Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Conceptos CLADOS Nuria Medina Medina

2
Genoma El genoma es todo el material genético contenido en las células de un organismo en particular. Por lo general, al hablar de genoma en los seres eucarióticos nos referimos sólo al ADN contenido en el núcleo, organizado en cromosomas.

3
Genoma Eucariótico Ser eucariota consiste básicamente en tener la mayor parte del genoma confinado en el núcleo.

4
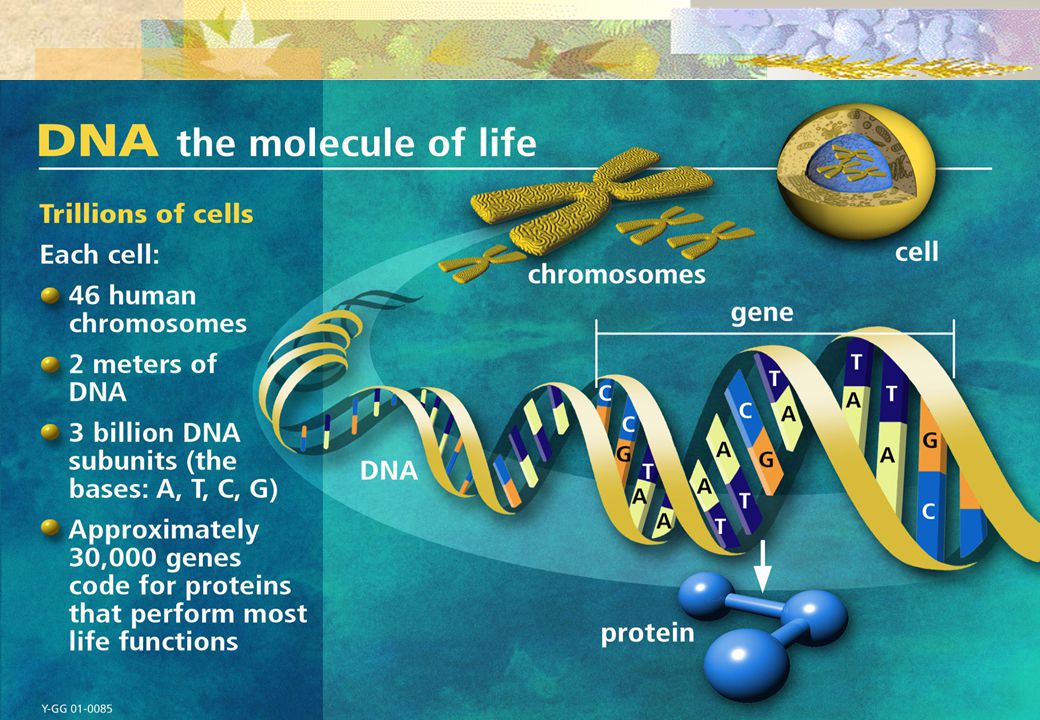
ADN El ácido desoxirribonucleico, frecuentemente abreviado como ADN (y también DNA, del inglés DeoxyriboNucleic Acid), es un tipo de ácido nucleico, una macromolécula que forma parte de todas las células. Contiene la información genética usada en el desarrollo y el funcionamiento de los organismos vivos conocidos, siendo el responsable de su transmisión hereditaria.
, es un tipo de ácido nucleico, una macromolécula que forma parte de todas las células. Contiene la información genética usada en el desarrollo y el funcionamiento de los organismos vivos conocidos, siendo el responsable de su transmisión hereditaria.")
5
ADN Desde el punto de vista químico, el ADN es un polímero de nucleótidos, es decir, un polinucleótido. Un polímero es un compuesto formado por muchas unidades simples conectadas entre sí, como si fuera un largo tren formado por vagones. En el ADN, cada vagón es un nucleótido, y cada nucleótido, a su vez, está formado por un azúcar (la desoxirribosa), una base nitrogenada (que puede ser adenina→A, timina→T, citosina→C o guanina→G) y un grupo fosfato que actúa como enganche de cada vagón con el siguiente. Lo que distingue a un vagón (nucleótido) de otro es, entonces, la base nitrogenada, y por ello la secuencia del ADN se especifica nombrando sólo la secuencia de sus bases.
, una base nitrogenada (que puede ser adenina→A, timina→T, citosina→C o guanina→G) y un grupo fosfato que actúa como enganche de cada vagón con el siguiente. Lo que distingue a un vagón (nucleótido) de otro es, entonces, la base nitrogenada, y por ello la secuencia del ADN se especifica nombrando sólo la secuencia de sus bases.")
6
ADN Cada molécula de ADN está constituida por dos cadenas formadas por un elevado número de compuestos químicos llamados nucleótidos. Estas cadenas forman una especie de escalera retorcida que se llama doble hélice. Cada nucleótido está formado por tres unidades: una molécula de azúcar llamada desoxirribosa, un grupo fosfato y uno de cuatro posibles compuestos nitrogenados llamados bases: adenina (abreviada como A), guanina (G), timina (T) y citosina (C).
, guanina (G), timina (T) y citosina (C).")
7
ADN Cada base orgánica de cada cadena se une a la otra base de la otra cadena mediante un enlace o puente de hidrógeno El apareamiento de bases se da entre A y T y entre G y C, unidas por puentes de hidrógeno (líneas punteadas) Existen dos puentes entre tiamina y adenina, y tres entre citosa y guanina.
 Existen dos puentes entre tiamina y adenina, y tres entre citosa y guanina.")
8
Cromosoma Dentro de las células, el ADN está organizado dentro de estructuras llamadas cromosomas que, durante el ciclo celular, se duplican antes de que la célula se divida. En biología, se denomina cromosoma (del griego χρώμα, - τος chroma, color y σώμα, -τος soma, cuerpo o elemento) a cada uno de los pequeños cuerpos en forma de bastoncillos en que se organiza la cromatina del núcleo celular durante las divisiones celulares (mitosis y meiosis).
 a cada uno de los pequeños cuerpos en forma de bastoncillos en que se organiza la cromatina del núcleo celular durante las divisiones celulares (mitosis y meiosis).")
9
Cromosoma La cromatina es un material microscópico que lleva la información genética de los organismos eucariotas y está constituida por ADN asociado a proteínas especiales llamadas histonas. Las histonas se unen al ADN, ayudan a dar su forma a los cromosomas y ayudan a controlar la actividad de los genes.

