Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Deriva Génica Deriva, Ne, Efecto Fundador y Cuellos de Botella, Deriva+Selección

2
La deriva génica (random genetic drift) y el tamaño efectivo (Ne) de las poblaciones.
¿Qué es la deriva génica? El tamaño efectivo de las poblaciones, definiciones. Efecto de fundador y cuellos de botella. Deriva y Selección.

3
Deriva Génica: Azar en la evolución

4
Deriva Considerar una población en HW de la cual se toman individuos al azar para iniciar una colonia… Si hacemos esto n veces, la distribución de frecuencias alélicas de todas las colonias juntas estará representada por una distribución binomial de probabilidades.

6
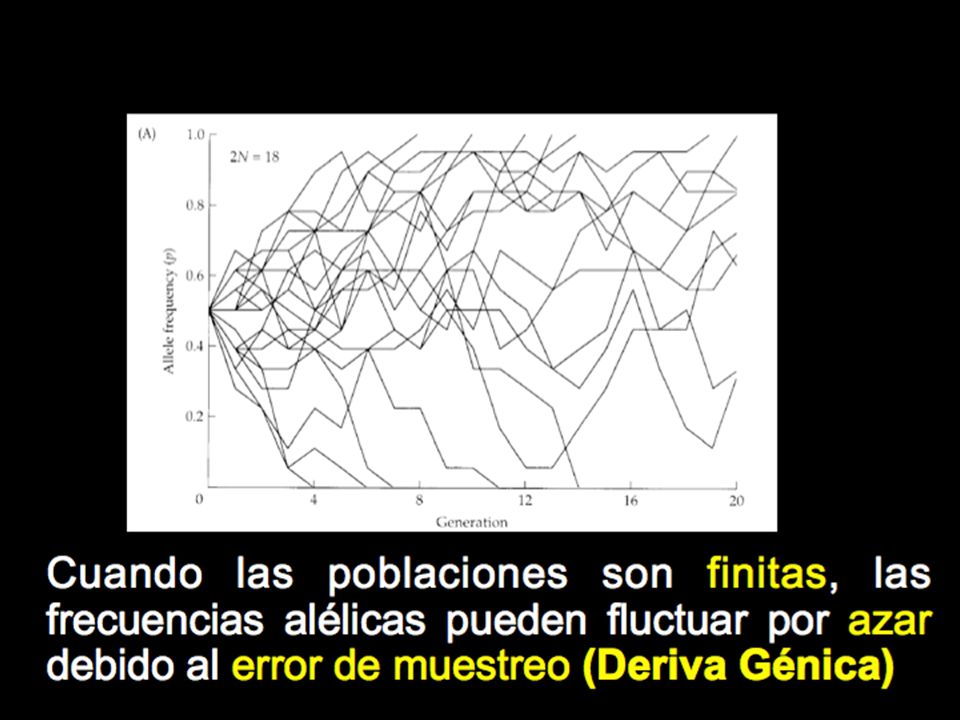
La representación gráfica de cómo varían las frecuencias alélicas en un modelo de deriva génica (Wright-Fisher)
")
7
Cada línea representa una colonia
de tamaño 2N

10
Cambios en las frecuencias alélicas
Deriva Génica: Azar en la evolución Cambios en las frecuencias alélicas fuerza dominante en poblaciones pequeñas y en el cambio molecular. Central en el estudio de la conservación! Su análisis es complicado y abstracto... Parámetro!: Tamaño efectivo de la población: Ne

11
depende del tamaño efectivo de la población, Ne
Deriva Génica: Dominante en poblaciones pequeñas (- de 100 individuos... pero...) depende del tamaño efectivo de la población, Ne Se pierde variación genética H Divergen las poblaciones (aumenta la Fst). En total (“n –replicas”) se mantiene la f. alélica q
 depende del tamaño efectivo de la población, Ne. Se pierde variación genética H. Divergen las poblaciones (aumenta la Fst). En total ( n –replicas ) se mantiene la f. alélica q.")
12
Ejemplo efectos Deriva Génica:
Tristan da Cunha: Isla en medio del Atlántico, a 2900 km de Sudáfrica: población pequeña, Deriva + Endogamia... 161 personas

13
Ejemplo efectos Deriva Génica:
Sólo genes de 7 fundadoras (mujeres), varias parientas entre ellas a) linaje materno, X, 5 linajes, mitocondria hm=0.768 b) 7 apellidos (linaje paternos, Y), 9 linajes, hy = 0.847
, varias parientas entre ellas. a) linaje materno, X, 5 linajes, mitocondria hm= b) 7 apellidos (linaje paternos, Y), 9 linajes, hy =")
14
mitocondria, linajes maternos:
5 y sólo 3 comunes, 1 raro...

