Descargar la presentación
La descarga está en progreso. Por favor, espere

1
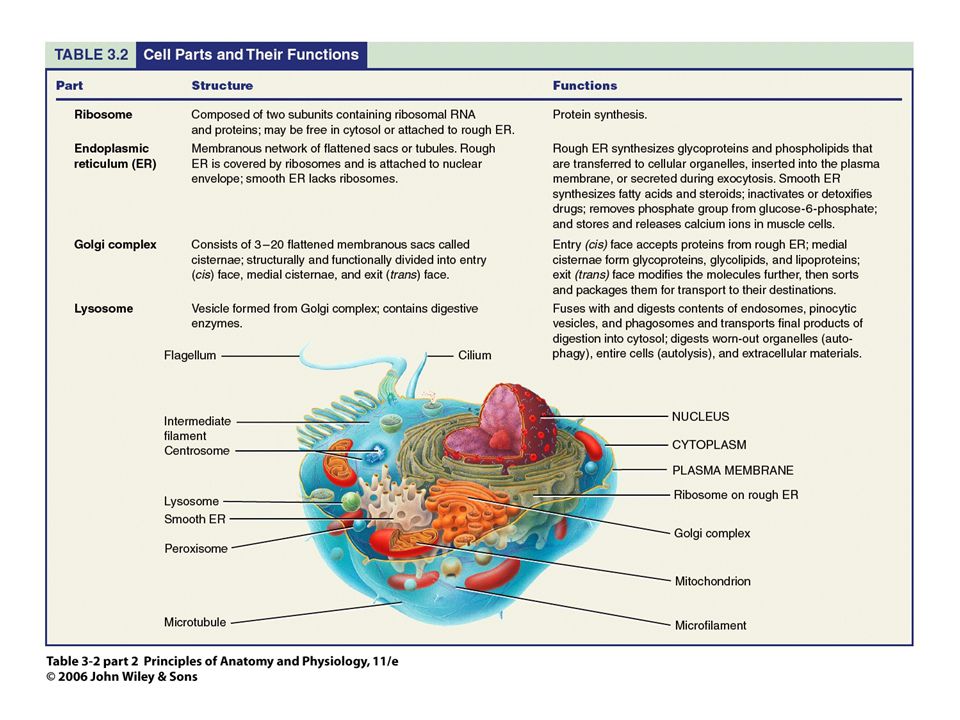
La célula Unidad viva básica del organismo Nº total ~ 100 billones
Características similares: metabolismo, consumo de O2, división celular… Organización de la célula Núcleo: ADN empaquetado (cromatina) rodeado de membrana nuclear Citoplasma (rodeado de membrana celular)
 rodeado de membrana nuclear. Citoplasma (rodeado de membrana celular)")
2
La célula Membranas Compuestas por: Tipos:
Lípidos (fosfolípidos, colesterol) y proteínas Tipos: Membrana plasmática Membrana nuclear Membrana del retículo endoplásmico Membrana mitocondrial Lisosomas Aparato de Golgi
 y proteínas. Tipos: Membrana plasmática. Membrana nuclear. Membrana del retículo endoplásmico. Membrana mitocondrial. Lisosomas. Aparato de Golgi.")
3
1. Membrana plasmática Barrera física entre el LIC y el LEC
Otras funciones: transporte, comunicación, reconocimiento, adhesión En este tema nos vamos a centrar en el estudio de la membrana plasmática. Si pensamos que tanto el medio intracelular como el extracelular son compartimentos acuosos nos daremos cuenta de la importancia de existencia de una barrera física que impida la salida o la pérdida de enzimas, nucleótidos, otras moléculas hidrosolubles y, en definitiva, que mantenga la diferente composición existente entre ambos compartimentos. Evidentemente esta barrera no puede estar constituida por moléculas hidrosolubles, de hecho la composición de la membrana celular es lipídica. De hecho, la composición de todas las membranas que rodean a los orgánulos están constituidas principalmente pos fosfolípidos y proteínas. Los fosfolípidos se caracterizan por poseer un grupo fosfato. El ejemplo más común de molécula fosfolipídica es el constituido por la unión de dos moléculas de ácido graso a dos de los átomos de carbono del glicerol. El tercer átomo de carbono se une al grupo fosfato, el cual puede unirse a otras moléculas, de este modo si se une a una molécula de colina la molécula generada es la fosfatidilcolina. De este modo dentro de la molécula se describen dos regiones una polar o hidrófila y otra apolar o hidrófoba (moléculas anfipáticas). En el entorno acuoso en el que se encuentran estos fosfolípidos hace que alejen, en todo lo posible, las cadenas de ácidos grasos del contacto con el agua por lo que forman bicapas lipídicas. Prácticamente todas las moléculas de fosfolípidos de las membranas biológicas tienen estructura de bicapa lipídica, de forma que la porción polar queda en contacto con el agua a la apolar resguardada de ese contacto. La zona media hidrófoba de la membrana es la que impide el paso de moléculas e iones hidrosulobles. Pero sin embargo sabemos que existen moléculas polares que atraviesan la membrana. Esta propiedad selectiva y especializada del transporte de membrana se deben al componente proteico de las mismas.