10
Ejemplo Podemos imaginar que nuestro genoma es como una gran biblioteca compuesta por 46 estanterías, organizadas en 23 pares, cada una de las cuales contiene muchos libros con información, como para –en sentido figurado– fabricar un ladrillo o una herramienta. Si imaginamos nuestro genoma como una gran biblioteca, decimos entonces que los pares de estantes son los cromosomas homólogos. Cada miembro de un par de cromosomas es similar, pero no idéntico, a su compañero. Los libros son los genes, en los cuales se guarda la información para fabricar una proteína (un ladrillo) o una enzima (una herramienta). El conjunto de genes de una especie determinada se llama genoma.
 o una enzima (una herramienta). El conjunto de genes de una especie determinada se llama genoma.")
11
ADN - Cromosoma En los organismos eucariontes, el ADN está organizado en cromosomas. Cada especie tiene un número característico: la cebolla tiene 16 (organizados en 8 pares), la mosca de la fruta Drosophila melanogaster, 8, y los seres humanos, 46. De esto no se desprende que una mayor cantidad de cromosomas equivale a ser “más inteligente” ya que las células que componen las papas tienen 48 cromosomas. Los seres humanos tenemos 23 pares de cromosomas: 22 de ellos se llaman cromosomas autonómicos y se heredan uno del padre y otro de la madre. Los cromosomas del par 23 se llaman cromosomas sexuales y son diferentes entre sí.
, la mosca de la fruta Drosophila melanogaster, 8, y los seres humanos, 46. De esto no se desprende que una mayor cantidad de cromosomas equivale a ser más inteligente ya que las células que componen las papas tienen 48 cromosomas. Los seres humanos tenemos 23 pares de cromosomas: 22 de ellos se llaman cromosomas autonómicos y se heredan uno del padre y otro de la madre. Los cromosomas del par 23 se llaman cromosomas sexuales y son diferentes entre sí.")
13
Genoma - Fenotipo El genotipo se refiere a la dotación génica del individuo, a su genoma, mientras que el fenotipo hace referencia a la manifestación del genotipo. Si el genotipo es el plan de instrucciones, el fenotipo constituye el resultado de la acción. Pero sólo el genotipo se hereda. El conjunto de genes de un individuo es el mismo desde su constitución en el zigoto, pero el fenotipo puede definirse a muy distintos niveles y es variable en el tiempo. El tipo celular, el tejido resultante del proceso de división, la morfología del organismo, su comportamiento…, son también fenotipos en la medida en que son expresión del genotipo. No existe una correspondencia biunívoca entre el genotipo y el fenotipo. Un mismo genotipo puede dar lugar a distintos fenotipos, y un mismo fenotipo puede ser consecuencia de distintos genotipos. Heterogeneidad: diferentes genotipos pueden producir fenotipos similares.

14
Gen Un gen es un segmento corto de ADN, que le dice al cuerpo cómo producir una proteína específica. Hay aproximadamente genes en cada célula del cuerpo humano y la combinación de todos los genes constituye el material hereditario para el cuerpo humano y sus funciones (genoma) Un gen es un subconjunto determinado de nucleótidos de uno de los lados de la escalera del cromosoma referenciado Un gen se encuentra en la misma posición en cada cromosoma
 Un gen es un subconjunto determinado de nucleótidos de uno de los lados de la escalera del cromosoma referenciado. Un gen se encuentra en la misma posición en cada cromosoma.")
15
Gen Los rasgos dominantes son controlados por un gen en el par
Los rasgos recesivos requieren que ambos genes en el par trabajen juntos para controlar el rasgo Muchas características personales, como la estatura, son determinadas por más de un gen. Sin embargo, algunas enfermedades, como la anemia drepanocítica, pueden ser ocasionadas por un cambio en un solo gen Si una enfermedad es autosómica dominante, quiere decir que la persona sólo necesita recibir el gen anormal de uno de los padres para heredar la enfermedad

