Descargar la presentación
La descarga está en progreso. Por favor, espere

1
12. Control de la expresión génica en eucariontes
Niveles: DNA transcripcional postranscripcional traduccional postraduccionales

2
12. Control de la expresión génica en eucariontes
contenido 12. Control de la expresión génica en eucariontes • niveles de control de la expresión génica • regulación transcripcional • regulación postranscripcional • regulación traduccional • regulación postraduccional • regulación a nivel de DNA • flujo de la información genética

3
niveles de control de la expresión génica

4
regulación transcripcional
RNA polimerasas cis: P mínimo, secuencias proximales, intensificadores y silenciadores trans: RNA polimerasas (I, II, III), TF, activadores y represores regulación hormonal
, TF, activadores y represores. regulación hormonal.")
5
regulación transcripcional
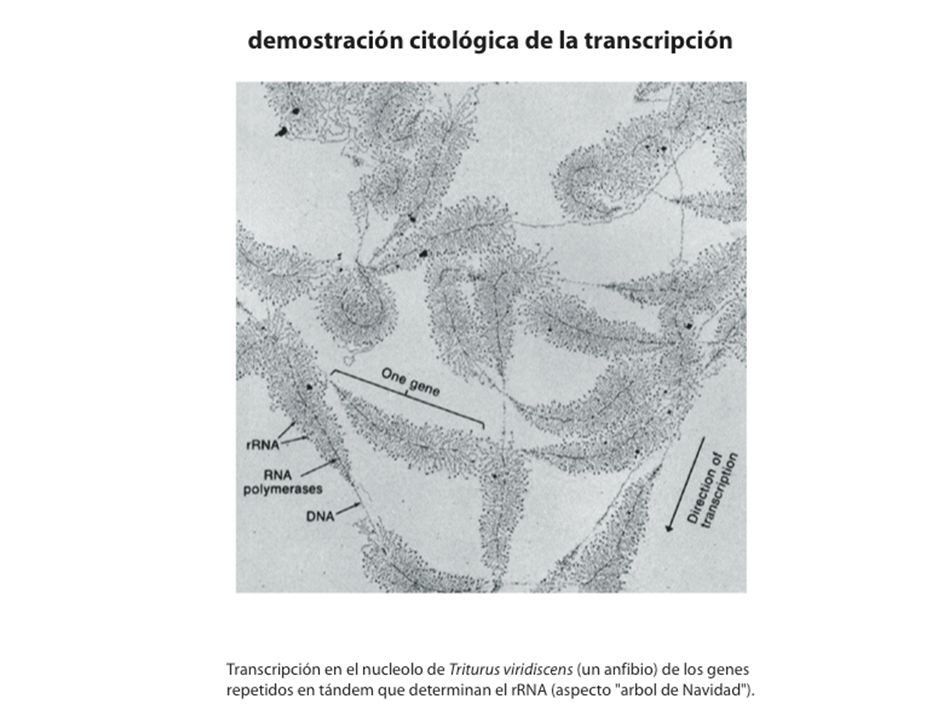
transcripción NO acoplada a traducción! mRNA monocistrónicos! (NO operones) RNA Polymerases in Eukaryotes There are three classes of eukaryotic RNA polymerases: I, II and III, each comprising two large subunits and smaller subunits. The two large subunits are homologous to the E. coli b and b' subunits. Two smaller subunits are similar to the E. coli a subunit. However, the eukaryotic RNA polymerase does not contain any subunit similar to the E. coli s factor. Therefore, in eukaryotes, transcriptional initiation should be mediated by other proteins. RNA polymerase II is involved in the transcription of all protein genes and most snRNA genes. It is undoubtedly the most important among the three classes of RNA polymerases. The other two classes transcribe only RNA genes. RNA polymerase I is located in the nucleolus, transcribing rRNA genes except 5S rRNA. RNA polymerase III is located outside the nucleolus, transcribing 5S rRNA, tRNA, U6 snRNA and some small RNA genes.
 RNA Polymerases in Eukaryotes. There are three classes of eukaryotic RNA polymerases: I, II and III, each comprising two large subunits and smaller subunits. The two large subunits are homologous to the E. coli b and b subunits. Two smaller subunits are similar to the E. coli a subunit. However, the eukaryotic RNA polymerase does not contain any subunit similar to the E. coli s factor. Therefore, in eukaryotes, transcriptional initiation should be mediated by other proteins. RNA polymerase II is involved in the transcription of all protein genes and most snRNA genes. It is undoubtedly the most important among the three classes of RNA polymerases. The other two classes transcribe only RNA genes. RNA polymerase I is located in the nucleolus, transcribing rRNA genes except 5S rRNA. RNA polymerase III is located outside the nucleolus, transcribing 5S rRNA, tRNA, U6 snRNA and some small RNA genes.")