. En el entorno acuoso en el que se encuentran estos fosfolípidos hace que alejen, en todo lo posible, las cadenas de ácidos grasos del contacto con el agua por lo que forman bicapas lipídicas. Prácticamente todas las moléculas de fosfolípidos de las membranas biológicas tienen estructura de bicapa lipídica, de forma que la porción polar queda en contacto con el agua a la apolar resguardada de ese contacto. La zona media hidrófoba de la membrana es la que impide el paso de moléculas e iones hidrosulobles. Pero sin embargo sabemos que existen moléculas polares que atraviesan la membrana. Esta propiedad selectiva y especializada del transporte de membrana se deben al componente proteico de las mismas.")
4
Fosfolípidos mayoritarios en las membranas eucariotas
1. Membrana plasmática Fosfolípidos mayoritarios en las membranas eucariotas Hay cuatro tipos de fosfolípidos en la membrana celular: fosfatidilcolina esfingomielina (en este fosfolípido la glicerina ha sido sustituída por un aminoalcohol llamado D-4-esfingenina) fosfatidilserina fosfatidiletanolamina La zona media hidrófoba de la membrana es la que impide el paso de moléculas e iones hidrosulobles. Pero sin embargo sabemos que existen moléculas polares que atraviesan la membrana. Esta propiedad selectiva y especializada del transporte de membrana se deben al componente proteico de las mismas. Note that only phosphatidylserine carries a net negative charge, the importance of which we discuss later; the other three are electrically neutral at physiological pH, carrying one positive and one negative charge. Together these four phospholipids constitute more than half the mass of lipid in most membranes. Other phospholipids, such as the inositol phospholipids, are present in smaller quantities but are functionally very important. The inositol phospholipids, for example, have a crucial role in cell signaling.
 fosfatidilserina. fosfatidiletanolamina La zona media hidrófoba de la membrana es la que impide el paso de moléculas e iones hidrosulobles. Pero sin embargo sabemos que existen moléculas polares que atraviesan la membrana. Esta propiedad selectiva y especializada del transporte de membrana se deben al componente proteico de las mismas. Note that only phosphatidylserine carries a net negative charge, the importance of which we discuss later; the other three are electrically neutral at physiological pH, carrying one positive and one negative charge. Together these four phospholipids constitute more than half the mass of lipid in most membranes. Other phospholipids, such as the inositol phospholipids, are present in smaller quantities but are functionally very important. The inositol phospholipids, for example, have a crucial role in cell signaling.")
5
1. Membrana plasmática COLESTEROL La fluidez de la membrana depende de su composición. En este aspecto el colesterol actúa como un amortiguador de la fluidez. El colesterol es un constituyente fundamental en las membranas celulares. Está constituido por una cabeza polar, un núcleo esteroideo, el cual se dispone paralelo a las cadenas de ácidos grasos de los fosfolípidos de la membrana y una cola hidrocarbonada. Si nos fijamos en el esquema veremos como las moléculas de colesterol se encuentran orientadas en la bicapa de forma que los grupos hidroxilos se encuentran próximos a las cabezas polares de los fosfolípidos. En esta posición el núcleo de anillos esteroideos interacciona e inmoviliza parcialmente las cadenas de hidrocarbono más cercanas a las cabezas polares. Al disminuir la movilidad de estos primeros grupos de CH2 de las cadenas de fosfolípidos el colesterol hace que, en esta región la membrana sea menos deformable y disminuye la permeabilidad de la bicapa a las moléculas de agua. De este modo puede decirse que el colesterol actúa como una amortiguador de la fluidez de la membrana. Las membranas eucariotas presentan grandes cantidades de colesterol (hasta una molécula de colesterol por cada una de fosfolípido, Although cholesterol tends to make lipid bilayers less fluid, at the high concentrations found in most eucaryotic plasma membranes, it also prevents the hydrocarbon chains from coming together and crystallizing. In this way, it inhibits possible phase transitions.) Además de los fosfolípidos, las proteínas y el colesterol existe otro integrante de las membranas celulares, los glicolípidos. El colesterol amortigua la fluidez de la MP (= menos deformable) Disminuye la permeabilidad de la MP al agua
 Además de los fosfolípidos, las proteínas y el colesterol existe otro integrante de las membranas celulares, los glicolípidos. El colesterol amortigua la fluidez de la MP (= menos deformable) Disminuye la permeabilidad de la MP al agua.")
