Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Estructura de la membrana celular y tipos de transporte
Esteban Osorio Cadavid Biología Celular y Bioquímica I Universidad del Valle

21
MEMBRANAS CELULARES Y TRANSPORTE
GENERALIDADES: Las membranas celulares son estructuras laminares con permeabilidad selectiva. Por medio de ellas se separa el medio intracelular altamente organizado, del medio extracelular, garantizando así la integridad celular y la independencia de la célula frente al medio que la rodea. Las membranas celulares son la base estructural de la compartimentación. Los organelos, delimitados por las membranas internas de la célula, mantienen una composición química y unas funciones específicas. A través de las membranas, tanto de la membrana plasmática (membrana externa) como de las endomembranas, se realizan intercambios de sustancias entre el interior y el exterior de la célula y de los organelos. Para la realización de estos intercambios las membranas poseen dispositivos especiales de transporte.
 como de las endomembranas, se realizan intercambios de sustancias entre el interior y el exterior de la célula y de los organelos. Para la realización de estos intercambios las membranas poseen dispositivos especiales de transporte.")
23
Las membranas facilitan la comunicación de la célula con el medio.
Receptores específicos permiten la percepción de estímulos de diversa índole: químicos, hormonales, físicos, etc. Algunas membranas pueden ellas mismas generar señales químicas y eléctricas. También, muchas membranas pueden generar gradientes de potencial electroquímicos que la célula utiliza para la producción de energía en la forma de ATP.

24
FUNCIONES CELULARES ASOCIADAS CON LAS MEMBRANAS
Mantenimiento de la integridad y de la organización celular Compartimentación estructural y funcional Transporte de sustancias Comunicación Generación y recepción de señales químicas y físicas Generación de energía libre

25
ESTRUCTURA Y COMPOSICIÓN DE LAS MEMBRANAS CELULARES
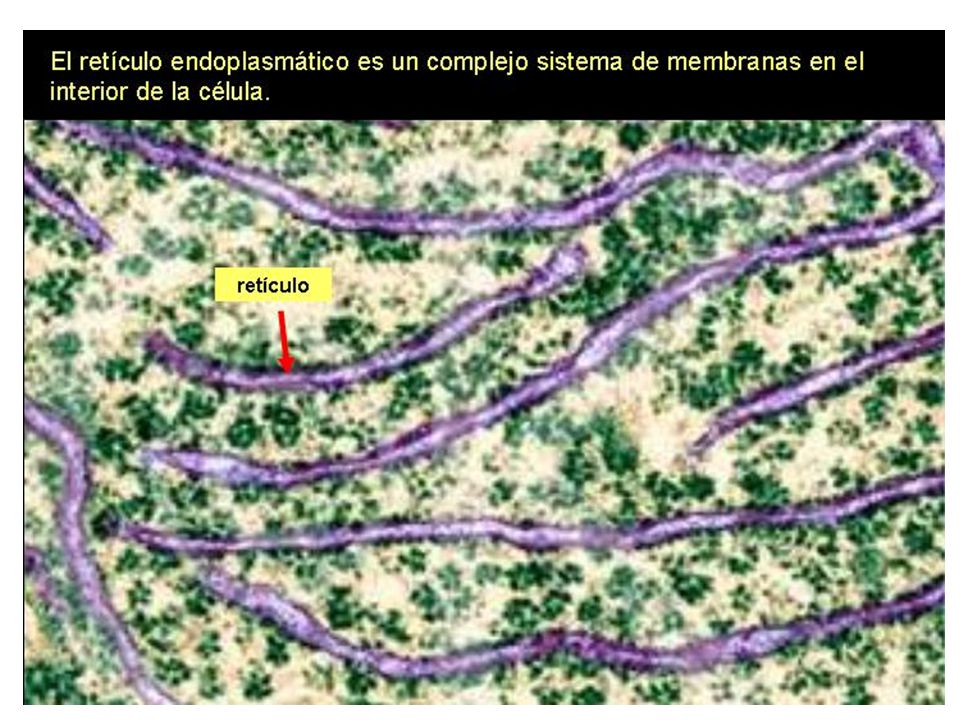
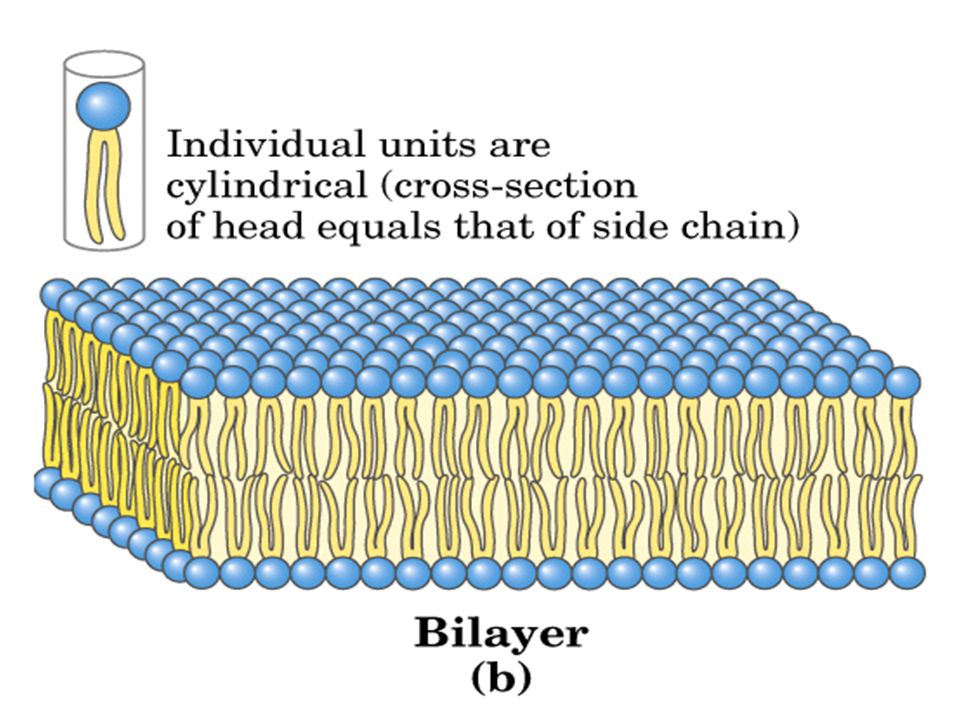
Los cortes de células y tejidos observados al microscopio electrónico, muestran una estructura trilaminar de las membranas, cuyo espesor varía entre 5 y 10 nm. En 1925, E. Gorter y F. Grendel extrajeron los lípidos de las membranas plasmáticas de eritrocitos de mamíferos. Al hacerlos flotar en agua, estos formaron una monocapa, que ocupó dos veces el área de las membranas originales. Concluyeron que, los lípidos se organizaban en forma de bicapas en estas membranas.