6
promotor eucariótico (RNApol II)
secuencias proximales promotor mínimo Figure: 16-13 The Promoter The promoter at the 5’-end of eukaryotic genes consists of several modular elements, including the TATA box (-30) the CAAT box (-70) and the GC box (about -110). Eukaryotic promoters include the TATA box that lies ~30 bps upstream of the coding region and is typically the binding site recognized by RNA polymerase (RNAP). Promoters also include binding sites for various transcription factors that reside close to the ORF, and enhancer elements that are usually found quite a distance upstream or downstream from the coding region. Eukaryotic promoters reside immediately upstream (5' ~ bp) of the coding region of a gene, meaning they act in cis in relation to the open reading frame (ORF). Key sequences of the promoter structure comprise a variety of binding sites, where RNA polymerase (RNAP), transcription factors, and other regulators of transcription bind to DNA. Promoters function not only to bind RNA polymerase and transcription factors, but also specify the places and times that transcription can occur from that gene. In additon, different promoters have different combinations of factor binding sites depending on how those ORFs are transcriptonally regulated. In general, these sites can be moved around or even flipped into the opposite orientation and there will still be promoter activity. In other words, it seems that it is necessary to have some number of sites in the immediate upstream region, but exactly where they are within this region and what they are seems to be unimportant for activity.
 the CAAT box (-70) and the GC box (about -110). Eukaryotic promoters include the TATA box that lies ~30 bps upstream of the coding region and is typically the binding site recognized by RNA polymerase (RNAP). Promoters also include binding sites for various transcription factors that reside close to the ORF, and enhancer elements that are usually found quite a distance upstream or downstream from the coding region. Eukaryotic promoters reside immediately upstream (5 ~ bp) of the coding region of a gene, meaning they act in cis in relation to the open reading frame (ORF). Key sequences of the promoter structure comprise a variety of binding sites, where RNA polymerase (RNAP), transcription factors, and other regulators of transcription bind to DNA. Promoters function not only to bind RNA polymerase and transcription factors, but also specify the places and times that transcription can occur from that gene. In additon, different promoters have different combinations of factor binding sites depending on how those ORFs are transcriptonally regulated. In general, these sites can be moved around or even flipped into the opposite orientation and there will still be promoter activity. In other words, it seems that it is necessary to have some number of sites in the immediate upstream region, but exactly where they are within this region and what they are seems to be unimportant for activity.")
7
complejo inicio : TBP y factores de transcripción
Figure: 16-18 Assembly of the Transcription Complex in Eukaryotes Assembly of the transcription complex in eukaryotes. The initial step involves binding TBP to the TATA box, which bends the DNA. A combination of transcription factors (TFIIA, TFIIB, etc.) and RNA polymerase II are added in stepwise fashion prior to the initiation of transcription. Transcription Initiation: Transcription initiation requires the interaction of RNA polymerase with promoter DNA. In eukaryotic cells, there are three different types of RNA polymerases, each having particular functions and properties. RNA polymerase I is found in the nucleolar region of the nucleus and is responsible for transcribing the large ribosomal RNAs; RNA polymerase II transcribes messenger RNA precursors; and RNA polymerase III transcribes small RNAs such as transfer RNA, 5S ribosomal RNA, and other small DNA sequences. None of the eukaryotic RNA polymerases can bind efficiently to DNA. Rather, there are families of DNA-binding proteins that first bind to DNA and, once bound, interact with RNA polymerase to initiate mRNA synthesis. Initiation of Transcription by RNA Polymerase II and Associated Transcription Factors TFIIs and TATA Binding Protein (TBP). Generally, the initiation of transcription by RNA polymerase II occurs as follows: First, transcription factors (TBPs and TFIIs) and RNA Polymerase are sequencially assembled on to DNA at the promoter site. Other TFIIs arrive next and DNA is unwound to form an open complex. This process requires ATP hydrolysis and promoter clearance during which the transcription initiation complex moves beyond the promoter site through to the coding region. After promoter clearance, some of the initiation factors disassociate with the RNAPII complex. The remaining TFIIs and the RNAPII enzyme are now considered part of the transcription elongation complex.
 and RNA polymerase II are added in stepwise fashion prior to the initiation of transcription. Transcription Initiation: Transcription initiation requires the interaction of RNA polymerase with promoter DNA. In eukaryotic cells, there are three different types of RNA polymerases, each having particular functions and properties. RNA polymerase I is found in the nucleolar region of the nucleus and is responsible for transcribing the large ribosomal RNAs; RNA polymerase II transcribes messenger RNA precursors; and RNA polymerase III transcribes small RNAs such as transfer RNA, 5S ribosomal RNA, and other small DNA sequences. None of the eukaryotic RNA polymerases can bind efficiently to DNA. Rather, there are families of DNA-binding proteins that first bind to DNA and, once bound, interact with RNA polymerase to initiate mRNA synthesis. Initiation of Transcription by RNA Polymerase II and Associated Transcription Factors TFIIs and. TATA Binding Protein (TBP). Generally, the initiation of transcription by RNA polymerase II occurs as follows: First, transcription factors (TBPs and TFIIs) and RNA Polymerase are sequencially assembled on to DNA at the promoter site. Other TFIIs arrive next and DNA is unwound to form an open complex. This process requires ATP hydrolysis and promoter clearance during which the transcription initiation complex moves beyond the promoter site through to the coding region. After promoter clearance, some of the initiation factors disassociate with the RNAPII complex. The remaining TFIIs and the RNAPII enzyme are now considered part of the transcription elongation complex.")
