Descargar la presentación
La descarga está en progreso. Por favor, espere

1
REMODELADO MITOTICO DEL REPLICON Y DE LA ESTRUCTURA CROMOSOMICA
Jean-Marc Lemaitre,1 Etienne Danis,1 Philippe Pasero,2 Yegor Vassetzky,1 and Marcel Me´ chali1,* Petignat, Camila Rochi, Lucía Sampayo, Rocío Toro, Ayelén

2
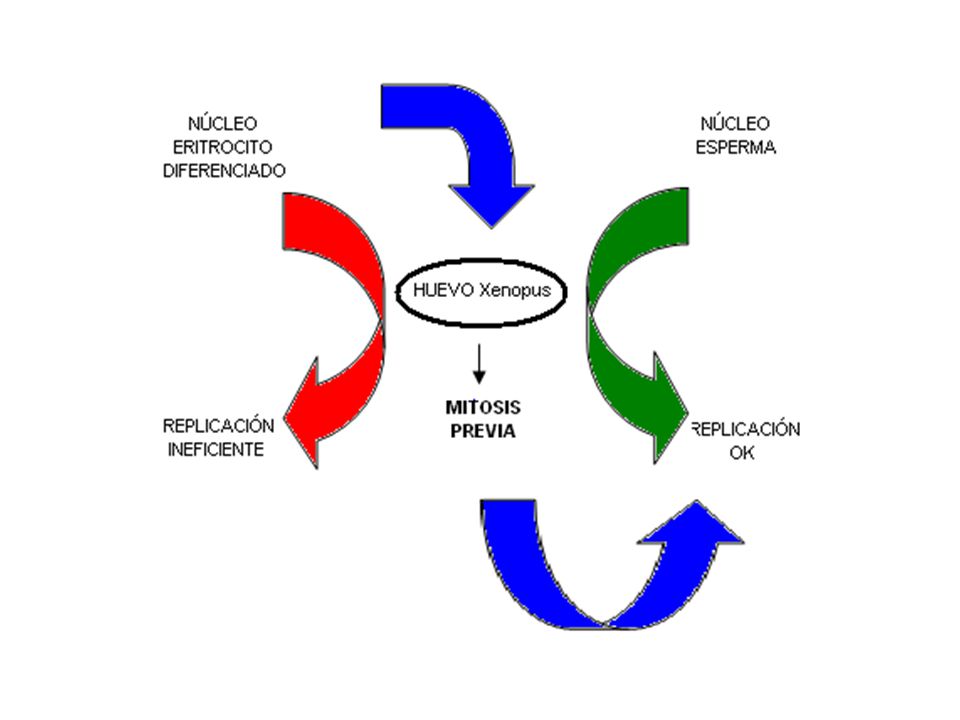
Núcleos adultos diferenciados No se adaptan bien a condiciones de
replicación Se usan espermas se observa que el núcleo está organizado en loops cortos antes de la fertilización Permite Replicación rápida Se trabaja en Xenopus esperma en extracto de huevo de Xenopus, se reconstituye núcleo Se organiza el núcleo. DNA se replica.

3
TRANSFERENCIA NUCLEAR
Permite clonación de animales y obtención de CÉLULAS MULTIPOTENTES de tejidos ya diferenciados Suele ser ineficiente ya que en núcleo adulto ya diferenciado es más complicada la replicación Se quiere identificar qué factores están involucrados en la remodelación

4
Baja eficiencia de clonación
Organización cromosómica del núcleo adulto no se adapta bien a las condiciones necesarias para que haya replicación

5
OBJETIVOS Determinar qué factores permiten que núcleo adulto se adapte a eventos de desarrollo temprano (Replicación rápida). DEMUESTRAN La MITOSIS es crucial en la reorganización nuclear en células adultas (que permite adaptarse a los patrones de replicación embrionarios)
. DEMUESTRAN. La MITOSIS es crucial en la reorganización nuclear en células adultas. (que permite adaptarse a los patrones de replicación embrionarios)")
7
RESULTADOS