6
1. Membrana plasmática La mayoría de las membranas celulares constituyen un “mosaico fluido” de fosfolípidos y proteínas. Los componentes proteicos de la membrana pueden dividirse en proteínas periféricas y proteínas integrales. Debido a que la membrana celular no es sólida y en su interior, fosfolípidos y proteínas pueden desplazarse lateralmente. Ello origina que las proteínas no presenten una distribución uniforme, sino que la distribución de las mismas presenta un patrón en mosaico que cambia constantemente la que se ha denominado modelo de mosaico fluido. Aunque la mayoría de lípidos y proteínas pueden moverse libremente por en el plano de la membrana el intercambio entre las monocapas es poco frecuente (es poco probable que una porción hidrofílica grande cambie de monocapa si tiene que ser arrastrada a través del interior no polar de la bicapa lipídica). Existen excepciones en los que los componentes de la membrana no pueden difundir libremente, por ejemplo, el secuestro de los receptores de acetilcolina (proteína integral de membrana) en la placa motora terminal de músculo esquelético. En este tipo de distribución el citoesqueleto parece fijar ciertas proteínas de membrana. Pero no sólo las proteínas desempeñan funciones celulares importantes. Determinados fosfolípidos presentes en diminutas proporciones en la membrana plasmática tienen importantes funciones en la transducción de señales. El bifosfato de fosfatidilinositol, cuando se hidroliza por una fosfolipasa C activada por un receptor, libera inositol 1,4,5-trifosfato (IP3) y diacilglicerol. El IP3 se libera en el citosol, donde actúa sobre receptores del retículo endoplásmico para provocar la liberación de Ca2+, lo que afecta a muchos procesos celulares. El diacilglicerol permanece en la membrana, donde junto con el Ca2+ interviene en la activación de la proteinkinasa C, una proteína muy importante en la transducción de señales. La fluidez de la membrana depende de su composición. En este aspecto el colesterol actúa como un amortiguador de la fluidez.
. Existen excepciones en los que los componentes de la membrana no pueden difundir libremente, por ejemplo, el secuestro de los receptores de acetilcolina (proteína integral de membrana) en la placa motora terminal de músculo esquelético. En este tipo de distribución el citoesqueleto parece fijar ciertas proteínas de membrana. Pero no sólo las proteínas desempeñan funciones celulares importantes. Determinados fosfolípidos presentes en diminutas proporciones en la membrana plasmática tienen importantes funciones en la transducción de señales. El bifosfato de fosfatidilinositol, cuando se hidroliza por una fosfolipasa C activada por un receptor, libera inositol 1,4,5-trifosfato (IP3) y diacilglicerol. El IP3 se libera en el citosol, donde actúa sobre receptores del retículo endoplásmico para provocar la liberación de Ca2+, lo que afecta a muchos procesos celulares. El diacilglicerol permanece en la membrana, donde junto con el Ca2+ interviene en la activación de la proteinkinasa C, una proteína muy importante en la transducción de señales. La fluidez de la membrana depende de su composición. En este aspecto el colesterol actúa como un amortiguador de la fluidez.")