8
intensificadores y silenciadores
Transcription Factor Binding Sites: Sites where regulatory transcription factors bind are often called X Regulatory Elements (XREs) or X Elements (XEs), where X is something to indicate what the regulation involves. For example: * ERE: Estrogen Response Element * HSE: Heat Shock Element * MRE: Metal Response Element * GRE: Glucocorticoid Response Element Enhancer and Silencer Regions: Regulators also bind to enhancer elements to upregulate transcription. Enhancers can activate transcription with upwards of s of base pairs of DNA between the enhancer and the rest of the promoter. Characteristics of Enhancers Enhancer elements act as protein binding sites for transcription factors, often the same ones that are found binding near the TATA box. Enhancers are found further away from the start site than typical promoter elements. They can be located upstream or downstream of the region being transcribed. They can work in either the 5' or 3' direction with respect to the coding region. Being such a far distance from the ORF suggests that the mechanism of enhancer action occurs by looping of the DNA. This change in DNA conformation would bring the enhancer elements in closer proximity to the main promoter region to exert an effect on transcription. The formation of the loop may also be facilitated by interactions between proteins binding the enhancer and proteins bound to regions closer to the ORF. Silencers act similarly to enhancers in that they regulate gene expression from a far distance. However, the function of the silencer, as its name implies, is to down-regulate gene expression. In other words, silencers keep gene expression off.
 or X Elements (XEs), where X is something to indicate what the regulation involves. For example: * ERE: Estrogen Response Element. * HSE: Heat Shock Element. * MRE: Metal Response Element. * GRE: Glucocorticoid Response Element. Enhancer and Silencer Regions: Regulators also bind to enhancer elements to upregulate transcription. Enhancers can activate transcription with upwards of s of base pairs of DNA between the enhancer and the rest of the promoter. Characteristics of Enhancers. Enhancer elements act as protein binding sites for transcription factors, often the same ones that are found binding near the TATA box. Enhancers are found further away from the start site than typical promoter elements. They can be located upstream or downstream of the region being transcribed. They can work in either the 5 or 3 direction with respect to the coding region. Being such a far distance from the ORF suggests that the mechanism of enhancer action occurs by looping of the DNA. This change in DNA conformation would bring the enhancer elements in closer proximity to the main promoter region to exert an effect on transcription. The formation of the loop may also be facilitated by interactions between proteins binding the enhancer and proteins bound to regions closer to the ORF. Silencers act similarly to enhancers in that they regulate gene expression from a far distance. However, the function of the silencer, as its name implies, is to down-regulate gene expression. In other words, silencers keep gene expression off.")
9
control de la transcripción eucariótica
ELEMENTOS EN CIS Y EN TRANS!! REVIEW Orchestrated response: a symphony of transcription factors for gene control Bryan Lemon,1 and Robert Tjian Genes Dev. 14: , October 15, 2000 Figure 1. Fundamental elements of eukaryotic transcriptional control. (A) Early studies led to the identification of multiple eukaryotic RNA polymerases and the mapping of promoter and enhancer DNA sequences. Core promoter elements (grey) that direct transcription by RNA Pol II (yellow, green shading) include the TATA box (TATA), the initiator (INR), and the downstream promoter element (DPE). Promoter-proximal basal level enhancer elements (BLE, GC box, CCAAT box, green) were found in many genes; however, gene-specific signal-responsive distal enhancer elements were also identified and are represented here as the hormone responsive element (HRE, purple) and nuclear factor element (NFE, blue). (B) Sequence-specific DNA-binding transcription factors that were isolated biochemically include specificity protein-1 (Sp1, green), families of activator proteins (i.e., AP-1, green), CCAAT enhancer-binding proteins (C/EBP), steroid receptors (i.e., glucocorticoid or GR, purple) and tissue-specific transcription factors (i.e., NF-B, blue). Components of the core initiation machinery required by RNA Pol II for promoter recognition and basal transcription from naked DNA were also fractionated biochemically, including TATA-binding protein, (TBP, red) and general transcription factors (TFIIA, TFIIB, TFIID, TFIIE, TFIIF and TFIIH, purple). (C) Because TBP is insufficient for activated transcription, discovery of TAF subunits (orange) within TFIID revealed a requirement for co-activators to mediate activator responsiveness. Depicted here is one example of a specific and functional interaction between Sp1 and Drosophila TAF110 (human TAF130). (D) Many co-activators and co-repressors were subsequently found to be required for mediating signals between sequence-specific transcription factors and the core machinery. For further discussion pertaining to the specific co-regulators depicted, see text and Table 1.
 Early studies led to the identification of multiple eukaryotic RNA polymerases and the mapping of promoter and enhancer DNA sequences. Core promoter elements (grey) that direct transcription by RNA Pol II (yellow, green shading) include the TATA box (TATA), the initiator (INR), and the downstream promoter element (DPE). Promoter-proximal basal level enhancer elements (BLE, GC box, CCAAT box, green) were found in many genes; however, gene-specific signal-responsive distal enhancer elements were also identified and are represented here as the hormone responsive element (HRE, purple) and nuclear factor element (NFE, blue). (B) Sequence-specific DNA-binding transcription factors that were isolated biochemically include specificity protein-1 (Sp1, green), families of activator proteins (i.e., AP-1, green), CCAAT enhancer-binding proteins (C/EBP), steroid receptors (i.e., glucocorticoid or GR, purple) and tissue-specific transcription factors (i.e., NF-B, blue). Components of the core initiation machinery required by RNA Pol II for promoter recognition and basal transcription from naked DNA were also fractionated biochemically, including TATA-binding protein, (TBP, red) and general transcription factors (TFIIA, TFIIB, TFIID, TFIIE, TFIIF and TFIIH, purple). (C) Because TBP is insufficient for activated transcription, discovery of TAF subunits (orange) within TFIID revealed a requirement for co-activators to mediate activator responsiveness. Depicted here is one example of a specific and functional interaction between Sp1 and Drosophila TAF110 (human TAF130). (D) Many co-activators and co-repressors were subsequently found to be required for mediating signals between sequence-specific transcription factors and the core machinery. For further discussion pertaining to the specific co-regulators depicted, see text and Table 1.")
