Descargar la presentación
La descarga está en progreso. Por favor, espere

1
(Ciclo del ácido cítrico)
Ciclo de Krebs (Ciclo del ácido cítrico) Dr. Juan Pablo Damián Área de Bioquímica Facultad de Veterinaria, Montevideo, Uruguay. UdelaR
 Dr. Juan Pablo Damián. Área de Bioquímica. Facultad de Veterinaria, Montevideo, Uruguay. UdelaR.")
2
Hans Adolf Krebs ciclo de la úrea Alemania Inglaterra
En 1932, se traslada a la Universidad de Friburgo, y en conjunto con el bioquímico Kurt Henseleit, descubre las reacciones involucradas en el ciclo de la úrea Alemania Inglaterra Urea synthesis in mammalian liver [5], Krebs, H.A. Nature 151 (3818), pp. 23
, pp. 23.")
3
Premio Nobel de Fisiología y Medicina, 1953 Krebs y Lipmann
Compuestos ricos en enegría

4
THE INTERMEDIATE METABOLISM OF CARBOHYDRATES
Krebs, H.A The Lancet 230 (5952), pp Cited by
, pp Cited by.")
5
Comprender la importancia del Ciclo de Krebs en el metabolismo.
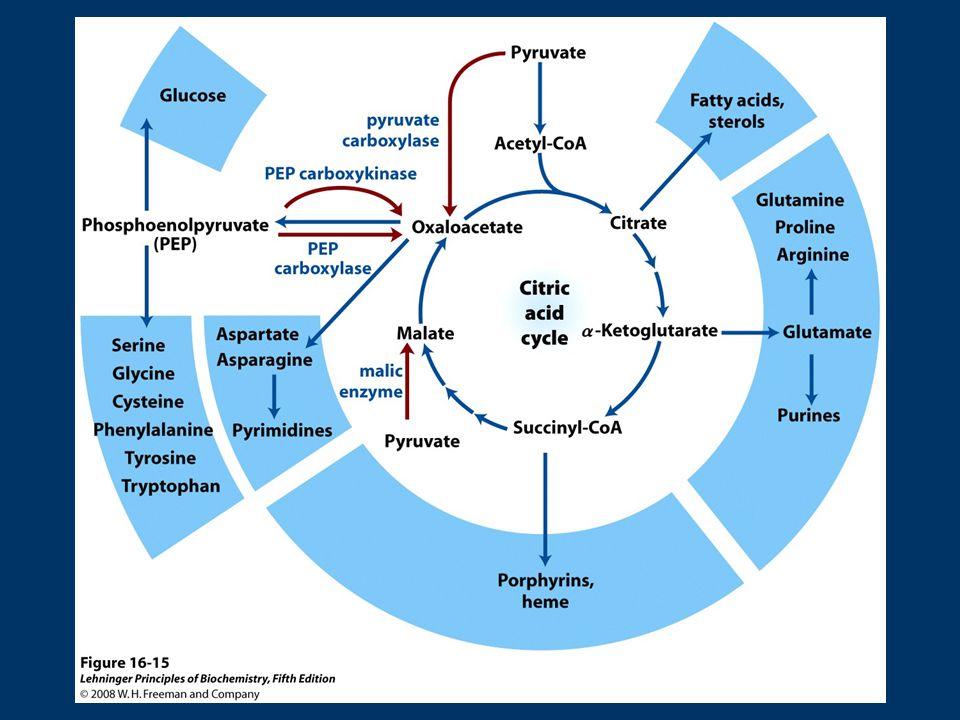
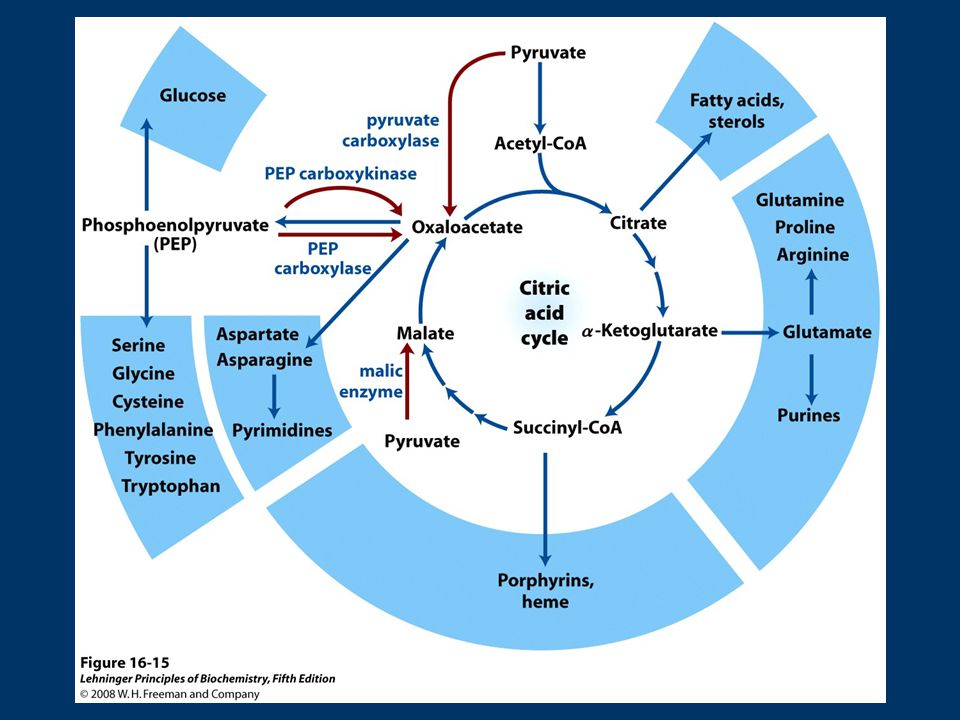
Objetivos: Comprender la importancia del Ciclo de Krebs en el metabolismo. Conocer las reacciones del Ciclo de Krebs y su regulación. 1) Introducción - Fuentes de acetato. - Ubicación del ciclo en el mapa metabólico y en la célula. - Condiciones energéticas 2) Reacciones del ciclo 3) Balance global 4) Regulación 5) Funciones anabólicas 6) Reacciones anapleróticas
 Introducción. - Fuentes de acetato. - Ubicación del ciclo en el mapa metabólico y en la célula. - Condiciones energéticas. 2) Reacciones del ciclo. 3) Balance global. 4) Regulación. 5) Funciones anabólicas. 6) Reacciones anapleróticas.")
6
Ubicación del Ciclo de Krebs en la respiración celular
1) Producción de Acetil-CoA 2) Oxidación de Acetil-CoA 3) Transferencia e- y fosforilación oxidativa Modificado Fig. 16-1 de Lehninger
 Producción de. Acetil-CoA. 2) Oxidación de. Acetil-CoA. 3) Transferencia e- y fosforilación oxidativa. Modificado Fig de Lehninger.")
7
¿En que ruta se oxida Acetil-CoA?
Ciclo de Krebs (ciclo del ácido cítrico) es una ruta cíclica constituida por una secuencia de 8 reacciones. La entrada de casi todos los “combustible” al Ciclo de Krebs es como Acetil-CoA (grupo acetilo activado). O CH3-C-S-CoA Acetil-CoA:
 es una ruta cíclica constituida por una secuencia de 8 reacciones. La entrada de casi todos los combustible al Ciclo de Krebs es como Acetil-CoA (grupo acetilo activado). O CH3-C-S-CoA. Acetil-CoA:")
8
3) Es el principal combustible del ciclo de Krebs.
O CH3-C-S-CoA que es el Acetil-CoA? producto de la degradación de los carbohidratos, lípidos y aminoácidos. 2) compuesto de “alta energía”, ∆G’o = kJ/mol (hidrólisis del enlace tioester, más exergónica que la del ATP que es KJ/mol). 3) Es el principal combustible del ciclo de Krebs.
 compuesto de alta energía , ∆G’o = kJ/mol (hidrólisis del enlace tioester, más exergónica que la del ATP que es KJ/mol). 3) Es el principal combustible del ciclo de Krebs.")
9
¿Donde se realiza el ciclo de Krebs?
Es en la mitocondria donde se realizan todas las reacciones del Ciclo de Krebs Lehninger ( )
")
10
Fases o Etapas de la respiración
Aas tb´dan Piruvato? Mitocondria