27
Composición química de las membranas:
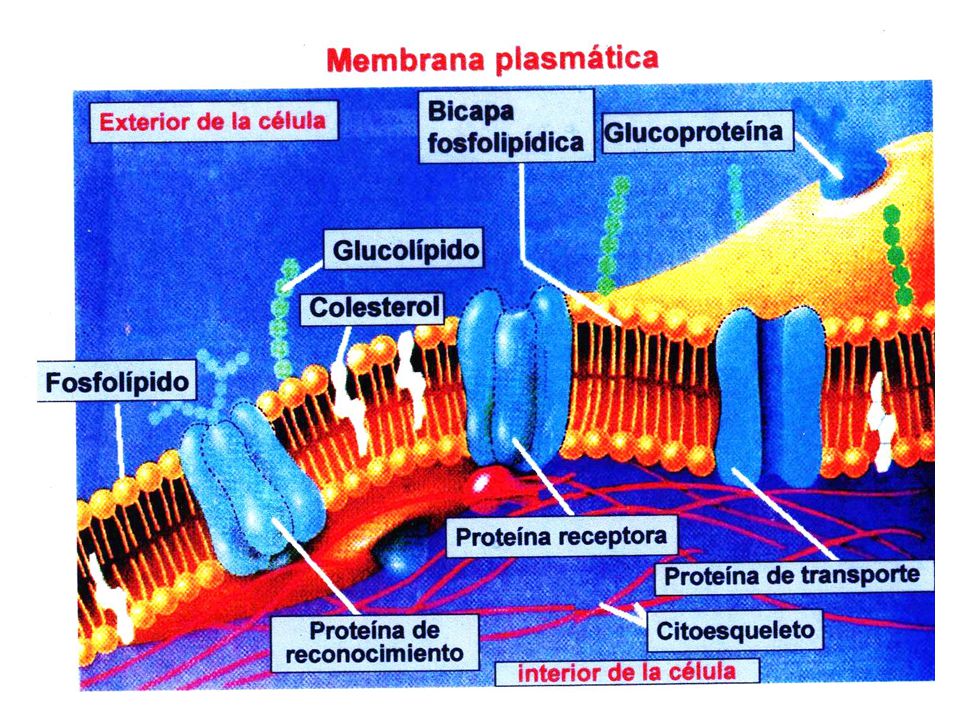
En diversos tipos de membranas se pueden encontrar las siguientes clases de componentes: lípidos, proteínas y oligosacáridos. Los lípidos de las membranas son todos del tipo anfipático: fosfoglicéridos, esfingomielinas, cerebrósidos, gangliósidos, sulfátidos, plasmalógenos, ácidos grasos (generalmente unidos a proteínas) y colesterol. Los oligosacáridos normalmente se encuentran unidos a proteínas (glucoproteínas) y a lípidos (glucolípidos).
 y colesterol. Los oligosacáridos normalmente se encuentran unidos a proteínas (glucoproteínas) y a lípidos (glucolípidos).")
28
Agregados formados por los lípidos anfipáticos en el agua

32
Modelos estructurales de membranas celulares:
Las primeras observaciones de cortes de membranas con el microscópio electrónico de transmisión mostraron, una estructura trilaminar: Una capa clara, en medio de dos láminas densas a los electrones (oscuras). Estas imágenes influyeron durante varias décadas sobre el diseño de diferentes modelos para explicar la organización de las membranas. 1. Modelo de Davson y Danielli (1935). Estos investigadores explicaron la estructura trilaminar de la siguiente manera: la parte central estaba constituida por una bicapa de lípidos enfrentados por sus colas no polares, y las dos láminas externas correspondían a capas de proteínas globulares, dispuestas de manera continua sobre y por debajo de la bicapa lipídica. Las dos capas proteicas explicaban, según ellos, las propiedades hidrofílicas de las membranas.
. Estas imágenes influyeron durante varias décadas sobre el diseño de diferentes modelos para explicar la organización de las membranas. 1. Modelo de Davson y Danielli (1935). Estos investigadores explicaron la estructura trilaminar de la siguiente manera: la parte central estaba constituida por una bicapa de lípidos enfrentados por sus colas no polares, y las dos láminas externas correspondían a capas de proteínas globulares, dispuestas de manera continua sobre y por debajo de la bicapa lipídica. Las dos capas proteicas explicaban, según ellos, las propiedades hidrofílicas de las membranas.")
33
2. Modelo de J.D. Robertson (1950-1960).
Con base en el grosor de aproximadamente 7 a 8 nm para la estructura trilaminar, observada por medio de la microscopía electrónica, Robertson propuso un modelo con las dos capas proteicas de 2 nm de espesor cada una y una bicapa central lipídica de 3-4 nm. Para que estas dimensiones se cumplieran, las proteínas no estarían en forma globular sino extendida. Este modelo se denominó de “unidad de membrana”, debido a que se supuso que todas las membranas celulares, tanto procariotas como eucariotas, tendrían la misma organización. El modelo de Robertson, además introdujo un concepto importante: la asimetría de las membranas. Los glúcidos (oligosacáridos) sólo se encuentran en la superficie externa de las membranas. Estudios posteriores demostrarían que también existe asimetría en la distribución de los lípidos anfipáticos de la bicapa, por ejemplo, las lecitinas, las enfingomielinas y los glucoesfingolípidos predominan en la capa exterior.
 sólo se encuentran en la superficie externa de las membranas. Estudios posteriores demostrarían que también existe asimetría en la distribución de los lípidos anfipáticos de la bicapa, por ejemplo, las lecitinas, las enfingomielinas y los glucoesfingolípidos predominan en la capa exterior.")
34
Composición lipídica de la membrana plasmática y membranas de organelas de un hepatocito de rata

35
Distribución asimétrica de los fosfolípidos entre la monocapa interna y externa de la membrana plasmática de eritrocito

36
3. Modelo del mosaico fluido (1972).
Modelo propuesto por S.J. Singer y G.L. Nicholson con base en los datos proporcionados por nuevas técnicas aplicadas al estudio de las membranas. Empleando criofractura y criograbado, las imágenes microscópicas mostraron estructuras granuladas complementarias, en la fractura de la bicapa. Las protuberancias en un lado de la fractura tenían concavidades complementarias en la cara opuesta. Otras técnicas novedosas, como La espectroscopia de resonancia de spin electrónico (ESR), que mide la movilidad de los lípidos en la membrana, y la calorimetría de barrido diferencial (DSC), que mide las transiciones de gel a estados más fluidos mediante la absorción de calor en función de la temperatura, mostraron la existencia de una bicapa lipídica fluida, que permite la movilidad de sus componentes.
, que mide la movilidad de los lípidos en la membrana, y la calorimetría de barrido diferencial (DSC), que mide las transiciones de gel a estados más fluidos mediante la absorción de calor en función de la temperatura, mostraron la existencia de una bicapa lipídica fluida, que permite la movilidad de sus componentes.")
37
Corte de la bicapa lipídica por la técnica freeze-fracture

