Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Glicobiologia El estudio de la estructura, química, bioquímica y función biológica de los carbohidratos complejos (glicanos).
.")
2
Glicobiología mas del 50% de todas las proteínas conocidas así como mas del 80% de las proteínas de membrana están glicosiladas. El contenido de carbohidrato de una glicoproteina varia entre el 1 y el 80% del peso de la misma. Los glicanos o glicoconjugados son el biopolímero mas abundante en la naturaleza. Mas del 1.5% de todo el genoma humano codifica enzimas involucradas en la biosíntesis y modificación de glicoconjugados, en ciertos protozoos este porcentaje se eleva a casi el 12%. La enzima Hidroxilasa del CMP-acido sialico es el único gene identificado que no se encuentra en humanos pero si en chimpance. La O-acetilglucosaminilacion es una modificación regulatoria mas abundante que la Fosforilación

3
Glicobiología Different sizes, chemical, physical and antigenic properties of glycans determine the variety of biological functions which are now slowly being unraveled. The overwhelming body of evidence indicates that the carbohydrates attached to glycoconjugates at the surface of cells play pivotal roles in the early stages of infection by pathogenic invaders, such as bacteria and viruses. They are important for defence and inflammation, for fertilization and development, for memory consolidation and rejection of xenografts. They explain many storage deseases. They have also been highlighted as structures of primary importance in metastasis and the progression of cancer thus providing a gauge of malignancy. In the more recent past, immunologists and physicians have begun to realise that the absence of the normal saccharide portions from recombinant therapeutics and adjuvants provokes an immune response from some patients during trials. Thus sugars, it emerged, were laying claim to important physiological and pharmaceutical properties, and could no longer be ignored. In 1988 the very appropriate neologisrn, `glycobiology,' was coined. (Rademacher, Parekh, Dweck).
.")
4
GLICOCONJUGADOS Proteoglicanos Glicoproteinas Glicolipidos
Glicosilfosfatidil-inositol anclajes Monosacarido Proteina Lipido Etanolamina fosfato Inositol fosfato

5
Glicobio(pato)logia El estudio de la estructura, química, bioquímica y función biológica de los carbohidratos complejos (glicanos) y su implicación causal en el desarrollo de patologías, diagnosis y terapia.
 y su implicación causal en el desarrollo de patologías, diagnosis y terapia.")
6
Hexasacarido Hexapeptido Nº Isomeros > 1012 64 106
Diferencias en la composición de isomeros entre oligopeptidos y oligosacaridos Hexasacarido Hexapeptido Nº Isomeros > (librería 6 hexopiranosas) (20 aminoácidos) Cantidad > 100 nmol < 100pmol Síntesis semanas h (robot) For example, the number of all possible linear and branched isomers of a pentasaccharide can be calculated to be greater than 2 x 106 as opposed to 120. The table presents parally the isomeric possibilities of comparable sequences of oligopeptide and oligosaccharide. Of course, chemists have been able to recreate glycosides in the laboratory since the outset of this century, but the inherent polyvalency of the constituent saccharides and ensuring that glycosides are preferentially formed in the desired anomeric configuration, makes laboratory preparation of synthetic oligosaccharides of adequate size and complexity for mimicking their natural counterparts a highly labour-intensive task, the task that has been the subject of considerable effort by carbohydrate chemists. Introduction of “antigen expression cloning” in 1989 has opened a new way of correct glycan synthesis. Antigen deficient cell line is transfected with exogenous DNA library and resultant transfectants screened for their ability to synthesyze appropriate cell surface antigens. Nevertheless, the synthesis of most types of glycosidic linkage with stereochemical precision is now fairly routine, although still far from trivial in the case of the larger oligosaccharides. Esto implica que no hay método analítico que permita un análisis estructural completo (determinación de secuencia) en cantidades de 100 nmol como se puede hacer con 100 pmol de proteínas u oligonucleotidos. No hay método analítico que permita distinguir entre 1012 estructuras con la misma masa.
 (20 aminoácidos) Cantidad > 100 nmol < 100pmol. Síntesis 20 semanas 3 h (robot) For example, the number of all possible linear and branched isomers of a pentasaccharide can be calculated to be greater than 2 x 106 as opposed to 120. The table presents parally the isomeric possibilities of comparable sequences of oligopeptide and oligosaccharide. Of course, chemists have been able to recreate glycosides in the laboratory since the outset of this century, but the inherent polyvalency of the constituent saccharides and ensuring that glycosides are preferentially formed in the desired anomeric configuration, makes laboratory preparation of synthetic oligosaccharides of adequate size and complexity for mimicking their natural counterparts a highly labour-intensive task, the task that has been the subject of considerable effort by carbohydrate chemists. Introduction of antigen expression cloning in 1989 has opened a new way of correct glycan synthesis. Antigen deficient cell line is transfected with exogenous DNA library and resultant transfectants screened for their ability to synthesyze appropriate cell surface antigens. Nevertheless, the synthesis of most types of glycosidic linkage with stereochemical precision is now fairly routine, although still far from trivial in the case of the larger oligosaccharides. Esto implica que no hay método analítico que permita un análisis estructural completo (determinación de secuencia) en cantidades de 100 nmol como se puede hacer con 100 pmol de proteínas u oligonucleotidos. No hay método analítico que permita distinguir entre 1012 estructuras con la misma masa.")
7
Origen de la Diversidad Estructural y Regulación de Afinidad
Composición de monosacáridos Al menos 7 monosacáridos distintos son comunes de todos los glicanos, aunque hay mas de 30 que pueden formar parte de glicanos en la naturaleza. NeuAc Gal Glc Man Fuc GalNAc GlcNAc Enlace glicosidico (secuencia) Entre el carbono aldehido/ceto y cualquier grupo hidroxilo del otro monosacarido Anomericidad Configuracion a o b del carbono que contiene el grupo aldehido/ceto Ramificación: numero y posición Tamaño del anillo: furanosa o piranosa “Cluster” con cadenas vecinas en la misma glicoproteina (mucinas) o en la superficie celular (“rafts”).
 Entre el carbono aldehido/ceto y cualquier grupo hidroxilo del otro monosacarido. Anomericidad. Configuracion a o b del carbono que contiene el grupo aldehido/ceto. Ramificación: numero y posición. Tamaño del anillo: furanosa o piranosa. Cluster con cadenas vecinas en. la misma glicoproteina (mucinas) o en la superficie celular ( rafts ).")
8
H O C 2 N 3 a H O C 2 a 1 O H C 2 3 6 O H C 2 3 H O C 2 b 1