11
Ciclo de Krebs (ciclo del ácido cítrico):
Aas tb´dan Piruvato? Ciclo de Krebs (ciclo del ácido cítrico): es una ruta cíclica constituida por una secuencia de 8 reacciones, mediante el cual se oxida el AcetilCoA y se conserva la energía de oxidación en forma de coenzimas reducidos.
: es una ruta cíclica constituida por una secuencia de 8 reacciones, mediante el cual se oxida el AcetilCoA y se conserva la energía de oxidación en forma de coenzimas reducidos.")
12
Ciclo de Krebs Localización: mitocondria 1948 - Kennedy & Lehninger;
Todas las reacciones del ciclo de Krebs se ubican en la mitocondria Donde se producen la mayor parte de las reacciones oxidativas que liberan energía y se acoplan a la síntesis de ATP. Membrana externa Membrana interna Lehninger ( ) Citosol Membrana externa Membrana interna Matriz Cara externa Cara interna Crestas Membrana externa Membrana interna AG a AcetilCoA Degradación Ox de aas Cte- y FOx Matriz Complejo de la Piruvato Deshidrogenasa Enzimas del Ciclo de Krebs (7 matriz y 1 membrana interna)
 Citosol. Membrana externa. Membrana interna. Matriz. Cara externa. Cara interna. Crestas. Membrana externa. Membrana interna. AG a AcetilCoA. Degradación Ox de aas. Cte- y FOx. Matriz. Complejo de la Piruvato Deshidrogenasa. Enzimas del Ciclo de Krebs. (7 matriz y 1 membrana interna)")
13
Membrana externa mitocondria Membrana interna Matriz

14
EN LA MITOCONDRIA, ¿DONDE SE LOCALIZAN LAS ENZIMAS DEL CICLO DE KREBS?
Es en la mitocondria donde se realizan todas las reacciones del Ciclo de Krebs Lehninger ( ) Kennedy & Lehninger 1 en membrana mitocondrial interna 7 en matriz mitocondrial Complejo Piruvato Deshidrogena también en matriz mitocondrial
 Kennedy & Lehninger. 1 en membrana mitocondrial interna. 7 en matriz mitocondrial. Complejo Piruvato Deshidrogena. también en matriz mitocondrial.")
15
Reacciones del Ciclo de Krebs

16
NADH Panorámica del ciclo de Krebs 1 2 8 3 7 4 6 5 oxalacetato
CH3 C SCoA Acetil CoA COO- O O CH2 C 1 2 3 4 5 6 7 8 O C O- HO C COO- oxalacetato Condensación CH2 H C COO- COO- Deshidrogenación H NADH CH2 COO- Isomerización H CH COO- COO- HO C COO- HO C ―H H HC ―H Descarboxilación Oxidativa CO2 COO- CH2 COO- Fosforilación a nivel de sustrato Hidratación COO- CH2 FADH2 COO- C COO- CH COO- GTP CH2 O CH Descarboxilación oxidativa CH2 CH2 COO- CH2 C SCoA Deshidrogenación COO- O CO2

17
Enzima homodiméricacada subunidad tiene 2 dominios, Grande y rígido
Condensación Acetil CoA S-CoA CH3 C CoA-SH H2O O O Citrato sintasa CH2 C Oxalacetato O C COO- CH2 O- 1 Citrato HO C COO- Grupo carbonílico C2 CH2 COO- ∆G’o = kJ/mol Enzima homodiméricacada subunidad tiene 2 dominios, Grande y rígido Y otro pequeño y flexible, el sitio activo se sitúa entre los dos. Unión al OA Forma abierta Forma cerrada

18
Citrato sintasa: se cristalizo y se dedujo su estructura por difracción con rayos X
Forma cerrada Forma abierta 1er sustrato Cada subunidad: dos dominios 1 rígido y otro mas pequeño flexible, SA situado entre los dos Unión de OA induce cambio conformación en el flexible, aparición de sitio de unión para Acetil coA

19
El oxalacetato, primer sustrato que se une al enzima, induce un cambio conformacional en el dominio flexible, que provoca la aparición de un sitio de unión para el segundo sustrato, el acetil-CoA. El AcetilCoA al perder o ceder un protón al Asp 375 se Convierte en un intermediario enolato, este interm. se estabiliza por formación de puentes de hidrógeno con la His274. El enolato se reordena para atacar el carbono carbonílico del oxalacetato, la Hist 320 actúa como Un ácido gral. La condensación restante genera citril-CoA Se forma “citril-CoA” (intermediario) cuya hidrolisis hace que la reacción sea muy exergonica. El tioéster se hidroliza a continuación, regenerando CoA-SH y produciendo citrato.
 cuya hidrolisis. hace que la reacción sea muy exergonica. El tioéster se hidroliza a continuación, regenerando CoA-SH y produciendo citrato.")
20
flechas=movimiento de e- 3 cadenas laterales se orientan p/catalisis
unión tioéster del Acetil-CoA activa los H metilicos y Asp capta protón del grupo metilo dando un intermediario Enolato estabilizado por puentes de H y/o protonización de His El enolato se reordena y ataca el C carbonilo del oxalacetato con la His en posición de captar el protón que había cedido (His actúa como un ácido general) La condensación resultante genera citril CoA
 La condensación resultante genera citril CoA.")
21
Hidrólisis del tioéster regenerando CoA-SH y dando citrato

22
1) Condensación de OA (4 C) y acetil-CoA (2 C) para dar citrato (6 C).
Unión al OA Acetil CoA S-CoA CH3 C CoA-SH H2O O O Citrato sintasa CH2 C Oxalacetato (OA) O C COO- CH2 O- 1 Citrato C HO COO- CH2 COO- ∆G’o = kJ/mol Unión de OA induce cambio conformacional, provoca aparición del sitio de unión para acetil-CoA. Se forma “citril-CoA” (intermediario) cuya hidrolisis hace que la reaccion sea muy exergonica. Reacción fundamental para el funcionamiento global del ciclo debido a la baja concentración de oxalacetato (<10-6M). 1) Condensación de OA (4 C) y acetil-CoA (2 C) para dar citrato (6 C).
 O. C. COO- CH2. O- 1. Citrato. C. HO. COO- CH2. COO- ∆G’o = kJ/mol. Unión de OA induce cambio conformacional, provoca aparición del sitio de unión para acetil-CoA. Se forma citril-CoA (intermediario) cuya hidrolisis hace que la reaccion sea muy exergonica. Reacción fundamental para el funcionamiento global del ciclo debido a la baja concentración de oxalacetato (<10-6M). 1) Condensación de OA (4 C) y acetil-CoA (2 C) para dar citrato (6 C).")
23
Panorámica del ciclo de Krebs
CH3 C SCoA Acetil CoA COO- O O CH2 C O C 1 2 O- HO C COO- Citrato oxalacetato Condensación CH2 H C COO- COO- H CH2 COO- Isomerización H CH COO- Aconitasa (aconitato hidratasa) HO C COO- H Isocitrato
 HO. C. COO- H. Isocitrato.")
24
Isomerización 2 H2O O CH2 C CH2 COO- CH2 COO- O- HO C COO- C COO- H CH COO- H C COO- C COO- HO C COO- aconitasa aconitasa H H H Citrato Isocitrato cis-Aconitato ∆G’o = kJ/mol La aconitasa hidratasa cataliza la transformación reversible del citrato en isocitrato a través de la formación intermedia de ác tricarboxílico cis-aconitato. Que normalmente no se disocia del centro activo Aconitasa cambia OH del C2 al C3 (se forma el intermediario “Cis-aconitato”que no se desprende de la enzima). En la célula la reacción transcurre a la derecha gracias a la rapidez del consumo de isocitrato en el siguiente paso del ciclo, entonces disminuye su concentración
. En la célula la reacción transcurre a la derecha gracias a la rapidez del consumo de isocitrato en el siguiente paso del ciclo, entonces disminuye su concentración.")
25
La aconitasa contiene un Centro ferro-sulfurado que
Isomerización 2 H2O O CH2 C CH2 COO- CH2 COO- O- HO C COO- C COO- H CH COO- H C COO- C COO- HO C COO- aconitasa aconitasa H H H Citrato Isocitrato cis-Aconitato ∆G’o = kJ/mol La aconitasa contiene un Centro ferro-sulfurado que actúa como centro de fijación de sustratos en el centro activo, catalítico y como en la adición o eliminación de agua.

