Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Termodinámica molecular I. http://einstein.ciencias.uchile.cl/ BioFisicoQuimicaMacroMolecular2008/Clases/Clase3 2008

2
2007 Fundamentos de la simulación de computacional de macromoléculas.

3
Simplificaciones para tratar un problema complejo Conjunto dinámico de moléculas en solución

4
Simplificaciones para tratar un problema complejo Conjunto dinámico de moléculas en solución Caja periódica

5
Simplificaciones para tratar un problema complejo Conjunto dinámico de moléculas en solución Caja periódica Molécula dinámica

6
Simplificaciones para tratar un problema complejo Conjunto dinámico de moléculas en solución Caja periódica Molécula dinámica Molécula estática

7
En un l una solución 1mM de insulina hay 6·10 14 moléculas de insulina. Cada molécula de insulina tiene 760 átomos, está constantemente cambiando de forma, e interactuando con 10 8 moléculas de agua. Primera simplificación. Cajas periódicas. El comportamiento promedio de un conjunto de macromoléculas se puede representar por un solo ejemplar de la macromolécula. (Se pierde las interacciones entre las macromoléculas) Molécula dinámica en una caja periódica llena de moléculas de solvente En el computador (in silico) se construye un modelo que simula un sistema en que una macromolécula está confinada en una caja llena de moléculas de solvente y se simula la trayectoria de la estructura en el tiempo. Dinámica molecular. http://en.wikipedia.org/wiki/In_silico
 Molécula dinámica en una caja periódica llena de moléculas de solvente En el computador (in silico) se construye un modelo que simula un sistema en que una macromolécula está confinada en una caja llena de moléculas de solvente y se simula la trayectoria de la estructura en el tiempo. Dinámica molecular.")
8
Primera simplificación. Cajas periódicas. El comportamiento promedio de un conjunto de macromoléculas se puede representar por un solo ejemplar de la macromolécula. (Se pierde las interacciones entre las macromoléculas) Molécula dinámica en una caja periódica llena de moléculas de solvente Se simula un arreglo tridimensional de cajas todas iguales. Se copia en todas las otras cajas lo que ocurra en la caja central. Como todas las cajas son iguales, la molécula entra a la caja de donde salió por el lado izquierdo. Así se simula que esta caja es parte de una conjunto grande. Si una molécula de solvente sale por el lado derecho de una caja entra a la vecina por el lado izquierdo.
 Molécula dinámica en una caja periódica llena de moléculas de solvente Se simula un arreglo tridimensional de cajas todas iguales. Se copia en todas las otras cajas lo que ocurra en la caja central. Como todas las cajas son iguales, la molécula entra a la caja de donde salió por el lado izquierdo. Así se simula que esta caja es parte de una conjunto grande. Si una molécula de solvente sale por el lado derecho de una caja entra a la vecina por el lado izquierdo..")
9
Primera simplificación. Cajas periódicas. El comportamiento promedio de un conjunto de macromoléculas se puede representar por un solo ejemplar de la macromolécula. (Se pierde las interacciones entre las macromoléculas) Molécula dinámica en una caja periódica llena de moléculas de solvente El sistema que tenía inicialmente 6·10 14 moléculas de insulina y 3,3 10 20 moléculas de agua se reduce a 760 átomos de la macromolécula y unas 2300 moléculas de agua. (caja de 20x20x20 Å)
 Molécula dinámica en una caja periódica llena de moléculas de solvente El sistema que tenía inicialmente 6·10 14 moléculas de insulina y 3,3 moléculas de agua se reduce a 760 átomos de la macromolécula y unas 2300 moléculas de agua. (caja de 20x20x20 Å).")
10
Primera simplificación. Cajas periódicas. El comportamiento promedio de un conjunto de macromoléculas se puede representar por un solo ejemplar de la macromolécula. Segunda simplificación. El solvente como un continuo Se elimina la mayor parte de las moléculas del solvente. Sólo quedan las moléculas de solvente asociadas fuertemente a la macromolécula. El solvente es representado por un medio continuo caracterizado por propiedades macroscópicas, como la constante dieléctrica. No hacen falta las cajas periódicas. Molécula dinámica inmersa en un continuo sin estructura El sistema se reduce a 760 átomos + 20 moléculas de agua.