9
Complejidad del Glicoma
GENOMA PROTEOMA DNA RNA Proteína Célula Organismo Enzimas Carbohidratos Glycoconjugados TRANSCRIPTOMA GLICOMA Unlike the template driven biosynthesis of nucleic acids and proteins, deprived of opportunities for molecular proofreading, biosynthesis of glyacans in glycoconjugates which relies on posttranslational biosynthetic machinary i.e. on a series of biosynthetic enzymes, termed glycozymes . Their activities depend on many highly variable factors. Cáncer Desarrollo Inflamación Diferenciación Activación Respuesta infecciones Dinámico Variaciones espaciales y temporales Respuesta al ambiente

10
Modulación del plegamiento
Desarrollo secuencial y cambios de importancia de las funciones de los glicanos Procariotes Eucariotes Unicelular Multicelular Estructural Plegamiento de proteínas I would like to end my talk by some evolutionary ideas, that emerged from the recent studies of glycoconjugates. The full discussion of the evolving roles of glycosylation is to be found in the K. Drickamer paper The natural philosophy of economizing with energy , molecular arsenal and space, with coded, compressed and zipped information, does not apply for building a glycoconjugate of 14 sugars just to trim it down to 5 or 8. Since this is what happens, we may look upon the function of glycoconjugates from an evolutionary point of view . Modulación del plegamiento “Tagging” Extracelular

11
Funciones Biológicas de los Glicanos
Físicas/Estructurales Reconocimiento Intrínseco = Propio Reconocimiento Extrínseco = No Propio INTRINSIC RECEPTOR EXTRINSIC RECEPTOR SELF P SELF Mimetismo molecular P = Patogeno OLIGOSACCHARIDE =

12
Los Glicanos actúan como señales vía Lectinas

13
ANIMAL AND PLANT GLYCOPROTEINS SHOW VARIOUS SUGAR CONTENTS
GLYCOPROTEIN Present in Mr % sugar content ENZYMES Alkaline phosphatase Murin liver Carboxypeptidase Y Yeast HORMONES AND CYTOKINES Chorionic gonadotropin Hum. urine Erythropoietin Hum. Urine Interferon g Hum. WBC LECTINS Soybean lectin Potato lectin The content of sugar varies from almost negligible to dominant as seen from th slide.

14
EXAMPLES OF GLYCOPROTEINS IN ANIMALS AND PLANTS
Cell-surface and viral coat glycoproteins Serum glycoproteins Structural glycoproteins

15
Oligosaccharides of the cell surface are attached to membrane embedded proteins and certain lipid
glycolipid hydrophobic helix integral protein phospholipid cholesterol The mistaken idea that biologically important molecules belong to mutually exclusive four categories (A. Kossel) were certainly no help in glycoconjugate research. While purifying proteins people tried to get rid of sugars, and muccopolysaccharides were found stubbornly contaminated with proteins. The contaminants were considered the consequenece of poor purification methods. 1958 revealed the first described covalent linkage between sugar and protein (H. Muir, Biochem. J.) The new era of the biology of carbohydrates started in the sixties. In 1968 the sugar chains of serum glycoproteins were found responsible for signalling the readiness for the removal of these molecules from the blood stream of higher animals. The discovery sparked a sudden interest amongst scientists. In retrospect this finding had major implications for the design of glycoprotein drugs. For some time we are aware that the great majority of secrtory and membrane bound proteins are glycosylated. So are many many mambrane bound lipids. Recent advances in glycobiology have denostrated that glycosylation is not limited to surface and extracellular proteins only, but is widespread in the nucleus and cytoplasm of the cell.
 were certainly no help in glycoconjugate research. While purifying proteins people tried to get rid of sugars, and muccopolysaccharides were found stubbornly contaminated with proteins. The contaminants were considered the consequenece of poor purification methods revealed the first described covalent linkage between sugar and protein (H. Muir, Biochem. J.) The new era of the biology of carbohydrates started in the sixties. In 1968 the sugar chains of serum glycoproteins were found responsible for signalling the readiness for the removal of these molecules from the blood stream of higher animals. The discovery sparked a sudden interest amongst scientists. In retrospect this finding had major implications for the design of glycoprotein drugs. For some time we are aware that the great majority of secrtory and membrane bound proteins are glycosylated. So are many many mambrane bound lipids. Recent advances in glycobiology have denostrated that glycosylation is not limited to surface and extracellular proteins only, but is widespread in the nucleus and cytoplasm of the cell.")
16
X-ray structure of murine antibody against canine lymphoma
Molecular model, based on the crystal structures of IgG Fab and IgG Fc. Fab glycosylation sites are in the hyper-variable region and occupied approx. 40% of the time (Dweck, 1995) X-ray structure of murine antibody against canine lymphoma Textbook presentation 1998
 X-ray structure of murine antibody against canine lymphoma. Textbook presentation")
17
New methods revolutionised the study of glycoconjugate structures
CD2 , surface glycoprotein involved in cell-adhesion, bears a single sugar attachment. CD2 mediates signal transduction through an interaction with CD58. If sugar is removed, protein unfolds and loses its activity. Substitution of Lys 61 with Glu makes structural effect of sugar redundant. Data, pointing to carbohydrates as moieties performing various, thus far unsuspected, important biological functions, started to pore from many laboratories. 3D structures revealed detailed insight of the structural impact of glycans on the glycosylated molecules. Sugar attachments were found ensure certain immediate environmental advantages on the glycoconjugate molecules such as favourable solvation properties , greater stability and improved resistance to degradation. Many proteins bear a sugar attachment such as 13.6 kDa T Lymphocyte Adhesion Domain of Human CD2

18
Specific glycoprotein - lectin recognition defines many physiological processes
Successful fertilisation depends on binding of spermal lectins to glycoproteins of egg membrane Fertilisation Development and differentiation Activity regulation of hormones Immune response and inflammation Memory consolidation and many more Cel-cell recognition relies on the specific glycoprotein-lectin interaction capable to decode information carried in the presented oligosaccharide motive.