26
Aconitasa Residuo básico de E
Centro ferro-sulfurado Residuo básico de E colabora (posiciona Citrato en sitio activo) 3 Residuos de Cys unidos a Fe, 1 al carboxilo (también interactúa con OH) del citrato carencia de F: pierde centro ferro-sulfurado
 3 Residuos de Cys unidos a Fe, 1 al carboxilo (también interactúa con OH) del citrato. carencia de F: pierde centro ferro-sulfurado.")
27
Isocitrato deshidrogenasa
Panorámica del ciclo de Krebs CH3 C SCoA Acetil CoA COO- O O CH2 C O C 1 2 3 O- HO C COO- Citrato oxalacetato Condensación CH2 H C COO- COO- H CH2 COO- Isomerización H CH COO- HO C COO- NADH H Isocitrato NAD+ CO2 Descarboxilación Oxidativa CH2 COO- Isocitrato deshidrogenasa CH2 3) Descarboxilacion oxidativa C COO- O α-cetoglutarato se oxida el isocitrato y se reduce el NAD+, un grupo carboxilo sale como CO2.
 Descarboxilacion oxidativa. C. COO- O. α-cetoglutarato. se oxida el isocitrato y se reduce el NAD+, un grupo carboxilo sale como CO2.")
28
Isocitrato deshigrogenasa
3. Oxidación del isocitrato a a-cetoglutarato y CO2 eliminación de e- por Mn2+ unido facilita descarboxilación Descarboxilación oxidativa del isocitrato NADH + H+ CO2 intermediario enol NAD+ CH2 COO- CH2 COO- CH2 COO- CH2 COO- H CH COO- H C COO- H C H+ H C H HO C H 1 C O 2 C O- 3 C O- Mn2+ Mn2+ C C C C O- O- O- O- O O O O a-cetoglutarato Isocitrato Oxalosuccinato Isocitrato se oxida a través de la transferencia de un hidruro al NAD+ ∆G’o = -20,9 kJ/mol Reordenamiento del intermediario enol genera α-cetoglutarato Oxidación Transferencia de Ion hidruro Isocitrato deshigrogenasa Eliminación de e- por Mn2+ unido facilita descarboxilación Reordenamiento del intermediario enol genera α-cetoglutarato Existen dos formas diferentes de isocitrato deshidrogenasa: • NAD dependiente (matriz mitocondrial) • NADP dependiente (matriz mitocondrial y citosol)
 • NADP dependiente (matriz mitocondrial y citosol)")
29
NADH Panorámica del ciclo de Krebs 1 2 3 NAD+ 4 NAD+ Citrato
CH3 C SCoA Acetil CoA COO- O O CH2 C O C 1 2 3 O- HO C COO- Citrato oxalacetato Condensación CH2 H C COO- COO- H CH2 COO- Isomerización H CH COO- HO C COO- NADH H Isocitrato Descarboxilación Oxidativa CO2 NAD+ CH2 COO- CH2 COO- COO- C COO- 4 CH2 CH2 O α-cetoglutarato CH2 CH2 CO2 NAD+ CoA-SH C C SCoA SCoA Succinil-CoA O O

30
4. Oxidación de a-cetoglutarato a succinil-CoA y CO2
Descarboxilación oxidativa del a-cetoglutarato NADH + H+ CoA-SH NAD+ CH2 COO- CH2 COO- H C H H C H CO2 C O- + C S-CoA C O- O O ∆G’o = kJ/mol Succinil-CoA a-cetoglutarato Complejo multienzimático de la “α-cetoglutarato deshidrogenasa” Posee 3 enzimas homólogos a (E1, E2, E3) de la piruvato desh 5 Cofactores (TPP, lipoato, FAD, CoA y NAD) Paso 4: Descarboxilación oxidativa El complejo de la α-cetoglutarato deshidrogenasa es muy parecido al complejo piruvato deshidrogenasa, tanto en estructura como en función
 de la piruvato desh. 5 Cofactores (TPP, lipoato, FAD, CoA y NAD) Paso 4: Descarboxilación oxidativa. El complejo de la α-cetoglutarato. deshidrogenasa es muy parecido al. complejo piruvato deshidrogenasa, tanto en estructura como en función.")
31
NADH Panorámica del ciclo de Krebs 1 2 3 NAD+ 4 Succinil-CoA sintetasa
CH3 C SCoA Acetil CoA COO- O O CH2 C O C 1 2 3 O- HO C COO- Citrato oxalacetato Condensación CH2 H C COO- COO- H CH2 COO- Isomerización H CH COO- HO C COO- NADH H Isocitrato Descarboxilación Oxidativa CO2 NAD+ CH2 COO- Fosforilación a nivel de sustrato CH2 COO- COO- C COO- COO- 4 GTP CH2 CH2 O α-cetoglutarato CH2 CH2 CH2 CO2 Succinil-CoA sintetasa 5 NAD+ CH2 CoA-SH C C SCoA SCoA Succinato COO- Succinil-CoA GDP O O

32
5. Conversión del Succinil-CoA e Succinato
Fosforilación a nivel de sustrato CoA-SH GDP + Pi GTP CH2 COO- COO- H C H H C H C S-CoA C H2 O ∆G’o = -2,9 kJ/mol COO- Succinil-CoA Succinato Succinil-CoA sintetasa La formación acoplada de GTP (o ATP) a expensas de la energía liberada por la decarboxilación oxidativa del α-ceto ceto-glutarato glutarato es otro ejemplo de fosforilación a nivel del sustrato
 a. expensas de la energía liberada por la. decarboxilación oxidativa del. α-ceto ceto-glutarato glutarato es otro ejemplo de fosforilación a nivel del sustrato.")
33
5. Conversión del Succinil-CoA en Succinato
CoA-SH GDP + Pi GTP CH2 COO- COO- H C H H C H C S-CoA C H2 O ∆G’o = -2,9 kJ/mol COO- Succinil-CoA Succinato Succinil-CoA sintetasa El succinil-CoA tiene un enlace tioéster con una energía libre de estándar de Hidrólisis que es altamente negativa (∆G’o = -36 kJ/mol). La energía liberada en La rotura de este enlace se utiliza para promover la síntesis de un enlace Fosfoanhídrido del GTP o del ATP con una variación neta de ∆G’o = -2,9 kJ/mol La formación acoplada de GTP (o ATP) a expensas de la energía liberada por la decarboxilación oxidativa del α-ceto ceto-glutarato glutarato es otro ejemplo de fosforilación a nivel del sustrato
. La energía liberada en. La rotura de este enlace se utiliza para promover la síntesis de un enlace. Fosfoanhídrido del GTP o del ATP con una variación neta de ∆G’o = -2,9 kJ/mol. La formación acoplada de GTP (o ATP) a. expensas de la energía liberada por la. decarboxilación oxidativa del. α-ceto ceto-glutarato glutarato es otro ejemplo de fosforilación a nivel del sustrato.")
34
Succinil-CoA sintetasa
En tres pasos: El sucinil-CoA se une al enzima, y un grupo fosforilo sustituye al CoA del succinil-CoA, y se forma un acil fosfato de alta energía. El succinil fosfato dona el grupo fosforilo a un residuo de His del enzima, y se forma un fosfohistidil enzima de elevada energía El grupo fosforilo es transferido desde el res His al fosfato terminal de GDP formando GTP.

35
5. Conversión del Succinil-CoA e Succinato
Fosforilación a nivel de sustrato CoA-SH GDP + Pi GTP CH2 COO- COO- H C H H C H C S-CoA C H2 O ∆G’o = -2,9 kJ/mol COO- Succinil-CoA Succinato Succinil-CoA sintetasa Nucleosido difosfatasa quinasa GTP + ADP GDP + ATP ΔG´°= 0 kJ/mo

36
NADH Panorámica del ciclo de Krebs 1 2 3 Succinato deshidrogenasa
CH3 C SCoA Acetil CoA COO- O O CH2 C O C 1 2 3 O- HO C COO- Citrato oxalacetato Condensación CH2 H C COO- COO- H CH2 COO- Isomerización H CH COO- HO C COO- NADH H Isocitrato Succinato deshidrogenasa membrana mitocondrial interna Descarboxilación Oxidativa CO2 6) Oxido-reducción. NAD+ CH2 COO- Fosforilación a nivel de sustrato COO- CH2 FADH2 COO- COO- C COO- CH COO- 4 GTP CH2 CH2 O α-cetoglutarato CH 6 CH2 Fumarato CH2 CH2 COO- CO2 5 NAD+ CH2 CoA-SH C C SCoA SCoA Succinato COO- Succinil-CoA GDP O O
 Oxido-reducción. NAD+ CH2. COO- Fosforilación a. nivel de sustrato. COO- CH2. FADH2. COO- COO- C. COO- CH. COO- 4. GTP. CH2. CH2. O. α-cetoglutarato. CH. 6. CH2. Fumarato. CH2. CH2. COO- CO2. 5. NAD+ CH2. CoA-SH. C. C. SCoA. SCoA. Succinato. COO- Succinil-CoA. GDP. O. O.")
37
6. Oxidación del Succinato a Fumarato
FAD FADH2 COO- COO- C H2 C H C H2 Succinato deshidrogenasa H C COO- COO- Succinato Fumarato ∆G’o = 0 kJ/mol En eucariotas, la succinato deshidrogenasa se encuentra unida a la membrana mitocondrial interna, contiene tres centros hierro-azufre diferentes y una molécula de FAD unida covalentemente.

38
6. Oxidación del Succinato a Fumarato
FAD FADH2 COO- COO- C H2 C H C H2 Succinato deshidrogenasa H C COO- COO- Succinato Fumarato ∆G’o = 0 kJ/mol Membrana externa Cara externa Citosol Cara interna En eucariotas, la succinato deshidrogenasa se encuentra unida a la membrana mitocondrial interna, contiene tres centros hierro-azufre diferentes y una molécula de FAD unida covalentemente. Cara externa Cara interna Matriz

39
Complejo II: Succinato Deshidrogenasa
Ciclo de Krebs Succinato Fumarato FAD C CH2 O O- CH FADH2 Q QH2

40
Complejo II: Succinato Deshidrogenasa
FADH2 + Q FAD + QH2 Succinato Fumarato ΔG´°= -16,4 Kj/mol Succinato Fumarato FAD C CH2 O O- CH FADH2 Q QH2

41
6. Oxidación del Succinato a Fumarato
FAD FADH2 COO- CH2 COO- C H2 C H C H2 Succinato deshidrogenasa H C COO- COO- Succinato Fumarato En eucariotas, la succinato deshidrogenasa se encuentra unida a la membrana mitocondrial interna, contiene tres centros hierro-azufre diferentes y una molécula de FAD unida covalentemente.