11
Primera simplificación. Cajas periódicas. El comportamiento promedio de un conjunto de macromoléculas se puede representar por un solo ejemplar de la macromolécula. Segunda simplificación. Solvernte momo un continuo. Se elimina la mayor parte de las moléculas del solvente. Como la macromolécula está en el vacío, no hacen falta las caja periódicas. Sólo quedan las moléculas de solvente asociadas fuertemente a la macromolécula. Tercera simplificación. Molécula estática. La estructura nativa de la macromolécula es la de menor energía. Se elimina las propiedades dinámicas de la macromolécula. Mecánica Molecular. Minimización de energía. Molécula estática

12
Energía potencial y energía cinética. La energía E del sistema es la suma de su energía cinética K más su energía potencial U. La energía cinética corresponde al movimiento de los átomos. La energía potencial corresponde a la energía de la interacción entre los diferentes átomos de la macromolécula. El movimiento de los átomos obedece a las leyes de Newton. La fuerza F en la dirección de la trayectoria de un móvil se relaciona con su masa, m, y su aceleración, a, a lo largo de la trayectoria, según esta ley:

13
La fuerza ejercida en la dirección del vector de la trayectoria, r, se relaciona con el gradiente local de energía potencial: El sistema está en equilibrio cuando F = 0, es decir en un mínimo de energía potencial.

14
La fuerza ejercida en la dirección de r se relaciona con el gradiente local de energía potencial: El sistema está en equilibrio cuando F = 0, es decir en un mínimo de energía. Este es el principio sobre el cual se basan los métodos de minimización de energía, que nos buscan la estructura más estable. Con este método se obtiene la imagen estática de la molécula. Las propiedades dinámicas de la molécula se simulan a través de la energía cinética de sus átomos. La energía cinética es función de la temperatura.

15
Las propiedades dinámicas de la molécula se simulan a través de la energía cinética de sus átomos. La energía cinética es función de la temperatura. La velocidad al cuadrado promedio es: La velocidad cuadrática media es: La distribución de la velocidad debe ser compatible con la distribución de Boltzmann de la energía: es la distribución de Maxwell-Boltzmann. http://hyperphysics.phy-astr.gsu.edu/hbase/kinetic/maxspe.html#c1

16
Potenciales moleculares. La energía potencial de una molécula con N átomos es la suma de la energía potencial de cada uno de sus átomos. La energía potencial del átomo i es la suma de la energía de las interacciones de este átomo con el resto de los átomos del sistema. La energía potencial de las interacciones obedece a reglas diferentes dependiendo de si los átomos están o no unidos por enlaces covalentes.

17
Continuación 13 ago 2008

18
La energía potencial de las interacciones obedece a reglas diferentes dependiendo de si los átomos están o no unidos por enlaces covalentes Interacciones covalentes Estiramiento de enlaces Deformación de los ángulos de enlace Torsiones Torsiones impropias Interacciones no covalentes Interacciones de van der Waals Interacciones carga-carga Interacciones dipolo-dipolo Interacciones covalentes Estiramiento de enlaces Deformación de los ángulos de enlace Torsiones Torsiones impropias Interacciones no covalentes Interacciones de van der Waals Interacciones carga-carga Interacciones dipolo-dipolo

19
Potenciales de las interacciones covalentes. Energía guardada al deformar un resorte: x0x0 x Ley de Hooke: la relación entre la fuerza F y la longitud x del resorte es: La energía potencial que se acumula en una deformación infinitesimal del resorte sigue esta ley: La energía acumulada al deformar el resorte desde x i = x 0 a x f = x: La energía potencial acumulada en el resorte es siempre positiva, ya sea para x f > x 0 o x f < x 0. http://webphysics.davidson.edu/applets/animator4/demo_hook.html

