Descargar la presentación
La descarga está en progreso. Por favor, espere

1
NATURALEZA DEL MATERIAL HEREDITARIO

2
Friedrich Miescher Aisló varias moléculas ricas en fosfatos, a las cuales llamó nucleínas (actualmente ácidos nucleicos), a partir del núcleo de los glóbulos blancos procedentes de vendajes quirúrjicos en 1869 El DNA fue aislado y estudiado por primera vez por el suizo Friedrich Miescher en En su investigación intentaba digerir proteínas de las células del pus, observando que el núcleo de esas células no era digerido, por lo que lo que había allí no eran proteínas, sino otra sustancia a la que llamó "nucleína" por su localización en el núcleo celular. Más tarde, al comprobarse su carácter ácido, recibió el nombre de "ácido nucleico".
, a partir del núcleo de los glóbulos blancos procedentes de vendajes quirúrjicos en El DNA fue aislado y estudiado por primera vez por el suizo Friedrich Miescher en En su investigación intentaba digerir proteínas de las células del pus, observando que el núcleo de esas células no era digerido, por lo que lo que había allí no eran proteínas, sino otra sustancia a la que llamó nucleína por su localización en el núcleo celular. Más tarde, al comprobarse su carácter ácido, recibió el nombre de ácido nucleico .")
3
El alemán Felix Hoppe-Seyler aisló un ácido nucleico de levaduras que difería en sus propiedades del ácido nucleico de Mieschler, al que se denominó ácido timonucleico (por su facilidad para ser extraído del timo de animales) para diferenciarlo del de levaduras. El alemán Felix Hoppe-Seyler aisló un ácido nucleico de levaduras que difería en sus propiedades del ácido nucleico de Mieschler, al que se denominó ácido timonucleico (por su facilidad para ser extraído del timo de animales) para diferenciarlo del de levaduras
 para diferenciarlo del de levaduras.")
4
Hacia 1890, Albrecht Kossel, hidrolizó el ácido nucleico, descubriendo la existencia de hidratos de carbono y de unos compuestos o bases nitrogenadas a las que dio los nombres de “adenina”, “guanina”, “citosina” y “timina”. Kossel recibió el premio Nobel de Fisiología y Medicina en 1910 Hacia 1890, el químico alemán Albrecht Kossel, hidrolizó el ácido nucleico, descubriendo la existencia de hidratos de carbono y de unos compuestos o bases nitrogenadas a las que dio los nombres de "adenina", "guanina", "citosina" y "timina". Kossel recibió el premio Nobel de Fisiología y Medicina en 1910.

5
En 1911, Theodore Leven, demostró que los hidratos de carbono eran pentosas, posteriormente se observó que el ácido nucleico de levadura poseía, como pentosa, la ribosa, mientras que el timonucleico poseía la desoxirribosa, recibiendo a partir de entonces los nombres de ácido RIBONUCLEICO (RNA), y ácido DESOXIRRIBONUCLEICO (DNA). Se descubrió también que el RNA no poseía timina, pero a cambio poseía uracilo