8
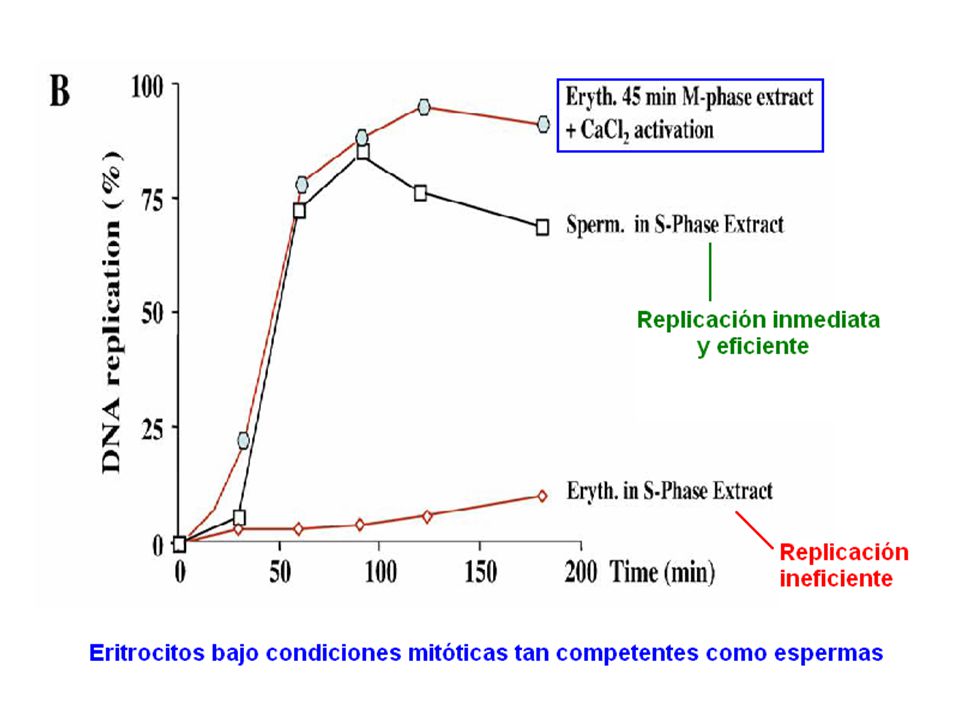
Para que núcleo adulto pueda replicar es necesario paso previo por mitosis.

10
Análisis de focos de inicio de replicación en eritrocitos:
Aumenta número de focos con paso previo por mitosis

11
¿Remodelación mitótica afecta el tamaño del replicón?
DNA combing (BrdUTP) Eritrocitos en extracto fase S la distancia entre los orígenes varía entre 30 y 230 Kbp El 97% de los replicones son mayores a 30 Kbp La lenta replicación observada en núcleos de eritrocitos se debe a una baja frecuencia de inicios de replicación
 Eritrocitos en extracto fase S la distancia entre los orígenes varía entre 30 y 230 Kbp. El 97% de los replicones son mayores a 30 Kbp. La lenta replicación observada en núcleos de eritrocitos se debe a una baja frecuencia de inicios de replicación.")
12
En eritrocitos previamente expuestos a extracto en fase M el 74% de los replicones es menor a 30 Kbp
El pasaje por la mitosis acortó las distancias entre los orígenes, siendo ahora similares a lo observado en esperma.

13
Organización del nucleosoma
La organización del nucleosoma es similar Igual nivel de proteinas del complejo pre-RC Estas proteinas no se pegan a la cromatina durante la mitosis

14
La acetilación de histonas puede contribuir a la modificación de los orígenes de replicación
TSA: inhibe desacetilasas CTPB: activa acetiltransferasas AA: inhibe acetiltransferasas Acetilación de histonas no necesaria para replicación nivel de acetilación del extracto es suficiente para la máxima replicación

15
CTPB restituye la replicación
AA disminuye la repliación La acetilación de histonas es necesaria para la rápida replicación observada durante el desarrollo temprano y el nivel de acetilación en el huevo es suficiente para obtener el máximo de replicación.

16
La acetilación de las histonas influye en la replicación de los eritrocitos, siempre y cuando hayan sido expuestos primero a un extracto en fase M, de manera que se permita la formación de los cromosomas mitóticos.

17
Entonces…¿El remodelado del replicón en fase M podría deberse a una reorganización de la cromatina?
N. Eritrocito incubado en extracto M o S o en solución control LIS: extracción de DNA sin histonas ni otras proteínas, pero mantiene los sitios de anclaje del DNA a la matriz nuclear Sondas de DNA para hibridar en geles de agarosa