7
1. Membrana plasmática Las características funcionales de la MP dependen de las proteínas que contiene. Muchas proteínas de membrana son glucoproteínas. Tipos (por la forma en la que están dispuestas en la MP): - Periféricas: incluidas de manera parcial en una de las superficies de la membrana, unidas covalentemente a lípidos o asociadas a ellos mediante un dominio hidrofóbico. - Integrales: abarcan todo el espesor de la membrana. Son anfipáticas. Aunque la estructura básica de la membrana celular sea la bicapa lipídica la mayoría de funciones específicas se llevan a cabo mediante la acción de proteínas. Su tipo y cantidad son muy variables en función del tipo celular. Como los lípidos, las proteínas de membrana a menudo se encuentran asociadas a cadenas de oligosacáridos (presentes en el exterior celular). De este modo la superficie celular presenta una cubierta rica en carbohidratos, el glicocálix, que veremos posteriormente. Existen diferentes formas de asociación entre la bicapa lipídica y las proteínas: Proteínas transmembrana (1,2,3,): Pueden atravesar completamente la bicapa, teniendo porciones de la proteína en contacto con el exterior y el interior celular. Al igual que sus vecinos los lípidos, estas proteínas transmembrana son moléculas anfipáticas (la región hidrofóbica se localiza en el interior de la membrana e interacciona con las colas hidrofóbicas de los lípidos). La hidrofobicidad de algunas de estas proteínas se aumenta por la formación de interacciones covalentes con cadenas de ácidos grasos que se insertan desde la monocapa citosólica (1). Proteínas localizadas en el citosol (4, 5, 7): ya sea por asociación con la monocapa lipídica por una hélice alfa anfipática (4) o por una o más uniones covalentes lipídicas por ejemplo por cadena de ácidos grasos (5) (son proteínas traducidas en citosol). Otras proteínas sólo se localizan en la superficie celular unidas mediante un oligosacárido por un puente covalente al fosfatidilinositol (6)(son proteinas traducidas en retículo endoplasmático). Otras proteínas se unen a la membrana mediante interacciones no covalentes con otras proteínas. Se les deomina proteínas periféricas de membrana (pueden liberarse de la membrana exponiendo a las células a medios hipo o hipertónicos o modificando el pH). El resto de proteínas transmembrana o ancladas a la bicapa mediante grupos lipídicos o con otras proteínas mediante inusuales uniones estrechas (pero que no pueden separarse por estos medios) se denominan proteínas integrales. El tipo de unión en la membrana refleja la función de la proteína. Solo las transmembrana actúan a ambos lados de la membrana o pueden transportar moléculas a su través. Los receptores de superficie son proteínas transmembrana que unen moléculas señal en el exterior celular y generan señales intracelulares. Las proteínas que sólo funcionan en uno de los lados normalmente se asocian con otras proteínas o lípidos que también actúan en ese lado. (esto es una tontería). La mayoría de las proteínas transmembrana atraviensan la bicapa como una alpha hélice simple (1), a hélice multiple (2) o como barriles β (3). Además las proteínas siempre tiene una única orientación en la membrana. Esta asimetría refleja el proceso de síntesis e inserción desde RE y las diferentes funciones de dominios extra e intracelulares. Los residuos proteicos que atraviesan la bicapa lipídica son aminoácidos con cadenas laterales no polares. Debido a que en enlace peptídico es polar y al ambiente hidrofóbico en que se encuentran estos enlaces peptídicos están dirigidos por puentes de hidrógeno, función de los cuales se optimiza al formarse la alpha hélice. Los barriles beta son más rígidos y cristalizan más rápido que las alpha hélices. El número de cadenas que atraviesan la membran puede oscilar de 8 a 22 y son abundantes en mitocondria, Cloroplastos y bacterias. Algunas forman poros llenos de agua que posibilitan el paso de sustancias hidrofílicas, otras actúan como receptores o incluso como enzimas (mayoritariamente restringidos a mitocondria, Cloroplastos y bacterias). En células eucariotas la mayoría de proteínas transmembrana son alpha hélice. The helices within these proteins can slide against each other, allowing the protein to undergo conformational changes that can be exploited to open and shut ion channels, transport solutes, or transduce extracellular signals into intracellular ones. In β barrel proteins, by contrast, each β strand is bound rigidly to its neighbors by hydrogen bonds, making conformational changes of the barrel itself unlikely. Al igual que los lípidos de membrana las proteínas pueden estar glicosiladas. De hecho, la gran mayoría de las proteínas en células animaleslo están. Al igual que ocurría en lípidos los oligosacátidos se añaden en el lumen de retículo y golgi. Por esta razón los oligos siempre se encuentran en la cara externa de la membrana. Otra diferencia que existe entre la cara citosólica y la externa en las proteínas de membrana es que el citosol es un ambiente reductor por lo que en la cara externa se formarán intra y entre proteínas puentes disulfuro entre la cys. Estos puentes son muy importantes en el mantenimiento de la conformación estructural o en la asociación con otros polipéptidos.