20
La energía acumulada en el resorte es siempre positiva, ya sea para x f > x 0 o x f < x 0. Este potencial se usa para describir la energía acumulada por estirar o acortar la distancia interatómica de dos átomos unidos por un enlace covalente. U B ( B de bond = enlace ) Tiene forma de parábola y se llama potencial armónico. La parábola pasa por cero para r = r 0. La distancia r 0 para enlaces covalentes típicos se obtiene de mediciones cristalográficas en moléculas pequeñas. El coeficiente k B se obtiene de datos obtenidos de mediciones de espectros de las fluctuaciones de las distancias de enlace en moléculas pequeñas. Se supone que todos los enlaces de una misma clase tienen iguales propiedades independiente de la molécula en que se encuentren. r
 Tiene forma de parábola y se llama potencial armónico. La parábola pasa por cero para r = r 0. La distancia r 0 para enlaces covalentes típicos se obtiene de mediciones cristalográficas en moléculas pequeñas. El coeficiente k B se obtiene de datos obtenidos de mediciones de espectros de las fluctuaciones de las distancias de enlace en moléculas pequeñas. Se supone que todos los enlaces de una misma clase tienen iguales propiedades independiente de la molécula en que se encuentren. r.")
21
r Calcular la energía cinética suponiendo U+K = 100 J? Energía. J Calcular la velocidad?

22
r Calcular la energía cinética suponiendo U+K = 100 J? ¿Calcular la velocidad? ¿Cuál es la distancia promedio? ¿Cuál es la distancia más probable?

23
r Tarea: Trazar la trayectoria r(t) para U + K = cte. Calcular la energía cinética suponiendo U + K = 100 J? Calcular la velocidad? Cuál es la distancia promedio? Cuál es la distancia más probable?

24
http://www.edu.aytolacoruna.es/aula/fisica/teoria/A_Franco/oscilaciones/mas/mas.htm
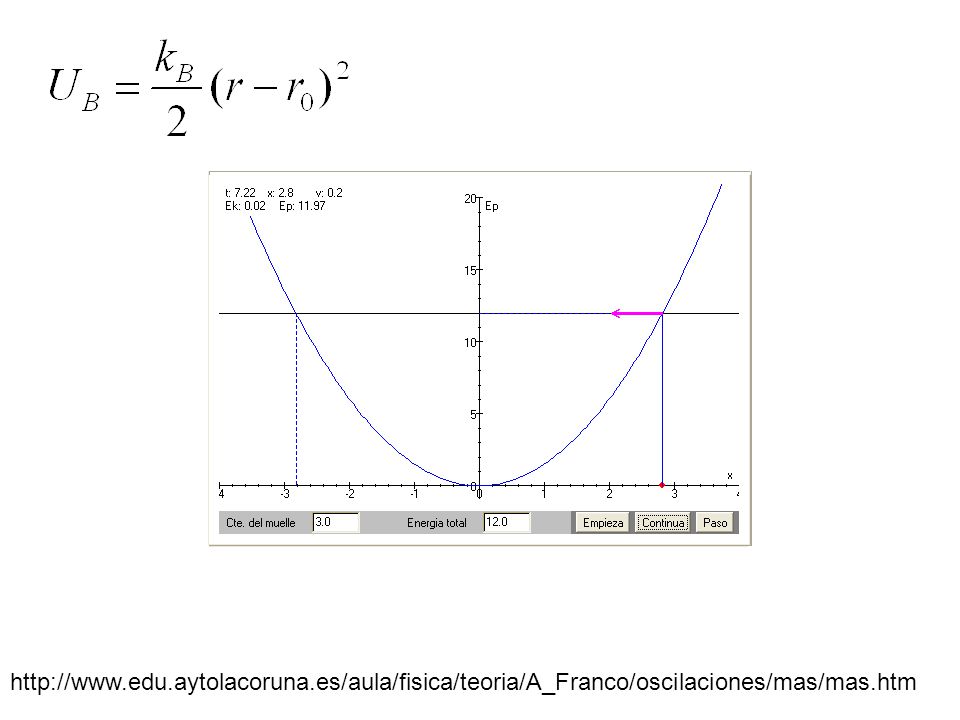
25
C-H2681.099 C-C1101.490 C-N(amida)4031.278 C=05951.200 C-N2011.457 N-H4050.980 k B /2 kcal/mol Å 2 r0År0Å Parámetros para el potencial armónico del estiramiento de enlaces. Arieh Warshel. “Computer Modeling of Chemical Reactions in Enzymes and Solutions”. John Wiley and Sons, Inc, New York. 1991 Table 4.1 pag 112 1 caloría = 4,184 joule

26
¿Qué energía se necesita para romper el enlace? La energía del potencial armónico crece sin límite al estirar el enlace y nunca se rompe. El potencial armónico no sirve para simular reacciones químicas en que se rompen y crean enlaces. Para estas simulaciones de usa el potencial de Morse que permite romper el enlace.