19
Glycoproteins are found responsible for
Xenografts rejection Cancer progression and metastasis Storage diseases Immune response towards recombinant therapeutics - and many more

20
STUDYING GLYCOCONJUGATES
In our research we approach glycocojugates being focussed either on the structure, biosynthesis or functions of oligosaccharide parts of the investigated molecules, but our ultimate goal is to understand the impact of the glycan on the structure and function of the integral complex molecule. Each of these approaches uses different and specific rechniques. We are studying changes of glycosylation related to diseases. We may percieve possible application as diagnostic markers or as intervening drugs. At the start let us get acquainted with the monosaccharide building blocks and more complex oligosaccharide structures that present them self in glycoconjugates . EVOLUTION DIAGNOSIS THERAPY PHARMACEUTICAL INDUSTRY ANALYTICAL TOOLS

21
ALDOSE: CH2(OH)(CHOH)n-2HCO KETOSE: CH2(OH)(CHOH)n-3COCH2OH
D-Glucose Cyclic hemiacetal forms Simple sugars are either ketoses or aldoses predominatly in semiacetal or semiketal cyclic configuration. By cyclisation C1 has become a new chiral center, thus two anomeric forms exsist: a or b . Each form may adopt one of two conformotations. If two diastereomers differ only in the configuration about C-atom 1, they are called anomers, e.g. - and -glucose. Epimers are diastereomeric monosaccharides which have the opposite configuration of the hydroxyl group at only one position, e.g. D-glucose and D-mannose at C-atom 2, or D-galactose and D-glucose at C-atom 4. Hydrocarbons may contain additional structural elements. They may be reduced, aminated, oxydised, sulphated , phosphorylated, carboxylated or methylated. The stereochemistry and nomenclature of sugars will be discussed in details by Professor Kamerling. All functions of linked oligosaccharides are made possible by the characteristic chemical features of carbohydrates: 1.the existence of at least one and often two or more asymmetric centers, 2.the ability to exist either in linear or ring structures, 3.the capacity to form polymeric structures via glycosidic bonds, and 4.the potential to form multiple hydrogen bonds with water or other molecules in their environment

22
Glycosidic link Sugar1-OH + Sugar2-OH Sugar1-O- Sugar2 + H2O
16 kJ ENZYME Sugar1-OH + Sugar2-OH Sugar1-O- Sugar2 + H2O UDP GDP CMP GalNAc Fucose Sialic a. GlcNAc Mannose Galactose Glucose Glucuronic a. Xylose The presence of several nucleophilic hydroxyl groups and an electrophylic carbonyl centre renders sugars ideal molecules for polymerisation, in much the same way that amino acids are able to link together via peptide bonds to form proteins, and nucleotides through phosphodiester bonds to produce RNA and DNA. Coupling between two monosaccharides occurs by chemical condensation of the anomeric functional group of the glycosyl donor, on the one hand, and a hydroxyl group of the glycosyl acceptor on the other, resulting in the formation of a glycoside (Fig.). This reaction seals the anomeric fate of the glycosyl donor, since it has become a cyclic acetal that is resistant to ring-opening save by treatment with strong aqueous acid. However, such linkages do not occur spontaneously in Nature and require the participation of activated glycosyl donors, in the shape of nucleoside phosphate sugars, and of highly specialised enzymes in order to surmount the considerable kinetic and thermodynamic barriers to bond formation. Enzymes involved occur naturaly in minute concentrations and are extremely difficult to be isolated and analysed. One of the major goals of glycobiologists is to determine the crystal structure of glycosyltransferase. “Knocking out” genes leaves questions of regulation, and introduces new isoforms or targeting. The course offers a very priviladged opportunity to attend the lecture of J. Shaper, who pioneered the cloning of b-(1-4) galactsyltransferase. Enzyme catalyzed reaction seals the anomeric configuration.
. This reaction seals the anomeric fate of the glycosyl donor, since it has become a cyclic acetal that is resistant to ring-opening save by treatment with strong aqueous acid. However, such linkages do not occur spontaneously in Nature and require the participation of activated glycosyl donors, in the shape of nucleoside phosphate sugars, and of highly specialised enzymes in order to surmount the considerable kinetic and thermodynamic barriers to bond formation. Enzymes involved occur naturaly in minute concentrations and are extremely difficult to be isolated and analysed. One of the major goals of glycobiologists is to determine the crystal structure of glycosyltransferase. Knocking out genes leaves questions of regulation, and introduces new isoforms or targeting. The course offers a very priviladged opportunity to attend the lecture of J. Shaper, who pioneered the cloning of b-(1-4) galactsyltransferase. Enzyme catalyzed reaction seals. the anomeric configuration.")
23
Laine (1988) Pentasacarido lineal, monosacaridos no repetidos. n! x 2na x 2nr x 4n-1 n=numero de monosacaridos a=anomericidad r=tamaño del anillo 4n-1= enlaces Con cinco monosacáridos distintos el total es: estructuras posibles Con seis monosacáridos distintos el total es: Para Proteínas el total es 20n. Si n= Para oligonucleotidos el total es 4n . Si n=5 1024 Si incluimos posibles ramificaciones el total para un n=6 llega a: = 2.6 x109 sin considerar las formas D o L. El total de formas posible para un hexasacarido (el tamaño medio involucrado en reconocimiento) es de 1.05 x 1012. Esto implica que no hay metodo analitico que permita un análisis estructural completo (determinacion de secuencia) en cantidades de 100 nmol como se puede hacer con 100 pmol de proteinas u oligonucleotidos. No hay metodo analitico que permita distinguir entre 1012 estructuras con la misma masa.
 es de 1.05 x Esto implica que no hay metodo analitico que permita un análisis estructural completo (determinacion de secuencia) en cantidades de 100 nmol como se puede hacer con 100 pmol de proteinas u oligonucleotidos. No hay metodo analitico que permita distinguir entre 1012 estructuras con la misma masa.")
24
Comparable sequences of oligopeptide and oligosaccharide differ drammatically in isomeric
possibilities For example, the number of all possible linear and branched isomers of a pentasaccharide can be calculated to be greater than 2 x 106 as opposed to 120. The table presents parally the isomeric possibilities of comparable sequences of oligopeptide and oligosaccharide. Of course, chemists have been able to recreate glycosides in the laboratory since the outset of this century, but the inherent polyvalency of the constituent saccharides and ensuring that glycosides are preferentially formed in the desired anomeric configuration, makes laboratory preparation of synthetic oligosaccharides of adequate size and complexity for mimicking their natural counterparts a highly labour-intensive task, the task that has been the subject of considerable effort by carbohydrate chemists. Introduction of “antigen expression cloning” in 1989 has opened a new way of correct glycan synthesis. Antigen deficient cell line is transfected with exogenous DNA library and resultant transfectants screened for their ability to synthesyze appropriate cell surface antigens. Nevertheless, the synthesis of most types of glycosidic linkage with stereochemical precision is now fairly routine, although still far from trivial in the case of the larger oligosaccharides. R.R. Schmidt, Angew. Chem. 1986,