: - Periféricas: incluidas de manera parcial en una de las superficies de la membrana, unidas covalentemente a lípidos o asociadas a ellos mediante un dominio hidrofóbico. - Integrales: abarcan todo el espesor de la membrana. Son anfipáticas. Aunque la estructura básica de la membrana celular sea la bicapa lipídica la mayoría de funciones específicas se llevan a cabo mediante la acción de proteínas. Su tipo y cantidad son muy variables en función del tipo celular. Como los lípidos, las proteínas de membrana a menudo se encuentran asociadas a cadenas de oligosacáridos (presentes en el exterior celular). De este modo la superficie celular presenta una cubierta rica en carbohidratos, el glicocálix, que veremos posteriormente. Existen diferentes formas de asociación entre la bicapa lipídica y las proteínas: Proteínas transmembrana (1,2,3,): Pueden atravesar completamente la bicapa, teniendo porciones de la proteína en contacto con el exterior y el interior celular. Al igual que sus vecinos los lípidos, estas proteínas transmembrana son moléculas anfipáticas (la región hidrofóbica se localiza en el interior de la membrana e interacciona con las colas hidrofóbicas de los lípidos). La hidrofobicidad de algunas de estas proteínas se aumenta por la formación de interacciones covalentes con cadenas de ácidos grasos que se insertan desde la monocapa citosólica (1). Proteínas localizadas en el citosol (4, 5, 7): ya sea por asociación con la monocapa lipídica por una hélice alfa anfipática (4) o por una o más uniones covalentes lipídicas por ejemplo por cadena de ácidos grasos (5) (son proteínas traducidas en citosol). Otras proteínas sólo se localizan en la superficie celular unidas mediante un oligosacárido por un puente covalente al fosfatidilinositol (6)(son proteinas traducidas en retículo endoplasmático). Otras proteínas se unen a la membrana mediante interacciones no covalentes con otras proteínas. Se les deomina proteínas periféricas de membrana (pueden liberarse de la membrana exponiendo a las células a medios hipo o hipertónicos o modificando el pH). El resto de proteínas transmembrana o ancladas a la bicapa mediante grupos lipídicos o con otras proteínas mediante inusuales uniones estrechas (pero que no pueden separarse por estos medios) se denominan proteínas integrales. El tipo de unión en la membrana refleja la función de la proteína. Solo las transmembrana actúan a ambos lados de la membrana o pueden transportar moléculas a su través. Los receptores de superficie son proteínas transmembrana que unen moléculas señal en el exterior celular y generan señales intracelulares. Las proteínas que sólo funcionan en uno de los lados normalmente se asocian con otras proteínas o lípidos que también actúan en ese lado. (esto es una tontería). La mayoría de las proteínas transmembrana atraviensan la bicapa como una alpha hélice simple (1), a hélice multiple (2) o como barriles β (3). Además las proteínas siempre tiene una única orientación en la membrana. Esta asimetría refleja el proceso de síntesis e inserción desde RE y las diferentes funciones de dominios extra e intracelulares. Los residuos proteicos que atraviesan la bicapa lipídica son aminoácidos con cadenas laterales no polares. Debido a que en enlace peptídico es polar y al ambiente hidrofóbico en que se encuentran estos enlaces peptídicos están dirigidos por puentes de hidrógeno, función de los cuales se optimiza al formarse la alpha hélice. Los barriles beta son más rígidos y cristalizan más rápido que las alpha hélices. El número de cadenas que atraviesan la membran puede oscilar de 8 a 22 y son abundantes en mitocondria, Cloroplastos y bacterias. Algunas forman poros llenos de agua que posibilitan el paso de sustancias hidrofílicas, otras actúan como receptores o incluso como enzimas (mayoritariamente restringidos a mitocondria, Cloroplastos y bacterias). En células eucariotas la mayoría de proteínas transmembrana son alpha hélice. The helices within these proteins can slide against each other, allowing the protein to undergo conformational changes that can be exploited to open and shut ion channels, transport solutes, or transduce extracellular signals into intracellular ones. In β barrel proteins, by contrast, each β strand is bound rigidly to its neighbors by hydrogen bonds, making conformational changes of the barrel itself unlikely. Al igual que los lípidos de membrana las proteínas pueden estar glicosiladas. De hecho, la gran mayoría de las proteínas en células animaleslo están. Al igual que ocurría en lípidos los oligosacátidos se añaden en el lumen de retículo y golgi. Por esta razón los oligos siempre se encuentran en la cara externa de la membrana. Otra diferencia que existe entre la cara citosólica y la externa en las proteínas de membrana es que el citosol es un ambiente reductor por lo que en la cara externa se formarán intra y entre proteínas puentes disulfuro entre la cys. Estos puentes son muy importantes en el mantenimiento de la conformación estructural o en la asociación con otros polipéptidos.")