6
BASES NITROGENADAS

7
Streptococcus pneumoniae
Trabajos de Grifith (1928) Streptococcus pneumoniae El material empleado por Griffith (médico inglés) fue la bacteria Diplococcus pneumoniae o neumococo y los ratones. Cuando se inyecta a un ratón con el esputo de una persona enferma (con neumonía) dicho ratón muere de septicemia a las 24 horas. Esta capacidad virulenta de los neumococos se debe a la presencia de una cápsula de polisacáridos (polímeros de glucosa + ácido glucurónico) que envuelve a la bacteria y la protege de la fagocitosis. Para poder realizar cualquier estudio genético es necesaria la existencia de variabilidad para el carácter analizado. Griffith observó la existencia de diferentes tipos de neumococos: neumococos virulentos de tipo S que dan lugar a colonias con aspecto liso y brillante (producen la cápsula azucarada que los protege de la fagocitosis del huésped) y neumococos no virulentos (avirulentos) de tipo R que dan lugar a colonias de tipo rugoso y mate (carecen de la cápsula azucarada protectora). Colonia de tipo S Cepa S Cepa R
 Streptococcus pneumoniae. El material empleado por Griffith (médico inglés) fue la bacteria Diplococcus pneumoniae o neumococo y los ratones. Cuando se inyecta a un ratón con el esputo de una persona enferma (con neumonía) dicho ratón muere de septicemia a las 24 horas. Esta capacidad virulenta de los neumococos se debe a la presencia de una cápsula de polisacáridos (polímeros de glucosa + ácido glucurónico) que envuelve a la bacteria y la protege de la fagocitosis. Para poder realizar cualquier estudio genético es necesaria la existencia de variabilidad para el carácter analizado. Griffith observó la existencia de diferentes tipos de neumococos: neumococos virulentos de tipo S que dan lugar a colonias con aspecto liso y brillante (producen la cápsula azucarada que los protege de la fagocitosis del huésped) y neumococos no virulentos (avirulentos) de tipo R que dan lugar a colonias de tipo rugoso y mate (carecen de la cápsula azucarada protectora). Colonia de tipo S. Cepa S Cepa R.")
8
Cepa R Rugosas Sin cápsulas No virulentas Cepa S Lisas Con cápsula Virulentas Griffith nuca había observado que los neumococos RII (no virulentos) mutarán o cambiarán a SIII (virulentos). Griffith observó que si inyectaba a los ratones con neumococos de tipo RII (avirulentos) a las 24 horas seguían vivos (Figura A), mientras que si los inyectaba con neumococos SIII (virulentos) a las 24 horas los ratones morían (Figura B). Entonces decidió calentar los neumococos SIII (virulentos) para destruirlos y posteriormente inyectarlos a los ratones, encontrando que los ratones seguían vivos después de 24 horas (Figura C). Por consiguiente el calor, destruía el poder infectivo de los neumococos SIII. Por último, inyectó a los ratones una mezcla de neumococos RII vivos (no virulentos) y de SIII (virulentos) previamente muertos por calor, encontrado que los ratones morían a las 24 horas y extrayendo de su sangre neumococos SIII vivos Figura D). puesto que los neumococos RII (avirulentos) nunca mutan a SIII (virulentos), en el último experimento (Figura D) se demuestra la existencia de una sustancia presente en los extractos de neumococos SIII muertos por calor que es capaz de transformar a los neumocos RII vivos en SIII vivos, dicha sustancia fue denominada por Griffith el Principio Transformante. Esto sólo se podía explicar si algo de las lisas muertas había pasado a las rugosas vivas y las había transformado se llamó “principio transformante”. La cuestión era averiguar su naturaleza .
 mutarán o cambiarán a SIII (virulentos). Griffith observó que si inyectaba a los ratones con neumococos de tipo RII (avirulentos) a las 24 horas seguían vivos (Figura A), mientras que si los inyectaba con neumococos SIII (virulentos) a las 24 horas los ratones morían (Figura B). Entonces decidió calentar los neumococos SIII (virulentos) para destruirlos y posteriormente inyectarlos a los ratones, encontrando que los ratones seguían vivos después de 24 horas (Figura C). Por consiguiente el calor, destruía el poder infectivo de los neumococos SIII. Por último, inyectó a los ratones una mezcla de neumococos RII vivos (no virulentos) y de SIII (virulentos) previamente muertos por calor, encontrado que los ratones morían a las 24 horas y extrayendo de su sangre neumococos SIII vivos Figura D). puesto que los neumococos RII (avirulentos) nunca mutan a SIII (virulentos), en el último experimento (Figura D) se demuestra la existencia de una sustancia presente en los extractos de neumococos SIII muertos por calor que es capaz de transformar a los neumocos RII vivos en SIII vivos, dicha sustancia fue denominada por Griffith el Principio Transformante. Esto sólo se podía explicar si algo de las lisas muertas había pasado a las rugosas vivas y las había transformado se llamó principio transformante . La cuestión era averiguar su naturaleza .")
9
Estudios posteriores pusieron de manifiesto que la transformación de neumocos RII en SIII se podía realizar en tubo de ensayo sin necesidad de utilizar ratones en el experimento. Es decir, se puede mezclar en el mismo medio de cultivo líquido neumocos RII vivos con neumococos SIII previamente muertos por calor y obtener neumococos SIII vivos y virulentos