25
Diversidad de enlaces sacárido-proteína en las glicoproteínas

26
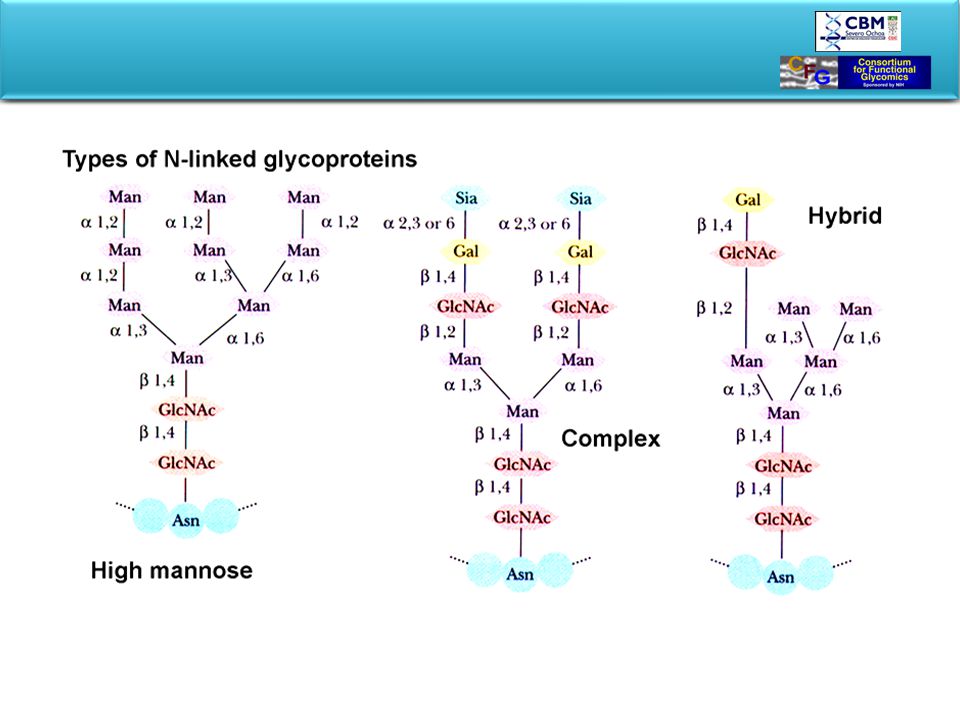
Oligosaccharides are linked to proteins by O- or N-glycosidic bonds
In N-linked oligosaccharides N-acetylglucosamine at the C1 position of reducing end of sugar binds to Asn within Asn-X-Ser/Thr sequence. O-glycosidic bonds are formed between C1 position of reducing terminus of monosaccharide or oligosaccharide and hydroxyamino acid, Ser or Thr. (N-acetylgalactosamine is often at the non-reducing end.)
")
28
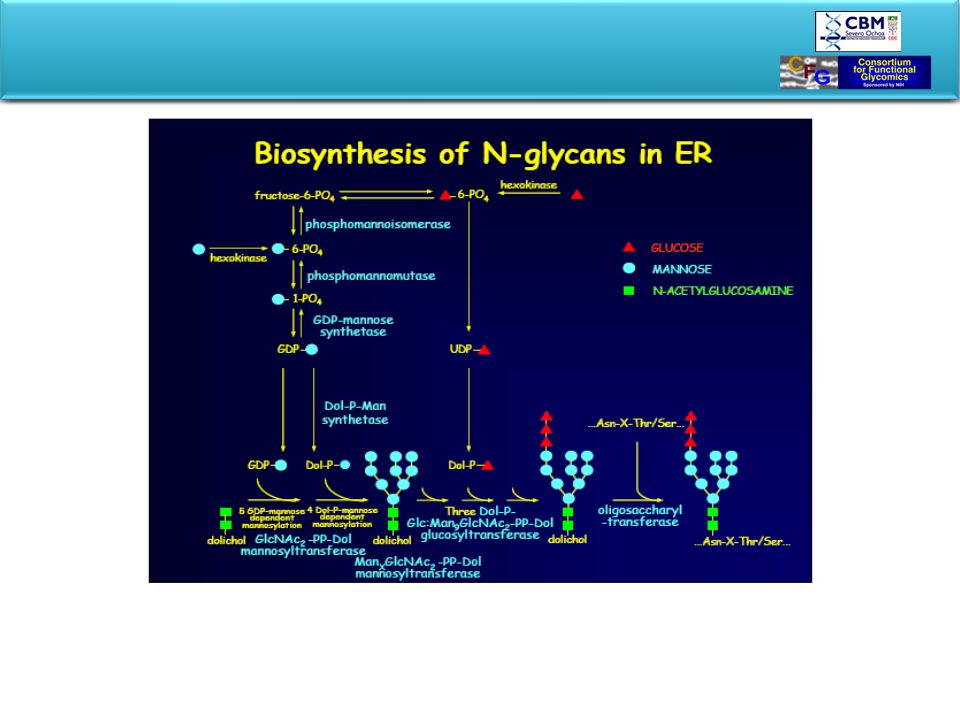
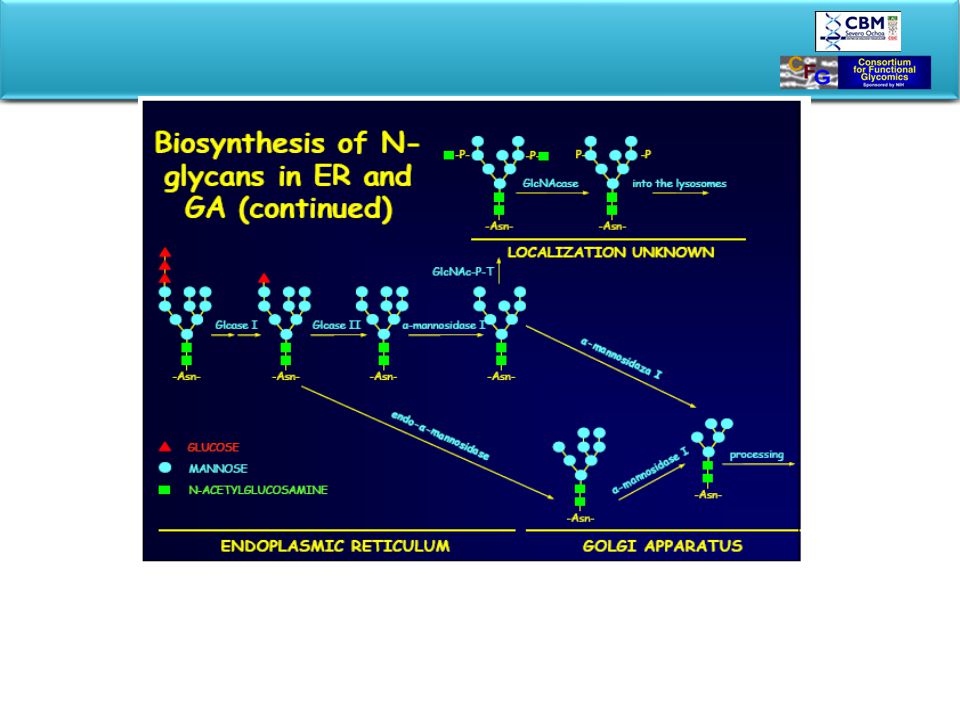
BIOSYNTHESIS OF GLYCOPROTEINS STARTS IN ER
In ER oligosaccharide is preassembled on a lipid carrier and transferred “en block” to the nascent protein. Glycan processing and tagging occurs in Golgi. Glycosylated product is packed and exported to final destination. Rough Er-ribosomes are in charge to produce secreted proteins, plasma membrane proteins, proteins of ER, of Golgy apparatus and lysosomes. Proteins penetrate through ER-mebrane into lumen. After passing ER they are ferried to the Golgy by transfer vesicle that bud of ER-membrane and eventually fuse with Golgi. Golgi packages the protein into secretory vesicles which fuse with the plasma membrane to release their content by exocytosis. The road to the perifery is also the road of posttranslational processing called glycosylation.