10
inducción de la expresión génica por hormonas esteroides
Figure: 16-19 The Effect of a Steroid Hormone on Gene Expression Las hormonas esteroideas, moleculas hidrofóbicas, se difunden libremente en el interior de las células. Sus células "diana" contienen proteínas citoplásmicas y/o nucleares que funcionan como receptores hormonales. La hormona se une al receptor y el complejo actúa como un factor de transcripción estimulando o frenando a los “diana”, de los genes. las hormonas esteroides son: * Corticoides: glucocorticoides y mineralocorticoides. Existen múltiples fármacos con actividad corticoide, como la prednisona. * Hormonas sexuales masculinas: son los andrógenos como la testosterona y sus derivados, los anabolizantes androgénicos esteroides; estos últimos llamados simplemente esteroides. * Hormonas sexuales femeninas. * Vitamina D y sus derivados. Hormonas placentarias Estrógenos Esteroides Funciones: Mantenimiento del embarazo Progestinas Funciones: Imitan la acción de la progesterona Hormonas gonadales Estrógenos (ováricas) Esteroides, estradiol y estrona Funciones: Maduración y función de los órganos sexuales secundarios Progestinas (ováricas) Esteroide, progesterona Funciones: Implantación del óvulo y mantenimiento del embarazao Andrógenos (testicular) Esteroide, testosterona Hormonas cortico-suprarrenales Glucocorticoides Esteroides, cortisol y corticosterona Funciones: Diveros efectos sobre la inflamación y la síntesis de proteína Mineralocorticoides Esteroides, aldosterona Funciones: Mantenimiento del balance de la sal Hormone regulation - another good example of short-term regulation of transcription: Cells of higher eukaryotes are specialized and generally shielded from rapid changes in the external environment. Hormone signals are one mechanism for regulating transcription in response to demands of the environment. Hormones act as inducers produced by one cell and cause a physiological response in another cell. Hormones act only on target cells with hormone specific receptors, and levels of hormones are maintained by feedback pathways. Hormones deliver signals in two different ways: Steroid hormones pass through the cell membrane and bind cytoplasmic receptors, which together bind directly to DNA and regulate gene expression. Polypeptide hormones bind at the cell surface and activate transmembrane enzymes to produce second messengers (such as cAMP) that activate gene transcription. The biological effects of steroid hormones are mediated by receptors associated with the plasma membrane as well as located inside of target cells. This perspective focuses on recent advances in our understanding of the integration that occurs between membrane-associated rapid signaling events and various changes in gene transcription that modulate the function and phenotype of steroid-responsive cells. Three frequently studied members of the steroid hormone receptor superfamily, the estrogen receptors, the thyroid hormone receptors, and the vitamin D receptors, are included to illustrate the emerging concepts. Each of these hormones has been conclusively shown to function at multiple subcellular sites leading to a continuum of signals intimately linked by intracellular cross talk. Understanding the molecular mechanisms by which these steroid hormones and their receptors transduce cellular signals will allow us to create new pharmacologic therapies aimed at treatment of a variety of human diseases affecting the cardiovascular system, the reproductive system, the skeletal system, the nervous system, the mammary gland, and many others.
 Esteroides, estradiol y estrona. Funciones: Maduración y función de los órganos sexuales secundarios. Progestinas (ováricas) Esteroide, progesterona. Funciones: Implantación del óvulo y mantenimiento del embarazao. Andrógenos (testicular) Esteroide, testosterona. Hormonas cortico-suprarrenales. Glucocorticoides. Esteroides, cortisol y corticosterona. Funciones: Diveros efectos sobre la inflamación y la síntesis de proteína. Mineralocorticoides. Esteroides, aldosterona. Funciones: Mantenimiento del balance de la sal. Hormone regulation - another good example of short-term regulation of transcription: Cells of higher eukaryotes are specialized and generally shielded from rapid changes in the external environment. Hormone signals are one mechanism for regulating transcription in response to demands of the environment. Hormones act as inducers produced by one cell and cause a physiological response in another cell. Hormones act only on target cells with hormone specific receptors, and levels of hormones are maintained by feedback pathways. Hormones deliver signals in two different ways: Steroid hormones pass through the cell membrane and bind cytoplasmic receptors, which together bind directly to DNA and regulate gene expression. Polypeptide hormones bind at the cell surface and activate transmembrane enzymes to produce second messengers (such as cAMP) that activate gene transcription. The biological effects of steroid hormones are mediated by receptors associated with the plasma membrane as well as located inside of target cells. This perspective focuses on recent advances in our understanding of the integration that occurs between membrane-associated rapid signaling events and various changes in gene transcription that modulate the function and phenotype of steroid-responsive cells. Three frequently studied members of the steroid hormone receptor superfamily, the estrogen receptors, the thyroid hormone receptors, and the vitamin D receptors, are included to illustrate the emerging concepts. Each of these hormones has been conclusively shown to function at multiple subcellular sites leading to a continuum of signals intimately linked by intracellular cross talk. Understanding the molecular mechanisms by which these steroid hormones and their receptors transduce cellular signals will allow us to create new pharmacologic therapies aimed at treatment of a variety of human diseases affecting the cardiovascular system, the reproductive system, the skeletal system, the nervous system, the mammary gland, and many others.")