18
Sonda: DNA total de eritrocito (digerido)
En los geles corren 5 fragmentos de DNA, producidos por clonado en plásmidos, de regiones de DNA codificante para ribosomas (rDNA) varias copias en el genoma de Xenopus posibilidad de hibridar Plásmido Sonda: DNA total de eritrocito (digerido) Sondas: MARs En solución control: sólo se asocia a matriz rDNA de región “1” S: no modifica la organización nuclear del rDNA M: los 5 dominios de rDNA se detectan = los 5 se asocian a matriz M produce reorganización de sitios de anclaje del rDNA a matriz Ctrol. plásmido Sondas de DNA
 varias copias en el genoma de Xenopus + posibilidad de hibridar. Plásmido. Sonda: DNA total de eritrocito (digerido) Sondas: MARs. En solución control: sólo se asocia a matriz rDNA de región 1 S: no modifica la organización nuclear del rDNA. M: los 5 dominios de rDNA se detectan = los 5 se asocian a matriz. M produce reorganización de sitios de anclaje del rDNA a matriz. Ctrol. plásmido. Sondas de DNA.")
19
Para explicar estos resultados se investigó si M afecta el tamaño de los “loop” de DNA (en los dominios de rDNA) Medición del tamaño de los “loop” por la técnica de Radio del Halo de Máxima Fluorescencia (MFHR) Buffer: [sales] DNA loops forman halo fluorescente rodeando al esqueleto nuclear residual Loop size = 2x MHFR Tinción + radiación UV Tamaño DNA lineal: 1um = 2.3kbp
 Buffer: [sales] DNA loops forman halo fluorescente rodeando al esqueleto nuclear residual. Loop size = 2x MHFR. Tinción + radiación UV. Tamaño DNA lineal: 1um = 2.3kbp.")
20
Se distingue el halo del esqueleto nuclear
ubicando a éste último por inmunolocalización N. Eritr a) Solución control b) Extracto S (interfase) c) Extracto M Ca++ N. Esperma d) Extracto S (interfase) N. Eritr. tiene un tamaño promedio de loop ≈ otras células somáticas S: no cambia el tamaño medio del loop M: disminuye el tamaño medio del loop! ≈ tamaño del N.esperma en S
 Solución control. b) Extracto S (interfase) c) Extracto M Ca++ N. Esperma d) Extracto S (interfase) N. Eritr. tiene un tamaño promedio de loop ≈ otras células somáticas. S: no cambia el tamaño medio del loop. M: disminuye el tamaño medio del loop! ≈ tamaño del N.esperma en S.")
21
…Por lo tanto, hasta ahora sabemos que:
pasar por fase M (mitosis) induce 2 tipos de rearreglos en el N. eritrocito: Reduce el tamaño medio de los loop de DNA (esto sería consistente con una mayor cantidad de sitios de anclaje a la matriz nuclear) en proporciones paralelas Reduce el espacio medio entre orígenes de replicación Y, en ambos casos ≈ N.esperma en fase S
 induce 2 tipos de rearreglos en el N. eritrocito: Reduce el tamaño medio de los loop de DNA (esto sería. consistente con una mayor cantidad de sitios de anclaje a la. matriz nuclear) en proporciones. paralelas. Reduce el espacio medio entre orígenes de replicación. Y, en ambos casos ≈ N.esperma en fase S.")
22
Esperma + ovocito en M Condensan cromosomas (metafase)
topoisom II Esperma + ovocito en M Condensan cromosomas (metafase) Entonces, ¿Condensación cromosómica en M (vía topII) está involucrada en el reseteo del N.eritrocito? Ca ++ Esperma N.eritr. Ca ++ N.eritr ’ M con ICRF(inhhib. topoisom. II) S 45’ M S Ca ++ o directamente transferencia M S rápida rápida Eritr. requiere topII = condensación cromosómica se necesita NO de la transic M-S sino de la condens. de los cromos ( o sea, terminar metafase). pero sólo en eritr. no en sperm lenta lenta Ca ++ Esperma ’ M con ICRF S rápida
 Entonces, ¿Condensación cromosómica en M (vía topII) está involucrada en el reseteo del N.eritrocito Ca ++ Esperma. N.eritr. Ca ++ N.eritr. 45’ M con ICRF(inhhib. topoisom. II) S. 45’ M S. Ca ++ o directamente transferencia. M S. rápida. rápida. Eritr. requiere topII = condensación cromosómica. se necesita NO de la transic M-S sino de la condens. de los cromos ( o sea, terminar metafase). pero sólo en eritr. no en sperm. lenta. lenta. Ca ++ Esperma 45’ M con ICRF S. rápida.")
23
Esperma desmembranados, incubados en extracto M +
N. Eritr ICRF; tinción de DNA con Hoechst Medición de actividad de H1-kinasa Extractos con ICRF estaban en mitosis