40
Movimiento de los lípidos de membrana

41
Demostración de la difusión lateral de las proteínas de membrana

43
Membrane Structure

45
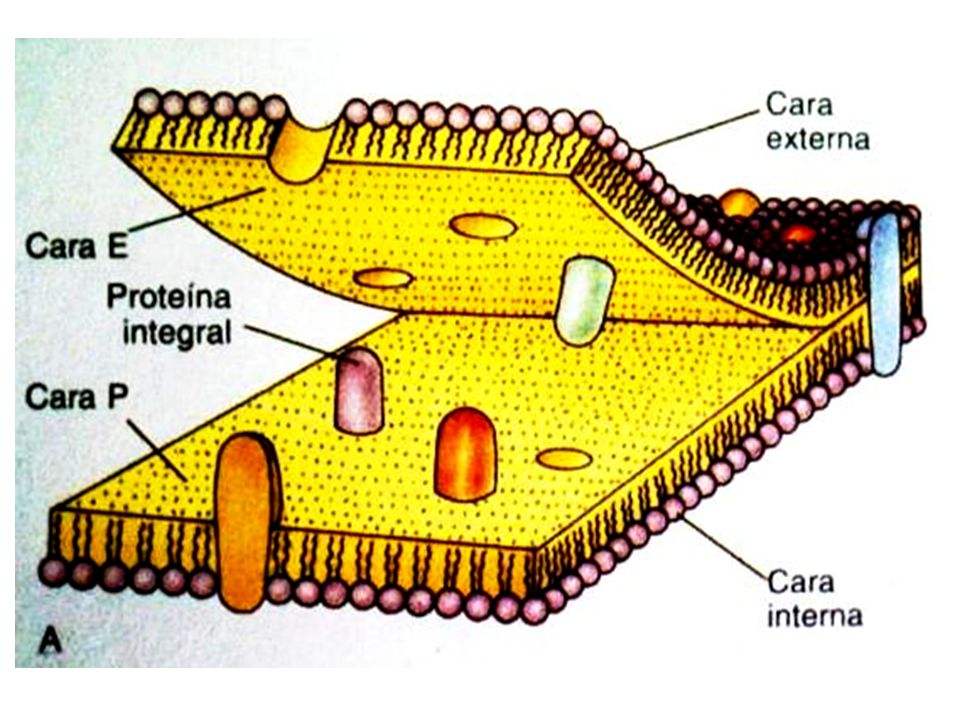
Esquemáticamente, el modelo del mosaico fluido considera que las membranas celulares están constituidas por una matriz, a manera de esqueleto aglutinante, conformada por la bicapa de los lípidos anfipáticos. Las proteínas están asociadas a la bicapa lipídica, ya sea unidas por la parte externa o por el lado citoplasmático (proteínas extrínsecas o periféricas), o incluidas atravesando la bicapa (proteínas intrínsecas, integrales o transmembranales). En el logro de esta asociación se presentan diversos tipos de interacciones moleculares: lípido-proteína, proteína-proteína, interacciones hidrofóbicas, interacciones polares y aún covalentes: como por ejemplo, proteína-ácido graso, o proteína-carbohidrato. La fluidez de la matriz lipídica implica la ocurrencia de movilidad en los lípidos y en las proteínas, condición fundamental para que las membranas puedan desarrollar sus funciones.
, o incluidas atravesando la bicapa (proteínas intrínsecas, integrales o transmembranales). En el logro de esta asociación se presentan diversos tipos de interacciones moleculares: lípido-proteína, proteína-proteína, interacciones hidrofóbicas, interacciones polares y aún covalentes: como por ejemplo, proteína-ácido graso, o proteína-carbohidrato. La fluidez de la matriz lipídica implica la ocurrencia de movilidad en los lípidos y en las proteínas, condición fundamental para que las membranas puedan desarrollar sus funciones.")
47
TRANSPORTE A TRAVÉS DE MEMBRANAS
Una de las funciones más importantes de las membranas es su capacidad para transportar sustancias desde y hacia el interior de las células. Las funciones de transporte permiten a las células: Regular el volumen celular, mantener el pH y conservar la composición iónica intracelular dentro de los límites adecuados para una eficiente actividad enzimática. -Extraer y concentrar combustibles metabólicos y precursores del ambiente y eliminar desechos. -Generar gradientes iónicos esenciales para la excitabilidad de células especializadas, como por ejemplo, las nerviosas, musculares y secretoras. -Generar los gradientes electroquímicos que posibilitan la obtención de energía libre, básicamente en la forma de ATP.

55
DIFUSIÓN Es el movimiento pasivo de moléculas de una concentración superior a una inferior hasta alcanzar el equilibrio. Los Gases se mueven a través de la membrana plasmática por difusión.

58
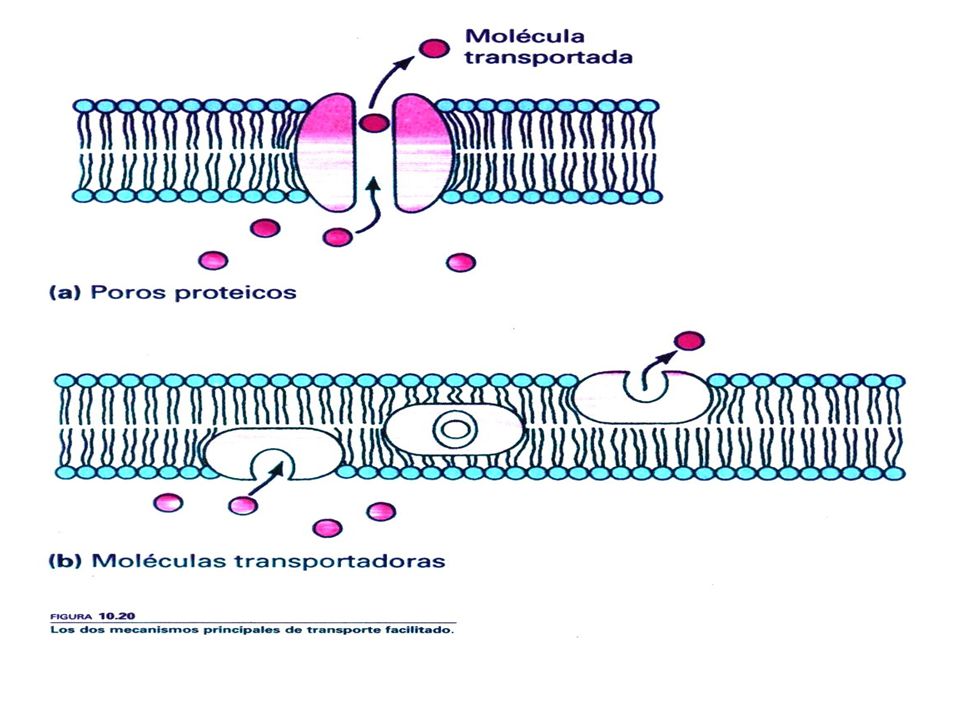
Proceso de Transporte Facilitado
Proteína se acopla con la molécula La forma de la proteína cambia La molécula se mueve a través de la membrana.