15
9 linajes paternos, varios “colados”
un haplotipo Y, el cinco mucho más común...

25
Efectos de la Deriva Génica
Errores de muestreo... similar en todos los loci Unos dejan más hijos que otros por azar... mayor en poblaciones pequeñas

26
Efectos de la Deriva Génica
promedio

27
Efectos de la Deriva Génica N=9, fijación desde la generación 4 N= 50, no se ha fijado ninguna en 20 generaciones

28
2) Se va perdiendo variación (H) en el tiempo
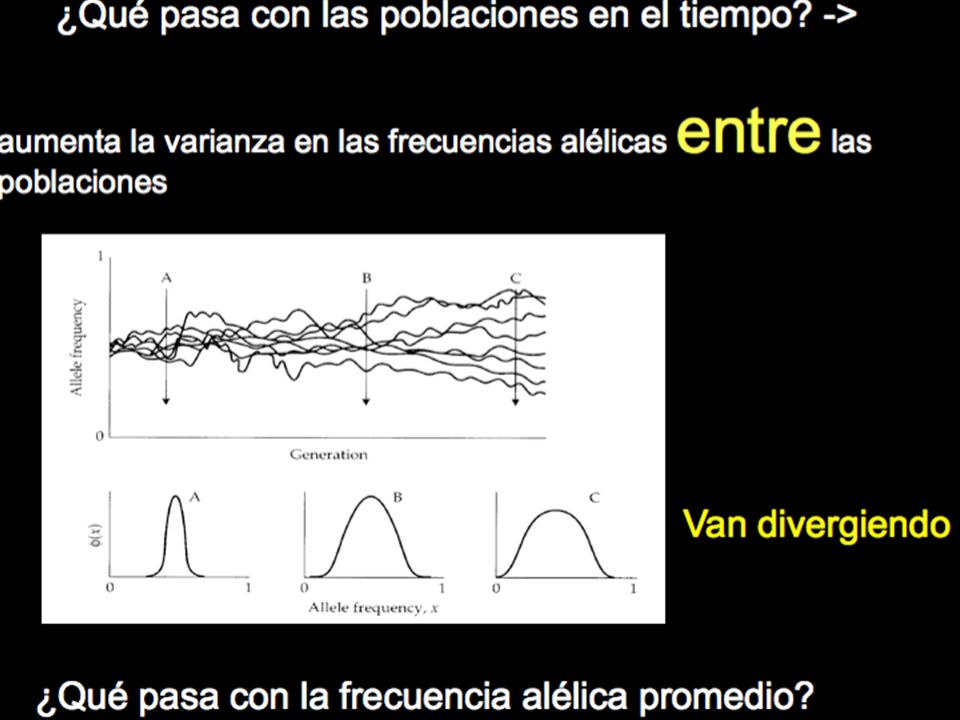
Entre más pequeño el N, más violentos (rápido y abruptos) son los cambios (si N infinito no hay cambio, H.-W.!) 2) Se va perdiendo variación (H) en el tiempo 3) Se pierde toda la variación (se fija un alelo) en dos poblaciones, un q=1 en t= 20, en otra q= 0 en t= 28 4) Van divergiendo las poblaciones en el tiempo, o sea aumenta la varianza en la f. alélicas entre las poblaciones! 5) La q se mantiene constante en promedio!
 son los cambios (si N infinito no hay cambio, H.-W.!) 2) Se va perdiendo variación (H) en el tiempo. 3) Se pierde toda la variación (se fija un alelo) en dos poblaciones, un q=1 en t= 20, en otra q= 0 en t= 28. 4) Van divergiendo las poblaciones en el tiempo, o sea aumenta la varianza en la f. alélicas entre las poblaciones! 5) La q se mantiene constante en promedio!")
29
4) Van divergiendo las poblaciones en el tiempo,
o sea aumenta la varianza en la f. alélicas entre las poblaciones!

30
N= 20 5) La q se mantiene constante en promedio!
 La q se mantiene constante en promedio!")
31
Probabilidad de fijación: igual que su f. alélica inicial! (u = 0.5)
")
32
pr. de fijación igual a la frec. alélica inicial:
más fácil de fijarte si ya es común el alelo

33
Experimento de Buri 1956 2 alelos neutros, bw75 y bw, locus
brown, color de ojos en D. melanogaster N = 16 por réplica. 107 replicas, p=q=0.5 iniciales 19 generaciones. No se muestran las poblaciones que se fijan al t= 19, 30 en bw y 28 en bw75.

