= m(1-(1/RR)) susceptible, RR>1 w=1-s resisten. RR<1 w=1/1-s
) susceptible, RR>1 w=1-s resisten. RR<1 w=1/1-s")
110
s= m(1-(1/RR)) w=1-s (s) m=0.1 es sólo un poco de talacha w=1/1-s
susceptible,RR>1 w=1-s resistentes RR<1 w=1/1-s (s) (0.051) (0.029) (0.073) (0.092) m=0.1 es sólo un poco de talacha
 (0.051) (0.029) (0.073) (0.092) m=0.1. es sólo un poco de talacha.")
111
CONCLUSIONES La estimación de la selección y sus parámetros w y s es complicada Se puede confundir con otros procesos Se necesitan diseños cuidadosos Las n necesitan ser muy grandes, especialmente si s es pequeña. Existen varios métodos disponibles, vimos cinco: 1chi-cuadradas, 2 f. genotípicas viabilidades, 3 desv. H.W., 4 cruzas (experimentos) controladas, 5 RR
 controladas, 5 RR.")
112
La selección puede ser MUY intensa
en varios casos la s se ha estimado de más de 0.1, y hasta 0.5 y más: cambio x selección en decenas de generaciones!! También puede ser débil, pero entre más débil es más difícil estimarla de manera directa, pero puede deja una huella molecular, como luego veremos.

113
Otros direccional, como Biston betularia,
Varios casos de selección balanceadora, no sólo anemia falciforme!: kuru, HLA sudamérica, MHC salmón vs. virus. Otros direccional, como Biston betularia, o complicados como Zoarces viviparus o en Hordeum vulgare, que parece que cambia en el tiempo, coadaptaciones, “underdominace” Próxima clase: complicaciones a la SN (sel. sexual, factores ecológicos, apareamientos clasificados, etc.!)
")
114
Artículos de selección: Martes 18 septiembre
1) Allison AC Anemia, British Med. J. (1954) Ana Teresa 2) Allison AC Anemia 2, Ann. H. Genetics (1954) Nancy Jueves 20: 3) Edward y Chapman 2011, TREE, Selección sexual. Victor Argaez 4) Lenski y Travisano Pnas Alejandra G. 5) Rainey y Trevisano Natures G. Yaxal minutos… todos leer y resumen EXAMEN PRIMERA PARTE: MARTES 25 DE SEPTIEMBRE
 Allison AC Anemia, British Med. J. (1954) Ana Teresa. 2) Allison AC Anemia 2, Ann. H. Genetics (1954) Nancy. Jueves 20: 3) Edward y Chapman 2011, TREE, Selección sexual. Victor Argaez. 4) Lenski y Travisano Pnas Alejandra G. 5) Rainey y Trevisano Natures G. Yaxal minutos… todos leer y resumen. EXAMEN PRIMERA PARTE: MARTES 25 DE SEPTIEMBRE.")
115
(pensar en el trabajo semestral)
fin!

Presentaciones similares