62
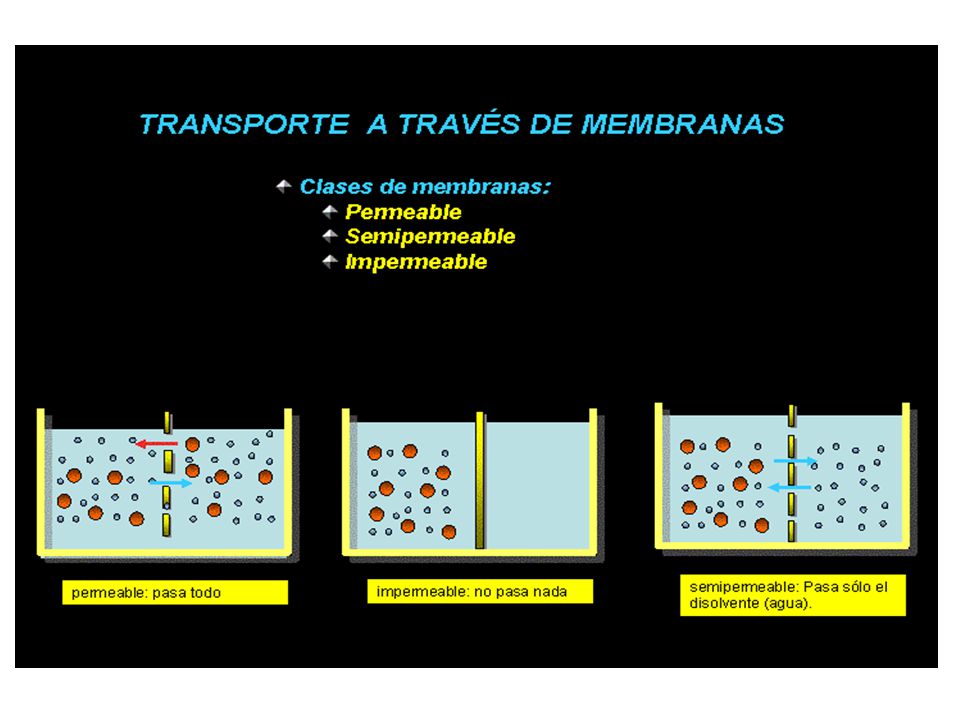
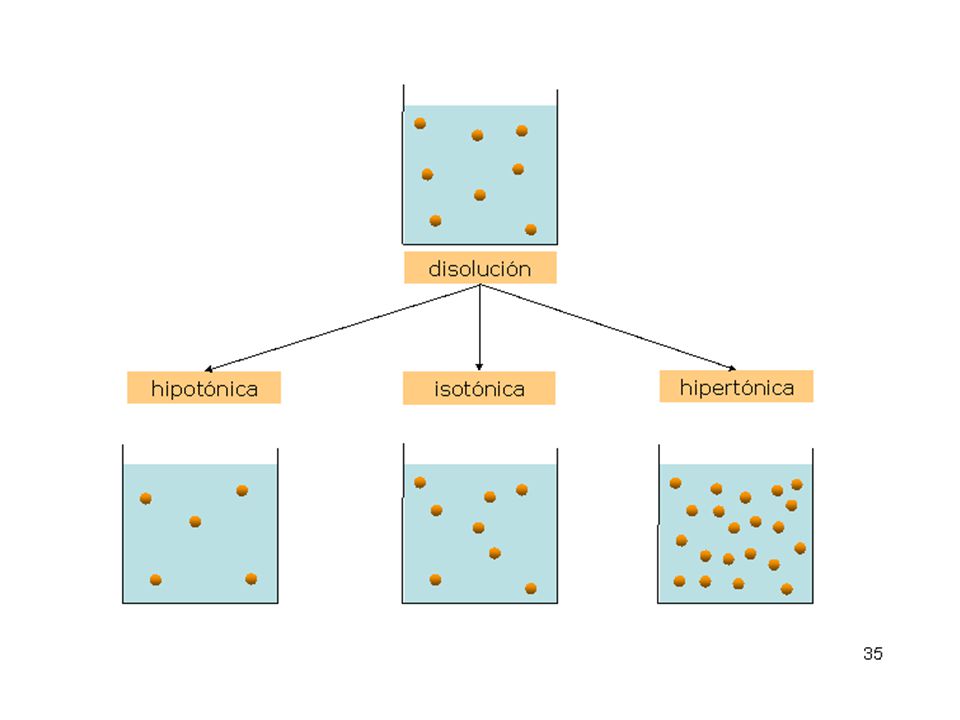
membrana semipermeable
Comportamiento de dos disoluciones separadas por una membrana semipermeable. Presión osmótica membrana semipermeable hipertónica hipotónica Para explicar la ósmosis platearemos el siguiente modelo teórico...

63
Dejémonos de moscas y pasemos a las moléculas.

65
membrana semipermeable
Seguro que ahora habrás comprendido por qué aumenta la cantidad de líquido en la disolución hipertónica. Si aún no es así tendremos que explicarlo más detenidamente….. membrana semipermeable hipertónica hipotónica

69
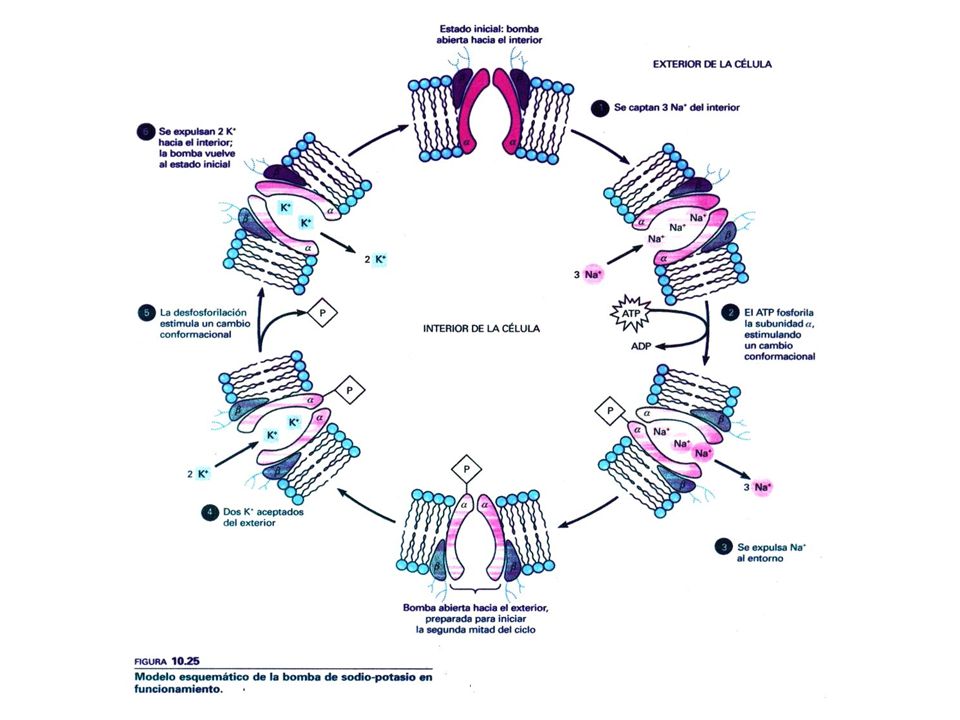
TRANSPORTE ACTIVO Movimiento Molecular
Requiere energía (en contra de gradiente) Ejemplo: Bomba sodio-potasio
 Ejemplo: Bomba sodio-potasio.")
73
El transporte pasivo presenta diferentes modalidades: DIFUSIÓN SIMPLE y DIFUSIÓN FACILITADA
La difusión simple ocurre cuando las moléculas atraviesan la membrana a favor del gradiente de concentración sin ayuda alguna. El paso puede ocurrir a través de la bicapa lipídica, si son moléculas no polares o gases; o a través de poros o canales estructurales, generalmente de naturaleza proteica, por donde pueden pasar las moléculas polares y los iones. La difusión facilitada requiere de moléculas transportadoras que ayuden a atravesar la membrana. Estos transportadores son generalmente proteínas transmembranales, que por su especificidad y características cinéticas similares a las enzimas, han recibido el nombre genérico de PERMEASAS. Se han identificado permeasas para numerosos nutrientes celulares, entre ellas las de animoácidos y las de diversos monosacáridos.