27
http://www.sc.ehu.es/sbweb/fisica/oscilaciones/morse/morse.htm Potencial de Morse U B,Morse = -D para r = r 0 D es la energía necesaria para romper el enlace. Discutir sobre “ el error de los enlaces ricos en energía”. r - r 0 http://einstein.ciencias.uchile.cl/BioFisicoQuimicaMacroMolecular2008/ Lecturas/ElErrorDeLosEnlacesRicosEnEnergia

28
Fragmento de un archivo PDB (Protein Data Bank) de una molécula de polialanina ATOM1NALAA1-34.19234.97573.362 ATOM2CAALAA1-33.01034.68372.517 ATOM3CALAA1-33.28433.75671.315 ATOM4OALAA1-34.36033.17071.214 ATOM5CBALAA1-31.89434.10473.392 ATOM6NALAA2-32.23333.48170.540 ATOM7CAALAA2-32.29532.61369.344 ATOM8CALAA2-30.94732.44768.614 ATOM9OALAA2-29.90832.86169.125 ATOM10CBALAA2-33.35533.16068.383 ATOM11NALAA3-30.96231.67167.529 ATOM12CAALAA3-29.76731.39766.704 ATOM13CALAA3-30.06830.47065.511 ATOM14OALAA3-31.20630.04165.329 ATOM15CBALAA3-28.66530.80067.585 Para saber más sobre el formato PDB visite: http://www.rcsb.org/pdb/docs/format/pdbguide2.2/guide2.2_frame.html Descripción del contenido de la línea (ATOM, HETATM, TER, END, REMARK, etc)
 de una molécula de polialanina ATOM1NALAA ATOM2CAALAA ATOM3CALAA ATOM4OALAA ATOM5CBALAA ATOM6NALAA ATOM7CAALAA ATOM8CALAA ATOM9OALAA ATOM10CBALAA ATOM11NALAA ATOM12CAALAA ATOM13CALAA ATOM14OALAA ATOM15CBALAA Para saber más sobre el formato PDB visite: Descripción del contenido de la línea (ATOM, HETATM, TER, END, REMARK, etc)")
29
Fragmento de un archivo PDB (Protein Data Bank) de una molécula de polialanina ATOM1NALAA1-34.19234.97573.362 ATOM2CAALAA1-33.01034.68372.517 ATOM3CALAA1-33.28433.75671.315 ATOM4OALAA1-34.36033.17071.214 ATOM5CBALAA1-31.89434.10473.392 ATOM6NALAA2-32.23333.48170.540 ATOM7CAALAA2-32.29532.61369.344 ATOM8CALAA2-30.94732.44768.614 ATOM9OALAA2-29.90832.86169.125 ATOM10CBALAA2-33.35533.16068.383 ATOM11NALAA3-30.96231.67167.529 ATOM12CAALAA3-29.76731.39766.704 ATOM13CALAA3-30.06830.47065.511 ATOM14OALAA3-31.20630.04165.329 ATOM15CBALAA3-28.66530.80067.585 Para saber más sobre el formato PDB visite: http://www.rcsb.org/pdb/docs/format/pdbguide2.2/guide2.2_frame.html Número del átomo
 de una molécula de polialanina ATOM1NALAA ATOM2CAALAA ATOM3CALAA ATOM4OALAA ATOM5CBALAA ATOM6NALAA ATOM7CAALAA ATOM8CALAA ATOM9OALAA ATOM10CBALAA ATOM11NALAA ATOM12CAALAA ATOM13CALAA ATOM14OALAA ATOM15CBALAA Para saber más sobre el formato PDB visite: Número del átomo")