29
Biosintesis compleja pero conservada evolutivamente
…en principio no diana terapeutica EndoH sensitive EndoH resistant

33
Lectin-like chaperons mediate and control proper protein folding
Lectin-like chaperonnes, calnexin and soluble calreticulin mediate proper folding and control the quality of protein glycosylation. The mechanism is unader debate: Helenius, A Spiro include mannosidase B: Bergeron Mechanisms may differ according to substrate-protein. Calnexin and calreticulin bind misfolded or unfolded monoglucosylated glycoproteins to prevent final deglucosylation by -glucosidase II, which signalises the export to ER, in case of impaired protein folding.

34
Evolucion de complejidad
Levaduras Hongos Insectos Plantas Vertebrados

35
Oligosaccharide transfer
Origin of Glycoheterogeneity O-linked peptide Ser/Thr N-linked peptide A-X-S/T Oligosaccharide transfer Branching enzymes Processing enzymes Initial monosaccharide Terminal transferases Branching enzymes Glycoforms Terminal transferases

36
Glycosaminoglycans attached to proteoglycans are formed
from repeating disaccharide units A class of polysaccharides known as glycosaminoglycans is involved in a variety of extracellular (and sometimes intracellular) functions. Glycosaminoglycans are fundamental constituents of proteoglycans . Glycosaminoglycans consist of linear chains of repeating disaccharides in which one of the monosaccharide units is an amino sugar and one (or both) of the monosaccharide units contains at least one negatively charged sulfate or carboxylate group. The repeating disaccharide structures found commonly in glycosaminoglycans are shown. Heparin, with the highest net negative charge of the disaccharides shown, is a natural anticoagulant substance. It binds strongly to antithrombin III (a protein involved in terminating the clotting process) and inhibits blood clotting. Hyaluronate molecules may consist of as many as 25,000 disaccharide units, with molecular weights of' up to 107. Hyaluronates are important components of the vitreous humor in the eye and of synovial fluid, the lubricant fluid of joints in the body. The chondroitins and keratan sulfate are found in tendons, cartilage, and other connective tissue, whereas dermatan sulfate, as its name implies, is a component of the extracellular matrix of skin.
 functions. Glycosaminoglycans are fundamental constituents of proteoglycans . Glycosaminoglycans consist of linear chains of repeating disaccharides in which one of the monosaccharide units is an amino sugar and one (or both) of the monosaccharide units contains at least one negatively charged sulfate or carboxylate group. The repeating disaccharide structures found commonly in glycosaminoglycans are shown. Heparin, with the highest net negative charge of the disaccharides shown, is a natural anticoagulant substance. It binds strongly to antithrombin III (a protein involved in terminating the clotting process) and inhibits blood clotting. Hyaluronate molecules may consist of as many as 25,000 disaccharide units, with molecular weights of up to 107. Hyaluronates are important components of the vitreous humor in the eye and of synovial fluid, the lubricant fluid of joints in the body. The chondroitins and keratan sulfate are found in tendons, cartilage, and other connective tissue, whereas dermatan sulfate, as its name implies, is a component of the extracellular matrix of skin.")
37
Cartilage proteoglycan *Mr 220 952 Versican *Mr 264 048 Decorin
PROTEOGLYCAN STRUCTURES are predominantly glycosaminoglycans O-linked to Ser Secreted or extracellular Cartilage proteoglycan *Mr Versican *Mr Decorin * Mr Intracellular granule Serglycin *Mr Membrane intercalated Syndecan * Mr *Protein Mr Proteoglycans are a family of glycoproteins whose carbohydrate moieties are predominantly glycosaminoglycans. The structures of only a few proteoglycans are known, and even these few display considerable diversity They range in size from serglycin, having 104 amino acid residues (10.2 kD) to versican, having 2409 residues (265 kD). Each of these proteoglycans contains one or two types of covalently linked glycosaminoglycans. In the known proteoglycans, the glycosaminoglycan units are O-linked to serine residues of Ser-Gly dipeptide sequences. Serglycin is named for a unique central domain of 49 amino acids composed of' alternating serine and glycine residues. Decorin, a small proteoglycan secreted by fibroblasts and found in the extracellular matrix of connective tissues, contains only three Ser-Gly pairs, only one of which is normally glycosylated. The cartilage matrix proteoglycan contains 117 Ser-Gly pairs to which chondroitin sulfates attach. Proteoglycans may be soluble and located in the extracellular matrix, as is the case for serglycin, versican, and the cartilage matrix proteoglycan, or they may be integral transmembrane proteins, such as syndecan. Both types of proteoglycan appear to function by interacting with a variety of other molecules through their glycosaminoglycan components and through specific receptor domains in the polypeptide itself. In addition to glycosaminoglycan units, proteoglycans may also contain other N-linked and O-linked oligosaccharide groups.