14
regulación postranscripcional (RNA)
• modificación (CAP y poliA) y procesamiento de intrones (splicing) transporte al citoplasma vida media siRNA mRNA transport control: Eukaryote mRNA transport is regulated. Some experiments show ~1/2 of primary transcripts never leave the nucleus and are degraded. Mature mRNAs exit through the nuclear pores. mRNA degradation control: All RNAs in the cytoplasm are subject to degradation. tRNAs and rRNAs usually are very stable; mRNAs vary considerably (minutes to months) -a diferencia de los mRNA procariótiucos que suelen durar sólo minutos!, aquí la variación de la vida media es mayor-. Stability may change in response to regulatory signals and is thought to be a major regulatory control point. Various sequences and processes affect mRNA half-life: AU-rich elements Secondary structure Deadenylation enzymes remove As from poly(A) tail 5’ de-capping Internal cleavage of mRNA and fragment degradation
 y. procesamiento de intrones (splicing) transporte al citoplasma. vida media. siRNA. mRNA transport control: Eukaryote mRNA transport is regulated. Some experiments show ~1/2 of primary transcripts never leave the nucleus and are degraded. Mature mRNAs exit through the nuclear pores. mRNA degradation control: All RNAs in the cytoplasm are subject to degradation. tRNAs and rRNAs usually are very stable; mRNAs vary considerably (minutes to months) -a diferencia de los mRNA procariótiucos que suelen durar sólo minutos!, aquí la variación de la vida media es mayor-. Stability may change in response to regulatory signals and is thought to be a major regulatory control point. Various sequences and processes affect mRNA half-life: AU-rich elements. Secondary structure. Deadenylation enzymes remove As from poly(A) tail. 5’ de-capping. Internal cleavage of mRNA and fragment degradation.")
15
adición de la caperuza (CAP) al 5’
RNA nuclear: hnRNA o transcrito pimario (uno de ellos pre-mRNA) caperuza (CAP): 7-metil-guanosina unida al 5’ se añade en la dirección opuesta! 3 funciones de CAP: transporte al citoplasma, estabilidad frente a degradación, reconocimiento ribosoma
 caperuza (CAP): 7-metil-guanosina unida al 5’ se añade en la dirección opuesta! 3 funciones de CAP: transporte al citoplasma, estabilidad frente a degradación, reconocimiento ribosoma.")
16
CAP The 5' end of all eukaryotic mRNAs are capped with a unique 5' -----> 5' linkage to a 7-methylguanosine residue. The capped end of the mRNA is thus, protected from exonucleases and more importantly is recognized by specific proteins of the translational machinery.

17
adición de la cola poliA al 3’
señal de poliadenilación la endonucleasa corta unos 20nt más alla de la señal de poliadenilación cola poliA: A finalmente: mRNA primario completo 2 funciones de cola poliA: transporte al citoplasma y estabilidad frente a degradación a veces a partir de un mismo preRNA ocurren poliadenilaciones alternativas, dando lugar a diferentes mRNa primarios

18
pre-mRNA primario -> mRNA primario completo -> mRNA funcional

19
maduración del mRNA eucariótico
ovoalbúmina: proteína de la clara del huevo eliminación de intrones

21
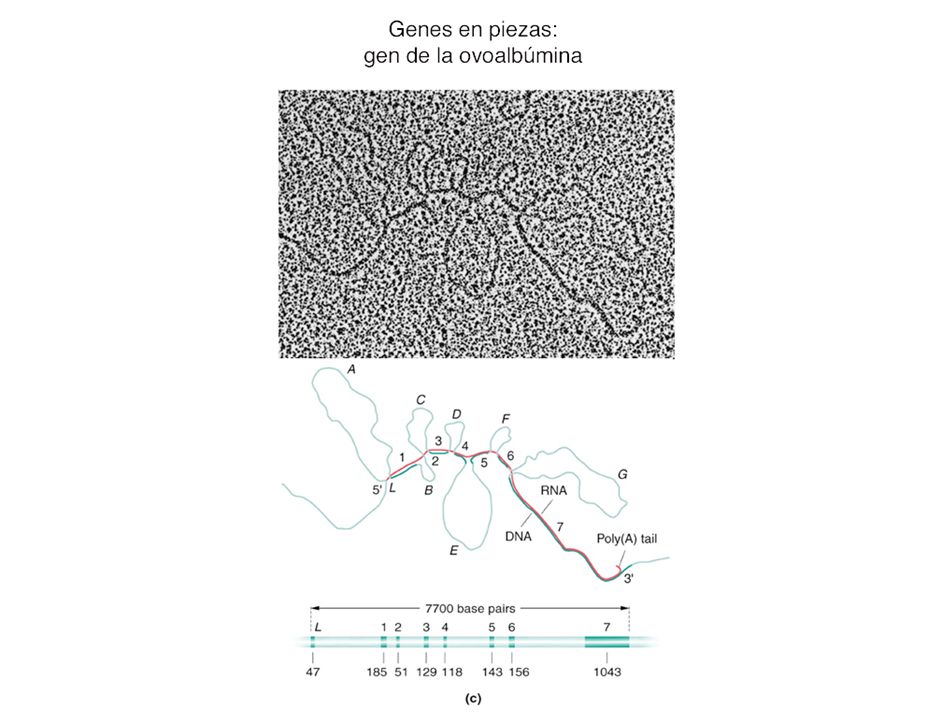
genes fragmentados o en piezas
Figure: Intervening Sequences in Various Eukaryotic Genes Intervening sequences in various eukaryotic genes. The numbers indicate the number of nucleotides present in various intron and exon regions.