42
NADH Panorámica del ciclo de Krebs 1 2 3 Succinato deshidrogenasa NAD+
CH3 C SCoA Acetil CoA COO- O O CH2 C O C 1 2 3 O- HO C COO- Citrato oxalacetato Condensación CH2 H C COO- COO- H CH2 COO- Isomerización COO- Malonato (análogo succinato) CH2 H CH COO- inhibidor competitivo HO C COO- NADH H Isocitrato Succinato deshidrogenasa Descarboxilación Oxidativa CO2 NAD+ CH2 COO- Fosforilación a nivel de sustrato COO- CH2 FADH2 COO- COO- C COO- CH COO- 4 GTP CH2 CH2 O α-cetoglutarato CH 6 CH2 Fumarato CH2 CH2 COO- CO2 5 NAD+ CH2 CoA-SH C C SCoA SCoA Succinato COO- Succinil-CoA GDP O O
 CH2. H. CH. COO- inhibidor competitivo. HO. C. COO- NADH. H. Isocitrato. Succinato deshidrogenasa. Descarboxilación. Oxidativa. CO2. NAD+ CH2. COO- Fosforilación a. nivel de sustrato. COO- CH2. FADH2. COO- COO- C. COO- CH. COO- 4. GTP. CH2. CH2. O. α-cetoglutarato. CH. 6. CH2. Fumarato. CH2. CH2. COO- CO2. 5. NAD+ CH2. CoA-SH. C. C. SCoA. SCoA. Succinato. COO- Succinil-CoA. GDP. O. O.")
43
NADH Panorámica del ciclo de Krebs 1 2 3
CH3 C SCoA Acetil CoA COO- O O CH2 C O C 1 2 3 O- HO C COO- Citrato oxalacetato Condensación CH2 H C COO- COO- H Fumarasa (fumarato hidratasa), estereoespecífica CH2 COO- Isomerización H CH COO- COO- HO C COO- NADH HO C ―H H Isocitrato HC ―H L-Malato Descarboxilación Oxidativa CO2 COO- NAD+ 7 CH2 COO- Fosforilación a nivel de sustrato Hidratación COO- CH2 FADH2 COO- COO- C COO- CH COO- 4 GTP CH2 CH2 O α-cetoglutarato CH 6 CH2 Fumarato CH2 CH2 COO- CO2 5 NAD+ CH2 CoA-SH C C SCoA SCoA Succinato COO- Succinil-CoA GDP O O
, estereoespecífica. CH2. COO- Isomerización. H. CH. COO- COO- HO. C. COO- NADH. HO. C ―H. H. Isocitrato. HC ―H. L-Malato. Descarboxilación. Oxidativa. CO2. COO- NAD+ 7. CH2. COO- Fosforilación a. nivel de sustrato. Hidratación. COO- CH2. FADH2. COO- COO- C. COO- CH. COO- 4. GTP. CH2. CH2. O. α-cetoglutarato. CH. 6. CH2. Fumarato. CH2. CH2. COO- CO2. 5. NAD+ CH2. CoA-SH. C. C. SCoA. SCoA. Succinato. COO- Succinil-CoA. GDP. O. O.")
44
7. Hidratación del Fumarato a Malato.
COO- COO- H2O C H HO C H H C H C H COO- Fumarasa COO- Fumarato L-Malato ∆G’o = -3,8 kJ/mol Cataliza la hidratación del doble enlace en trans del fumarato Esta enzima es específica para el fumarato y el L-malato

45
Cis Trans

46
7. Hidratación del Fumarato a Malato.
COO- COO- H2O C H HO C H H C H C H COO- Fumarasa COO- Fumarato L-Malato ∆G’o = -3,8 kJ/mol COO- COO- OH- COO- H+ C H HO C H HO C H H C C H C H H COO- Fumarasa COO- COO- Fumarato L-Malato Estado de transición carbanión Esta enzima es específica para el fumarato y el L-malato

47
NADH Panorámica del ciclo de Krebs 1 2 3 NAD+ 8 NAD+ 7 4 6 5 NAD+
CH3 C SCoA Acetil CoA COO- O O CH2 C O C 1 2 3 O- HO C COO- Citrato oxalacetato Condensación CH2 NAD+ H C COO- COO- H Deshidrogenación CH2 COO- 8 Isomerización H CH COO- COO- HO C COO- NADH HO C ―H H Isocitrato HC ―H L-Malato Descarboxilación Oxidativa CO2 COO- NAD+ 7 CH2 COO- Fosforilación a nivel de sustrato Hidratación COO- CH2 FADH2 COO- COO- C COO- CH COO- 4 GTP CH2 CH2 O α-cetoglutarato CH 6 CH2 Fumarato CH2 CH2 COO- CO2 5 NAD+ CH2 CoA-SH C C SCoA SCoA Succinato COO- Succinil-CoA GDP O O

48
8. Oxidación del malato a oxalacetato
NADH + H+ NAD+ COO- COO- O C HO C H H C H H C H COO- COO- L- Malato deshidrogenasa Oxalacetato L-Malato ∆G’o = 29,7 kJ/mol El Oxalacetato es eliminado continuamente por la reacción altamente exergónica de la citrato sintasa. Esto mantiene la concentración de oxalacetato relativamente baja <106M, lo que empuja la reacción de la malato deshidrogenasa hacia la formación de oxalacetato 1 sola molécula de Oxalacetato es usada para catalizar un gran número de Acetil CoA

49
equilibrio de reacción desplazado a la formación de malato
Panorámica del ciclo de Krebs CH3 C SCoA Acetil CoA COO- O O CH2 C O C 1 2 3 O- HO C COO- Citrato oxalacetato Condensación CH2 NAD+ ∆G’o = kJ/mol H C COO- COO- H Deshidrogenación CH2 COO- 8 Isomerización ∆G’o = 29,7 kJ/mol H CH COO- COO- HO C COO- equilibrio de reacción desplazado a la formación de malato [oxalacetato] (<10-6M) si es utilizado continuamente (citrato sintasa) empuja la reacción a la formación de oxalacetato NADH HO C ―H H Isocitrato HC ―H L-Malato Descarboxilación Oxidativa CO2 COO- NAD+ 7 CH2 COO- Fosforilación a nivel de sustrato Hidratación COO- CH2 FADH2 COO- COO- C COO- CH COO- 4 GTP CH2 CH2 O α-cetoglutarato CH 6 CH2 Fumarato CH2 CH2 COO- CO2 5 NAD+ CH2 CoA-SH C C SCoA SCoA Succinato COO- Succinil-CoA GDP O O
 si es utilizado continuamente (citrato sintasa) empuja la reacción a la formación de oxalacetato. NADH. HO. C ―H. H. Isocitrato. HC ―H. L-Malato. Descarboxilación. Oxidativa. CO2. COO- NAD+ 7. CH2. COO- Fosforilación a. nivel de sustrato. Hidratación. COO- CH2. FADH2. COO- COO- C. COO- CH. COO- 4. GTP. CH2. CH2. O. α-cetoglutarato. CH. 6. CH2. Fumarato. CH2. CH2. COO- CO2. 5. NAD+ CH2. CoA-SH. C. C. SCoA. SCoA. Succinato. COO- Succinil-CoA. GDP. O. O.")
51
Comprender la importancia del Ciclo de Krebs en el metabolismo.
Objetivos: Comprender la importancia del Ciclo de Krebs en el metabolismo. Conocer las reacciones del Ciclo de Krebs y su regulación. 1) Introducción - Fuentes de acetato. - Ubicación del ciclo en el mapa metabólico y en la célula. - Condiciones energéticas 2) Reacciones del ciclo 3) Balance global 4) Regulación 5) Funciones anabólicas 6) Reacciones anapleróticas
 Introducción. - Fuentes de acetato. - Ubicación del ciclo en el mapa metabólico y en la célula. - Condiciones energéticas. 2) Reacciones del ciclo. 3) Balance global. 4) Regulación. 5) Funciones anabólicas. 6) Reacciones anapleróticas.")
52
Descarboxilación oxidativa
proteínas aminoácidos glucógeno glucosa triacilglicéridos ácidos grasos Acetil CoA piruvato Descarboxilación oxidativa del Piruvato 2C Ciclo de Krebs 4 C 6 C 4C + 1 sola molécula de Oxalacetato es usada para catalizar un gran número de Acetil-CoA CO2 CO2