34
Experimento de Buri 1956 tiempo

35
La media de q se mantiene
Experimento de Buri 1956 La media de q se mantiene La varianza aumenta según lo esperado (para una Ne de 9!)
")
36
Variación Genética: ¿como se pierde la H por Deriva?
La prob. de que un mismo alelo se tome dos veces es: 1/2N (o sea su freq.) La prob. De que dos alelos sean diferentes es 1-1/2N Aún siendo diferentes, pueden ser idénticos por descendencia (inbreeding coefficient) = ft
 La prob. De que dos alelos sean diferentes. es 1-1/2N. Aún siendo diferentes, pueden ser idénticos. por descendencia (inbreeding coefficient) = ft.")
37
la autocigosis anterior... incremento debido a que la pobl. es finita

38
rearreglando y sumando 1 en los dos lados como ya vimos sustituyendo:

39
Si Ne es infinito, Ht+1= Ht, no hay cambios, estamos en H.-W.
Si Ne es 1 (lo menos posible) la Ht+1 es la mitad de la Ht Este es el máximo cambio posible por Deriva Génica
 la Ht+1 es la mitad de la Ht. Este es el máximo cambio posible por Deriva Génica.")
40
Generalizando para t generaciones: que es aproxima- damente:

41
¿en cuanto tiempo se pierde un alelo? x= Ht / Ho : que tanto queda de la variación genética inicial

42
¿en cuanto tiempo se pierde un alelo? Para que se pierda la mitad de la var. genética, se necesita un tiempo del orden de magnitud del Ne (sólo es rápido si son apenas decenas de individuos)
")
43
Incremento en la Varianza q:

44
la divergencia entre poblaciones, la Varianza en q, depende de la var
la divergencia entre poblaciones, la Varianza en q, depende de la var. gen. inicial, de N y de t: mayor: entre más var. génica, más t y menor N

45
Incremento en la Varianza q:
entre mayor sea el Ne, menor es la varianza entre las poblaciones

46
Simulación de Deriva con
20 individuos, matriz 41 x 41

47
0.08 fija 0.08 fija casi “plana”

49
N=16 Ne= ca. 9 n= 20

50
¿En cuanto tiempo se fija un alelo por Deriva Génica?
Obviamente, debe de depender de la Ne y la frec. alélica inicial

51
se itera hasta que todas las poblaciones
se fijan... aburrido

52
T. de fijación, aproximación de Kimura y Ohta (1971)
el t de fijación función lineal de Ne y decrece si la p inicial es mayor si q= 0.2, t= 3.57 N, si q = 0.8, t= 1.61N; si q=.01, t=3.979

54
tiempo de fijación (coalescencia) promedio de
un nuevo alelo neutro, cuatro veces su Ne...

55
en realidad el t a la fijación
es una distribución de probabilidades tpromedio =1.81N tpromedio =3.57N q=0.2

56
Experimento de Buri tiempos de fijación observados y esperados, matriz 2N=18

57
El Tamaño efectivo de la población Ne:
El tamaño de una población ideal que experimenta los mismos efectos debidos a la DERIVA GENICA que la población real que estudiamos... generalmente se sugiere que debe de ser similar al número de adultos reproductivos, pero es más complejo...

58
Factores que lo complican: Variación en la relación de los sexos de individuos reproductivos (dioicos) El número de descendientes por individuos Número de individuos reproductivos en diferentes generaciones Tipo de reproducción Mezcla de individuos reproductivos, pre-reproductivos, post-reproductivos en un mismo censo poblacional
 El número de descendientes por individuos Número de individuos reproductivos en diferentes generaciones Tipo de reproducción Mezcla de individuos reproductivos, pre-reproductivos, post-reproductivos en un mismo censo poblacional")
59
Ne: el tamaño censal (real) se “convierte” al
de una población ideal donde todo los progenitores tiene las mismas probabilidad de reproducirse (dif. aleatorias)
")
60
Población ideal: 1) Diploide,
2) Hermafrodita (monoico), autofertilización 3) Varianza fecundidad al azar.
 Hermafrodita (monoico), autofertilización. 3) Varianza fecundidad al azar.")
61
Con estos supuestos, la distribución de número de gametos (progenie) por padre (k) se aproxima a una distribución de Poisson cuando la N es grande. El punto es estimar Ne o ¿cómo se desvía N del modelo ideal?