10
No sería hasta 1944 cuando Avery, McLeod y McCarthy repitieron los experimentos de Griffith y demostraron que el "Principio transformante" que convertía a las bacterias rugosas en lisas era, precisamente, el DNA. Avery McCarthy McLeod Avery, McLeod y McCarthy mediante analísis químicos, enzimáticos y serológicos y utilizando técnicas de electroforesis, ultracentrifugación y espectroscopía aislaron a partir de los extractos de neumococos SIII (virulentos) muertos por calor cinco fracciones distintas con el mayor grado de pureza posible en la época. Estas cinco fracciones diferentes fueron una correspondiente a Polisacáridos, otra de Lípidos, una de Proteínas, otra de ARN y otra de ADN.
 muertos por calor cinco fracciones distintas con el mayor grado de pureza posible en la época. Estas cinco fracciones diferentes fueron una correspondiente a Polisacáridos, otra de Lípidos, una de Proteínas, otra de ARN y otra de ADN.")
11
Comprobaron que ninguna de las fracciones era capaz
Aislaron a partir de los extractos de neumococos virulentos muertos por calor cinco fracciones distintas Polisacáridos, Lípidos, Proteínas, ARN y ADN. Con cada una de estas fracciones intentaron transformar las células no virulentas vivas en virulentas. Comprobaron que ninguna de las fracciones era capaz de llevar a cabo la transformación excepto la fracción químicamente pura que contenía ADN Con cada una de estas fracciones procedentes de SIII muertos por calor intentaron transformar las células RII vivas en SIII. Comprobaron que ninguna de las fracciones era capaz de transformar los neumococos RII en SIII excepto la fracción químicamente pura que contenía ADN (ácido desoxirribonucleico). Para asegurarse de que solamente la fracción de ADN era capaz de transformar los neumococos RII en SIII, emplearon enzimas de degradan o digieren específicamente el ADN. Cuando trataban la fracción de ADN con estas enzimas y después intentaban transformar las células RII en SIII, no lo conseguían. Si trataban la fracción de ADN con enzimas que degradan específicamente el ARN y después intentaban la transformación, las células RII se transformaban en SIII. Si la fracción de ADN se trataba previamente con proteasas (enzimas que degradan las proteínas) también conseguían transformar los neumococos RII en SIII. Conclusiones de Avery, McLeod y McCarthy: Teniendo en cuenta que la única fracción químicamente pura de los neumococos SIII muertos por calor que puede transformar los neumococos RII en SIII es el ADN, el Principio Transformante detectado por Griffith debe ser el ADN. Por tanto, la molécula responsable de convertir los neumococos no virulentos en virulentos es el ADN y, por consiguiente, en él debe residir la información genética La primera demostración de que el ADN es el material hereditario se debe, por tanto, a Avery, McLeod y McCarthy en 1944, pero la comunidad científica, en ese momento, no estaba preparada para aceptar sus resultados, ya que pensaban que el ADN era una molécula monótona que consistía en la repetición de un tetranucleótido y que no podía ser la molécula que almacenaba la información genética ya que no disponía de la variabilidad suficiente. Sin embargo, las proteínas eran muy variables y si eran consideradas como candidatos a ser el material hereditario. La transformación en bacterias: para conseguir que una bacteria se transforme es necesario que el ADN exógeno o transformante penetre en su interior, posteriormente, el ADN exógeno o transformante debe integrarse en el ADN bacteriano, luego debe expresarse y, por último, tiene que transmitirse de una bacteria a otra. Por tanto, la molécula responsable de convertir los neumococos no virulentos en virulentos es el ADN y, por consiguiente, en él debe residir la información genética.
. Para asegurarse de que solamente la fracción de ADN era capaz de transformar los neumococos RII en SIII, emplearon enzimas de degradan o digieren específicamente el ADN. Cuando trataban la fracción de ADN con estas enzimas y después intentaban transformar las células RII en SIII, no lo conseguían. Si trataban la fracción de ADN con enzimas que degradan específicamente el ARN y después intentaban la transformación, las células RII se transformaban en SIII. Si la fracción de ADN se trataba previamente con proteasas (enzimas que degradan las proteínas) también conseguían transformar los neumococos RII en SIII. Conclusiones de Avery, McLeod y McCarthy: Teniendo en cuenta que la única fracción químicamente pura de los neumococos SIII muertos por calor que puede transformar los neumococos RII en SIII es el ADN, el Principio Transformante detectado por Griffith debe ser el ADN. Por tanto, la molécula responsable de convertir los neumococos no virulentos en virulentos es el ADN y, por consiguiente, en él debe residir la información genética. La primera demostración de que el ADN es el material hereditario se debe, por tanto, a Avery, McLeod y McCarthy en 1944, pero la comunidad científica, en ese momento, no estaba preparada para aceptar sus resultados, ya que pensaban que el ADN era una molécula monótona que consistía en la repetición de un tetranucleótido y que no podía ser la molécula que almacenaba la información genética ya que no disponía de la variabilidad suficiente. Sin embargo, las proteínas eran muy variables y si eran consideradas como candidatos a ser el material hereditario. La transformación en bacterias: para conseguir que una bacteria se transforme es necesario que el ADN exógeno o transformante penetre en su interior, posteriormente, el ADN exógeno o transformante debe integrarse en el ADN bacteriano, luego debe expresarse y, por último, tiene que transmitirse de una bacteria a otra. Por tanto, la molécula responsable de convertir los neumococos no virulentos en virulentos es el ADN y, por consiguiente, en él debe residir la información genética.")
12
CONCLUSIÓN La primera demostración de que el ADN es el material hereditario se debe, por tanto, a Avery, McLeod y McCarthy en 1944, pero la comunidad científica, en ese momento, no estaba preparada para aceptar sus resultados, ya que pensaban que el ADN era una molécula monótona que consistía en la repetición de un tetranucleótido y que no podía ser la molécula que almacenaba la información genética ya que no disponía de la variabilidad suficiente. Sin embargo, las proteínas eran muy variables y si eran consideradas como candidatos a ser el material hereditario.