53
NADH Panorámica del ciclo de Krebs 1 2 3 NAD+ 8 NAD+ 7 4 6 5 NAD+
CH3 C SCoA Acetil CoA COO- O O CH2 C O C 1 2 3 O- HO C COO- Citrato oxalacetato Condensación CH2 NAD+ H C COO- COO- H Deshidrogenación CH2 COO- 8 Isomerización H CH COO- COO- HO C COO- NADH HO C ―H H Isocitrato HC ―H L-Malato Descarboxilación Oxidativa CO2 COO- NAD+ 7 CH2 COO- Fosforilación a nivel de sustrato Hidratación COO- CH2 FADH2 COO- COO- C COO- CH COO- 4 GTP CH2 CH2 O α-cetoglutarato CH 6 CH2 Fumarato CH2 CH2 COO- CO2 5 NAD+ CH2 CoA-SH C C SCoA SCoA Succinato COO- Succinil-CoA GDP O O

54
NADH Panorámica del ciclo de Krebs 1 2 3 NAD+ 8 NAD+ 7 4 6 5 NAD+
CH3 C SCoA Acetil CoA COO- O O CH2 C O C 1 2 3 O- HO C COO- Citrato oxalacetato Condensación CH2 NAD+ H C COO- COO- H Deshidrogenación CH2 COO- 8 Isomerización H CH COO- COO- HO C COO- NADH HO C ―H H Isocitrato HC ―H L-Malato Descarboxilación Oxidativa CO2 COO- NAD+ 7 CH2 COO- Fosforilación a nivel de sustrato Hidratación COO- CH2 FADH2 COO- COO- C COO- CH COO- 4 GTP CH2 CH2 O α-cetoglutarato CH 6 CH2 Fumarato CH2 CH2 COO- CO2 5 NAD+ CH2 CoA-SH C C SCoA SCoA Succinato COO- Succinil-CoA GDP O O

55
Balance global del Ciclo de Krebs

56
NADH Panorámica del ciclo de Krebs 1 2 3 NAD+ 8 NAD+ 7 4 6 5 NAD+
CH3 C SCoA Acetil CoA COO- O O CH2 C O C 1 2 3 O- HO C COO- Citrato oxalacetato Condensación CH2 NAD+ H C COO- COO- H Deshidrogenación CH2 COO- 8 Isomerización H CH COO- COO- HO C COO- NADH HO C ―H H Isocitrato HC ―H L-Malato Descarboxilación Oxidativa CO2 COO- NAD+ 7 CH2 COO- Fosforilación a nivel de sustrato Hidratación COO- CH2 FADH2 COO- COO- C COO- CH COO- 4 GTP CH2 CH2 O α-cetoglutarato CH 6 CH2 Fumarato CH2 CH2 COO- CO2 5 NAD+ CH2 CoA-SH C C SCoA SCoA Succinato COO- Succinil-CoA GDP O O

57
Ecuaciones igualadas para cada reacción del Ciclo de Krebs
Entradas salidas 1) Oxalacetato + Acetil-CoA + H2O Citrato + CoA 2) Citrato + H2O Isocitrato + H2O 3) Isocitrato + NAD+ α-cetoglutarato + NADH + H+ + CO2 4) α-cetoglutarato + NAD+ + CoA Succinil-CoA + CO2 + NADH + H+ 5) Succinil-CoA + GDP + Pi Succinato + GTP + CoA 6) Succinato + FAD Fumarato + FADH2 7) Fumarato + H2O L-malato 8) L-malato + NAD+ Oxalacetato + NADH + H+
 Oxalacetato + Acetil-CoA + H2O. Citrato + CoA. 2) Citrato + H2O. Isocitrato + H2O. 3) Isocitrato + NAD+ α-cetoglutarato + NADH + H+ + CO2. 4) α-cetoglutarato. + NAD+ + CoA. Succinil-CoA + CO2. + NADH + H+ 5) Succinil-CoA + GDP + Pi. Succinato + GTP + CoA. 6) Succinato + FAD. Fumarato + FADH2. 7) Fumarato + H2O. L-malato. 8) L-malato + NAD+ Oxalacetato + NADH + H+")
58
Entradas: Acetil-CoA + 3 NAD+ + FAD + GDP + Pi + 2 H2O
Ecuaciones igualadas para cada reacción del Ciclo de Krebs Entradas salidas 1) Oxalacetato + Acetil-CoA + H2O Citrato + CoA 2) Citrato + H2O Isocitrato + H2O 3) Isocitrato + NAD+ α-cetoglutarato + NADH + H+ + CO2 4) α-cetoglutarato + NAD+ + CoA Succinil-CoA + CO2 + NADH + H+ 5) Succinil-CoA + GDP + Pi Succinato + GTP + CoA 6) Succinato + FAD Fumarato + FADH2 7) Fumarato + H2O L-malato 8) L-malato + NAD+ Oxalacetato + NADH + H+ Entradas: Acetil-CoA + 3 NAD+ + FAD + GDP + Pi + 2 H2O Salidas: 3 NADH + 3 H+ + FADH2 + 2 CO2 + GTP + CoA
 Oxalacetato + Acetil-CoA + H2O. Citrato + CoA. 2) Citrato + H2O. Isocitrato + H2O. 3) Isocitrato + NAD+ α-cetoglutarato + NADH + H+ + CO2. 4) α-cetoglutarato. + NAD+ + CoA. Succinil-CoA + CO2. + NADH + H+ 5) Succinil-CoA + GDP + Pi. Succinato + GTP + CoA. 6) Succinato + FAD. Fumarato + FADH2. 7) Fumarato + H2O. L-malato. 8) L-malato + NAD+ Oxalacetato + NADH + H+ Entradas: Acetil-CoA + 3 NAD+ + FAD + GDP + Pi + 2 H2O. Salidas: 3 NADH + 3 H+ + FADH2 + 2 CO2 + GTP + CoA.")
59
Balance Global Energía Acetil CoA + 3 NAD+ + FAD + GDP + Pi + 2 H2O
Acetil CoA (2 C) Oxalacetato (4 C) Citrato (6 C) NADH +H+ NAD+ Balance Global Isocitrato (6 C) L-Malato (4 C) Acetil CoA + 3 NAD+ + FAD + GDP + Pi + 2 H2O CoA + 2 CO2 + 3 NADH + FADH2 + GTP NAD+ H2O Energía 3 NADH + FADH2 + GTP NADH +H+ CO2 FADH2 Fumarato (4 C) -cetoglutarato (5 C) FAD NAD+ GTP GDP + Pi CoASH NADH +H+ Succinato (4 C) CoASH CO2 Succinil CoA (4 C)
 Oxalacetato. (4 C) Citrato (6 C) NADH +H+ NAD+ Balance Global. Isocitrato (6 C) L-Malato (4 C) Acetil CoA + 3 NAD+ + FAD + GDP + Pi + 2 H2O. CoA + 2 CO2 + 3 NADH + FADH2 + GTP. NAD+ H2O. Energía. 3 NADH + FADH2 + GTP. NADH +H+ CO2. FADH2. Fumarato (4 C) -cetoglutarato. (5 C) FAD. NAD+ GTP. GDP + Pi. CoASH. NADH +H+ Succinato (4 C) CoASH. CO2. Succinil CoA (4 C)")
60
Destino de los Carbonos
Panorámica del ciclo de Krebs CH3 C SCoA Acetil CoA COO- O O CH2 C O C 1 2 3 O- HO C COO- Citrato oxalacetato CH2 H C COO- COO- H CH2 COO- 8 H CH COO- Destino de los Carbonos del Acetil CoA COO- HO C COO- HO C ―H H Isocitrato HC ―H L-Malato Los 2 C del Acetil CoA salen en una vuelta del ciclo? CO2 COO- 7 CH2 COO- COO- CH2 COO- COO- C COO- CH COO- 4 CH2 CH2 O α-cetoglutarato CH 6 CH2 Fumarato CH2 CH2 COO- CO2 5 CH2 C C SCoA SCoA Succinato COO- Succinil-CoA O O

61
y fosforilación oxidativa
Respiración celular 2) Oxidación de Acetil-CoA produce NADH FADH2 ¿Donde se reoxidan? Cadena de transporte de e- 3) Transferencia e- y fosforilación oxidativa produce ATP Modificado Fig de Lehninger
 Oxidación de. Acetil-CoA. produce. NADH FADH2. ¿Donde se reoxidan Cadena de transporte de e- 3) Transferencia e- y fosforilación oxidativa. produce. ATP. Modificado Fig de Lehninger.")
62
Modelo quimiosmótico: e- del NADH, FADH2 y otros sustratos oxidables se reoxidan en la CT e-

63
Rendimiento energético en cada vuelta del Cilo de Krebs
3 NADH + 1 FADH2 +1 GTP 3 x 2.5 ATP + 1 x 1.5 ATP +1 ATP 10 ATP En la Fosforilación Oxidativa: 1 NADH ,5 ATP 1 FADH ,5 ATP