16
Alelo Un alelo es cada una de las formas alternativas que puede tener un gen (se diferencian en su secuencia) y que se puede manifestar en modificaciones concretas de la función de ese gen Al ser la mayoría de los mamíferos diploides estos poseen dos alelos de cada gen, uno de ellos procedente del padre y el otro de la madre Por regla general se conocen varias formas alélicas de cada gen; el alelo más extendido de una población se denomina "alelo normal, salvaje o silvestre", mientras que los otros más escasos, se conocen como "alelos mutantes"
 y que se puede manifestar en modificaciones concretas de la función de ese gen. Al ser la mayoría de los mamíferos diploides estos poseen dos alelos de cada gen, uno de ellos procedente del padre y el otro de la madre. Por regla general se conocen varias formas alélicas de cada gen; el alelo más extendido de una población se denomina alelo normal, salvaje o silvestre , mientras que los otros más escasos, se conocen como alelos mutantes")
17
Ejemplo Alelo Pensemos que un gen es como como una página de un libro. En esta página existen las instrucciones completas para fabricar un producto. Imaginemos también que todos los individuos de una especie tienen esta página en su 'biblioteca genetica' (genoma). De un individuo a otro puede haber pequeñas diferencias en las letras que aparecen en estas páginas dadas. Pensemos, por ejemplo, que hay 99 letras en la página. Todas las letras pueden ser iguales de una página de un individuo a otro. Pero, puede haber una, o unas pocas letras, en la pagina que pueden ser diferentes de un individuo a otro. Esta variación se llama un alelo.
. De un individuo a otro puede haber pequeñas diferencias en las letras que aparecen en estas páginas dadas. Pensemos, por ejemplo, que hay 99 letras en la página. Todas las letras pueden ser iguales de una página de un individuo a otro. Pero, puede haber una, o unas pocas letras, en la pagina que pueden ser diferentes de un individuo a otro. Esta variación se llama un alelo.")
18
Ejemplo Alelo Organismos que tienen típicamente dos copias de la misma página en su biblioteca se llaman organismos diploides Cuando un individuo tiene dos copias idénticas de la misma página le llamamos organismo homogeneo Cuando un individuo tiene dos copias de la página ligeramente diferentes entonces le llamamos individuo heterogeneo

19
Dos alelos del mismo gen
Ejemplo Alelo Los conejos son diploides por lo que tienen dos copias de la misma página. Supongamos que la página del ejemplo es el gen que codifica el color del pelo, entonces sería diferente para los dos conejos. Dos alelos del mismo gen

20
Carotipo

21
Carotipo En la figura se presentan todos los cromosomas mitóticos de una niña, ordenados por parejas de homólogos y por su longitud, lo que se denomina cariotipo. Puede observarse que en ese cariotipo hay 46 cromosomas (o sea, 2n=46) que es el número cromosómico de la especie humana. Se puede advertir, también, que cada cromosoma tiene una estructura doble, con dos cromátidas hermanas que yacen paralelas entre sí y unidas por un único centrómero. Durante la mitosis las cromátidas hermanas, que son idénticas, se separan una de otra hacia dos nuevas células. Las parejas de cromosomas homólogos que se observan en la imagen tienen, además, una semejanza genética fundamental: presentan los mismos genes situados en los mismos lugares a lo largo del cromosoma (tales lugares se denominan locus o loci en plural). Esto indica que cada miembro del par de homólogos lleva información genética para las mismas características del organismo.
 que es el número cromosómico de la especie humana. Se puede advertir, también, que cada cromosoma tiene una estructura doble, con dos cromátidas hermanas que yacen paralelas entre sí y unidas por un único centrómero. Durante la mitosis las cromátidas hermanas, que son idénticas, se separan una de otra hacia dos nuevas células. Las parejas de cromosomas homólogos que se observan en la imagen tienen, además, una semejanza genética fundamental: presentan los mismos genes situados en los mismos lugares a lo largo del cromosoma (tales lugares se denominan locus o loci en plural). Esto indica que cada miembro del par de homólogos lleva información genética para las mismas características del organismo.")
22
Carotipo En organismos con reproducción sexual, uno de los miembros del par de cromosomas homólogos proviene de la madre (a través del óvulo) y el otro del padre (a través del espermatozoide). Por ello, y como consecuencia de la herencia biparental, cada organismo diploide tiene dos copias de cada uno de los genes, cada una ubicada en uno de los cromosomas homólogos Una excepción importante en el concepto de parejas de cromosomas homólogos es que en muchas especies los miembros de una pareja, los cromosomas que determinan el sexo o cromosomas sexuales, no tienen usualmente el mismo tamaño, igual situación del centrómero, la misma proporción entre los brazos o, incluso, los mismos loci. En la imagen puede observarse, por ejemplo, que el cromosoma Y (que determina el sexo masculino en humanos) es de menor tamaño y carece de la mayoría de los loci que se encuentran en el cromosoma X
 y el otro del padre (a través del espermatozoide). Por ello, y como consecuencia de la herencia biparental, cada organismo diploide tiene dos copias de cada uno de los genes, cada una ubicada en uno de los cromosomas homólogos. Una excepción importante en el concepto de parejas de cromosomas homólogos es que en muchas especies los miembros de una pareja, los cromosomas que determinan el sexo o cromosomas sexuales, no tienen usualmente el mismo tamaño, igual situación del centrómero, la misma proporción entre los brazos o, incluso, los mismos loci. En la imagen puede observarse, por ejemplo, que el cromosoma Y (que determina el sexo masculino en humanos) es de menor tamaño y carece de la mayoría de los loci que se encuentran en el cromosoma X.")
23
Locus (plural loci) En biología locus se utiliza para identificar posiciones de interés sobre determinadas secuencias Un locus (del latín locus, lugar; plural loci) es una posición fija sobre un cromosoma, como la posición de un gen La lista ordenada de locus conocidos para un genoma particular se denomina mapa genético
 es una posición fija sobre un cromosoma, como la posición de un gen. La lista ordenada de locus conocidos para un genoma particular se denomina mapa genético.")
24
Haplotiplo Un haplotipo (del griego haploos = simple) es una combinación de alelos ligados a múltiples locus que se transmiten juntos El haplotipo se puede referir a un solo locus, a varios o a un genoma completo. En el caso de organismos diploides como el hombre, un haplotipo del genoma comprende solo un miembro del par de alelos en cada locus Es decir, cada persona tiene un haplotipo materno y un haplotipo paterno