62
El tamaño efectivo de las poblaciones es el número de individuos en una población ideal que tienen la misma magnitud de deriva que la población real. Hay 3 aproximaciones para calcular estimar esta “magnitud”: Cambios en endogamia (inbreeding) = inbreeding effective size Cambios en la varianza en frecuencias alélicas = variance effective size Estimar la tasa de pérdida de heterocigosis = eigenvalue effective size.
 = inbreeding effective size. Cambios en la varianza en frecuencias alélicas = variance effective size. Estimar la tasa de pérdida de heterocigosis = eigenvalue effective size.")
63
Fecundidad al azar: dist. de Poisson
To estimate Ne How deviates from the ideal model? (Hartl and Clark, 1989): 1) Diploid organism 2) Sexual reproduction 3) Non-overlapping generations 4) Many independent subpopulation, each of constant size N 5) Random mating within each subpopulation 6) No migration between subpopulations 7) No selection Fecundidad al azar: dist. de Poisson
: 1) Diploid organism. 2) Sexual reproduction. 3) Non-overlapping generations. 4) Many independent subpopulation, each of constant size N. 5) Random mating within each subpopulation. 6) No migration between subpopulations. 7) No selection. Fecundidad al azar: dist. de. Poisson.")
66
si Nf=Nm Ne= Nf+Nm (N total) si un sexo domina Ne se hace mínimo
 si un sexo domina Ne se hace mínimo")
67
Cuando un sexo domina, la Ne es mínima

68
a mayores diferencia aleatorias en la
fecundidad, mayor deriva génica

69
La media y la varianza en el número de hijos decreció ,
al punto en el que Ne es mayor que N (censal)!!!
!!!")
70
¿como son Ne/N en la naturaleza?
entre 0.69 y 0.95 humanos... pero en bichos con mucha fecundidad, como moluscos, peces, árboles puede se MUY pequeño!

71
media armónica de Ne afectada por valores bajos de N

72
variación y cuellos de botella en poblaciones humanas (Egipto)
")
73
Efecto fundador y cuellos de botella
Una población puede descender de unos cuantos individuos ya sea porque: - La población fue iniciada por unos pocos individuos (efecto fundador) - Sólo un pequeño número de de individuos sobrevivieron en una generación en particular (cuello de botella)
 - Sólo un pequeño número de de individuos sobrevivieron en una generación en particular (cuello de botella)")
74
Resultados de reducciones dramáticas en Ne deriva
Cambios al azar de las variación genética Disminución de heterocigosis Pérdida de alelos

75
Efecto de fundador y cuellos de botella.
Veamos los efectos de esta reducción en el tamaño en la var. genética: despejando donde Ht and Ht+1 son las heterocigosis en la población original y en el grupo fundador

76
si hay n generaciones con una N pequeña,
Efecto de fundador y cuellos de botella. si hay n generaciones con una N pequeña, como cuando hay cuellos de botella donde la Hs es la heterocigosis antes del cuello de botella. si Ho = 0.7, Ht = 0.6, y t= 5, Ne = 16.4. ENTRE MAYOR o más larga sea LA REDUCCION EN LA H, MENOR Ne

78
Efecto de fundador y cuellos de botella.
Una población puede descender de pocos individuos por a) la población fue iniciada por pocos: efecto de fundador b) porque solo sobrevivieron unos pocos en cuellos de botella Esto genera cambios al azar en las frecuencia alelicas, disminuyendo la heterocigosis y dejando poco alelos!
 la población fue iniciada por pocos: efecto de fundador. b) porque solo sobrevivieron unos pocos en cuellos de botella. Esto genera cambios al azar en las frecuencia alelicas, disminuyendo la heterocigosis y dejando poco alelos!")
79
Efecto de fundador y cuellos de botella.
Veamos los efectos de esta reducción en el tamaño en la var. genética: despejando donde Ht and Ht+1 son las heterocigosis en la población original y en el grupo fundador

80
si hay n generaciones con una N pequeña,
Efecto de fundador y cuellos de botella. si hay n generaciones con una N pequeña, como cuando hay cuellos de botella donde la Hs es la heterocigosis antes del cuello de botella. si Ho = 0.7, Ht = 0.6, y t= 5, Ne = 16.4. ENTRE MAYOR o más larga sea LA REDUCCION EN LA H, MENOR Ne