64
2) Fosfoenolpiruvato a 2) piruvato 2) Piruvato a 2) acetil-CoA 5
Reducción de CoE y formación de ATP en oxidación aeróbica de la glucosa (glucólisis, descarboxilación oxidativa del pir., CK, Cte y FO) Reacción ATP/Co E ATP Glucosa a G-6-P - 1 ATP -1 F-6-P a F1,6BP 2) GAP a 2) 1,3BPG 2 NADH 5 (3) 2) 1,3BPG a 2) 3PG 2 ATP 2 2) Fosfoenolpiruvato a 2) piruvato 2) Piruvato a 2) acetil-CoA 5 2) Isocitrato a 2) α-cetoglutarato 2) α-cetoglutarato a 2) succinil-CoA 2) succinil-CoA a 2) succinato 2 ATP (2GTP) 2) Succinato a 2) fumarato 2 FADH2 3 2) Malato a 2) oxalacetato Total 30-32 Glucólisis 5 o 7 ATP PDH 10 ATP/vuelta C. K. X 2= 20 ATP
 Reacción. ATP/Co E. ATP. Glucosa a G-6-P. - 1 ATP. -1. F-6-P a F1,6BP. 2) GAP a 2) 1,3BPG. 2 NADH. 5 (3) 2) 1,3BPG a 2) 3PG. 2 ATP. 2. 2) Fosfoenolpiruvato a 2) piruvato. 2) Piruvato a 2) acetil-CoA. 5. 2) Isocitrato a 2) α-cetoglutarato. 2) α-cetoglutarato a 2) succinil-CoA. 2) succinil-CoA a 2) succinato. 2 ATP (2GTP) 2) Succinato a 2) fumarato. 2 FADH ) Malato a 2) oxalacetato. Total Glucólisis. 5 o 7 ATP. PDH. 10 ATP/vuelta. C. K. X 2= 20 ATP.")
65
Fases o Etapas de la respiración
Aas tb´dan Piruvato? Mitocondria

66
Sin Oxígeno, el piruvato no entra en la mitocondria
Oxidándose de forma incompleta a lactato. ATP ATP Aas tb´dan Piruvato? Lactato

67
Fases o Etapas de la respiración
En presencia de Oxígeno…. Aas tb´dan Piruvato? ATP ATP ATP ATP ATP ATP ATP ATP ATP ATP ATP ATP ATP ATP ATP ATP ATP ATP ATP ATP ATP ATP ATP ATP ATP ATP ATP ATP ATP ATP ATP ATP ATP

68
energía de oxidaciones de C K se conserva eficientemente!!
Partiendo de glucosa se generan ATP ¿Qué porcentaje de la “energía de la glucosa” se conserva en forma de ATP? Datos: glucosa a CO2 y H2O en calorímetro ∆G’o = kJ/mol Hidrólisis de ATP a ADP + Pi ∆G’o = kJ/mol Síntesis de 1 ATP … kJ/mol 32 ATP …… X = 976 kJ/mol 2840 kJ/mol ….. 100% 976 kJ/mol …... X = 34 % energía de oxidaciones de C K se conserva eficientemente!! 34 % en condiciones estándar 65 % en condiciones fisiológicas

69
Regulación del Ciclo de Krebs

70
Reacciones 1, 3 y 4 son irreversibles reacciones de control del flujo
Variaciones de energía libre (kJ/mol) en condiciones estándar y fisiológicas de las reacciones del Ciclo de Krebs Enzima ∆G’o ∆G’ 1 citrato sintasa -31.5 negativo 2 aconitasa +5 0 3 isocitrato deshidrogenasa -21 4 complejo α-cetoglutarato deshidrogenasa -33 5 succinil-CoA sintetasa -2.1 6 succinato dehidrogenasa +6 7 fumarasa -3.4 8 malato deshidrogenasa +29.7 Reacciones 1, 3 y 4 son irreversibles reacciones de control del flujo
 en condiciones estándar y fisiológicas de las reacciones del Ciclo de Krebs. Enzima. ∆G’o. ∆G’ 1. citrato sintasa negativo. 2. aconitasa. +5. isocitrato deshidrogenasa complejo α-cetoglutarato deshidrogenasa succinil-CoA sintetasa succinato dehidrogenasa fumarasa malato deshidrogenasa Reacciones 1, 3 y 4 son irreversibles reacciones de control del flujo.")
71
Reacciones alejadas del equilibrio
Acetil CoA (2 C) 1 2 3 4 5 6 7 8 Citrato Sintasa Aconitasa Isocitrato desidrogenasa Complejo de la Cetoglutarato deshidrogenasa Succinil CoA Sintetasa Succinato Deshidrogenasa Malato Deshidrogenasa Fumarasa Oxalacetato (4 C) Citrato (6 C) Reacciones alejadas del equilibrio NADH FADH2 Isocitrato (6 C) ∆G’ negativo L-Malato (4 C) 3 reacciones irreversibles 5 reacciones reversibles CO2 -cetoglutarato (5 C) GTP Fumarato (4 C) CO2 Succinato (4 C) Succinil CoA (4 C)
 Citrato Sintasa. Aconitasa. Isocitrato. desidrogenasa. Complejo de la. Cetoglutarato. deshidrogenasa. Succinil CoA Sintetasa. Succinato Deshidrogenasa. Malato. Deshidrogenasa. Fumarasa. Oxalacetato. (4 C) Citrato (6 C) Reacciones alejadas del equilibrio. NADH. FADH2. Isocitrato (6 C) ∆G’ negativo. L-Malato (4 C) 3 reacciones irreversibles. 5 reacciones reversibles. CO2. -cetoglutarato. (5 C) GTP. Fumarato (4 C) CO2. Succinato (4 C) Succinil CoA (4 C)")
72
Regulación estricta pero sencilla
Consumo de O2 reoxidación del NADH están acopladas producción de ATP Mecanismo de regulación por retroalimentación que coordina producción de NADH con consumo energético. Regulación por: - disponibilidad de sustrato - inhibición por producto - inhibición competitiva por intermediarios del ciclo Acetil-CoA y oxalacetato NADH Citrato, succinil-CoA, ATP [acetil-Coa] y [oxalacetato] normalmente no saturan la citrato sintasa

73
Reacción 1, E Citrato sintasa:
inhibida por: alta [NADH], [ATP], [citrato], [succinil-CoA] inhibidores competitivos: citrato de oxalacetato succinil-CoA de CoA aumenta actividad por: [acetil-CoA] y [ oxalacetato] (sustratos, no saturan E) Reacción 3, E Isocitrato deshidrogenasa: inhibida por: alta [NADH], [ATP] (- producto) activada por: alta [ADP], [Ca2+] Reacción 4, E Complejo α-cetoglutarato deshidrogenasa inhibida por: alta [NADH] y [succinil-CoA] (- producto) activada por: alta [ADP], [Ca2+]. Estas enzimas constituyen puntos en los que se regula el flujo a través del ciclo
 Reacción 3, E Isocitrato deshidrogenasa: inhibida por: alta [NADH], [ATP] (- producto) activada por: alta [ADP], [Ca2+] Reacción 4, E Complejo α-cetoglutarato deshidrogenasa. inhibida por: alta [NADH] y [succinil-CoA] (- producto) activada por: alta [ADP], [Ca2+]. Estas enzimas constituyen puntos en los que se regula el flujo a través del ciclo.")
74
NADH Regulación covalente Regulación alostérica Piruvato
Complejo de la Deshidrogenasa E2 E1 E3 P I II Protein kinasa fosfatasa E2 E1 E3 OH I II Protein kinasa fosfatasa CoA,NAD+, AMP, Ca2+ ATP NADH, Acetil CoA NADH, ATP, Succinil CoA, citrato ATP, Succinil CoA, NADH Acetil CoA ADP, Ca2+ NADH Citrato sintasa Isocitrato deshidrogenasa cetoglutarato Oxalacetato ADP, Acetil CoA, Oxalacetato Citrato Inactiva E1 Activa E1 Isocitrato L-Malato NADH -cetoglutarato Fumarato FADH2 GTP Succinato Succinil CoA

75
Canalización de sustratos contribuye a la catálisis
En C de K Puede darse canalización de sustratos Elevada concentración de E favorece su asociación Extracto de células rotas la dilución en buffer disminuye la 100 a 1000 veces

76
Por que es tan complicada la oxidación del Acetato a CO2?
Ciclo de Krebs es el núcleo del metabolismo intermediario. Productos de muchos procesos catabólicos alimentan el Ciclo. Intermediarios del ciclo son utilizados como precursores en distintas vías metabólicas!!!! No representa la vía más corta de Acetato a CO2 pero es la que a través del tiempo ha conferido mayores ventajas selectivas.