75
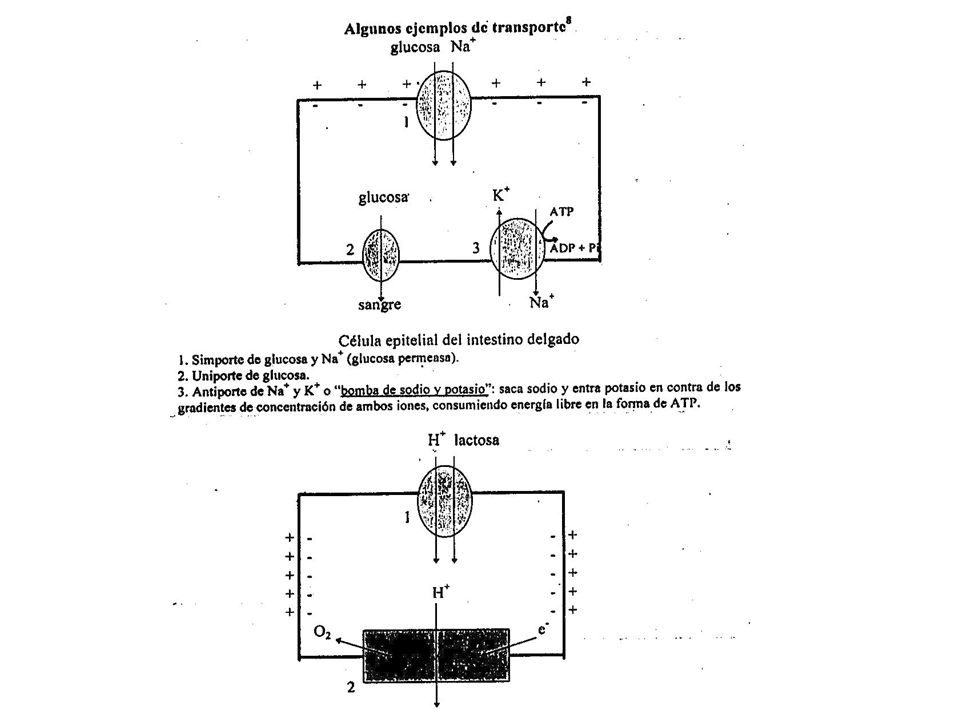
TIPOS DE TRANSPORTE El transporte puede ser UNIPORTE si se transporta una sola molécula o ión, o COTRANSPORTE si se movilizan simultáneamente dos o más especies moleculares, en la misma dirección (SIMPORTE) o en direcciones opuestas (ANTIPORTE). Dependiendo de los cambios de energía libre durante el transporte, éste se clasifica en PASIVO o ACTIVO. El transporte es pasivo cuando el cambio de energía libre que ocurre durante el proceso es negativo, indicando que el movimiento es espontáneo en la dirección planteada. Se considera activo, cuando el cambio de energía libre del proceso es positivo, no espontáneo, indicando que se requiere suministro de energía libre para que el transporte ocurra en la dirección planteada.
 o en direcciones opuestas (ANTIPORTE). Dependiendo de los cambios de energía libre durante el transporte, éste se clasifica en PASIVO o ACTIVO. El transporte es pasivo cuando el cambio de energía libre que ocurre durante el proceso es negativo, indicando que el movimiento es espontáneo en la dirección planteada. Se considera activo, cuando el cambio de energía libre del proceso es positivo, no espontáneo, indicando que se requiere suministro de energía libre para que el transporte ocurra en la dirección planteada.")
77
Tres clases generales de sistemas de transporte
77

90
COMUNICACIÓN INTERCELULAR
Una de las principales funciones de las membranas celulares es la mediación en la comunicación de señales entre células y entre éstas y el medio. La comunicación entre células puede efectuarse por medio de tres vías principales: Por medio de señales químicas entre células situadas a alguna distancia. 2. Por medio de señales químicas entre células situadas muy próximas. 3. A través de las llamadas uniones de comunicación (conexones), las cuales ponen en contacto los citoplasmas de células adyacentes.
, las cuales ponen en contacto los citoplasmas de células adyacentes.")
93
La vía de comunicación intercelular mejor conocida y ampliamente estudiada, es la que se hace por medio de SEÑALES QUÍMICAS entre células situadas a alguna distancia (variable). Algunos ejemplos son los siguientes: 1. Mediadores químicos locales: son tomados o destruidos muy rápidamente. Actúan sólo sobre células adyacentes. Ejemplos típicos son: los interferones, las linfocinas, las prostaglandinas y los tromboxanos. 2. Hormonas: son secretadas por células endocrinas especializadas, distribuidas luego por los fluidos del organismo (sangre, linfa, savia) a todo lugar y captadas por las células diana por medio de receptores específicos. Las células diana pueden estar a gran distancia de las células secretoras. 3. Neurotransmisores: son secretadas por células del sistema nervioso y actúan solamente sobre las células diana que están contiguas.
 a todo lugar y captadas por las células diana por medio de receptores específicos. Las células diana pueden estar a gran distancia de las células secretoras. 3. Neurotransmisores: son secretadas por células del sistema nervioso y actúan solamente sobre las células diana que están contiguas.")
94
LOS RECEPTORES La capacidad de una célula para responder a un mensajero particular de señalización extracelular, depende de que posea proteínas especializadas llamadas receptores, que unan o liguen la molécula señal. Los receptores son proteínas de las membranas o del citoplasma. Las moléculas señal que se unen a los receptores se denominan LIGANDOS. Muchas de estas moléculas actúan a concentraciones muy bajas (por ejemplo, 10-8 Molar). No obstante los receptores son capaces de reconocerlas y unirlas, generalmente con una muy alta afinidad. Los ligandos de naturaleza polar como los neurotransmisores, las hormonas proteicas y los factores de crecimiento, se unen a proteínas receptoras específicas situadas en la superficie de las células blanco. La unión receptor-ligando convierte este evento extracelular en una señal intracelular, que modifica el comportamiento funcional de la célula diana. La transducción de la señal, desde el receptor hasta el interior de la célula, usualmente es mediada por otras proteínas de la membrana plasmática.
. No obstante los receptores son capaces de reconocerlas y unirlas, generalmente con una muy alta afinidad. Los ligandos de naturaleza polar como los neurotransmisores, las hormonas proteicas y los factores de crecimiento, se unen a proteínas receptoras específicas situadas en la superficie de las células blanco. La unión receptor-ligando convierte este evento extracelular en una señal intracelular, que modifica el comportamiento funcional de la célula diana. La transducción de la señal, desde el receptor hasta el interior de la célula, usualmente es mediada por otras proteínas de la membrana plasmática.")
95
El número de receptores puede variar entre 500 y 100. 000 por célula
El número de receptores puede variar entre 500 y por célula. Se pueden encontrar difusos o acumulados en determinadas regiones de la membrana plasmática. La cinética de unión entre receptor-ligando es muy similar a la de Michaelis-Menten para las enzimas y por lo tanto puede llegar a la saturación. Generalmente, la unión del ligando al receptor induce cambios conformacionales que conducen a la generación de una señal intracelular, que modifica el comportamiento metabólico y funcional de la célula blanco. Las moléculas señal intracelulares, generadas por la unión receptor-ligando, se denominan SEGUNDOS MENSAJEROS. Por ejemplo, el AMP cíclico (AMPc) es el segundo mensajero en numerosas interacciones entre receptores y ligandos.
 es el segundo mensajero en numerosas interacciones entre receptores y ligandos.")
96
Algunas funciones celulares que dependen de interacciones receptor-ligando.
-Respuesta a estimulación hormonal Respuesta a neurotransmisores Percepción de olores Reconocimiento entre células iguales Reconocimiento parásito-hospedero Estimulación de la división celular por factores de crecimiento Captación de nutrientes y otras moléculas Endocitosis de macromoléculas, partículas y células

97
ENDOCITOSIS MEDIADA POR RECEPTORES.
Citosis: transporte de sustancias, partículas y células a través de vesículas constituidas a partir de las membranas celulares. Puede ser de dos clases: Endocitosis: transporte hacia el interior de la célula. Exocitosis: transporte hacia el exterior de la célula.