 to versican, having 2409 residues (265 kD). Each of these proteoglycans contains one or two types of covalently linked glycosaminoglycans. In the known proteoglycans, the glycosaminoglycan units are O-linked to serine residues of Ser-Gly dipeptide sequences. Serglycin is named for a unique central domain of 49 amino acids composed of alternating serine and glycine residues. Decorin, a small proteoglycan secreted by fibroblasts and found in the extracellular matrix of connective tissues, contains only three Ser-Gly pairs, only one of which is normally glycosylated. The cartilage matrix proteoglycan contains 117 Ser-Gly pairs to which chondroitin sulfates attach. Proteoglycans may be soluble and located in the extracellular matrix, as is the case for serglycin, versican, and the cartilage matrix proteoglycan, or they may be integral transmembrane proteins, such as syndecan. Both types of proteoglycan appear to function by interacting with a variety of other molecules through their glycosaminoglycan components and through specific receptor domains in the polypeptide itself. In addition to glycosaminoglycan units, proteoglycans may also contain other N-linked and O-linked oligosaccharide groups.")
38
O-linked saccharides extend functional protein domains above membrane surface
There appear to be two structural motifs for membrane glycoproteins containing O- linked saccharides. Certain glycoproteins, such as leukosialin, are O-glycosylated throughout much or most of their extracellular domain Leukosialin, like mucin, adopts a highly extended conformation, allowing it to project great distances above the membrane surface, perhaps protecting the cell from unwanted interactions with macromolecules or other cells. The second structural motif is exemplified by the low density lipoprotein (LDL) receptor and by decay accelerating factor (DAF). These proteins contain a highly O-glycosylated stem region that separates the transmembrane domain from the globular, functional extracellular domain. The O- glycosylated stem serves to raise the functional domain of the protein far enough above the membrane surface to make it accessible to the extracellular macromolecules with which it interacts.
 receptor and by decay accelerating factor (DAF). These proteins contain a highly O-glycosylated stem region that separates the transmembrane domain from the globular, functional extracellular domain. The O- glycosylated stem serves to raise the functional domain of the protein far enough above the membrane surface to make it accessible to the extracellular macromolecules with which it interacts.")
39
PROTEOGLYCAN STRUCTURE OF BOVINE CARTILAGE
Cartilage matrix proteoglycan is responsible for the flexibility and resilience of cartilage tissue in the body. Long filaments of hyaluronic acid are studded or coated with proteoglycan molecules. The hyaluronate chains can be as long as 4 m and can coordinate 100 or more proteoglycan units. Cartilage proteoglycan possesses a hyaluronic acid binding domain on the NH2-terminal portion of the polypeptide, which binds to hyaluronate with the assistance of a link protein. The proteoglycanhyaluronate aggregates can have molecular weights of 2 million or more. The proteoglycan structures are enmeshed in a network of collagen fibers. The proteoglycan-hyaluronate aggregates are highly hydrated by virtue of strong interactions between water molecules and the polyanionic complex. Release of water by these structures is resposponsible for shock-absorbing properties of cartilage tissue. When cartilage is compressed (such as when joints absorb the impact of walking or running), water is briefly squeezed out of the cartilage tissue and then reabsorbed when the stress is diminished. This reversible hydration gives cartilage its flexible, qualities and cushions the joints during physical activities that might otherwise injure the involved tissues. Hyaluronate (up to 4 m), coated with proteo-glycans, forms the backbone of a complex polyanionic structure (Mr>2x106) which is highly hydrated and enmeshed in a network of collagen fibers. Link protein assists the noncovalent attachment of proteoglycan core proteins.
, water is briefly squeezed out of the cartilage tissue and then reabsorbed when the stress is diminished. This reversible hydration gives cartilage its flexible, qualities and cushions the joints during physical activities that might otherwise injure the involved tissues. Hyaluronate (up to 4 m), coated with proteo-glycans, forms the backbone of a complex polyanionic structure (Mr>2x106) which is highly hydrated and enmeshed in a network of collagen fibers. Link protein assists the noncovalent. attachment of proteoglycan core proteins.")
40
EVOLVING FUNCTIONS OF OLIGOSACCHARIDES
“Structural effects arose adventitiously and were selected for. Direct effects of glycosylation on behavior of glycoproteins evolved recently.” T. Drickhamer, 1998 ADVENTITIOUS General properties and structural role N-linked cores are added early in the life of glycoproteins. INTERMEDIATE Sorting role Transient glucose signals for sorting. Oligomeric proteins assemble before entering Golgi. COLLATERAL and RECENT Substitutional roles for amino acids Extensions and O-linked sugars Recognition markers Potential for adaptation Terminal elaborations Supporting facts for the Hypothesis of sequential evolution of glycosylation: Two strategies : “En bloc” vs.., stepwise ER-Golgi shuttling protein ERGIC-53, similar to L-type lectins, recognizes the same structures as calnexin/calreticulin Core sugar bound to a proximal Asn in the vicinity of a hydrophobic region of trypanosomes coating protein makes redundant the protective -helix segment. Various proteins coat the trypanosome parasites: in one variety, close to the hydrophobic base there is a short a-helical stretch which , in an other variety, is substituted by high-mannose N-glycoside attached to an Asn in the vicinity.. The philosophy behind is that if the sugar exists, the -helix can be omitted.This sugar has been attached by the preexisting glycosylation machinery. The selection process works on the glycosylated protein. If the oligosaccharide is present, -helix can be omitted.This sugar has been attached by the preexisting machinary and -helix may be omitted, but then it becomes an essential part of the structure.