24
Esperma 45’ M purif.cromatina + PAGE de proteínas: IB con ac.anti-ORC2
¿Organización cromosómica dependiente de topoisomerasa II controla eficiencia del reclutamiento de ORC a la cromatina? Esperma ’ M purif.cromatina + PAGE de proteínas: IB con ac.anti-ORC2 N. Eritr Ca ++ 30’ 45’ M + ICRF S purif.cromatina + PAGE de proteínas: IB con ac.anti-ORC2 ORC2 no se recluta ORC2 no se recluta en ICRF + ORC2 se recluta en ICRF + y - El remodelado, en M, de los cromosomas dependiente de topo II, en N.eritr., se requiere para el reclutamiento apropiado de proteínas ORC en S Remodelado en M dependiente de topo.II se requiere, en eritrocitos, para el reclutamiento de ORC en S

25
Xenopus sp. Se va acumulando DNA en las sucesivas divisiones, en estos extractos in vitro
Esperma (a distintas conc. de DNA) ’ S purif.cromatina + PAGE de N.eritr proteínas: IB con ac.anti-ORC2 Se alcanza el máximo reclutamiento de ORC2 con 25ng de DNA Con 50ng de DNA aún no se alcanzó el máximo reclutamiento de ORC2 Reclutamiento de factores de iniciación no sólo es proporcional a la cantidad de cromatina disponible; también depende de la naturaleza de la misma
 30’ S purif.cromatina + PAGE de N.eritr. proteínas: IB con ac.anti-ORC2. Se alcanza el máximo reclutamiento de ORC2 con 25ng de DNA. Con 50ng de DNA aún no se alcanzó el máximo reclutamiento de ORC2. Reclutamiento de factores de iniciación no sólo es proporcional a la cantidad de cromatina disponible; también depende de la naturaleza de la misma.")
26
Disminución de los espacios entre replicones (inter-orígenes).
¿Qué elementos producirían la reorganización de la cromatina en el remodelado mitótico, hasta ahora? Disminución de los espacios entre replicones (inter-orígenes). Disminución en el tamaño del DNA loop por acción de la Topoisomerasa II; relacionado con un aumento del numero de sitios de anclaje no específico (random) del DNA a la matriz nuclear. Reclutamiento de factores de iniciación dependiente de la estructura cromosómica.
. Disminución en el tamaño del DNA loop por acción de la Topoisomerasa II; relacionado con un aumento del numero de sitios de anclaje no específico (random) del DNA a la matriz nuclear. Reclutamiento de factores de iniciación dependiente de la estructura cromosómica.")
27
Estos mismos, ¿ocurrirán durante el desarrollo temprano de los embriones ?!!!!!
IN VIVO

28
Remodelado de la estructura cromatínica por el ciclo celular en embriones tempranos de Xenopus.
Cariomeros en transición anafase-telofase e iniciando replicación del DNA antes de la reconstrucción del núcleo.

29
Núcleos POST-R de embriones.
Núcleos PRE-R de embriones (CARIOMEROS). Extracción de DNA manteniendo los sitios de anclaje a la matriz nuclear. Sondas de DNA (post-R/pre-R) para hibridar con dominios de rDNA.
. Extracción de DNA manteniendo los sitios de anclaje a la matriz nuclear. Sondas de DNA (post-R/pre-R) para hibridar con dominios de rDNA.")
30
Dominio rDNA de Xenopus.
Remodelado de la estructura cromatínica por el ciclo celular en embriones tempranos de Xenopus. Dominio rDNA de Xenopus. Electroforesis en geles de agarosa. Bromuro de etidio/ membranas de nylon. Hibridizado con DNA sonda total de Xenopus o MARs de cariomeros o MARs de núcleos post-replicativos de embriones.

31
IN VIVO también se observa la reorganización de la
estructura cromatínica luego de pasar por mitosis. Y ocurre en cada división del embrión temprano. Existen un gran nº de puntos de anclaje, aleatorios y espaciados cercanamente, entre la cromatina y la matriz nuclear que están involucrados en la rápida replicación del DNA en éste estadio de Xenopus. Entonces, mitosis reduce el tamaño del DNA loop. Cromatina está preparada para la fase S…. ¿?

32
Medición del DNA-loop por técnica del máximo halo fluorescente (MFHT)
Similar a Eryt + M + CaCl2 Similar al tamaño de replicón. Existencia de un proceso de REMODELADO POST- REPLICATIVO, que aumenta el espacio entre los sitios de anclaje a la matriz nuclear y se evidencia en el aumento del tamaño del DNA-loop.

33
Núcleos POST-R que NO pasaron por MITOSIS
Remodelado mitótico de los dominios Loop de la cromatina ocurre durante las fases S y M. Si la disminución del tamaño del DNA-loop (mitotic resetting of chromatin) fuese esencial para tasa de replicación. Núcleos POST-R que NO pasaron por MITOSIS tasa de replicación del DNA.
 fuese esencial para tasa de replicación. Núcleos POST-R que NO pasaron por MITOSIS. tasa de replicación del DNA.")
34
Remodelado mitótico de los dominios Loop de la cromatina ocurre durante las fases S y M.