98
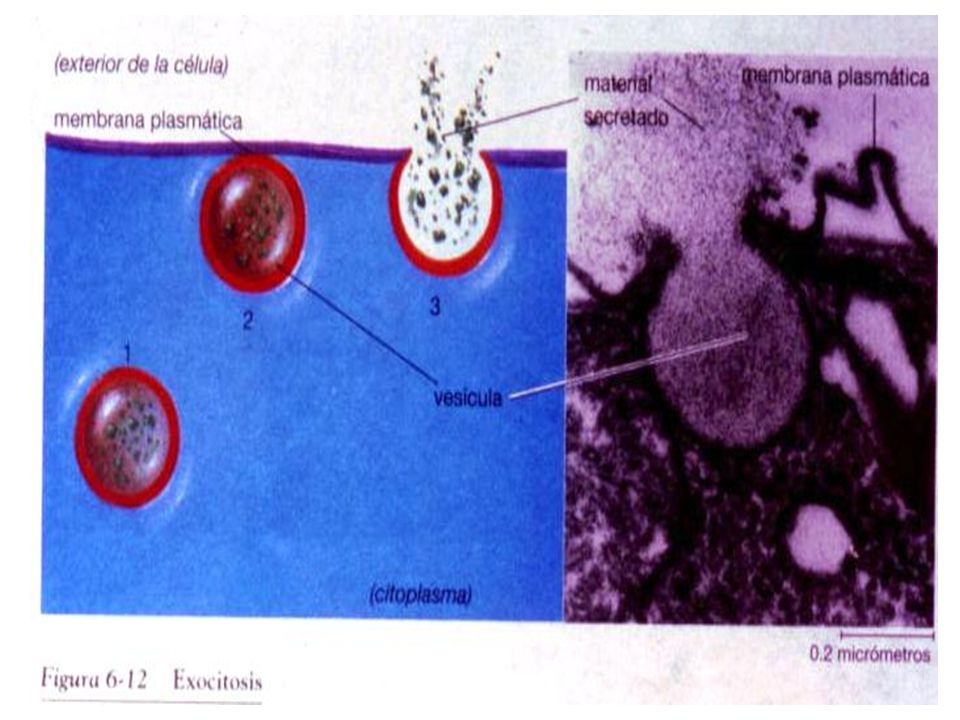
Durante la citosis la célula consume energía libre.
Cuando se aplican venenos metabólicos que impiden la producción de ATP, la citosis se detiene. Si se estimula la síntesis de ATP en células en cultivo, la citosis se hace más intensa. La exocitosis es fundamental en los procesos de secreción a partir del aparato de Golgi y para la excreción de desechos. Las vesículas que transportan las sustancias se fusionan con la membrana plasmática y descargan su contenido al exterior de la célula. La endocitosis permite la incorporación de sustancias, partículas y aún de células enteras, mediante la invaginación de la membrana plasmática y con la consiguiente formación de una vesícula endocítica.

99
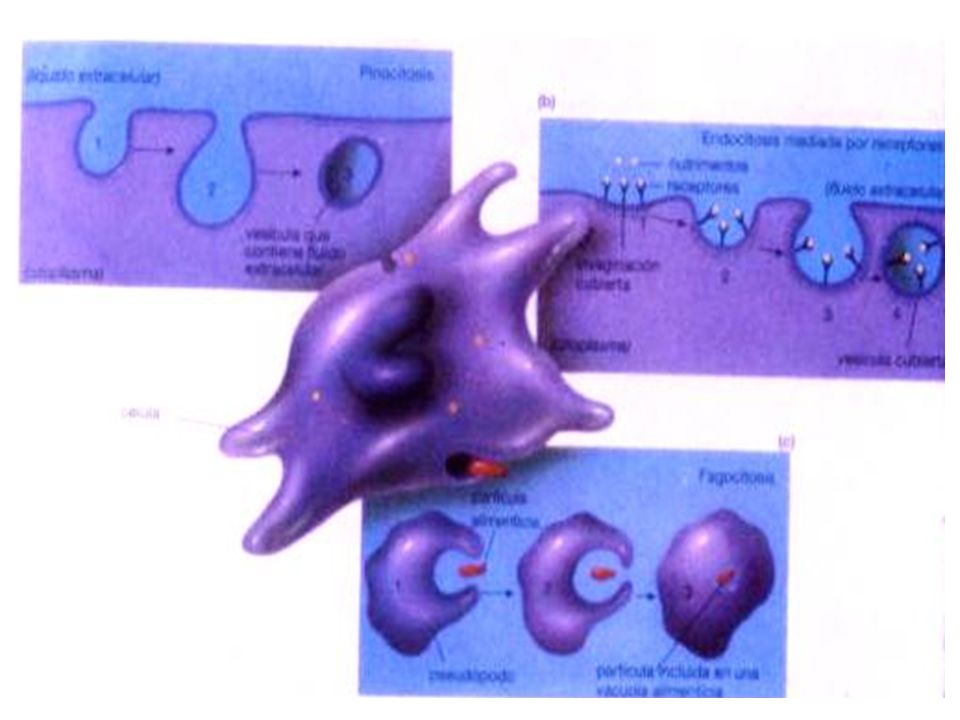
Pinocitosis: (del griego pinein, beber) si son sustancias líquidas.
Usualmente se reconocen dos clases de endocitosis de acuerdo al tipo de material transportado: Pinocitosis: (del griego pinein, beber) si son sustancias líquidas. 2. Fagocitosis: (del griego phagein, comer) si son partículas o células
 si son sustancias líquidas. 2. Fagocitosis: (del griego phagein, comer) si son partículas o células.")
100
Por medio de la endocitosis la célula puede internalizar macromoléculas de importancia nutricional y fisiológica, como por ejemplo: proteínas, polisacáridos, lípidos, lipoproteínas, transferrina, transcobalamina, hormonas polipeptídicas, inmunoglobulinas y enzimas lisosomales. Entre los mecanismos de transporte por endocitosis, el más estudiado por su importancia funcional, es la endocitosis mediada por receptores. Este es un proceso específico de transporte, en el que participan receptores de la membrana plasmática. Las macromoléculas y partículas se concentran en la superficie de la membrana unidas a sus receptores específicos. En unos pocos minutos los complejos formados por los receptores y los ligandos son internalizados.