25
Meiosis La meiosis es un proceso de división celular por el que a partir de una célula madre diploide (2n) se obtienen cuatro células hijas haploides (n) Durante la meiosis se producen dos divisiones celulares consecutivas conocidas como meiosis I y meiosis II. La primera de las divisiones, que es más compleja que la segunda, es una división reduccional en la cual se pasa de una célula diploide (con 2n cromosomas) a dos células haploides (con n cromosomas) cada una de ellas con 2n cromátidas La segunda división es mucho más sencilla y similar a una división mitótica, y en ella a partir de las dos células haploides (n) anteriormente formadas se obtienen cuatro células haploides (n) con n cromátidas cada una de ellas
 se obtienen cuatro células hijas haploides (n) Durante la meiosis se producen dos divisiones celulares consecutivas conocidas como meiosis I y meiosis II. La primera de las divisiones, que es más compleja que la segunda, es una división reduccional en la cual se pasa de una célula diploide (con 2n cromosomas) a dos células haploides (con n cromosomas) cada una de ellas con 2n cromátidas. La segunda división es mucho más sencilla y similar a una división mitótica, y en ella a partir de las dos células haploides (n) anteriormente formadas se obtienen cuatro células haploides (n) con n cromátidas cada una de ellas.")
26
Meiosis Dos pares de cromosomas Cada cromosoma se duplica
Cromosomas homólogos se enfrentan Cromosomas homólogos se recombinan Se dividen en dos: cada nueva célula tiene un cromosoma de cada tipo Se dividen en dos: cada cromosoma se separa en dos y cada parte va a una célula distinta

27
Meiosis

28
Recombinación genética
La recombinación genética es un proceso que lleva a la obtención de un nuevo genotipo a través del intercambio de material genético entre secuencias homólogas de DNA de dos orígenes diferentes Las secuencias homólogas de DNA tienen la misma secuencia o casi la misma; por consiguiente, puede ocurrir apareamiento de bases en una longitud extensa de las dos moléculas de DNA Los cromosomas homólogos tienen los mismos genes ubicados en el mismo sitio. Sin embargo, los genes, aunque similares, pueden no ser necesariamente idénticos como ocurre cuando existe una mutación en un gen.

29
Recombinación genética
Para que aparezcan nuevos genotipos como consecuencia de la recombinación, es esencial que las dos secuencias homólogas sean genéticamente diferentes. Tal es el caso en una célula eucariótica diploide, que tiene dos juegos de cromosomas, uno procedente de cada padre El punto donde los cromosomas se cruzan se denomina kiasma y el proceso de intercambio se llama entrecruzamiento

30
Recombinación genética
Entrecruzamiento La recombinación puede producirse con baja probabilidad en cualquier lugar del cromosoma (normalmente 3), la frecuencia de recombinación entre dos puntos depende de sus distancia
, la frecuencia de recombinación entre dos puntos depende de sus distancia.")
31
Recombinación genética
Recombinación (crossing over): proceso complejo por el cual dos moléculas de ADN, una materna y otra paterna, intercambian segmentos recíprocamente. Ocurre en la profase meiótica. Proceso de intercambio de parte del material cromosómico entre cromosomas homólogos durante la meiosis, que produce nuevas combinaciones de la información genética Fracción de recombinación (è): para un par de loci dados, la proporción de meiosis en las cuales son separados por recombinación. Usualmente denotada como è. Los valores de è van de 0,0 a 0,5. A mayor distancia entre los loci, mayor fracción de recombinación.
: proceso complejo por el cual dos moléculas de ADN, una materna y otra paterna, intercambian segmentos recíprocamente. Ocurre en la profase meiótica. Proceso de intercambio de parte del material cromosómico entre cromosomas homólogos durante la meiosis, que produce nuevas combinaciones de la información genética. Fracción de recombinación (è): para un par de loci dados, la proporción de meiosis en las cuales son separados por recombinación. Usualmente denotada como è. Los valores de è van de 0,0 a 0,5. A mayor distancia entre los loci, mayor fracción de recombinación.")
32
Recombinación genética
Ligamiento

33
Resolución del Haplotipo
El genotipo de un organismo puede no definir únivocamente su haplotipo Por ejemplo, consideramos un organismo diploide y dos locus bi-alélicos en el mismo cromosoma. El primer locus tiene alelos A y T con tres posibles genotipos AA, AT y TT, y el segundo locus tiene G y C, dando de nuevo tres posibles genotipos GG, GC y CC Para un individuo dado, hay entonces nueve configuraciones posibles para el genotipo en estos dos locus

34
Resolución del Haplotipo
Con A y T tiene tres posibles genotipos AA – que padre y madre tengan A en el locus TT – que padre y madre tengan T en el locus AT – que el padre tenga A y la madre T en el locus o que el padre tenga T y la madre A en el locus

35
Resolución del Haplotipo
Genotipos para el locus 1 locus 1 locus 2 T G C genotipo ó Genotipos para el locus 2 locus 1 locus 2 T C G haplotipo En ambos casos salen dos haplotipos TG y TC