8
1. Membrana plasmática Ejemplos de estructuras de proteínas de membrana Barriles formados por diferente número de cadenas que configuran un canal o poro Segmento hidrófobo Glicosilación de proteínas y formación depuentes disulfuro entre cisteínas

9
La célula Citoplasma Agua Proteínas disueltas
Electrolitos (Na+, Cl-, K+…) Glucosa Pequeñas cantidades de compuestos lipídicos Fibrillas de actina, tubulina… (citoesqueleto) Glóbulos de lípidos, gránulos de glucógeno, vesículas secretoras Orgánulos: Ribosomas, Retículo endoplásmico, aparato de Golgi, mitocondrias, lisosomas.
 Glucosa. Pequeñas cantidades de compuestos lipídicos. Fibrillas de actina, tubulina… (citoesqueleto) Glóbulos de lípidos, gránulos de glucógeno, vesículas secretoras. Orgánulos: Ribosomas, Retículo endoplásmico, aparato de Golgi, mitocondrias, lisosomas.")
10
Retículo Endoplasmico
La célula Retículo Endoplasmico Estructuras tubulares aplanadas interconectadas que rodean al núcleo Paredes formadas por bicapa lipídica membranosa El interior contiene la matriz endoplásmica. 2 tipos Con ribosomas: retículo endoplásmico rugoso (granular) Sin ribosomas: retículo endoplásmico agranular o liso
 Sin ribosomas: retículo endoplásmico agranular o liso.")
11
Ribosomas y retículo endoplásmico rugoso
La célula Ribosomas y retículo endoplásmico rugoso Ribosomas: Se encuentran anclados a las superficies externas de muchas regiones del retículo endoplásmico rugoso Compuestos por una mezcla de ARNr y proteínas Encargados de la síntesis proteica.

12
Retículo endoplásmico agranular o liso (REL)
La célula Retículo endoplásmico agranular o liso (REL) Carece de ribosomas acoplados Actúa en la síntesis de sustancias lipídicas, en el procesamiento de las proteínas y otros procesos enzimáticos celulares (p.e. glicosilación de proteínas, detoxificación de xenobióticos).
 Carece de ribosomas acoplados. Actúa en la síntesis de sustancias lipídicas, en el procesamiento de las proteínas y otros procesos enzimáticos celulares (p.e. glicosilación de proteínas, detoxificación de xenobióticos).")
13
La célula Aparato de Golgi
Intimamente relacionado con el retículo endoplásmico Posee membranas celulares parecidas a las del REL Consta de un apilamiento varias capas de vesículas cerradas y planas. Es importante en células secretoras. Estas expulsan las sustancias a secretar (p.e., hormonas). Conjuntamente con el retículo endoplásmico forman los lisosomas, vesículas secretoras u otros componentes citoplasmáticos.
. Conjuntamente con el retículo endoplásmico forman los lisosomas, vesículas secretoras u otros componentes citoplasmáticos.")