101
Se han reconocido más de 25 receptores específicos que participan en procesos de endocitosis mediada por receptores. Como ejemplos, podemos citar: 1. Receptores para el transporte de proteínas y partículas plasmáticas sanguíneas, que liberan al interior de la célula nutrientes. Como los receptores de las LDL, que permiten la obtención del colesterol y otros lípidos. Los receptores de transferrina, que permiten la obtención del hierro; y los receptores de la transcobalamina II, que posibilita la obtención de la vitamina B12. 2. Receptores de proteínas plasmáticas no transportadoras, tales como los de asialoglucoproteínas, de -2-macroglobulinas y de complejos inmunes (complejos antígeno-anticuerpo). 3. Receptores de factores de crecimiento proteicos y de hormonas polipeptídicas clásicas, tales como los receptores para los factores de crecimiento de las plaquetas (PDGF), factores de crecimiento epidérmico (EGF) y factores de crecimiento nervioso (NGF).
. 3. Receptores de factores de crecimiento proteicos y de hormonas polipeptídicas clásicas, tales como los receptores para los factores de crecimiento de las plaquetas (PDGF), factores de crecimiento epidérmico (EGF) y factores de crecimiento nervioso (NGF).")
102
RUTAS INTRACELULARES DE LOS COMPLEJOS RECEPTOR – LIGANDO (R-L) EN LA ENDOCITOSIS MEDIADA POR RECEPTORES. Los complejos receptor-ligando caen en invaginaciones denominadas huecos recubiertos. El recubrimiento del lado citoplasmático del hueco está constituido por redes proteicas, en las que el principal componente es la CLATRINA. (Proteína de peso molecular ). Debido a la fluidez de la matriz de las membranas, el hueco se desprende formando una vesícula recubierta o vesícula endocítica. La vesícula endocítica pierde su recubrimiento de clatrina y de algunas de las demás proteínas y se convierte en un endosoma. Entre las capacidades de algunos endosomas está la de “bombear” iones H+ desde el citoplasma hacia el interior del endosoma, rebajando el valor del pH interno al aumentar la concentración de hidrógeno [H+] en el interior del endosoma y convirtiendo a la vesícula en un compartimiento desacoplante del complejo receptor ligando. Representado por la sigla CURL (del inglés compartment of uncoupling of receptor-ligand).
. Debido a la fluidez de la matriz de las membranas, el hueco se desprende formando una vesícula recubierta o vesícula endocítica. La vesícula endocítica pierde su recubrimiento de clatrina y de algunas de las demás proteínas y se convierte en un endosoma. Entre las capacidades de algunos endosomas está la de bombear iones H+ desde el citoplasma hacia el interior del endosoma, rebajando el valor del pH interno al aumentar la concentración de hidrógeno [H+] en el interior del endosoma y convirtiendo a la vesícula en un compartimiento desacoplante del complejo receptor ligando. Representado por la sigla CURL (del inglés compartment of uncoupling of receptor-ligand).")
103
Luego se puede efectuar la fusión del endoplasma con lisosomas primarios, posibilitando la digestión de su contenido por las enzimas hidrolíticas lisosomales contenidas en estos endosomas primarios. En resumen, la ruta intracelular de los complejos receptor-ligando se podría esquematizar de la siguiente manera: COMPLEJO RECEPTOR- LIGANDO HUECO RECUBIERTO VESÍCULA RECUBIERTA ENDOSOMA CURL (¿) (¿) representa el siguiente paso, el cual varía dependiendo de los complejos receptor-ligando y del tipo de célula implicada.
 (¿) representa el siguiente paso, el cual varía dependiendo de los complejos receptor-ligando y del tipo de célula implicada.")
104
Como ejemplos ilustrativos se pueden citar los siguientes casos:
1. Los receptores reciclan y los ligandos se degradan en los lisosomas. Ejemplos: los receptores de LDL, asialoglucoproteínas, transcobalamina II, hormonas peptídicas y algunos virus. 2. Los receptores y los ligandos reciclan (no intervienen los lisosomas). Ejemplos: los receptores de transferrina, moléculas del MHC (complejo mayor de histocompatibilidad) clase I, en las células T y moléculas del MHC clase II, en los macrófagos. 3. Los receptores y los ligandos se degradan en los lisosomas. Ejemplos: los receptores del factor de crecimiento epidérmico (EGF) y algunos complejos inmunes (Fc o C3b). 4. Los receptores y los ligandos son transportados. Ejemplos: los receptores de las inmunoglobulinas IgG maternas e IgA secretoras.
. Ejemplos: los receptores de transferrina, moléculas del MHC (complejo mayor de histocompatibilidad) clase I, en las células T y moléculas del MHC clase II, en los macrófagos. 3. Los receptores y los ligandos se degradan en los lisosomas. Ejemplos: los receptores del factor de crecimiento epidérmico (EGF) y algunos complejos inmunes (Fc o C3b). 4. Los receptores y los ligandos son transportados. Ejemplos: los receptores de las inmunoglobulinas IgG maternas e IgA secretoras.")
105
IONOFOROS La acción de ciertos antibióticos denominados IONOFOROS, anula el potencial de las membranas y de esta forma anulan su capacidad de reacción. Estos antibióticos, sintetizados por microorganismos, aumentan la permeabilidad de las membranas a determinados iones, permitiendo que las concentraciones externa e interna se equilibren. Como ejemplos de estos antibióticos encontramos la Gramicidina A, la Valinomicina y la Nigericina. Sus estructuras moleculares se incrustan en la bicapa lipídica y forman un conducto a través del cual pasan los iones K+, Na+, y H+, en el caso de la Gramicidina A. K+ en el de la Valinomicina y, K+ y H+ en el caso de la Nigericina.

106
La Gramicidina es un péptido lineal de 15 residuos, con cadenas R hidrofóbicas que facilitan su solubilidad en la bicapa lipídica. El conducto transmembranal para los iones se formaría por la asociación de dos moléculas del antibiótico. La Valinomicina es un péptido cíclico formado por la repetición por tres veces de la secuencia L-lactato, L-valina, D-hidroxiisovalerato y D-valina. La parte hidrofóbica está constituida por las cadenas R de las valinas. Hacia el interior quedan varios átomos de oxígeno que dan el ambiente polar adecuado para el movimiento de los iones

109
La valinomicina, Un ionóforo peptídico que une al potasio
109

114
MEMBRANAS EXCITABLES Algunas de las membranas plasmáticas, tienen una gran capacidad de reacción ante estímulos mediante cambios transitorios de su potencial eléctrico. Tal es el caso de las membranas plasmáticas de las células nerviosas y las células musculares.