36
Haplotipo como herramienta
Como se indica anteriormente, un haplotipo es la constitución alélica de múltiples locus para un mismo cromosoma Dada la alta variabilidad alélica en el genoma humano, la probabilidad de que dos individuos no relacionados presenten un mismo haplotipo, es prácticamente nula Es por esto que el estudio de haplotipos se ha convertido en una herramienta útil en la determinación de relación génica entre individuos, y por lo tanto en el estudio del origen de mutaciones causantes de diversos padecimientos De este modo, podemos decir que una mutación dada, presente en dos individuos aparentemente no relacionados entre sí tiene un mismo origen, si al realizar un estudio de haplotipos, el haplotipo relacionado con la mutación en uno de los individuos es igual al haplotipo relacionado con la mutación en el otro individuo

37
SNP Un polimorfismo de un solo nucleótido o SNP (Single Nucleotide Polymorphism, pronunciado esnip) es una variación en la secuencia de ADN que afecta a una sola base (adenina (A), timina (T), citosina (C) o guanina (G)) de un a secuencia del genoma
 es una variación en la secuencia de ADN que afecta a una sola base (adenina (A), timina (T), citosina (C) o guanina (G)) de un a secuencia del genoma.")
38
Epidemiología Genética
La epidemiología genética es una disciplina relativamente reciente que estudia la interacción entre los factores genéticos y ambientales que dan origen a las enfermedades del ser humano. Valiéndose de marcadores genéticos desarrollados a través de la biología molecular, de complejos algoritmos almacenados en computadoras y de amplias bases de datos, la epidemiología genética se ha desarrollado notablemente durante los últimos años.

39
Epidemiología Genética
Estudios de Segregación Estudios en Familias Estudios de recurrencia familiar: Caso índice – caso control, Análisis de cohortes, Regresiones , Ecuaciones de estimación generalizables, Genética estadística Estudios en gemelos Estudios en parejas de hermanos afectados Estudios de ligamiento Estudios Poblacionales Estudios de asociación Estudios Combinados Test de haplotipos de riesgo (HRR) Test de distorsión de la trasmisión (TDT)
 Test de distorsión de la trasmisión (TDT)")
40
Análisis de Segregación
El Análisis de Segregación se basa en la estimación de la probabilidad de que los individuos de una población pertenezcan a un determinado genotipo a partir de unos datos observables. Para ellos se estudian los fenotipos de los individuos emparentados y se aplica la primera ley de Mendel (gen recesivo y gen dominante) para deducir si hay posible herencia genética.
 para deducir si hay posible herencia genética.")
41
Análisis del Ligamiento
Ligamiento genético puede definirse como la tendencia de genes u otras secuencias de ADN en loci específicos, a ser heredados juntos como consecuencia de su proximidad física en un solo cromosoma. Dos genes están ligados cuando no son transmitidos independientemente a la descendencia. Mientras más ligados estén dos loci, es más probable que sean heredados juntos; al medir el grado de recombinación entre ellos se puede averiguar la distancia que los separa.

42
Análisis del Ligamiento
Análisis de ligamiento - Linkage analysis (Análisis indirecto del ADN)
")
43
Análisis del Ligamiento

44
Análisis del Ligamiento
El estudio de ligamiento genético es un método indirecto que permite establecer la relación de una condición o enfermedad genética entre distintos miembros de una familia. Para establecer esta relación se utilizan marcadores genéticos localizados en la región cromosómica de interés, tratando de acotar el gen o la mutación asociada a la enfermedad o a la alteración investigada.

45
Análisis del Ligamiento
Marcador genético: Un segmento de ADN con una ubicación física identificable en un cromosoma y cuya herencia se puede rastrear. Un marcador puede ser un gen, o puede ser alguna sección del ADN sin función conocida. Dado que los segmentos del ADN que se encuentran contiguos en un cromosoma tienden a heredarse juntos, los marcadores se utilizan a menudo como formas indirectas de rastrear el patrón hereditario de un gen que todavía no ha sido identificado, pero cuya ubicación aproximada se conoce.

46
Análisis del Ligamiento
Desequilibrio de ligamiento (asociación alélica): cuando alelos en dos loci distintos están en los gametos más frecuentemente de lo esperado, según las frecuencias alélicas conocidas y la fracción de recombinación entre los dos loci.
: cuando alelos en dos loci distintos están en los gametos más frecuentemente de lo esperado, según las frecuencias alélicas conocidas y la fracción de recombinación entre los dos loci.")
47
Análisis del Ligamiento
Análisis paramétrico o basado en modelos: análisis de ligamiento que requiere un modelo genético predeterminado con variables que incluyen modo de herencia (autosómico dominante, recesivo, entre otros), penetrancia (completa e incompleta), frecuencias genotípicas, entre otras. Análisis no paramétrico: análisis de ligamiento que no requiere presuponer un modelo de herencia específico. Se usa para el análisis de enfermedades genéticamente complejas, que no presentan herencia mendeliana.
, penetrancia (completa e incompleta), frecuencias genotípicas, entre otras. Análisis no paramétrico: análisis de ligamiento que no requiere presuponer un modelo de herencia específico. Se usa para el análisis de enfermedades genéticamente complejas, que no presentan herencia mendeliana.")
48
Un ejemplo – Análisis de ligamiento
Las tres fases del análisis de ligamiento son las siguientes: Establecer los haplotipos de cada individuo: Se analizan los múltiples marcadores de ADN dispuestos a cada lado (marcadores flanqueantes) o dentro (marcadores intragénicos) de la región del gen de interés, con objeto de determinar el conjunto de marcadores (haplotipos) de cada miembro de la familia. Mediante la comparación de los haplotipos de los miembros de la familia con un estado genético conocido (es decir, afectados, no afectados), se puede identificar el haplotipo asociado con el alelo patológico. Determinar el estado genético: Una vez establecido el haplotipo asociado con la enfermedad, es posible determinar el estado genético de los miembros de una familia de riesgo.
 o dentro (marcadores intragénicos) de la región del gen de interés, con objeto de determinar el conjunto de marcadores (haplotipos) de cada miembro de la familia. Mediante la comparación de los haplotipos de los miembros de la familia con un estado genético conocido (es decir, afectados, no afectados), se puede identificar el haplotipo asociado con el alelo patológico. Determinar el estado genético: Una vez establecido el haplotipo asociado con la enfermedad, es posible determinar el estado genético de los miembros de una familia de riesgo.")
49
Un ejemplo – Análisis de ligamiento
Análisis de ligamiento para el diagnóstico prenatal de una enfermedad ligada al comosoma X.