45
Diversidad Evolutiva de las Lectinas

46
Oncogenic transformations are associated with numerous changes in glycosylation
different cancers are characterized by different changes appearance of novel carbohydrate structures changes in the activity of glycosyltransferases appearance of novel lectins novel carbohydrate structures and lectins are promising targets for cancer vaccines and drug targeting

47
NON-SPECIFIC ROLES OF GLYCANS BOUND TO PROTEIN
Folding, assembling Hepatic lipase and secretion Erythropoietin Vascular endothelial growth f immature GP aggregation HBV glycoproteins Conformational effects Yeast acid phospfatase increased thermal stability t-Plasminogen activator decreased backbone mobility Cell-surface CD Mucins (terminal Sia) Protective effects Fibronectin proteolytic degradation Ribonuclease B LDL-receptor (ovary cells) circulatory life time ceruloplasmin Functional effects and antithrombin binding Glycoprotein hormones kinetics Ribonuclesase A and B Proteins incorrectly folded or that fail to assemble into correct oligomeric complexes are not normally released from ER. E.g. T-cell receptor (TCR) composed of at least 7 subunits is never partially expressed as surface protein. (Helenius 1989). Proteolytic system of pre-Golgi degrades only unglycosylated proteins, yet sugars protect even incorrectly folded proteins from proteolytic degradation. General function of glycosylation is helping and controlling the folding, increase stability of the active conformation, provide the structure with the necessary rigidity, provide the active sites or adjust the activity of the active site as in case of ribonuclease B.
 Protective effects Fibronectin proteolytic degradation Ribonuclease B LDL-receptor (ovary cells) circulatory life time ceruloplasmin. Functional effects and antithrombin binding Glycoprotein hormones kinetics Ribonuclesase A and B. Proteins incorrectly folded or that fail to assemble into correct oligomeric complexes are not normally released from ER. E.g. T-cell receptor (TCR) composed of at least 7 subunits is never partially expressed as surface protein. (Helenius 1989). Proteolytic system of pre-Golgi degrades only unglycosylated proteins, yet sugars protect even incorrectly folded proteins from proteolytic degradation. General function of glycosylation is helping and controlling the folding, increase stability of the active conformation, provide the structure with the necessary rigidity, provide the active sites or adjust the activity of the active site as in case of ribonuclease B.")
48
Glicanos mas comunes en células animales
HIALURONANO GLICOSAMINO- GLICANOS HEPARAN SULFATO CONDROITIN SULFATO Glicanos mas comunes en células animales P S S S Ser-O- S S S S S -O-Ser NS NS Proteoglicano N- CHAINS O-LINKED CHAIN Ac ANCLAJE GLICOFOSFO- LIPIDO P Etn S P O N N Ser/Thr Asn NH Asn 2 Glicoproteina INOSITOL GLICOESFINGOLIPIDO Ac EXTERIOR P Acido Sialicos INTERIOR O-GlcNAc O Ser

49
HYALURONAN CHONDROITIN SULFATE Eliminacion de los glicanos mayoritarios permite la viabilidad celular in vitro GLYCOSAMINO- GLYCANS P S S S HEPARAN SULFATE Ser-O- S S S S S -O-Ser NS NS Proteoglycan N-LINKED CHAINS O-LINKED CHAIN Ac P GLYCOPHOSPHO- LIPID ANCHOR Etn S P O N N Ser/Thr Asn NH Asn 2 Glycoprotein INOSITOL GLYCOSPHINGOLIPID Ac OUTSIDE P Sialic Acids LETHAL INSIDE O-LINKED GlcNAc O Ser

50
Eliminación de los glicanos mayoritarios causa letalidad embrionaria in vivo
HYALURONAN CHONDROITIN SULFATE GLYCOSAMINO- GLYCANS P S S S HEPARAN SULFATE Ser-O- S S S S S -O-Ser NS NS Proteoglycan N-LINKED CHAINS O-LINKED CHAIN Ac P GLYCOPHOSPHO- LIPID ANCHOR Etn S P O N N Ser/Thr Asn NH Asn 2 Glycoprotein INOSITOL GLYCOSPHINGOLIPID Ac OUTSIDE P Sialic Acids INSIDE O-LINKED GlcNAc O Ser

52
Glicomica Comparativa: nueva aproximación para el establecimiento de las funciones endógenas de algunos oligosacaridos Species 1 Species 2 Species 3 Species 4 Species 5 = in situ localization of a specific oligosaccharide structure

53
Examples of Parasite Glycans

54
Examples of Parasite Glycans

55
Which class of Glycan recognition is more common?
Structural Structural OR Extrinsic Recognition Extrinsic Recognition Intrinsic Recognition Intrinsic Recognition The two classes of Glycan recognition are under different types and rates of evolutionary selection pressures

56
Glycans have probably been involved in an Ongoing Arms Race during Evolution
EXTRINSIC RECEPTOR INTRINSIC RECEPTOR M SELF M = Micro-organism Pathogen Toxin Symbiont OLIGOSACCHARIDE = How to Evade Microbial Recognition without loosing Endogenous Function?