13
En 1934 Levene aisló, a partir de ácidos nucleicos, unas moléculas más sencillas que estaban formadas por una pentosa, una base nitrogenada y una molécula de ácido fosfórico. A este conjunto se le dio el nombre de NUCLEÓTIDO. Levene pensó que los ácidos nucleicos estaban formados por cuatro nucleótidos, uno con cada una de las bases. Hasta mediados de los años 50 no se certificó que los ácidos nucleicos en realidad estaban formados por miles e, incluso, millones de nucleótidos y no por sólo cuatro

14
En la década de los 40, diversos investigadores (Feulgen, Caspersson, Mirsky, Sager y otros) desarrollaron técnicas que permitieron estudiar en qué lugares de las células aparecían los ácidos nucleicos. Se observó que el DNA solía aparecer casi exclusivamente en el núcleo y en pequeñas cantidades en algún orgánulo celular como las mitocondrias y los cloroplastos. Se comprobó también que existía DNA en los cromosomas, unido a proteínas, viéndose cómo la cantidad de DNA era siempre constante y propia de cada especie, lo que llevó a sospechar que tal vez existía relación entre el DNA de los cromosomas y los genes o factores hereditarios El RNA aparecía repartido por el citoplasma, sobre todo en los ribosomas, y en cierta cantidades también en el núcleo.