22
Figure: 13-T07 Table 13-7 Comparing Human Gene Size, mRNA Size, and the Number of Introns

23
patrones de maduración del hnRNA de la tropomisoina
tropomiosina de ratas: 1 mRNA = 14 formas de la proteína troponina T muscular: 1 mRNA = 64 formas de la proteína Drosophila: macho/hembra depende de una maduración de tra tropomiosina de ratas: 1 mRNA = 14 formas de la proteína troponina T muscular: 1 mRNA = 64 formas de la proteína Drosophila: macho/hembra depende de una maduración de tra el ejemplo de la imagen, es para diferenciación celular y además en desarrollo... A precursor-RNA may often be matured to mRNAs with alternative structures. An example where alternative splicing has a dramatic consequence is somatic sex determination in the fruit fly Drosophila melanogaster. In this system, the female-specific sxl-protein is a key regulator. It controls a cascade of alternative RNA splicing decisions that finally result in female flies. Sex in Drosophila is largely determined by alternative splicing otro ejemplo es el proceso de formación de diferentes anticuerpos a partir del mismo gen. Generamos unos 10E6 a 10E8 de anticuerpos diferentes y sólo tenemos unos 35 mil genes! Exon shuffling (barajamineto de exones) During evolution, DNA segments coding for modules or domains in proteins have been duplicated and rearranged. By shuffling modules between genes, protein families have evolved. Blood coagulation factors represent such a family. Its members contain similar domains in various combinations and numbers. P=protease domain, GF=growth factor domain, K="kringle"-domain. Splicing errors cause thalassemia Thalassemia, a form of anemia common in the Mediterrarean countries, is caused by errors in the splicing process. Normal red blood cells contain correctly spliced beta-globin, an important component in hemoglobin that takes up oxygen in the lungs. The red blood cells in thalassemia patients are distorted and sometimes immature, containing a nucleus. This is due to a point mutation in the beta-globin gene, which causes an error in splice site selection. A faulty beta-globin protein is made, leading to severe anemia. The Philadelphia chromosome and cancer The small Philadelphia chromosome is present in cancer cells of almost all patients with chronic myeloic leukemia, a form of cancer of the blood. This chromosome is generated by breaks in the long arms of chromosomes 9 and 22 followed by an exchange of the tips of the chromosomes (translocation). This results in the creation of a fusion gene in the Ph chromosome, which then via a spliced mRNA, gives rise to a fusion protein. This abnormal protein is believed to play a key role in causing leukemia.
 During evolution, DNA segments coding for modules or domains in proteins have been duplicated and rearranged. By shuffling modules between genes, protein families have evolved. Blood coagulation factors represent such a family. Its members contain similar domains in various combinations and numbers. P=protease domain, GF=growth factor domain, K= kringle -domain. Splicing errors cause thalassemia. Thalassemia, a form of anemia common in the Mediterrarean countries, is caused by errors in the splicing process. Normal red blood cells contain correctly spliced beta-globin, an important component in hemoglobin that takes up oxygen in the lungs. The red blood cells in thalassemia patients are distorted and sometimes immature, containing a nucleus. This is due to a point mutation in the beta-globin gene, which causes an error in splice site selection. A faulty beta-globin protein is made, leading to severe anemia. The Philadelphia chromosome and cancer. The small Philadelphia chromosome is present in cancer cells of almost all patients with chronic myeloic leukemia, a form of cancer of the blood. This chromosome is generated by breaks in the long arms of chromosomes 9 and 22 followed by an exchange of the tips of the chromosomes (translocation). This results in the creation of a fusion gene in the Ph chromosome, which then via a spliced mRNA, gives rise to a fusion protein. This abnormal protein is believed to play a key role in causing leukemia.")