77
Ciclo de Krebs tiene funciones catabólicas y anabólicas Vía Anfibólica
Involucra procesos de degradación y conservación de energía (catabolismo) Intermediarios del ciclo solo se requieren en concentraciones catalíticas para mantener la función degradativa del Ciclo Sin embargo… Varias vías biosintéticas utilizan intermediarios del ciclo como material de inicio en la biosíntesis (anabolismo) tiene funciones catabólicas y anabólicas Vía Anfibólica
 Intermediarios del ciclo solo se requieren en concentraciones catalíticas para mantener la función degradativa del Ciclo. Sin embargo… Varias vías biosintéticas utilizan intermediarios del ciclo como material de inicio en la biosíntesis. (anabolismo) tiene funciones catabólicas y anabólicas. Vía Anfibólica.")
78
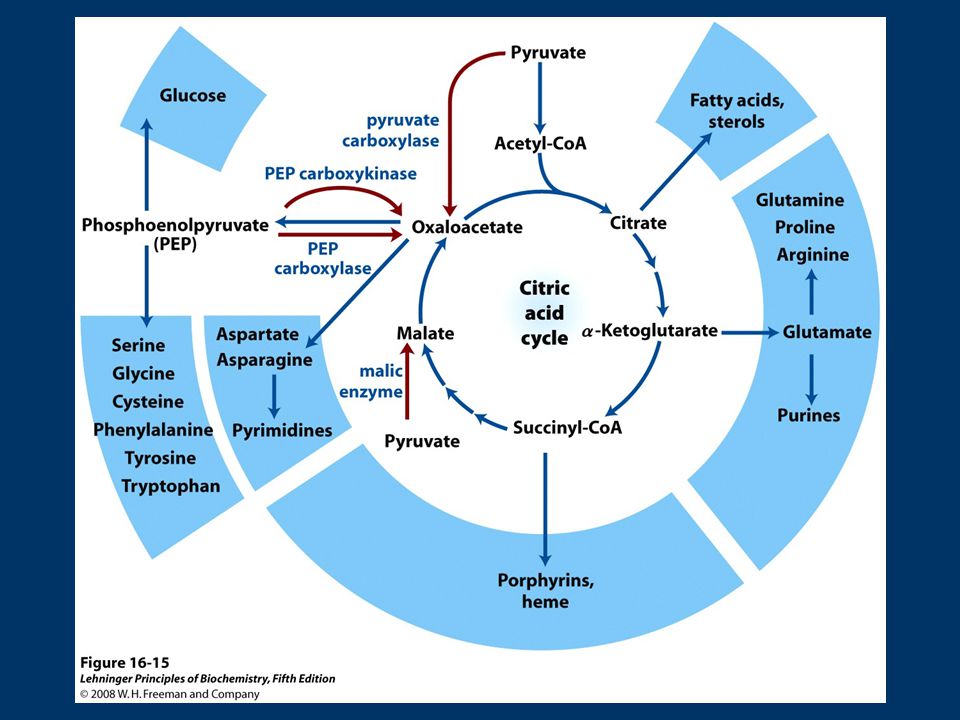
Ciclo de Krebs Las reacciones que utilizan (“agotan”) intermediarios del ciclo se denominan catapletoricas (del griego “cata” = descendente “pletorikos”= llenar) Gluconeogénesis (OA): biosíntesis de glucosa implica la conversión de OA en malato o aspartato para transportarse al citosol Biosíntesis de lípidos (citrato): síntesis de AG en citosol, requiere acetil-Co A que se produce en mitocondria, sale como citrato ATP+citrato+CoA ADP+Pi+OA+Acetil CoA Citrato liasa
 intermediarios del ciclo se denominan catapletoricas (del griego cata = descendente pletorikos = llenar) Gluconeogénesis (OA): biosíntesis de glucosa implica la conversión de OA en malato o aspartato para transportarse al citosol. Biosíntesis de lípidos (citrato): síntesis de AG en citosol, requiere acetil-Co A que se produce en mitocondria, sale como citrato. ATP+citrato+CoA ADP+Pi+OA+Acetil CoA. Citrato liasa.")
79
Biosíntesis de amino ácidos (α-cetoglutarato y OA):
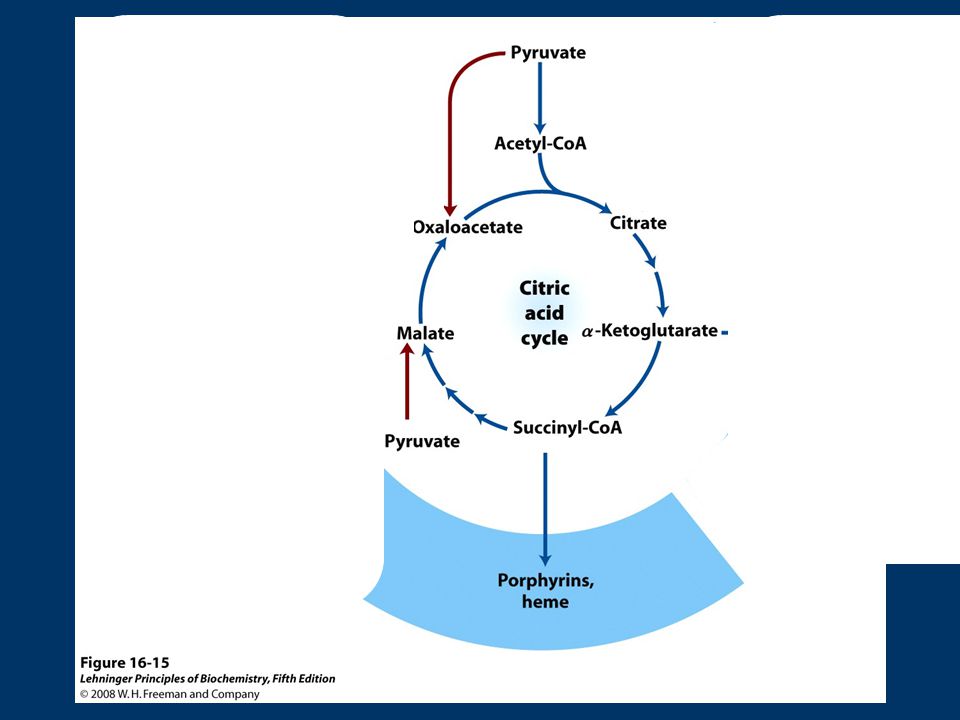
son iniciadores de la síntesis, desaminación de α-cetoglutarato+NADH+H + +NH Glu+NAD + +H2O Glu deshidrogenasa OA + Ala Asp. + Piruvato transaminación Biosíntesis de porfirinas (succinil-CoA): succinil-CoA es intermediario en síntesis del anillo de porfirina de los grupos hemo, importantes en transporte de O2 y electrones
: succinil-CoA es intermediario en síntesis del anillo de porfirina de los grupos hemo, importantes en transporte de O2 y electrones.")
81
Reacciones Anapleróticas
Los intermediarios que “salen” del ciclo de Krebs, se deben reponer!!!! Las reacciones que reponen intermediarios del ciclo se denominan anapleróticas Reacciones Anapleróticas Reacción Tejido/organismo Piruvato carboxilasa Hígado, Riñon Entrada Propionato Hígado (rumiantes) PEP Carboxiquinasa Corazón, Músc. esquelético Enzima Málico Todos,hepatocitos, adipocitos Reacción Tejido/organismo Piruvato carboxilasa Todos Hígado, Riñon Entrada Propionato Hígado (rumiantes) Enzima Málico Todos,hepatocitos, adipocitos
 PEP Carboxiquinasa. Corazón, Músc. esquelético. Enzima Málico. Todos,hepatocitos, adipocitos. Reacción. Tejido/organismo. Piruvato carboxilasa. Todos Hígado, Riñon. Entrada Propionato. Hígado (rumiantes) Enzima Málico. Todos,hepatocitos, adipocitos.")
83
4 subunidades idénticas
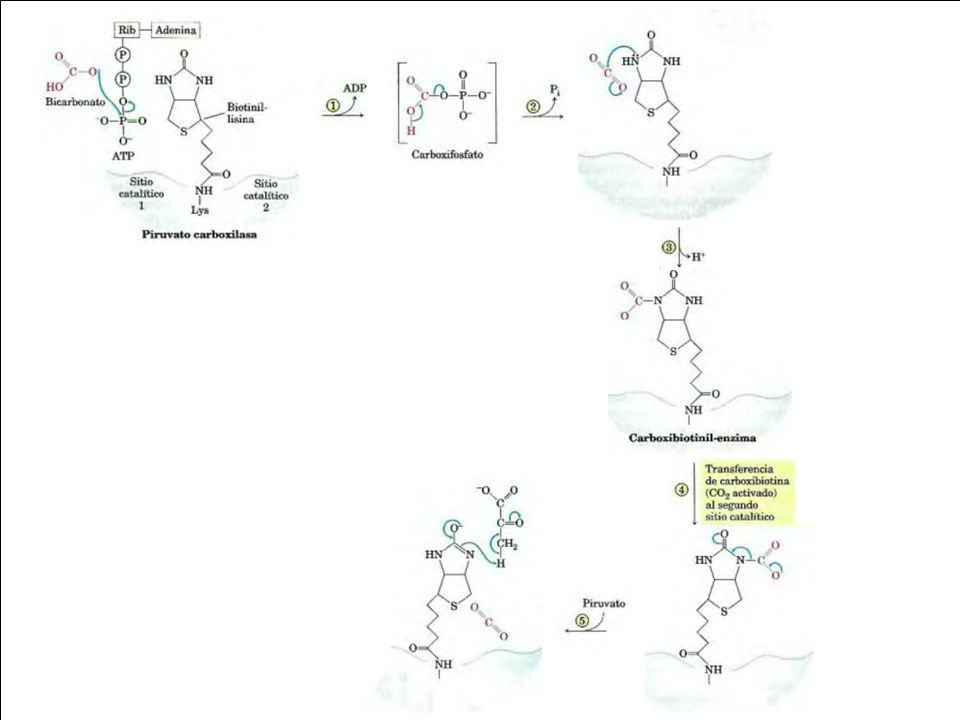
HCO3- ATP + ADP Pi 4 subunidades idénticas Cada una tiene Biotina (vit) como grupo prostético Biotina: transportador especializado de grupos monocarbonados en su forma más oxidada
 como grupo prostético. Biotina: transportador. especializado de grupos monocarbonados en su forma más oxidada.")
84
La PC tiene 4 subunidades idénticas, cada una tiene una molécula de biotina unida covalentemente a través de un enlace amida al grupo e-amino de un residuo específio de Lys en el sitio activo del enzima.