15
EXPERIMENTOS CON FAGOS RADIACTIVOS HERSHEY Y CHASE (1952)
El material que utilizaron Alfred Hershey y Marta Chase (1952) en sus estudios fue el virus T4 que infecta a la bacteria E. coli. Dicho virus mantiene una relación de tipo lítico con esta bacteria. Los virus de la serie T, como el T4 cuando se aíslan y se someten a un choque osmótico pueden reventarse liberándose el ADN que estaba en el interior de su cápside. Hershey y Chase pensaron en alguna forma para marcar los virus T4 solamente en su ADN y solamente en sus proteínas, se dieron cuenta de que el azufre (S) forma parte solamente de las proteínas pero no del ADN, y que el Fósforo (P) forma parte solamente del ADN pero no de las proteínas. Por tanto decidieron obtener dos colecciones de fagos diferentes, una colección de fagos T4 marcados solamente en su ADN con Fósforo radiactivo (P32) y otra colección de fagos T4 marcados solamente en sus proteínas con Azufre radiactivo (S35). el virus T4 que infecta a la bacteria E. coli
 en sus estudios fue el virus T4 que infecta a la bacteria E. coli. Dicho virus mantiene una relación de tipo lítico con esta bacteria. Los virus de la serie T, como el T4 cuando se aíslan y se someten a un choque osmótico pueden reventarse liberándose el ADN que estaba en el interior de su cápside. Hershey y Chase pensaron en alguna forma para marcar los virus T4 solamente en su ADN y solamente en sus proteínas, se dieron cuenta de que el azufre (S) forma parte solamente de las proteínas pero no del ADN, y que el Fósforo (P) forma parte solamente del ADN pero no de las proteínas. Por tanto decidieron obtener dos colecciones de fagos diferentes, una colección de fagos T4 marcados solamente en su ADN con Fósforo radiactivo (P32) y otra colección de fagos T4 marcados solamente en sus proteínas con Azufre radiactivo (S35). el virus T4 que infecta a la bacteria E. coli.")
16
Sabiendo que el azufre (S) forma parte solamente de las proteínas pero no del ADN, y que el Fósforo (P) forma parte solamente del ADN pero no de las proteínas Para conseguir este tipo de fagos T4 llevaron a cabo dos experimentos, en uno de ellos infectaron bacterias de E. coli que crecían en un medio que con tenía P32, los virus descendientes de la infección tenían marcado su ADN con P32; en el otro experimento infectaron bacterias de E. coli que crecían en un medio con S35 , los virus descendientes de la infección tenían marcadas sus proteínas con S35 . Para comprobar que habían marcado correctamente los virus T4, unos solamente en el ADN y otra colección solamente en las proteínas, aislaron parte de los virus descendientes de las infecciones los sometieron a choque osmótico y centrifugaron en condiciones tales que las cápsides de los virus eran más pesadas y se iban al fondo del tubo después de la centrifugación, mientras que el ADN que había salido fuera de las cápsides, como consecuencia del choque osmótico, se quedaba en solución al finalizar la centrifugación. Cuando el experimento se hacia con virus marcados en su ADN con P32 el marcaje aparecía en solución y no en el fondo del tubo, mientras que si el experimento se realizaba con fagos marcados en sus proteínas con S35 el marcaje aparecía en el fondo del tubo donde estaban las cápsides y no en solución.

17
En primer lugar, infectaron bacterias de E
En primer lugar, infectaron bacterias de E. coli que crecían en un medio normal con los fagos T4 marcados en su ADN con P32 e inmediatamente después de la infección agitaron violentamente la mezcla en una batidora, de esta manera trataban de separar las bacterias de los fagos. Posteriormente centrifugaban de forma que las bacterias al ser mas grandes y pesadas se depositaban en el fondo del tubo, mientras que las cápsides de los virus más pequeñas quedaban en solución. Después de centrifugar observaron que el marcaje correspondiente al P32 aparecía en el fondo del tubo donde estaban las bacterias, mientras que en la solución, donde estaban las cápsides de los fagos, no aparecía marcaje debido al P32 (Figura A). Algunos de los virus descendientes de esta infección tenían marcado su ADN con P32. Prácticamente lo único que entra en las bacterias después de la infección por el fago T4 es el ADN del fago, por dicho motivo, cuando se infecta con virus T4 marcados en su ADN el marcaje aparece en el fondo del tubo donde están las bacterias y no en la solución. Además en este caso, los virus descendientes están marcados en su ADN, lo que indica que el único nexo de unión entre dos generaciones de fago T4 es el ADN.
. Algunos de los virus descendientes de esta infección tenían marcado su ADN con P32. Prácticamente lo único que entra en las bacterias después de la infección por el fago T4 es el ADN del fago, por dicho motivo, cuando se infecta con virus T4 marcados en su ADN el marcaje aparece en el fondo del tubo donde están las bacterias y no en la solución. Además en este caso, los virus descendientes están marcados en su ADN, lo que indica que el único nexo de unión entre dos generaciones de fago T4 es el ADN.")
18
El segundo experimento consistió en infectar bacterias que crecían en un medio normal con fagos marcados en sus proteína con S35, agitar inmediatamente después de forma brusca en una batidora la mezcla y centrifugar. En este caso, la mayor parte del marcaje (80%) correspondiente al S35 estaba en la solución, donde se encontraban las cápsides; mientras que en el fondo del tubo, lugar en el que estaban las bacterias, había muy poco marcaje debido al S35 de las proteínas (Figura B). Los virus descendientes de esta infección no tenían marcadas sus cápsides con S35. cuando se infecta con virus marcados en las proteínas, se detecta muy poco marcaje en el fondo del tubo, donde están las bacterias, estando la mayoría del marcaje en la solución, lugar en que se localizan las cápsides proteicas de los virus y no se observan virus descendientes marcados en sus proteínas. Por consiguiente, la molécula portadora de la información en los virus T4 es el ADN y no las proteínas.