14
La célula Lisosomas Orgánulos vesiculares formados en el aparato de golgi y dispersas a lo largo del citoplasma. Proporcionan un sistema digestivo intracelular (sustancias y estructuras intracelulares, especialmente las dañadas, partículas alimenticias ingeridas por las celulas, bacterias…)
")
15
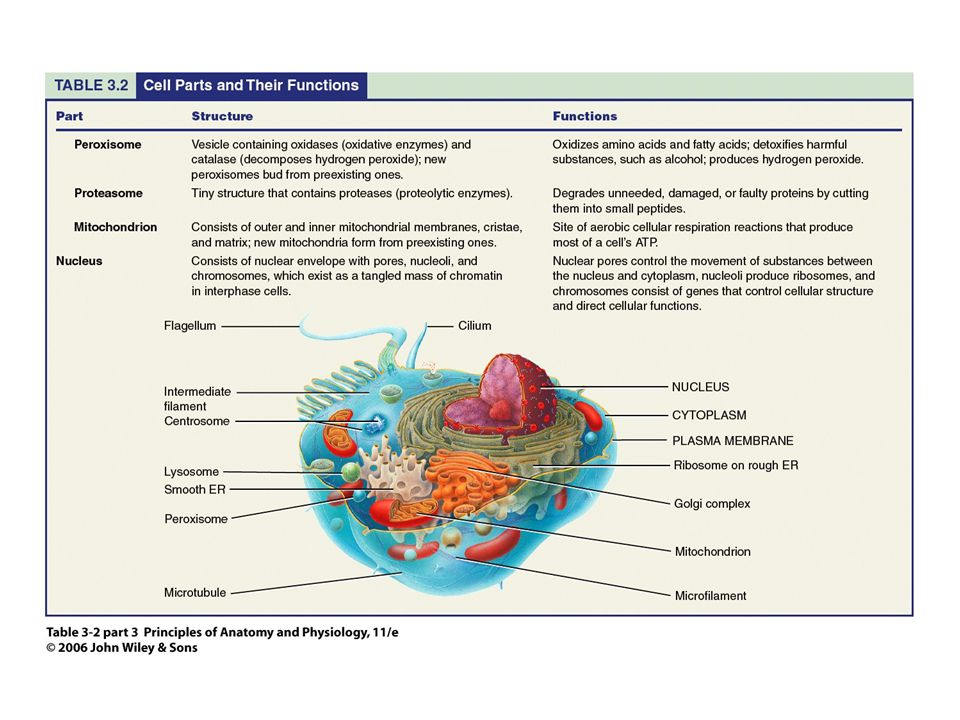
La célula Mitocondria Centrales energéticas
Respiración celular (oxidación de glucosa, AG y AAs) Cadena de transporte de electrones Síntesis de ATP: moneda energética celular
 Cadena de transporte de electrones. Síntesis de ATP: moneda energética celular.")
19
Diferenciación celular. Tejidos y órganos.
Las células han de diferenciarse para ejercer su función Diferenciación: cambios estructurales y funcionales de las células a medida que proliferan en el embrión para formar los diferentes tejidos y órganos. Consecuencia de la represión selectiva del genoma. Una célula normal expresa menos de la mitad de su genoma (nº genes ~ 25000). Células con funciones similares se agrupan formando TEJIDOS (NERVIOSO, EPITELIAL, MUSCULAR, CONJUNTIVO). Los ORGANOS están normalmente formados por los 4 tipos de tejido
. Células con funciones similares se agrupan formando TEJIDOS (NERVIOSO, EPITELIAL, MUSCULAR, CONJUNTIVO). Los ORGANOS están normalmente formados por los 4 tipos de tejido.")
20
Diferenciación celular. Tejidos y órganos.
Tejido muscular Miocitos especializados en la contracción→ función mecánica

21
Diferenciación celular. Tejidos y órganos.
Tejido epitelial Las células epiteliales forman estructuras membranosas. Funciones: - Revestimiento (epitelios simples o estratificados). - Secretoras ( glándulas de moco, hormonas…).
. - Secretoras ( glándulas de moco, hormonas…).")
22
Diferenciación celular. Tejidos y órganos.
Tejido conjuntivo (conectivo) Conjunto heterogéneo de tejidos derivados del mesodermo, formados por células rodeadas de grandes cantidades de material extracelular Función de sostén y separación de los diferentes elementos tisulares y también se convierte en un medio logístico (p.e. plasma de la sangre). Tipos: - No especializado (TC propiamente dicho) - Especializado (adiposo, cartílago, hueso, médula ósea, sangre)
 Conjunto heterogéneo de tejidos derivados del mesodermo, formados por células rodeadas de grandes cantidades de material extracelular. Función de sostén y separación de los diferentes elementos tisulares y también se convierte en un medio logístico (p.e. plasma de la sangre). Tipos: - No especializado (TC propiamente dicho) - Especializado (adiposo, cartílago, hueso, médula ósea, sangre)")
23
Diferenciación celular. Tejidos y órganos.
Tejido conjuntivo (conectivo)
")
24
Diferenciación celular. Tejidos y órganos.
Tejido Nervioso Neuronas, especializadas en generar y conducir impulsos eléctricos. Células de la glía: sostén

Presentaciones similares