85
Bicarbonato es activado por ATP formando Carboxifosfato
Anclaje de Biotina a E biotina (grupo prostetico de la E, transporta CO2)
")
86
Los grupos carboxilo son activados en una reacción que rompe el ATP y une CO2 (del bicarbonato) a la biotina ligada al enzima La carboxilación del piruvato tiene lugar en dos pasos: 1) un grupo carboxilo proveniente del bicarbonato se une a la biotina
 un grupo carboxilo proveniente del bicarbonato se une a la biotina.")
89
Y 2) a continuación el grupo carboxilo se transfiere al piruvato para formar oxalacetato. Estos dos pasos tienen lugar en sitios activos diferentes. Este CO2 (activado) se transfiere entonces al aceptor (piruvato) en un a reacción de carboxilación.
 se transfiere entonces al aceptor (piruvato) en un a reacción de carboxilación.")
90
. Se formar oxalacetato.

93
Glucosa Neoglucogénesis Propionato PropionilCoA

94
Tiocianasa, o Acil-CoA sintetasa C.K. Propionil-CoA carboxilasa
Glucosa RUMEN Glucólisis Piruvato Lactato deshidrogenasa oxalacetato Lactato malato Propionato Lactil-CoA Acrilil-CoA fumarato Otros tejidos sangre Propionil-CoA succinato ATP Hígado Propionato Propionato 1 AMP CoASH Glucosa Propionil-CoA ATP sangre CO2 Vía malato 2 AMP Tiocianasa, o Acil-CoA sintetasa 1 C.K. D-Metilmalonil-CoA oxalacetato 2 Propionil-CoA carboxilasa 3 -Metilmalonil-CoA 3 Metilmalonil-CoA racemasa Reorganización 4 4 Metilmalonil-CoA mutasa Succinil-CoA

95
Enzima málico Piruvato HCO3- + NAD(P)H NAD(P)+ L-Malato Todos,hepatocitos, adipocitos
H NAD(P)+ L-Malato Todos,hepatocitos, adipocitos")
96
TAG AG

97
Ciclo de Krebs CO2 CH3 C SCoA O O COO- CH2 C O C O- OH C COO- Citrato
Oxalacetato CH2 H C COO- COO- H CH2 COO- COO- Isocitrato Ciclo de Krebs H CH COO- HO C ―H HO C COO- H HC ―H CO2 COO- L-Malato CH2 COO- COO- CH2 COO- C COO- CH COO- CH2 O -cetoglutarato CH CH2 CH2 COO- CH2 C SCoA Fumarato CO2 COO- O Succinil CoA Succinato

98
Hepatocito Si el Acetil CoA no es utilizado en el ciclo de Krebs
Acetoacetato Β-hidroxibutirato Acetona Exportados Fuente de energía: Corazón Músculo esquelético Riñón cerebro Alta [CC] en: Inanición Diabetes Oveja gestación múltiple Vaca posparto (gran demanda energética) Gotas de lípidos Formación de cuerpos cetónicos Ácidos grasos Acetil-CoA Oxalacetato C de Krebs Exportada: combustible para cerebro y otros tejidos Glucosa
 Gotas de lípidos. Formación de cuerpos cetónicos. Ácidos grasos. Acetil-CoA. Oxalacetato. C de Krebs. Exportada: combustible para cerebro y otros tejidos. Glucosa.")
100
G6Pasa F16Pasa Glicerol OXA Glucosa TAG AG PEPCK Piruvato PC
SUSTRATO: metabolito/s que inicia una vía metabólica. PRECURSOR: metabolito/s que por modificaciones químicas aportan los sustratos o intermediarios de una vía metabólica. G6Pasa Glucosa Producto Glucosa-6-P Fructuosa-6-P F16Pasa TAG Fructuosa -1,6-BP Glicerol DHAP Gliceraldehído-3-P AG 1,3-B-P Lactato Los precursores más importantes son comp De tres carbonos, lact, piruvato, Glicerol. Músculo Propionato 3-PG Lactato 2-PG Intermediarios del C. K. PEPCK PEP OXA Piruvato aminoácidos PC Proteínas aminoácidos

101
Piruvato Acetil CoA Piruvato Carboxilasa Aminoácidos Citrato Oxalacetato Aminoácidos -cetoglutarato Glutamato Aminoácidos Succinil CoA L-Malato Fumarato Propionato Aminoácidos

103
Los aminoácidos estándar entran al ciclo de Krebs para ser oxidados.
Isocitrato Fumarato Alanina Cisteína Glicina Serina Triptófano -Cetoglutarato Piruvato Acetil-CoA Oxaloacetato Asparagina Aspartato Tirosina Fenilalanina Succinil-CoA Metionina Valina Treonina Isoleucina Arginina Histidina Glutamina Prolina Citrato Glutamato Leucina Lisina Tirosina Fenilalanina Triptófano Isoleucina Acetoacetil-CoA Cuerpos cetónicos Glucosa Ciclo de Krebs CO2 CO2 El ayuno conduce al catabolismo de aminoácidos glucogénicos.

104
El piruvato para ser oxidado en el ciclo de Krebs.
Debe ser convertido primero a oxaloacetato. Es convertido primero a acetil-CoA por descarboxilación oxidativa. Requiere bajos niveles de NAD porque este compuesto va inhibir el proceso. Es realizado por una enzima que lo convierte a Acetil CoA Requiere solamente TPP como cofactor Requiere de un Complejo Multienzimatico ubicado en la matriz mitocondrial Debe ser transportado desde el citosol a la mitocondria Requiere el “Complejo de la Piruvato Deshidrogenasa” que tiene TPP, Lipoato, FAD, NAD y CoA como cofactores. Debe ser descarboxilado en una reacción que en condiciones fisiológicas es reversible

105
El ciclo de Krebs: produce acetil-CoA consume GTP produce ATP indirectamente es catalizado por un único complejo multienzimático se realiza por E mitocondriales y citosolicas produce 3 NADH por cada vuelta del ciclo Salen 2 CO2 por vuelta del ciclo la conversión de succinato a fumarato es la única reacción catalizada por una enzima unida a la MMI Produce un GTP por fosforilación oxidativa es inhibido por altas concentraciones de NADH y de ATP Solo tiene funciones catabólicas Es una vía Anfibólica Es estimulado por altas concentraciones de Acetil-CoA, ADP y OA

106
El ciclo de Krebs: produce intermediarios biosintéticos no requiere reposición de los intermediarios biosintéticos la conversión de piruvato en oxalacetato es catalizada por la piruvato deshidrogenasa la principal forma de reponer intermediarios es transaminación de aa La piruvato carboxilasa es estimulada por altas concentraciones de acetil-CoA la piruvato carboxilasa no requiere cofactores

107
Síntesis El ciclo de Krebs es el centro del metabolismo
Se realiza en la mitocondria (7 E matriz, 1 MMI). Oxida intermediarios para dar coenzimas reducidas necesarias para la síntesis de ATP. Por cada vuelta del ciclo se producen 3 NADH, 1 FADH2 y 1 GTP (ATP) por fosforilación a nível de sustrato. Por cada vuelta del ciclo salen 2 carbonos como CO2. 3 (de 8) reacciones son irreversibles, 3 puntos de control de flujo a través del ciclo.
. Oxida intermediarios para dar coenzimas reducidas necesarias para la síntesis de ATP. Por cada vuelta del ciclo se producen 3 NADH, 1 FADH2 y 1 GTP (ATP) por fosforilación a nível de sustrato. Por cada vuelta del ciclo salen 2 carbonos como CO2. 3 (de 8) reacciones son irreversibles, 3 puntos de control de flujo a través del ciclo.")
108
Síntesis El ciclo de Krebs es el centro del metabolismo
altas [acetil-CoA] y [ oxalacetato] [ADP], [Ca2+] son estimuladoras del ciclo Altas [NADH], [ATP], [citrato], [succinil-CoA] son inhibidoras del ciclo Intermediarios son utilizados en la biosíntesis de constituyentes vitales para la célula: vía anfibólica. Reacciones anapleróticas reponen intermediarios. Principal reacción anaplerótica: Piruvato carboxilasa.

Presentaciones similares











.>")









>")