30
Fragmento de un archivo PDB (Protein Data Bank) de una molécula de polialanina ATOM1NALAA1-34.19234.97573.362 ATOM2CAALAA1-33.01034.68372.517 ATOM3CALAA1-33.28433.75671.315 ATOM4OALAA1-34.36033.17071.214 ATOM5CBALAA1-31.89434.10473.392 ATOM6NALAA2-32.23333.48170.540 ATOM7CAALAA2-32.29532.61369.344 ATOM8CALAA2-30.94732.44768.614 ATOM9OALAA2-29.90832.86169.125 ATOM10CBALAA2-33.35533.16068.383 ATOM11NALAA3-30.96231.67167.529 ATOM12CAALAA3-29.76731.39766.704 ATOM13CALAA3-30.06830.47065.511 ATOM14OALAA3-31.20630.04165.329 ATOM15CBALAA3-28.66530.80067.585 Tipo de átomo N=nitrógeno amida C=carbono carbonilo CA=Carbono alfa CB=Carbono beta Para saber más sobre el formato PDB visite: http://www.rcsb.org/pdb/docs/format/pdbguide2.2/guide2.2_frame.html
 de una molécula de polialanina ATOM1NALAA ATOM2CAALAA ATOM3CALAA ATOM4OALAA ATOM5CBALAA ATOM6NALAA ATOM7CAALAA ATOM8CALAA ATOM9OALAA ATOM10CBALAA ATOM11NALAA ATOM12CAALAA ATOM13CALAA ATOM14OALAA ATOM15CBALAA Tipo de átomo N=nitrógeno amida C=carbono carbonilo CA=Carbono alfa CB=Carbono beta Para saber más sobre el formato PDB visite:")
31
Fragmento de un archivo PDB (Protein Data Bank) de una molécula de polialanina ATOM1NALAA1-34.19234.97573.362 ATOM2CAALAA1-33.01034.68372.517 ATOM3CALAA1-33.28433.75671.315 ATOM4OALAA1-34.36033.17071.214 ATOM5CBALAA1-31.89434.10473.392 ATOM6NALAA2-32.23333.48170.540 ATOM7CAALAA2-32.29532.61369.344 ATOM8CALAA2-30.94732.44768.614 ATOM9OALAA2-29.90832.86169.125 ATOM10CBALAA2-33.35533.16068.383 ATOM11NALAA3-30.96231.67167.529 ATOM12CAALAA3-29.76731.39766.704 ATOM13CALAA3-30.06830.47065.511 ATOM14OALAA3-31.20630.04165.329 ATOM15CBALAA3-28.66530.80067.585 Para saber más sobre el formato PDB visite: http://www.rcsb.org/pdb/docs/format/pdbguide2.2/guide2.2_frame.html Tipo residuo en código de 3 letras
 de una molécula de polialanina ATOM1NALAA ATOM2CAALAA ATOM3CALAA ATOM4OALAA ATOM5CBALAA ATOM6NALAA ATOM7CAALAA ATOM8CALAA ATOM9OALAA ATOM10CBALAA ATOM11NALAA ATOM12CAALAA ATOM13CALAA ATOM14OALAA ATOM15CBALAA Para saber más sobre el formato PDB visite: Tipo residuo en código de 3 letras")
32
http://www.fao.org/documents/show_cdr.asp?url_file=/DOCREP/004/Y2775E/y2775e0e.htm