24
split genes = genes fragmentados o en piezas
secuencias intercaladas o intrones descubiertos en 1977 Richard J. Roberts – Autobiography ... In 1974, Richard Gelinas, whom I had first met at Harvard, joined my laboratory to characterize the initiation and termination signals for an Adenovirus-2 mRNA. The idea was to sequence the 5'-end of an mRNA, map its location on a restriction fragment, and then sequence the upstream region. This would be the promoter. Shortly after beginning the project, mRNA caps were discovered and we developed an assay for capped oligonucleotides. All seemed well until we came up with the startling finding that all late mRNAs seemed to begin with the same capped oligonucleotide, which was not encoded on the DNA next to the main body of the mRNA. We had excellent biochemical evidence for this, but real proof was elusive. In March, 1977, I hit on the right experiment to show that our proposed split structure for Adenovirus-2 mRNAs was correct. Louise Chow and Tom Broker, two talented electron microscopists, agreed to collaborate with us on the crucial experiment. We hoped to visualize the split structure by hybridizing an intact mRNA to its two different coding regions. Based on a guess about the location of the coding region for the 5'-end, we made appropriate DNA fragments. The reason for our guess turned out to be wrong, but luckily the fragment worked anyway! Finally, by direct visualization we could see the split genes in the electron microscope.

25
secuencias consenso en las uniones exón-intrón
para ‘corte y empalme’ Conservadas en intrón: 5’-GU y AG-3’

26
autoprocesamiento procesamiento por espliceosoma
autoprocesamiento: por formación de un lazo y trans-esterificación espliceosoma: snRNA + prt = small nuclear RiboNucleoProteína (snRNP) o snurps
 o snurps.")
28
genes en piezas y evolución: intrones temparnos
aparecen tempano en la evolución (en el RNA world, antes de las primeras células), pero los poerden los procariotas -aunque aparecen en ciertos fagos, cianobacterias y archebacterias- y sólo los eucariotas los mantienen 2.intrones tardíos (aparacen tras separacion proc-euc): barajamineto de exones (necesidad futura!! no evolución!) Exon shuffling (barajamineto de exones) -> explica la existencia de la variedad de proteínas During evolution, DNA segments coding for modules or domains in proteins have been duplicated and rearranged. By shuffling modules between genes, protein families have evolved. Blood coagulation factors represent such a family. Its members contain similar domains in various combinations and numbers. P=protease domain, GF=growth factor domain, K="kringle"-domain. 3. intrones = DNA egoísta??!! Splicing errors cause thalassemia Thalassemia, a form of anemia common in the Mediterrarean countries, is caused by errors in the splicing process. Normal red blood cells contain correctly spliced beta-globin, an important component in hemoglobin that takes up oxygen in the lungs. The red blood cells in thalassemia patients are distorted and sometimes immature, containing a nucleus. This is due to a point mutation in the beta-globin gene, which causes an error in splice site selection. A faulty beta-globin protein is made, leading to severe anemia. The Philadelphia chromosome and cancer The small Philadelphia chromosome is present in cancer cells of almost all patients with chronic myeloic leukemia, a form of cancer of the blood. This chromosome is generated by breaks in the long arms of chromosomes 9 and 22 followed by an exchange of the tips of the chromosomes (translocation). This results in the creation of a fusion gene in the Ph chromosome, which then via a spliced mRNA, gives rise to a fusion protein. This abnormal protein is believed to play a key role in causing leukemia.
, pero los poerden los procariotas -aunque aparecen en ciertos fagos, cianobacterias y archebacterias- y sólo los eucariotas los mantienen. 2.intrones tardíos (aparacen tras separacion proc-euc): barajamineto de exones (necesidad futura!! no evolución!) Exon shuffling (barajamineto de exones) -> explica la existencia de la variedad de proteínas. During evolution, DNA segments coding for modules or domains in proteins have been duplicated and rearranged. By shuffling modules between genes, protein families have evolved. Blood coagulation factors represent such a family. Its members contain similar domains in various combinations and numbers. P=protease domain, GF=growth factor domain, K= kringle -domain. 3. intrones = DNA egoísta !! Splicing errors cause thalassemia. Thalassemia, a form of anemia common in the Mediterrarean countries, is caused by errors in the splicing process. Normal red blood cells contain correctly spliced beta-globin, an important component in hemoglobin that takes up oxygen in the lungs. The red blood cells in thalassemia patients are distorted and sometimes immature, containing a nucleus. This is due to a point mutation in the beta-globin gene, which causes an error in splice site selection. A faulty beta-globin protein is made, leading to severe anemia. The Philadelphia chromosome and cancer. The small Philadelphia chromosome is present in cancer cells of almost all patients with chronic myeloic leukemia, a form of cancer of the blood. This chromosome is generated by breaks in the long arms of chromosomes 9 and 22 followed by an exchange of the tips of the chromosomes (translocation). This results in the creation of a fusion gene in the Ph chromosome, which then via a spliced mRNA, gives rise to a fusion protein. This abnormal protein is believed to play a key role in causing leukemia.")
29
descubren el autoprocesamiento del RNA!
RNA con actividad catalítica: ribozimas! Los intrones actúan como ribozimas. Descubierto en 1982 DNA no autocatálisis, pero sí RNA! -> RNA world: RNA fue el material genético original en el inicio de la vida en la Tierra

30
transporte al citoplasma
mRNA transport control: Eukaryote mRNA transport is regulated. Some experiments show ~1/2 of primary transcripts never leave the nucleus and are degraded. Mature mRNAs exit through the nuclear pores.