81
BORREGO CIMARRON: El borrego cimarrón (Ovis canadensis) ha disminuído mucho en número de individuos y distribución en el siglo pasado, debido a enfermedad, cacería y otros factores. Como resultado, ha habido un número de introducciones en el oeste de Norte América, a manera de esfuerzos para establecer poblaciones más viables.
 ha disminuído mucho en número de individuos y distribución en el siglo pasado, debido a enfermedad, cacería y otros factores. Como resultado, ha habido un número de introducciones en el oeste de Norte América, a manera de esfuerzos para establecer poblaciones más viables.")
82
BORREGO CIMARRON: isla Tiburón
Por ejemplo, en 1975, For example, in early 1975, 20 borregos (4 machos y 16 hembras) fueron capturados en Sonora y trasladados a Isla Tiburón en el Mar de Cortés. La población traslocada creció rápidamente y en 1999, se estimaron 650 borregos en la isla, todos descendientes de el pequeño grupo fundador.
 fueron capturados en Sonora y trasladados a Isla Tiburón en el Mar de Cortés. La población traslocada creció rápidamente y en 1999, se estimaron 650 borregos en la isla, todos descendientes de el pequeño grupo fundador.")
83
El número de alelos (n) y la heterocigosis (H) en la población introducida de borrego cimarrón de la isla y aquéllos vaores para tres poblaciones de la misma especie usando diez loci de microsatélites y un locus MHC locus MENOS VAR. GENÉTICA

84
= 0.57/ (2(0.57-0.42)) Si comparamos la heterocigosis de los micros de
Arizona vs. la Isla Tiburón como Ht and Ht+t, respectivmente, nos da, Ne= 1.9, MUY BAJO!!! = 0.57/ (2( ))
)")
85
Los cuatro machos tenían edades de 1, 1, 2 y 7 años, lo que sugiere que el mayor de ellos pudo hacer una mayor contribución inicial que el resto. Se asume que el macho más viejo era el único macho reproductivo en la primera “cohorte” de machos, entonces el tamaño efectivo de la generación fundadora pudo ser, usando la expresión: Ne= 4(16)(1)/(16 + 1) = 3.8.
(1)/(16 + 1) = 3.8.")
86
Ne= 4(16)(1)/(16 + 1) = 3.8. Aunque esto no explica toda la pérdida de heterocisgosis, puede explicar la mayoría.

87
EJEMPLO Elefante marino (ELEPHANT SEAL):
Se suponía que el elefante marino del norte había sido cazado hasta la extinción hacia finales del siglo XIX cuando los “últimos” 153 individuos fueron matados por colectores en 1884. Pero aparentemente algunos animales sobrevivieron en una playa remota en Isla Guadalupe, Mexico, y sus descendientes fueron redescubiertos en 1892. Sin embargo, los ancestros de la población de hoy en día (200,000) pueden haber sido tan sólo 20 (Bonnell and Selander, I974).
 pueden haber sido tan sólo 20 (Bonnell and Selander, I974).")
88
EJEMPLO ELEPHANT SEAL:
Hoelzel et aI. (2002) encontró dos haplotipos de mtDNA con frecuencias estimadas de 0.27 and 0.73 en las poblaciones contemporáneas dando una diversidad haplotípica de H= 0.40. También determinaron los haplotipos pre-cuello de botella usando ejemplares de museos H= 0.80.
 encontró dos haplotipos de mtDNA con frecuencias estimadas de 0.27 and 0.73 en las poblaciones contemporáneas dando una diversidad haplotípica de H= También determinaron los haplotipos pre-cuello de botella usando ejemplares de museos H=")
89
Para determinar el tamaño del cuello de botella que
resultó en esta pérdidad de diversidad, se asume que la pérdida puede ser descrita con esta expresión: En donde la diversidad de mtDNA original es Ho=0.80, la diversidad observada contemporánea es H1=0.40 y Nef.i es el tamaño efectivo de hembras en la pob.

90
Usando la aproximación de Hedrick (1995b) y examinando varios tamaños y duración de cuellos de botella, pero permitiendo a la población crecer a los niveles del censo de 1922 y 1960, un cuello de botella de una generación con tamaño de 15 es consistente con la pérdidad de variación observada en mtDNA.
 y examinando varios tamaños y duración de cuellos de botella, pero permitiendo a la población crecer a los niveles del censo de 1922 y 1960, un cuello de botella de una generación con tamaño de 15 es consistente con la pérdidad de variación observada en mtDNA.")
91
PERDIDA DE ALELOS: En loci muy variables, se pierde más rápido los alelos que la heterocigosis... mejor medida de cambio Locus de microsatélites con 20 alelos en un cuello de botella de 5 individuos(2N= 10) pierde cuando menos la mitad de los alelos, pero sólo se espera que pierda el 10% de su heterocigosis
 pierde cuando menos la mitad de los alelos, pero sólo se espera que pierda el 10% de su heterocigosis.")
92
Selección en poblaciones finitas:
Deriva sola: Cuando no hay diferencia en selección en un locus, un alelo puede fijarse o perderse como resultado de de deriva. La probabilidad de fijación de un alelo es igual a la frecuencia inicial, por lo que cuando un alelo es raro su probabilidad de fijación es muy baja. Selección sola: En contraste, en una población infinita, que por definición no tiene deriva, un alelo favorable siempre incrementa y se avanza asintóticamente a la fijación