57
Como evadir el reconocimiento microbiano sin perder la función endógena
Cambiar enlace Anadir modificación Enmascarar Añadir ramificación Sustitución M SELF Ac

58
Glicomica Comparativa

59
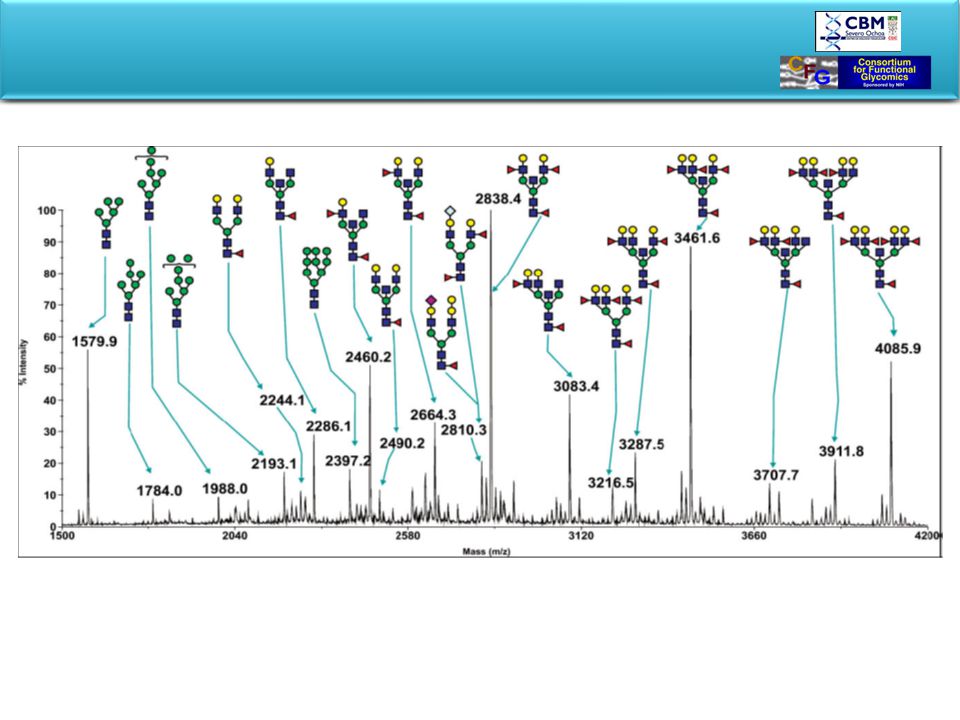
Glycan Arrays

60
Glycan Arrays Potencial de uso:
1.- Perfil inmunologico anti-glicanos para la identificacion del “target” en distintas condiciones patologicas (infecciones, cancer, autoinmunidad, xenotransplantes, etc). 2.- Identificacion de anticuerpos neutralizantes. 3.- Identificacion de nuevas familias de proteinas de union a carbohidratos tanto en el hospedador como en el agente infeccioso. Esto permitira identificar candidatos terapeuticos (Vacunas) y/o diagnosticos, asi como tambien ser critico para el entrendimiento de la relacion hospedador-agente infeccioso y la patogenesis d4e la inefccion. Tambien es posible establecer la firma glicano de los agentes ineecciosos. Es de destacar la capacidad de identificar (en combinacion con MALDI-TOF) antigenos no detctados por medios convencionales como WB.
. 2.- Identificacion de anticuerpos neutralizantes. 3.- Identificacion de nuevas familias de proteinas de union a carbohidratos tanto en el hospedador como en el agente infeccioso. Esto permitira identificar candidatos terapeuticos (Vacunas) y/o diagnosticos, asi como tambien ser critico para el entrendimiento de la relacion hospedador-agente infeccioso y la patogenesis d4e la inefccion. Tambien es posible establecer la firma glicano de los agentes ineecciosos. Es de destacar la capacidad de identificar (en combinacion con MALDI-TOF) antigenos no detctados por medios convencionales como WB.")
61
Glycan Arrays Chagas Serum Altamente reproducible en la identificacion de anticuerpos anti-glicanos en sueros

63
Concluding A great majority of proteins and many of membrane lipids are glycosylated. Many of their structures are to be revealed and their distinct function understood. Oligosaccharides serve various general and specific purposes, and may have regulatory properties. Information embedded in oligosaccharide structures, decoded by specific receptors, mediates many vital biological processes. Patterns of glycosylation depend on acting glycozymes. More information are needed about glycozymes. Diversity of glycotypes depends on tissue specific glycosylation machinery, yet various glycoforms are a challenging issue of biology. The role of glycosylation has been changing during evolution. Nonetheless, despite the diversity of important, more or less complex, structural motifs, the interlinking of a limited repertoire of simple monosaccharide building blocks leads to a very wide spectrum of biological oligosaccharides that will continue to present a formidable challenge to our understanding of the important processes in which they play a role. A great majority of proteins and many of membrane lipids are glycosylated. Many of their structures are to be revealed and their distinct function understood. Oligosaccharides serve various general and specific purposes, and may have regulatory properties. Information embedded in oligosaccharide structures and decoded by specific receptors mediates many vital biological processes. Patterns of glycosylation depend on acting glycozymes. We need more information about glycozymes. Diversity of glycotypes depends on tissue specific glycosyltion machinery, yet various glycoforms are a challenging issue of biology. The role of glycosylation has been changing during evolution. Microheterogeneity is common in glycoconjugates with elaborated glycans, while proteins involved in the synthesis of DNA and RNA bear unelaborated O-glycosylated attachments.

64
Concluding Impaired glycosylation may cause diseases and be disease specific. Surface oligosaccharides show antigenic properties and are receptor specific. Sugar structures change during development and aging, as well as in response to environmental factors. Understanding glycobiology will help understanding and treating diseases May the lectures on the challenging issues of glycobiology mediate your transformation into genuine glyco fan. Impaired glycosylation may cause diseases and be disease specific. Surface oligosaccharides show antigenic properties and are receptor specific. structures change during development and aging, as well as in response to environmental factors. Understanding glycobiology will have a huge impact on understanding and treating diseases The lectures that follow will bring you to the cutting edge of knowledge about the fascinating world of glycoconjugates and invite you to get involved in solving challenging issues of glycobiology

65
Quo vadis? Glicobiologia es un área en la que se requiere estudios sistemáticos descriptivos no mecanisticos (al menos por ahora). Esta claro que la exploración de la diversificación en estructuras de glicanos en patógenos y sus hospedadores puede educarnos acerca de su función. Beneficios: mejor conocimiento acerca de las susceptibilidades a infecciones y a un mejor diseño terapéutico, identificando glicanos con funciones criticas durante el desarrollo y diversas condiciones patológicas

Presentaciones similares