19
Conclusiones de Hershey y Chase.
Lo único que entra en las bacterias después de la infección por el fago T4 es el ADN del fago. Por tanto, cuando se infecta con virus T4 marcados en su ADN el marcaje aparece en el fondo del tubo donde están las bacterias y no en la solución. Además en este caso, los virus descendientes están marcados en su ADN, lo que indica que el único nexo de unión entre dos generaciones de fago T4 es el ADN. Sin embargo, cuando se infecta con virus marcados en las proteínas, no se detecta marcaje en el fondo del tubo, donde están las bacterias. La mayoría del marcaje aparece en la suspensión, donde se localizan las cápsidas proteicas de los virus y no se observan virus descendientes marcados en sus proteínas.

20
Conclusión Por consiguiente, la molécula portadora de la información en los virus T4 es el ADN y no las proteínas.. Alfred Hersey recibió el Premio Nobel por sus trabajos con fagos en 1969 junto con Max Delbrück y Salvador Luria.

21
A principios de los años 50 Chargaff tomó
muestras de ADN de distintas células y obtuvo: La proporción relativa de las cuatro bases variaba de unas especies a otras y se mantenía en valores similares en individuos de la misma especie. La cantidad de A=T y cantidad de C=G

22
Maurice Wilkins y Rosalind Franklin utilizaban métodos de difracción de Rayos X para determinar la estructura física del ADN puro. Estos métodos se basan en la propiedad que tienen los átomos de cualquier sustancia química de desviar el haz de Rayos X. Midiendo distancias y ángulos se pueden calcular las posiciones relativas de los átomos en las moléculas Maurice Wilkins y Rosalind Franklin utilizaban métodos de difracción de Rayos X para determinar la estructura física del ADN puro. Estos métodos se basan en la propiedad que tienen los átomos de cualquier sustancia química de desviar el haz de Rayos X. La estructura de la sustancia es la que establece la desviación, que resulta característica para ella. Al colocar la placa radiográfica detrás de una sustancia y bombardearla con rayos X, los haces derivados impresionana la emulsión produciendo puntos y líneas características. Midiendo distancias y ángulos se pueden calcular las posiciones relativas de los átomos en las moléculas

23
Patrón de difracción de una molécula de ADN cristalizada
Modelo cruzado en el medio es característico de una molécula helicoidal Las amplias bandas en la parte superior e inferior dan alguna indicación de periodicidad: una vuelta cada 3,4 nm Diámetro de la hélice 2nm 10 nucleótidos por vuelta