93
Selección + Deriva: En una población finita, un alelo favorable no siempre se fija, ya que puede perderse por efectos de azar (deriva). La probabilidad de fijación de un alelo favorable en una población finita u(p), es una función de la frecuencia inicial, la intensidad de la selección que lo favorece, y el tamaño de la población.
, es una función de la frecuencia inicial, la intensidad de la selección que lo favorece, y el tamaño de la población.")
94
Selección direccional,
Adecuaciones relativas de los genotipos A1A1,=1 +s, A1A2 = 1 + hs A2A2 =1, La ecuación genral de difusión (Kimura, 1962, Kimura y Otha, 1971) Se vuelve:
 Se vuelve:")
95
azar (deriva) p de fijación mucho más alta que
al azar si hay sel. dir. azar (deriva)
")
96
Cuando Ns<< l, s << 1/N, el cambio en las frecuencias alélicas es principalmente determindo por DERIVA: SI HAY POCA SELECIÓN Y/O Ne ES PEQUEÑO, LA EVOLUCION ES POR AZAR Cuando Ns>> 1, el cambio en las frecuencias alélicas es determinado por SELECCIÓN. SI LA SEL. ES INTENSA O Ne GRANDE, SOLO OPERA LA SELECCIÓN…

97
DERIVA SELECCION

98
Ne=20 sin sel. con sel s =0.1

99
En una población infinita, un alelo desfavorable siempre disminuye su frecuencia hasta perderse.
En contraste, en una población finita, un alelo desfavorable, particularmente si sus efectos no son graves, puede incrementar su frecuencia por azar y, potencialmente fijarse. Este efecto, en el que un alelo detrimental se comporta como neutral en una población pequeña, fue señalado por Wright (1931).
.")
100
Población finita: en una población finita, un alelo desfavorable, particularmente si sus efectos no son graves, puede incrementar su frecuencia por azar y, potencialmente fijarse. Ohta (1973) discutió este fenómeno en términos de evolución molecular y lo describió como el modelo CASI neutral.
 discutió este fenómeno en términos de evolución molecular y lo describió como el modelo CASI neutral.")
101
Ohta (1973) nearly neutral model.
Sugirió que el impacto relativo de la deriva génica y la selección varía con el tamaño poblacional, de tal modo que las variantes detrimentales pueden ser efectivamente neutras en poblaciones pequeñas, mientras que en poblaciones grandes son seleccionadas negativamente (en contra). POBLACIONES CHICAS, ACUMULAN VAR. GEN.DELETEREA!!!
. POBLACIONES CHICAS, ACUMULAN VAR. GEN.DELETEREA!!!")
102
ESPECIES AMENAZADAS: Algunas preocupaciones importantes para muchas especies amenazadas son que las poblaciones existentes son usualmente pequeñas, las especies han sufrido cuellos de botella en generaciones recientes o las poblaciones descienden de unos cuantos individuos. Todos estos factores pueden causar deriva génica importante con el potencial de pérdida de variacón genética “útil “ para cambios adaptativos futuros. Además, tamaños efectivos pequeños, pueden resultar en aumentos azarosos de alelos detrimentales porque los valores de Ns son muy bajos.

103
ESPECIES AMENAZADAS: Ejemplos: - La población de lobos escandinavos en cautiverio, inició con 4 fundadores con alta frecuencia de ceguera hereditaria (Laikre et al. 1993). - La población en cautiverio de cóndor de California, iniciada con 14 fundadores, con alta frecuencia de una forma letal de enanismo (Ralls et al.,2000). A pesar de que una estrategia de manejo que seleccione negativamente a los portadores puede reducir la frecuencia de estos alelos, también puede reducir la variación en otros genes!!!
. - La población en cautiverio de cóndor de California, iniciada con 14 fundadores, con alta frecuencia de una forma letal de enanismo (Ralls et al.,2000). A pesar de que una estrategia de manejo que seleccione negativamente a los portadores puede reducir la frecuencia de estos alelos, también puede reducir la variación en otros genes!!!")
104
Deriva Génica: ¡¡¡FIN!!!