115
POTENCIAL ELECTRICO DE LAS MEMBRANAS.
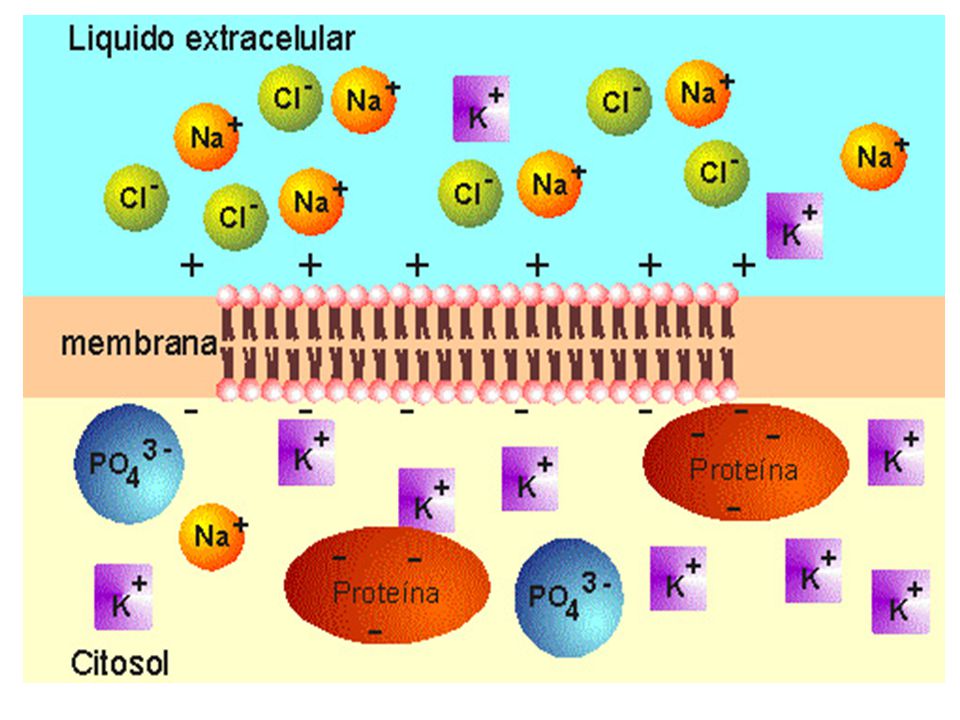
Las membranas son estructuras con polaridad eléctrica a causa de la desigual distribución de las cargas eléctricas entre los dos lados de la membrana. En general, las membranas plasmáticas son NEGATIVAS del lado citoplasmático y POSITIVAS del lado externo. El interior celular negativo, depende de aniones tales como: las proteínas, los ácidos nucleicos, los nucleótidos, y los iones HCO3-, H2PO4-, HPO4-2, SO3-, COO- y Cl-. El exterior celular positivo depende principalmente del Na+ y en menor grado al Mg+ y al Ca+2. El potencial eléctrico de cada membrana resultará de la suma de los potenciales de los iones presentes. Para el mantenimiento de los potenciales eléctricos de las membranas celulares (potenciales de reposo), es indispensable que los gradientes de concentración de los iones, a través de las membranas se mantengan.
, es indispensable que los gradientes de concentración de los iones, a través de las membranas se mantengan.")
117
V = 0.058 z . log Cext / Cint (voltios)
El potencial eléctrico a través de las membranas es una función de la carga y de la concentración de los iones en el interior y el exterior celular: V = z . log Cext / Cint (voltios) Esta ecuación se denomina Ecuación de Nernst, en la que: V = diferencia de potencial (potencial eléctrico) en voltios z = carga del ión Cext = concentración externa del ión Cint = concentración interna del ión La Ecuación de Nernst, deriva de la ecuación del transporte de una molécula cargada eléctricamente: G = RT log C2 (int) / C1 (ext) + zFV
 Esta ecuación se denomina Ecuación de Nernst, en la que: V = diferencia de potencial (potencial eléctrico) en voltios. z = carga del ión. Cext = concentración externa del ión. Cint = concentración interna del ión. La Ecuación de Nernst, deriva de la ecuación del transporte de una molécula cargada eléctricamente: G = RT log C2 (int) / C1 (ext) + zFV.")
119
POTENCIAL DE ACCIÓN El potencial de reposo de diversas membranas se encuentra entre –50 y –100 milivoltios, con el interior negativo respecto al exterior celular. Una membrana excitable es capaz de generar y conducir un impulso eléctrico, en la forma de un cambio repentino y temporal de su potencial eléctrico. La súbita reducción de la diferencia de cargas entre el interior y el exterior, se denomina despolarización. Esto se manifiesta por el repentino cambio del potencial de la membrana, pudiendo llegar a ser positivo dentro y negativo afuera (inversión de la polaridad). La despolarización sólo dura un milisegundo y recibe el nombre de POTENCIAL DE ACCIÓN, o de POTENCIAL EN ESPIGA y es la manifestación, de la excitabilidad de las membranas especializadas en este tipo de reacciones (las células musculares y las células nerviosas).
. La despolarización sólo dura un milisegundo y recibe el nombre de POTENCIAL DE ACCIÓN, o de POTENCIAL EN ESPIGA y es la manifestación, de la excitabilidad de las membranas especializadas en este tipo de reacciones (las células musculares y las células nerviosas).")
120
Pasado el milisegundo, el potencial regresa rápidamente a sus valores originales, por la búsqueda del equilibrio celular, lo cual se denomina Repolarización. Frecuentemente, pasando por estados transitorios de hiperpolarización, o sea de potenciales más negativos, de los de que se tenían inicialmente en el reposo. Medidas de la permeabilidad de la membrana durante la ocurrencia de un potencial de acción, muestran que la despolarización coincide con un aumento de la conductancia del ión Na+, por la apertura de los canales de conducción para este ión. El cambio de la polaridad (inversión), por la entrada masiva de los iones Na+, provoca posteriormente el cierre de los canales para el Na+ e inmediatamente la apertura de los canales para el K+. La salida de los iones K+ contribuye a la repolarización de las membranas.
, por la entrada masiva de los iones Na+, provoca posteriormente el cierre de los canales para el Na+ e inmediatamente la apertura de los canales para el K+. La salida de los iones K+ contribuye a la repolarización de las membranas.")
121
La apertura y cierre de los canales de conducción para el Na+ y K+, principales protagonistas de los cambios de permeabilidad, que afectan sus concentraciones intra y extracelulares durante el potencial de acción, responden a dos tipos de estimulación: 1. Neurotransmisores, como por ejemplo la acetilcolina. El cambio conformacional que ocurre por la unión del neurotransmisor a su receptor específico provoca la apertura del canal iónico. Este es el caso de los canales para Na+ en las membranas post-sinápticas. 2. Cambios de voltaje. Las modificaciones en la polaridad de la membrana (cambios de potencial) provocan el cierre o la apertura de canales sensibles a los cambios de voltaje. Este es el mecanismo de funcionamiento de diversos tipos de canales iónicos, como los de K+ y de Ca+2 y es la manera como el llamado “impulso nervioso” se propaga a través de las membranas de las neuronas y de otras células, a partir de la región sináptica. Los cambios de voltaje en un punto de la membrana, inducirán potenciales de acción en los puntos adyacentes mediante el funcionamiento de los canales iónicos sensibles a los cambios de voltaje.
 provocan el cierre o la apertura de canales sensibles a los cambios de voltaje. Este es el mecanismo de funcionamiento de diversos tipos de canales iónicos, como los de K+ y de Ca+2 y es la manera como el llamado impulso nervioso se propaga a través de las membranas de las neuronas y de otras células, a partir de la región sináptica. Los cambios de voltaje en un punto de la membrana, inducirán potenciales de acción en los puntos adyacentes mediante el funcionamiento de los canales iónicos sensibles a los cambios de voltaje.")
Presentaciones similares







































































.>")

>")



.>")