35
Se confirma BAJA T. de REPLICACIÓN.
Remodelado mitótico de los dominios Loop de la cromatina ocurre durante las fases S y M. Similar al núcleo de Eritrocitos en fase S. Se confirma BAJA T. de REPLICACIÓN.

36
Replicones MUY CERCANOS
Remodelado mitótico de los dominios Loop de la cromatina ocurre durante las fases S y M. Sperm N: Shift a valores de alto PM. La unión del DNA replicado de REPLICONES adyacentes. Replicones MUY CERCANOS Post-R N: no se observa shift. Replicones ESPACIADOS

37
Unión del replicón y ORC2 en núcleos de esperma o núcleos POST-R de embriones.
La tasa de replicación del DNA durante el desarrollo temprano, depende de la organización del cromosoma y NO de la cantidad absoluta de ORC.

38
Esto es lo que estaría pasando IN VIVO …

39
¿El aumento del tamaño del Loop, es un evento post-replicativo u ocurre durante la síntesis del DNA?

40
Remodelado mitótico de los dominios Loop de la cromatina ocurre durante las fases S y M.

41
Aumento gradual del tamaño del Loop durante fase S.
Remodelado mitótico de los dominios Loop de la cromatina ocurre durante las fases S y M. Aumento gradual del tamaño del Loop durante fase S.

42
Si incubamos los núcleos POST-R de embriones de Xenopus en extractos mitóticos … ¿Se reseteara el tamaño del Loop?

43
Tamaño Loop se reduce sin inhibidor de la Topoisomerasa II.
Remodelado mitótico de los dominios Loop de la cromatina ocurre durante las fases S y M. Tamaño Loop se reduce sin inhibidor de la Topoisomerasa II.

44
Núcleos diferenciados (eritrocitos)
MITOSIS Disminuye tamaño del DNA Loop RESETEO DE LA CROMATINA Núcleos POST-R de embriones de Xenopus Núcleos diferenciados (eritrocitos)
")
45
DISCUSIÓN

46
Organización dinámica de estructuras nucleares para replicación del DNA durante desarrollo

47
Remodelado mitótico (RM) ocurre en ausencia de ORC, o sea, independientemente del pre-RC.
RM provoca disminución del tamaño del Loop, permitiendo aumento del reclutamiento de ORC a la cromatina. Mayor nº de sitios de unión de ORC, se corresponde con mayor nº de sitios de anclaje a la matriz. Podría explicar el aumento en la densidad de orígenes de replicación. (En Xenopus ORC se asocia preferentemente a regiones ricas en A-T, que coinciden con los sitios de asociación a la matriz). El éxito de la clonación aumenta con “serial transfer”, los resultados obtenidos en este trabajo apoyan eso ( formación de cromosomas mitóticos permiten rápida replicación). Modelo antes propuesto: reloj citoplasmático de Xenopus, las células se dividen cada 30 min. Núcleos sin reprogramación genética no siguen ritmo del ciclo celular. Reseteo de la organización del núcleo dado por el pasaje por Metafase (45 min) y no por la transición de Metafase a Anafase. Relacionado con la organización de los cromosomas en esa etapa (disminuye el tamaño del loop y del espacio interorigen; no afecta la cantidad de factores pre-RC; topoisomerasa II necesaria para reclutamiento de ORC).
. El éxito de la clonación aumenta con serial transfer , los resultados obtenidos en este trabajo apoyan eso ( formación de cromosomas mitóticos permiten rápida replicación). Modelo antes propuesto: reloj citoplasmático de Xenopus, las células se dividen cada 30 min. Núcleos sin reprogramación genética no siguen ritmo del ciclo celular. Reseteo de la organización del núcleo dado por el pasaje por Metafase (45 min) y no por la transición de Metafase a Anafase. Relacionado con la organización de los cromosomas en esa etapa (disminuye el tamaño del loop y del espacio interorigen; no afecta la cantidad de factores pre-RC; topoisomerasa II necesaria para reclutamiento de ORC).")
48
En Xenopus: Núcleo de esperma y ovocito se replican antes de cariogamia, tienen que estar preparados para una rápida replicación, antes de la primer división mitótica. De hecho, antes de la fertilización … el núcleo del ovocito está bloqueado en metafase y sus cromosomas mitóticos organizados el núcleo de esperma está organizado en Loops cortos Esto indica que son los huevos no fertilizados los capaces de preparar núcleos diferenciados para el desarrollo

Presentaciones similares










>")