105
mexicanum: pollinated by beetles An example:Astrocaryum
A tropical rain forest palm, Los Tuxtlas Veracruz, monoecious, pollinated by beetles Effective population size Ne

106
Astrocaryum mexicanum To estimate Ne
How deviates from the ideal model? (Hartl and Clark, 1989): 1) Diploid organism 2) Sexual reproduction 3) Non-overlapping generations * 4) Many independent subpopulation, each of constant size N 5) Random mating within each subpopulation * 6) No migration between subpopulations 7) No selection * = violated by A. mexicanum
: 1) Diploid organism. 2) Sexual reproduction. 3) Non-overlapping generations * 4) Many independent subpopulation, each of constant size N. 5) Random mating within each subpopulation * 6) No migration between subpopulations. 7) No selection. * = violated by A. mexicanum.")
107
Effective population We estimated its effective population size using direct and indirect methods. Direct methods: Analyzing how violates the ideal model: Non- random mating in the population: Neighborhood analysis Overlapping generations: Different methods

108
Neighborhood area (Wright 1946):
Direct methods: Neighborhood area (Wright 1946): Area that includes most (82 to 87%) of the parents of the individuals in the center of the area. Represents a more or less panmictic area, and an approximation of the effective population size would be: Neb = neighborhood area * effective density / m2
: Area that includes most (82 to 87%) of the parents of the individuals in the center of the area. Represents a more or less panmictic area, and an approximation of the effective population size would be: Neb = neighborhood area * effective density / m2.")
109
Neighborhood area = [AFp * 1/2Vp *t]+ [AFsp *Vsp * (1+t) /2] +[AFss *Vss * (1+t) /2]) p= pollen sp= seeds, primary dispersion ss= seed, secondary dispersion AF = Area correction factor for pollen, = 4 if normal V= Axial variance in dispersal distances t= outcrossing rate (0 to 1)
![Neighborhood area = [AFp * 1/2Vp *t]+ [AFsp *Vsp * (1+t) /2] +[AFss *Vss * (1+t) /2]) p= pollen.](http://slideplayer.es/slide/3516100/12/images/109/Neighborhood+area+%3D+%EF%81%B0%EF%80%A8+%5BAFp+%2A+1%2F2Vp+%2At%5D%2B+%5BAFsp+%2AVsp+%2A+%281%2Bt%29+%2F2%5D+%2B%5BAFss+%2AVss+%2A+%281%2Bt%29+%2F2%5D%29+p%3D+pollen..jpg "sp= seeds, primary dispersion. ss= seed, secondary dispersion. AF = Area correction factor for pollen, = 4 if normal. V= Axial variance in dispersal distances. t= outcrossing rate (0 to 1)")
110
Outcrossing rates and Pollen dispersal estimates
t= outcrossing rate Estimated using 5 loci, Ritland and Jain’s (1981) method, several sites (6) and 3 years: Is not different from 1. Pollen dispersal: Estimated using 3 methods: a) Dispersal of fluorescent dyes in two different years. b) Minimum distance between an active female and male inflorescence. c) Paternity analysis based upon progeny with a rare allele (LAP 3) and 4 other loci.
 method, several sites (6) and 3 years: Is not different from 1. Pollen dispersal: Estimated using 3 methods: a) Dispersal of fluorescent dyes in two different years. b) Minimum distance between an active female and male. inflorescence. c) Paternity analysis based upon progeny with a rare allele (LAP 3) and 4 other loci.")
111
Seed dispersal estimates:
Primary dispersal (seed fall under the mother): using isolated palms Secondary dispersal: attaching a nylon filament to evaluate dispersal by mammals
: using isolated palms. Secondary dispersal: attaching a nylon filament to evaluate dispersal by mammals.")
112
Total neigborhood = pollen dispersal + primary seed dispersal + secondary seed dispersal = 2551 m2 This has to be multiplied by the effective density. We used three methods to estimate it: Nei and Imaizumi (1966) Ne = Nr* L Crow and Kimura (1972) Ne = No *L *i Hill (1972) Ne =(4No-2)L/(Var(f) +2)
 Ne = Nr* L. Crow and Kimura (1972) Ne = No *L *i. Hill (1972) Ne =(4No-2)L/(Var(f) +2)")
113
Effective population size in Astrocaryum mexicanum, direct methods:
These methods gave the effective density by m2, and using it in the neigborhood: Ne = 234 (Nei and Imaizumi); Ne /N= 0.177 Ne = 560 (Hill); Ne /N=0.434 May be a gross subestimate, because, for instance, long distance gene dispersal is very difficult to estimate, but even if rare, it increases Ne dramatically.
; Ne /N= Ne = 560 (Hill); Ne /N= May be a gross subestimate, because, for instance, long distance gene dispersal is very difficult to estimate, but even if rare, it increases Ne dramatically.")
114
Other direct attempts to measure Ne in plants:
Using non-corrected neighborhood sizes (reviewed in Crawford, 1984; Eguiarte, 1990) Mean Neb herbs(20 studies) = 364 individuals (range ) Mean Neb trees (10 studies) = 292 individuals (range ) Ne = 560 (Hill); Ne /N=0.434 in A. mexicanum LARGE! Ne /N in Papaver dubium = 0.07 (Mackay, 1980; Crawford, 1994) [In humans range Ne /N 0.34 to 0.95, mean 0.60 ]
 Mean Neb herbs(20 studies) = 364 individuals (range ) Mean Neb trees (10 studies) = 292 individuals (range ) Ne = 560 (Hill); Ne /N=0.434 in A. mexicanum LARGE! Ne /N in Papaver dubium = 0.07 (Mackay, 1980; Crawford, 1994) [In humans range Ne /N 0.34 to 0.95, mean 0.60 ]")
115
Indirect methods to estimate Nm in A. mexicanum:
Fst: measure of genetic differentiation = (Ht - Hs)/ Ht Fst from 0 if there is no differences in allelic frequencies to 1 if the populations are fixed at different alleles. S. Wright found that Fst = 1/ 4 Ne m +1, and Crow and Aoki (1984) developed a more general model including mutation: Fst = (1/ (4 Ne ma +1)), where a= [n/n-1] 2 (if the number of populations is large, both estimates converge).
/ Ht. Fst from 0 if there is no differences in allelic frequencies to 1 if the populations are fixed at different alleles. S. Wright found that Fst = 1/ 4 Ne m +1, and Crow and Aoki (1984) developed a more general model including mutation: Fst = (1/ (4 Ne ma +1)), where a= [n/n-1] 2. (if the number of populations is large, both estimates converge).")
116
A. mexicanum, indirect analyses: Locus Fst Ne m m(Ne =560)
Mdh 6Pgd Pgi Adh Lap Low or little differentiation among sites Average Ne m 4.44: Ne m larger than 1. Las poblaciones se comportan como una gran población panmíctica...