24
BASES NITROGENADAS

25
NUCLEÓTIDO

26
MODELO A ESCALA DE WATSON Y CRICK

27
Modelo de Watson y Crick
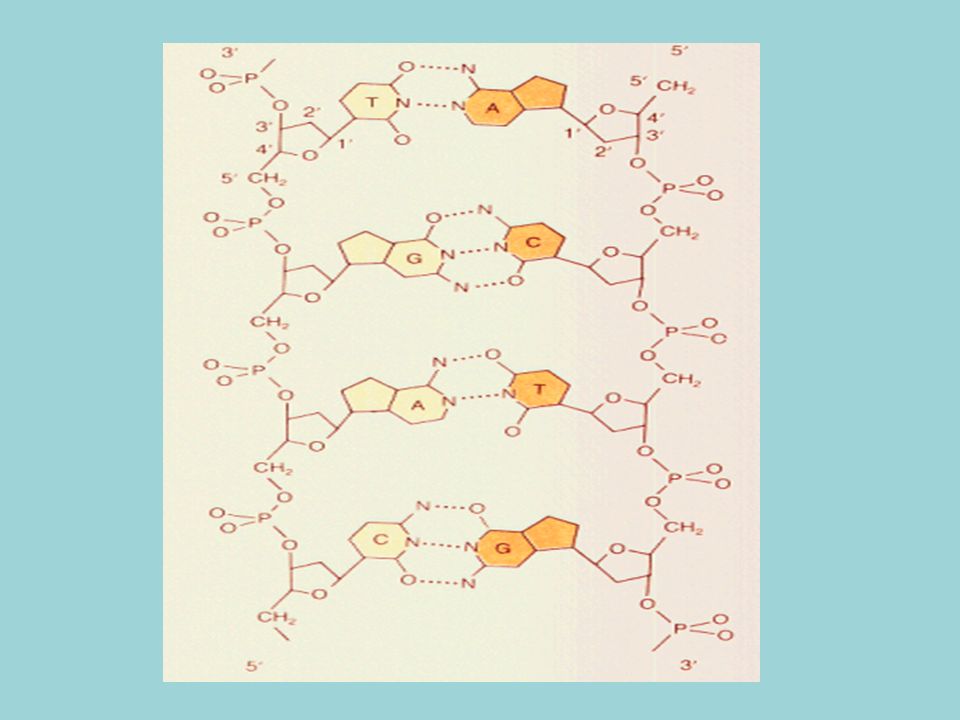
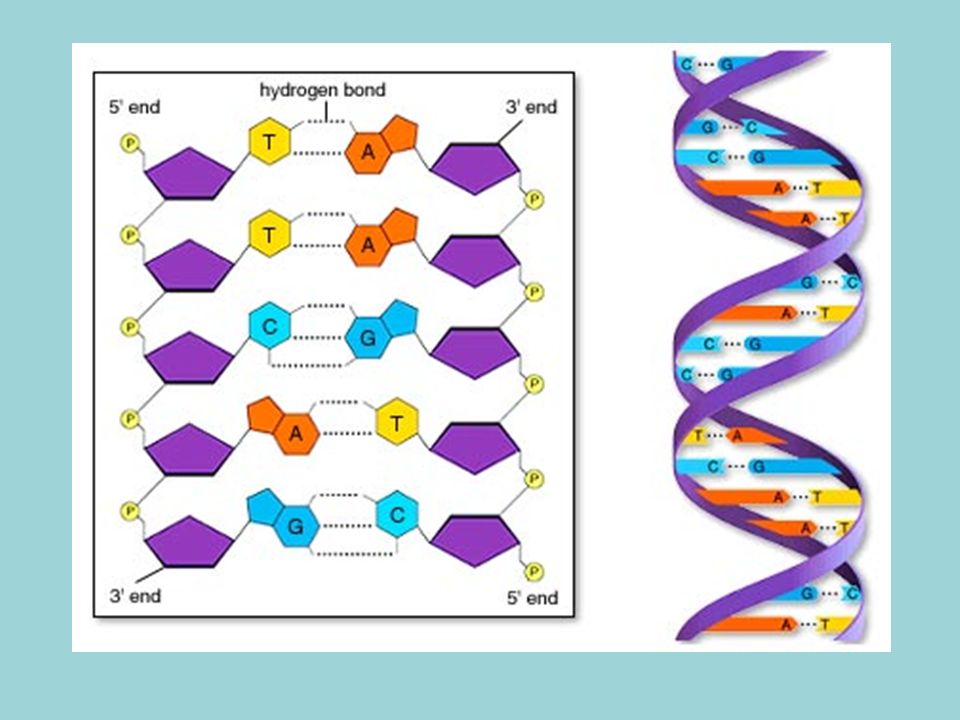
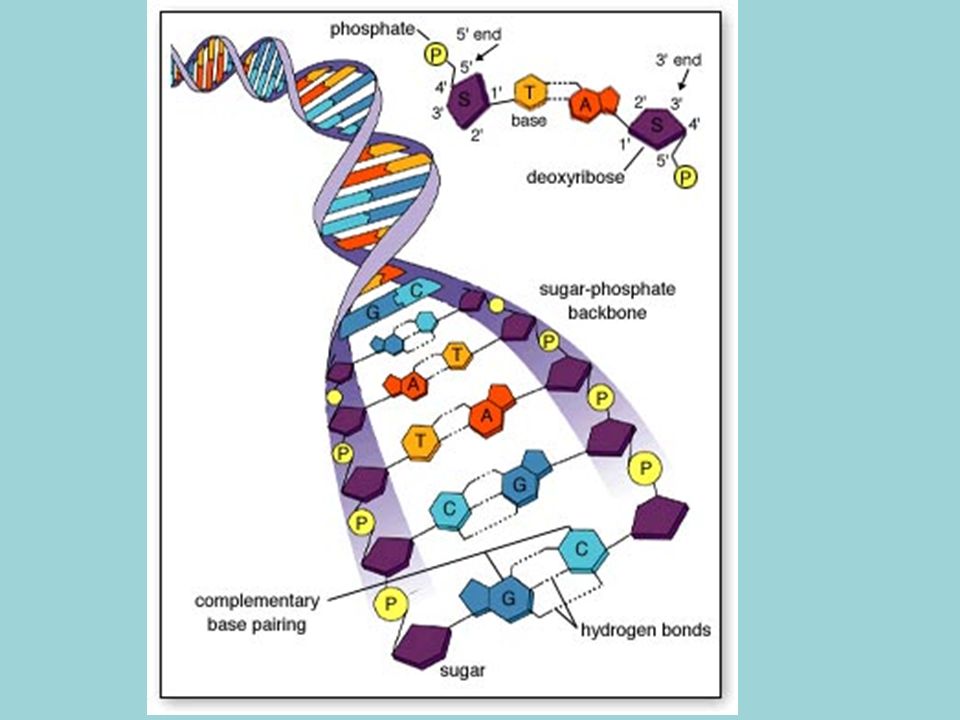
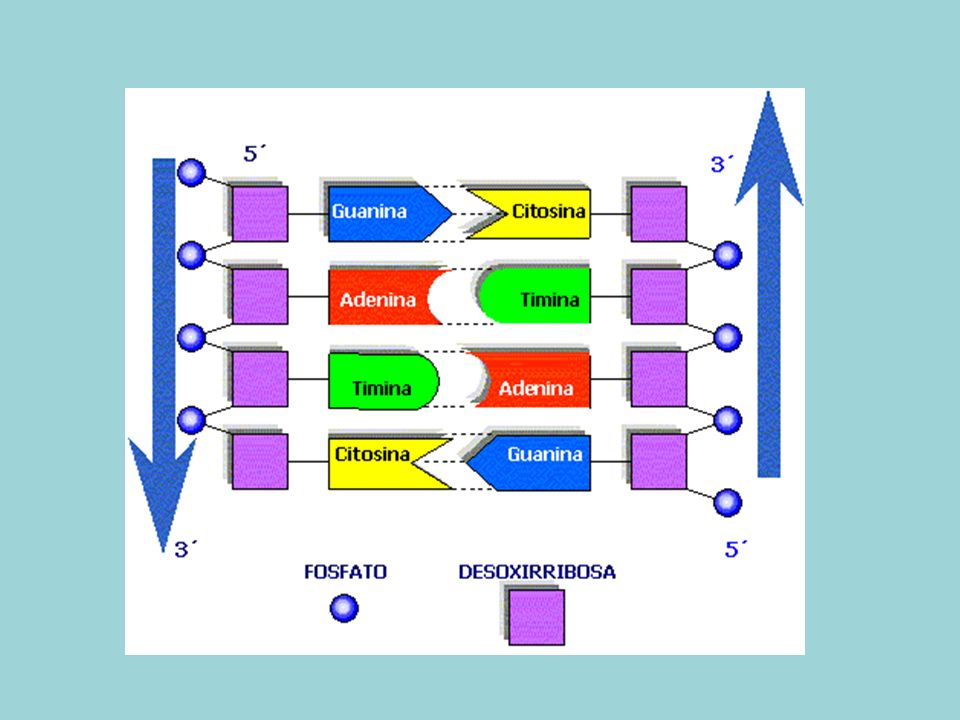
Doble hélice formada por dos cadenas de polinucleótidos enrollados a lo largo de un eje imaginario común. Una cadena va en sentido 5´-3´y la otra en sentido 3´-5´ (antiparalelas) Bases nitrogenadas situadas en el interior de la doble hélice, mientras que pentosa y fosfórico forman el esqueleto externo. Las dos cadenas están unidas mediante puentes de hidrógeno, formados entre A-T y C-G
 Bases nitrogenadas situadas en el interior de la doble hélice, mientras que pentosa y fosfórico forman el esqueleto externo. Las dos cadenas están unidas mediante puentes de hidrógeno, formados entre A-T y C-G.")
28
MOLÉCULA DE ADN

29
DOBLE CADENA DE ADN

31
Esquema de una molécula de ADN

35
MODELOS DE DOBLE HÉLICE

36
Este modelo fue asumido por la genética
molecular porque explica las dos funciones básicas para el material hereditario: Obtener réplicas que se transmiten a la descendencia Almacenar información codificada en un idioma de cuatro letras para la síntesis de proteínas.

37
Molécula de ARN

Presentaciones similares







 y Avery, McLeod y McCarthy (1944)>")