50
Análisis del Ligamiento Paramétrico
El análisis de ligamiento paramétrico se basa en valorar el logaritmo del cociente probabilidad de ligamiento/probabilidad de no ligamiento, es decir, el logaritmo de la razón de probabilidades [logarithmic odds ratio (lod score)] El cálculo del lod score se hace a partir de toda una serie de parámetros genéticos de la enfermedad y del marcador, que hay que especificar. El problema de este método es que su fiabilidad depende de manera crítica de que los valores de los parámetros que se introducen en el cálculo sean correctos. Esta es una de las razones por las que este método tiene muy poca aplicabilidad en los «rasgos complejos» ya que en ellos raramente se conocen con certeza dichos parámetros
] El cálculo del lod score se hace a partir de toda una serie de parámetros genéticos de la enfermedad y del marcador, que hay que especificar. El problema de este método es que su fiabilidad depende de manera crítica de que los valores de los parámetros que se introducen en el cálculo sean correctos. Esta es una de las razones por las que este método tiene muy poca aplicabilidad en los «rasgos complejos» ya que en ellos raramente se conocen con certeza dichos parámetros.")
51
Análisis del Ligamiento Paramétrico
LOD score (del inglés, logarithm of odds): medida de la probabilidad de que dos o más genes ó loci estén 'ligados'. La interpretación general de los valores de LOD score es la siguiente: LOD score<-2, evidencia en contra de ligamiento; LOD score entre -2 y 3, ligamiento probable, se requieren datos adicionales; LOD score=3, evidencia de ligamiento significativo.
: medida de la probabilidad de que dos o más genes ó loci estén ligados . La interpretación general de los valores de LOD score es la siguiente: LOD score<-2, evidencia en contra de ligamiento; LOD score entre -2 y 3, ligamiento probable, se requieren datos adicionales; LOD score=3, evidencia de ligamiento significativo.")
52
Análisis del Ligamiento no Paramétrico
Los métodos estadísticos de ligamiento no paramétricos no requieren especificar los parámetros genéticos de la enfermedad ni del marcador genético que se esté estudiando. Estos métodos se basan en la demostración de que los parientes afectados por una enfermedad dentro de una familia heredan una misma copia del marcador genético con más frecuencia de la que sería esperable si la transmisión de la enfermedad y del marcador fueran independientes. Ello se traduce en el hecho de que los parientes afectados presentan los mismos alelos del marcador con más frecuencia de la que corresponde al caso de transmisiones independientes de la enfermedad y del marcador, o, dicho de otro modo, en que hay un exceso de alelos compartidos por dichos parientes. Por ello, los métodosno paramétricos de ligamiento se llaman también allele-sharing methods (métodos de alelos compartidos).
.")
53
Análisis del Ligamiento no Paramétrico
Métodos de Ligamieno no Paramétrico: IBD (identical by descent): valoran como alelos compartidos los alelos que proceden del mismo gen progenitor. Los parientes afectados que se analizan pueden ser hermanos (sib pair analysis)o parientes con otro grado de parentesco, aplicándose en cada caso distintas pruebas estadísticas para saber si son o no significativas las desviaciones del número de alelos compartidos por los parientes con respecto a lo esperado cuando la enfermedad y el marcador no están relacionados Cuando dos parientes comparten un alelo de un marcador, pero no se puede saber si ambos alelos son IBD, lo que es frecuente, se habla de una identidad alélica IBS(identical by state)
: valoran como alelos compartidos los alelos que proceden del mismo gen progenitor. Los parientes afectados que se analizan pueden ser hermanos (sib pair analysis)o parientes con otro grado de parentesco, aplicándose en cada caso distintas pruebas estadísticas para saber si son o no significativas las desviaciones del número de alelos compartidos por los parientes con respecto a lo esperado cuando la enfermedad y el marcador no están relacionados. Cuando dos parientes comparten un alelo de un marcador, pero no se puede saber si ambos alelos son IBD, lo que es frecuente, se habla de una identidad alélica IBS(identical by state)")
54
Análisis del Ligamiento no Paramétrico
Métodos de Ligamieno no Paramétrico: APM (affected pedigree member): Estos métodos utilizan pruebas estadísticas diseñadas específicamente para trabajar con el número de alelos IBS Para llevar a cabo cualquiera de estos métodos estadísticos es preciso conocer la frecuencia de los alelos del marcador genético correspondiente en la población que se esté analizando, siendo los métodos APM especialmente sensibles a dicho parámetro
: Estos métodos utilizan pruebas estadísticas diseñadas específicamente para trabajar con el número de alelos IBS. Para llevar a cabo cualquiera de estos métodos estadísticos es preciso conocer la frecuencia de los alelos del marcador genético correspondiente en la población que se esté analizando, siendo los métodos APM especialmente sensibles a dicho parámetro.")
55
Mapa Genético Un mapa genético nos permite analizar las regiones cromosómicas donde puedan ubicarse genes responsables de una determinada característica y en su caso identificarlos Se muestra el mapa genético para cada cromosoma del arroz (Chr 1 – 12) en el lado izquierdo y los segmentos contiguos PAC/BAC a la derecha. La posición de los PAC/BAC es mostrada en verde. La escala de los mapas estimada en centimorgans (cM). Tomado de Nature 436:
 en el lado izquierdo y los segmentos contiguos PAC/BAC a la derecha. La posición de los PAC/BAC es mostrada en verde. La escala de los mapas estimada en centimorgans (cM). Tomado de Nature 436:")
56
Mapa Genético La cartografía genética es una disciplina de la genética que, mediante varias técnicas, busca asignar a los distintos genes de un genoma su lugar físico en aquél. Existen dos variantes fundamentales de mapas: los genéticos, definidos mediante unidades de frecuencia de recombinación, y los físicos, en los que las distancias entre loci se expresan en unidades de distancia en nucleótidos. Técnicas: FISH RFLP Cartografía mediante deleciones