33
Fragmento de un archivo PDB (Protein Data Bank) de una molécula de polialanina ATOM1NALAA1-34.19234.97573.362 ATOM2CAALAA1-33.01034.68372.517 ATOM3CALAA1-33.28433.75671.315 ATOM4OALAA1-34.36033.17071.214 ATOM5CBALAA1-31.89434.10473.392 ATOM6NALAA2-32.23333.48170.540 ATOM7CAALAA2-32.29532.61369.344 ATOM8CALAA2-30.94732.44768.614 ATOM9OALAA2-29.90832.86169.125 ATOM10CBALAA2-33.35533.16068.383 ATOM11NALAA3-30.96231.67167.529 ATOM12CAALAA3-29.76731.39766.704 ATOM13CALAA3-30.06830.47065.511 ATOM14OALAA3-31.20630.04165.329 ATOM15CBALAA3-28.66530.80067.585 Para saber más sobre el formato PDB visite: http://www.rcsb.org/pdb/docs/format/pdbguide2.2/guide2.2_frame.html Cadena: A, B, C etc
 de una molécula de polialanina ATOM1NALAA ATOM2CAALAA ATOM3CALAA ATOM4OALAA ATOM5CBALAA ATOM6NALAA ATOM7CAALAA ATOM8CALAA ATOM9OALAA ATOM10CBALAA ATOM11NALAA ATOM12CAALAA ATOM13CALAA ATOM14OALAA ATOM15CBALAA Para saber más sobre el formato PDB visite: Cadena: A, B, C etc")
34
Fragmento de un archivo PDB (Protein Data Bank) de una molécula de polialanina ATOM1NALAA1-34.19234.97573.362 ATOM2CAALAA1-33.01034.68372.517 ATOM3CALAA1-33.28433.75671.315 ATOM4OALAA1-34.36033.17071.214 ATOM5CBALAA1-31.89434.10473.392 ATOM6NALAA2-32.23333.48170.540 ATOM7CAALAA2-32.29532.61369.344 ATOM8CALAA2-30.94732.44768.614 ATOM9OALAA2-29.90832.86169.125 ATOM10CBALAA2-33.35533.16068.383 ATOM11NALAA3-30.96231.67167.529 ATOM12CAALAA3-29.76731.39766.704 ATOM13CALAA3-30.06830.47065.511 ATOM14OALAA3-31.20630.04165.329 ATOM15CBALAA3-28.66530.80067.585 Para saber más sobre el formato PDB visite: http://www.rcsb.org/pdb/docs/format/pdbguide2.2/guide2.2_frame.html Número del residuo
 de una molécula de polialanina ATOM1NALAA ATOM2CAALAA ATOM3CALAA ATOM4OALAA ATOM5CBALAA ATOM6NALAA ATOM7CAALAA ATOM8CALAA ATOM9OALAA ATOM10CBALAA ATOM11NALAA ATOM12CAALAA ATOM13CALAA ATOM14OALAA ATOM15CBALAA Para saber más sobre el formato PDB visite: Número del residuo")
35
Fragmento de un archivo PDB (Protein Data Bank) de una molécula de polialanina ATOM1NALAA1-34.19234.97573.362 ATOM2CAALAA1-33.01034.68372.517 ATOM3CALAA1-33.28433.75671.315 ATOM4OALAA1-34.36033.17071.214 ATOM5CBALAA1-31.89434.10473.392 ATOM6NALAA2-32.23333.48170.540 ATOM7CAALAA2-32.29532.61369.344 ATOM8CALAA2-30.94732.44768.614 ATOM9OALAA2-29.90832.86169.125 ATOM10CBALAA2-33.35533.16068.383 ATOM11NALAA3-30.96231.67167.529 ATOM12CAALAA3-29.76731.39766.704 ATOM13CALAA3-30.06830.47065.511 ATOM14OALAA3-31.20630.04165.329 ATOM15CBALAA3-28.66530.80067.585 Para saber más sobre el formato PDB visite: http://www.rcsb.org/pdb/docs/format/pdbguide2.2/guide2.2_frame.html Coordenadas xyz en Å
 de una molécula de polialanina ATOM1NALAA ATOM2CAALAA ATOM3CALAA ATOM4OALAA ATOM5CBALAA ATOM6NALAA ATOM7CAALAA ATOM8CALAA ATOM9OALAA ATOM10CBALAA ATOM11NALAA ATOM12CAALAA ATOM13CALAA ATOM14OALAA ATOM15CBALAA Para saber más sobre el formato PDB visite: Coordenadas xyz en Å")
36
El fragmento de la molécula de polialanina dibujado usando las coordenadas xyz del archivo PDB y el programa DS Visualizer 1.6 N CA CB O C http://www.accelrys.com/products/downloads/ds_visualizer/index.html Faltan los átomos de hidrógeno