117
Otra manera indirecta de estimar el Ne:
Infinite alleles model (Kimura) H= 4Ne/(4Ne+1) if (mutation rate) for allozymes is 10-6 in Zea spp. Doebly et al. (1984) reported a range H= Ne range 55,623-75,945 in 655 plant species, Hamrick et al. (1992) found a H=0.154 Ne = 45,508 higher than direct and ecological methods. Vecindades genéticas, entre Ne 1 a 4000!
 H= 4Ne/(4Ne+1) if (mutation rate) for allozymes is in Zea spp. Doebly et al. (1984) reported a range H= Ne range 55,623-75,945. in 655 plant species, Hamrick et al. (1992) found a H= Ne = 45,508. higher than direct and ecological methods. Vecindades genéticas, entre Ne 1 a 4000!")
118
Vecindades genéticas, entre Ne 1 a 4000!
H= 4Ne/(4Ne+1) Ne = 45, ,945 Vecindades genéticas, entre Ne 1 a 4000! The differences among estimates: may be due to the fact that the infinite alleles model may not describe their evolutionary process (Barbadilla et al. , 1996) estan midiendo cosas diferentes: vecindades el Ne local y ecológico, mientras que los alelos infinitos son el resultado de la evolución histórica de toda una especie, formada por n poblaciones ligadas por flujo génico...
 Ne = 45, ,945. Vecindades genéticas, entre Ne 1 a 4000! The differences among estimates: may be due to the fact that the infinite alleles model may not describe their evolutionary process (Barbadilla et al. , 1996) estan midiendo cosas diferentes: vecindades el Ne local y ecológico, mientras que los alelos infinitos son el resultado de la evolución histórica de toda una especie, formada por n poblaciones ligadas por flujo génico...")
119
ESPECIE poblaciones Vecindades el Ne local y ecológico.
Alelos infinitos son el resultado de la evolución histórica de toda una especie, formada por n poblaciones ligadas por flujo génico... ESPECIE poblaciones Aa AA aa

Presentaciones similares