57
Mapa Genético El mapa genético es la representación de la secuencia lineal de la información genética en cada cromosoma. Los mapas de ligamiento representan el orden de disposición de los genes y las fracciones de recombinación entre pares de ellos, que son proporcionales a la distancia física que los separa. Linkage Mapping: Elston-stewart method Lander-Green method SuperLink Los mapas genéticos miden la cantidad de recombinación entre dos loci. Las unidades de mapa genético son cuantificadas en porcentaje de recombinación (è) o en centimorgan (cM) entre dos loci.
 o en centimorgan (cM) entre dos loci.")
58
Mapa Genético La Frecuencia de Recombinación es un parámetro genético de cartografía genética, que, para dos loci dados, se emplea como indicador cuantitativo de la distancia que existe entre ambos. Su unidad, la unidad de mapa genético o centiMorgan (cM), en honor de Thomas Hunt Morgan, como la frecuencia de recombinación de 0,01, esto es, del 1%, lo que quiere decir que uno de cada 100 productos de meiosis es recombinante. Centimorgan (cM): unidad usada en los mapas de ligamiento, correspondiente a una distancia de separación entre dos genes que poseen una probabilidad de recombinar del 1%. En humanos, 1 cM equivale aproximadamente a 1 millón de pares de bases (1 Mb). Se usa como medida para expresar la posición de un marcador a partir del telómero del brazo corto del cromosoma.
, en honor de Thomas Hunt Morgan, como la frecuencia de recombinación de 0,01, esto es, del 1%, lo que quiere decir que uno de cada 100 productos de meiosis es recombinante. Centimorgan (cM): unidad usada en los mapas de ligamiento, correspondiente a una distancia de separación entre dos genes que poseen una probabilidad de recombinar del 1%. En humanos, 1 cM equivale aproximadamente a 1 millón de pares de bases (1 Mb). Se usa como medida para expresar la posición de un marcador a partir del telómero del brazo corto del cromosoma.")
59
Programas para Análisis de Ligamiento
MapMaker: Paquete para análisis de ligamiento con pequeños pedigrís, medios hermanos, caracteres recesivos con familias nucleares y análisis multipunto para pedigrís de tamaño moderado Genehunter NPL: Paquete de análisis de ligamiento paramétrico y no paramétrico Sim IBD: Paquete de análisis de ligamiento paramétrico y no paramétrico ...

60
Modelos Gráficos para Análisis de Ligamiento
Árboles filogenéticos Redes de recombinación ancestral Redes bayesianas Redes de Markov Markov Chan MonteCarlo (MCMC): familia de métodos iterativos basados en simulación probabilística
: familia de métodos iterativos basados en simulación probabilística.")
61
Árbol Filogenético La filogenia es la historia de la evolución de un grupo de organismos La filogenia se puede representar gráficamente mediante árboles filogenéticos En la base del tronco estaría el antepasado común de todos los organismos, y de él partirían unas ramas, de las cuales saldrían ramas más finas, y de éstas ramitas, etc.

62
Árbol Filogenético Hay dos tipos de árboles filogenéticos:
Árboles de especies: Muestra la evolución de especies Árboles de genes: Muestra las variaciones de secuencias de ADN o proteínas En organismos que se reproducen asexualmente el árbol de genes y e de especies es esencialmente lo mismo, pero en organismos sexuales, el árbol de especies evoluciona más lentamente que el árbol de especies

63
Árbol Filogenético La forma convencional de un árbol filogenético es la siguiente:

64
Árbol Filogenético El nodo raíz indica la posición de la divergencia base, y en ciencias de la computación suele situarse arriba en lugar de abajo del árbol Las ramas que parten de los nodos interiores indican divergencias genéticas y pueden pintarse también con forma rectangular

65
Árbol Filogenético La rotación de las ramas no importa, de modo que un mismo árbol permite distintas representaciones

66
Árbol Filogenético Otro ejemplo de árboles equivalentes:

67
Árbol Filogenético La raíz del árbol es importante porque da la dirección

68
Árbol Filogenético Existen distintos tipos de árboles:
Dendograma: indica sólo relaciones de parecido Cladograma: indica relaciones de parecido y evolución. Simplemente muestra la distancia al antecesor común en términos relativos. La longitud de las ramas no indican el tiempo evolutivo Serían equivalentes, porque en un cladograma el eje Y no tiene significado

69
Árbol Filogenético Filograma: indica relaciones de parecido, evolución y distancia, para ello contiene información adicional dada por la longitud de las ramas. Los números asociados con cada rama corresponden a un atributo de las secuencias, tal como cantidad de cambio evolutivo. Es aditivo Dendrograma: tipo especial de árbol aditivo en el cual los extremos del árbol son equidistantes de la raíz y son proporcionales al tiempo de divergencia

70
Construir el Árbol Filogenético
Los pasos generales para construir el árbol filogenético son: Definir conjunto de secuencias a analizar (DNA, RNA o proteínas) provenientes de distintos microorganismos Alinear correctamente esas secuencias Aplicar métodos adecuados para la construcción de árboles filogenéticos Evaluar estadísticamente el árbol filogenético obtenido
 provenientes de distintos microorganismos. Alinear correctamente esas secuencias. Aplicar métodos adecuados para la construcción de árboles filogenéticos. Evaluar estadísticamente el árbol filogenético obtenido.")
71
Construir el Árbol Filogenético
Un posible ejemplo de conjunto de secuencias sería: 1 posición del genoma (no está ordenado puede ser padre-madre o madre-padre) Cada fila es un individuo Información sobre el individuo y su familia
 Cada fila es un individuo. Información sobre el individuo y su familia.")
72
Construir el Árbol Filogenético
Existen distintos métodos para construir árboles filogenéticos: Métodos de distancia: Usando matrices de distancia: UPGMA: Unweighted Pair Group Method with Arithmetic Mean. NJ-Neighbour Joining. Vecino más cercano Minimum evolution. Utiliza el método de cuadrados mínimos. Métodos discretos: Operan directamente con las secuencias Parsimonia: usa un carácter. Criterio: buscar el menor número de cambios evolutivos requeridos Máxima verosimilitud (Maximum likelihood): utiliza el estado del carácter y la distancia
: utiliza el estado del carácter y la distancia.")
73
Construir el Árbol Filogenético
Métodos discretos: Analizan cada columna dentro del alineamiento y construyen el mejor árbol que se ajusta a esa condición Hay que usar hipótesis para cada columna dentro del alineamiento Suelen ser lentos

74
Construir el Árbol Filogenético
Método parsimonia: La idea es que la hipótesis más sencilla es la más probable Entonces el objetivo es encontrar el árbol que minimice el número de cambios

75
Construir el Árbol Filogenético
Método parsimonia (ejemplo): 10
: 10.")
76
Construir el Árbol Filogenético
Método parsimonia (ejemplo): Usando sólo sitios informativos (al menos dos valores distintos, y cada valor representado al menos 2 veces en alguna secuencia
: Usando sólo sitios informativos (al menos dos valores distintos, y cada valor representado al menos 2 veces en alguna secuencia.")
77
Evaluar el Árbol Filogenético
El test más simple para probar si el conjunto de datos soportan el árbol obtenido es el del bootstrap Es un método estadístico que puede estimar las distribuciones por creación repetida y análisis de conjuntos de datos artificiales

78
Otros términos Epistasis: interacción entre los productos de dos genes, en la cual un gen interfiere la expresión fenotípica del otro. Expresión génica: efecto del gen sobre el fenotipo; forma como el gen de una enfermedad se expresa por sí mismo. Expresividad variable: extensión e intensidad variable de rasgos fenotípicos entre personas con un genotipo dado. Fenocopia: cambio fenotípico inducido por factores ambientales, que remeda los efectos fenotípicos de una mutación. Heterogeneidad genética: cuando un trastorno tiene tipos diferentes de patrones de herencia; también tiende a interpretarse como heterogeneidad de locus. Heterogeneidad de locus: cuando diferentes genes producen el mismo fenotipo clínico en diferentes familias. Heterogeneidad alélica: cuando diferentes alelos (o mutaciones) en el mismo locus están presentes y resultan en el mismo fenotipo.
 en el mismo locus están presentes y resultan en el mismo fenotipo.")
79
Otros términos Intrón: región no codificante del gen.
Exón: región codificante del gen. NPL score: non parametric linkage score, medida de Lod score obtenida mediante métodos no paramétricos. Nomenclatura de marcadores: por ejemplo, D4S450: D=ADN; 4=cromosoma 4; S=single (secuencia única); 450=# de referencia. Método del miembro afectado del pedigrí (APM): método de análisis de ligamiento no paramétrico que no requiere asumir un modo de herencia específico. Penetrancia: frecuencia con la cual un genotipo se manifiesta por sí mismo en un fenotipo dado. La penetrancia es descrita como 'completa' o 'incompleta', de acuerdo con la frecuencia con que se expresa el genotipo en el fenotipo. Pleiotropismo: cuando un gen produce múltiples efectos.
; 450=# de referencia. Método del miembro afectado del pedigrí (APM): método de análisis de ligamiento no paramétrico que no requiere asumir un modo de herencia específico. Penetrancia: frecuencia con la cual un genotipo se manifiesta por sí mismo en un fenotipo dado. La penetrancia es descrita como completa o incompleta , de acuerdo con la frecuencia con que se expresa el genotipo en el fenotipo. Pleiotropismo: cuando un gen produce múltiples efectos.")
Presentaciones similares