37
Fragmento de un archivo PDB de una molécula de polialanina ATOM6NALAA2-32.23333.48170.540 ATOM7CAALAA2-32.29532.61369.344 ATOM8CALAA2-30.94732.44768.614 ATOM9OALAA2-29.90832.86169.125 ATOM10CBALAA2-33.35533.16068.383 Problema: Calcular la energía potencial del enlace C - C CA CB

38
ATOM7CAALAA2-32.29532.61369.344 ATOM10CBALAA2-33.35533.16068.383 x y z CC CC

39
x y z ATOM7CAALAA2-32.29532.61369.344 ATOM10CBALAA2-33.35533.16068.383

40
ATOM7CAALAA2-32.29532.61369.344 ATOM10CBALAA2-33.35533.16068.383 Å Datos de Warshel

41
Energía de los ángulos de enlace

42
k /2 kcal/mol 00 Parámetros para el potencial de los ángulos de enlace para amidas. Arieh Warshel. Computer Modeling of Chemical Reactions in Enzymes and Solutions. John Wiley and Sons, Inc, New York. 1991. Table 4.1 pag 112 H-C-H401.911 (109.5°) C-C-H251.911 C-C-C181.911 C’-N-H262.094 (120°) C’-N-C542.094 N-C’-O482.094 N-C’-C412.094 Linus Pauling The Chemical Bond, Cornell, NY, 1967
 C-C-H C-C-C C’-N-H (120°) C’-N-C N-C’-O N-C’-C Linus Pauling The Chemical Bond, Cornell, NY,")
43
Fragmento de un archivo PDB de una molécula de polialanina ATOM6NALAA2-32.23333.48170.540 ATOM7CAALAA2-32.29532.61369.344 ATOM8CALAA2-30.94732.44768.614 ATOM9OALAA2-29.90832.86169.125 ATOM10CBALAA2-33.35533.16068.383 Problema: Calcular la energía potencial del ángulo de enlace C - C - C C CA CB

44
CC CC C ATOM8CALAA2-30.94732.44768.614 ATOM7CAALAA2-32.29532.61369.344 ATOM10CBALAA2-33.35533.16068.383

45
CC CC C ATOM8CALAA2-30.94732.44768.614 ATOM7CAALAA2-32.29532.61369.344 ATOM10CBALAA2-33.35533.16068.383

46
ATOM8CALAA2-30.94732.44768.614 ATOM7CAALAA2-32.29532.61369.344 ATOM10CBALAA2-33.35533.16068.383

47
cos Para enlace C-C-C ATOM8CALAA2-30.94732.44768.614 ATOM7CAALAA2-32.29532.61369.344 ATOM10CBALAA2-33.35533.16068.383

48
Energía de las torsiones de enlaces A B C D A D ±180 A D -120 A D -60 A D 60 A D 120 A D 0 Proyecciones de Newman

49
A B C D El ángulo de torsión es el ángulo que hace el plano ABC con el plano BCD La energía de la torsión es una función periódica. El potencial de Rykaert-Ballemans:

50
= 0 = 180 1,2 dicloroetileno

51
1,2 dicloroetano =0 =60 =180

52
ATOM6NALAA2-32.23333.48170.540 ATOM7CAALAA2-32.29532.61369.344 ATOM8CALAA2-30.94732.44768.614 ATOM9OALAA2-29.90832.86169.125 ATOM10CBALAA2-33.35533.16068.383 ATOM11NALAA3-30.96231.67167.529 Fragmento de un archivo PDB de una molécula de polialanina Problema: Calcular la energía potencial de la torsión del enlace N - C - C - N+ N CA C N+

53
Determinación del vector perpendicular a un plano definido por los vectores A y B.

54
CC C N Determinación del vector perpendicular al plano N-C -C

55
ATOM6NALAA2-32.23333.48170.540 ATOM7CAALAA2-32.29532.61369.344 ATOM8CALAA2-30.94732.44768.614 ATOM11NALAA3-30.96231.67167.529 Determinación del vector perpendicular al plano N-C -C

56
Este es el vector perpendicular al plano N-C -C

57
ATOM6NALAA2-32.23333.48170.540 ATOM7CAALAA2-32.29532.61369.344 ATOM8CALAA2-30.94732.44768.614 ATOM11NALAA3-30.96231.67167.529 Determinación del vector perpendicular al plano C -C-N+

58
Éste es el vector perpendicular al plano C -C-N+

59
Éste es el vector perpendicular al plano N-C -C Éste es el vector perpendicular al plano C -C-N+

60
El ángulo de torsión es 180 grados, la energía es 0

61
Energía de las torsiones impropias L alaninaD alanina El campo de fuerza incluye una barrera de energía que impide el cambio de configuración L a D. Esta barrera se denomina torsión impropia

62
Energía de las torsiones impropias El campo de fuerza incluye una pozo de energía que mantiene en un solo plano los átomos del enlace peptídico. Esta es otra torsión impropia

63

Presentaciones similares






>")

 y líneas equipotenciales.>")

>")