31
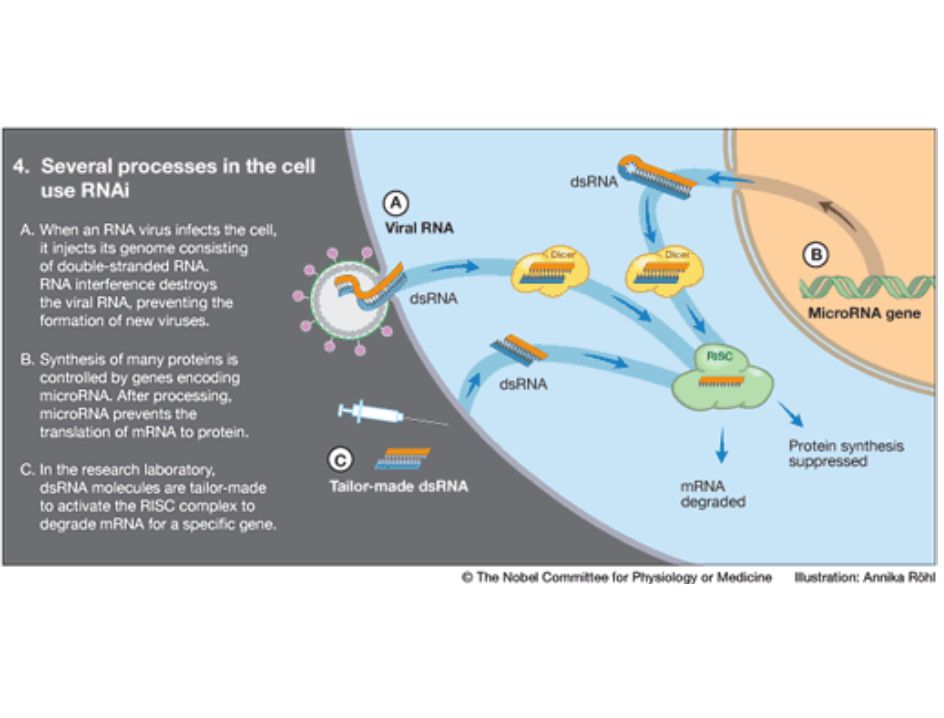
siRNA siRNA: small interfering RNA Dicer RISC: RNA-induced
silencing complex Dicer: prt q fragamenta el dsRNA RISC es reciclado y sigue actuando RISC esta formado por: helicasa + endonucleasa + exonucleasa + prt que ayuda a encontrar holomogia entre los RNS + RNa guía (el pequeño RNA que se genera tras la degradación del dsRNA) aunque este mecanismo no existe en bacterias, el equivalente sería el asRNA (RNA antisentido) The RNA interference machinery is unraveled The components of the RNAi machinery were identified during the following years (Fig 3). Double-stranded RNA binds to a protein complex, Dicer, which cleaves it into fragments. Another protein complex, RISC, binds these fragments. One of the RNA strands is eliminated but the other remains bound to the RISC complex and serves as a probe to detect mRNA molecules. When an mRNA molecule can pair with the RNA fragment on RISC, it is bound to the RISC complex, cleaved and degraded. The gene served by this particular mRNA has been silenced. 1. RNA interference – a defense against viruses and jumping genes 2. RNA interference regulates gene expression RNA interference is used to regulate gene expression in the cells of humans as well as worms (Fig 4B). Hundreds of genes in our genome encode small RNA molecules called microRNAs. They contain pieces of the code of other genes. Such a microRNA molecule can form a double-stranded structure and activate the RNA interference machinery to block protein synthesis. The expression of that particular gene is silenced. We now understand that genetic regulation by microRNAs plays an important role in the development of the organism and the control of cellular functions. 3. New opportunities in biomedical research, gene technology and health care siRNA How does RNAi work? Genetic and biochemical data indicate a possible two-step mechanism for RNA interference (RNAi): an initiation step and an effector step. a | In the first step, input double-stranded (ds) RNA is processed into 21–23-nucleotide 'guide sequences'. Whether they are single- or double-stranded remains an open question. An RNA amplification step (shaded box) has been suggested on the basis of the unusual properties of the interference phenomenon in whole animals, but this has not been reproduced definitively in vitro. b | The guide RNAs are incorporated into a nuclease complex, called the RNA-induced silencing complex (RISC), which acts in the second effector step to destroy mRNAs that are recognized by the guide RNAs through base-pairing interactions. We also suggest the incorporation of an active mechanism to search for homologous mRNAs. (Endo, endonucleolytic nuclease; exo, exonucleolytic nuclease; recA, homology-searching activity related to E. coli recA.)
 aunque este mecanismo no existe en bacterias, el equivalente sería el asRNA (RNA antisentido) The RNA interference machinery is unraveled. The components of the RNAi machinery were identified during the following years (Fig 3). Double-stranded RNA binds to a protein complex, Dicer, which cleaves it into fragments. Another protein complex, RISC, binds these fragments. One of the RNA strands is eliminated but the other remains bound to the RISC complex and serves as a probe to detect mRNA molecules. When an mRNA molecule can pair with the RNA fragment on RISC, it is bound to the RISC complex, cleaved and degraded. The gene served by this particular mRNA has been silenced. 1. RNA interference – a defense against viruses and jumping genes. 2. RNA interference regulates gene expression. RNA interference is used to regulate gene expression in the cells of humans as well as worms (Fig 4B). Hundreds of genes in our genome encode small RNA molecules called microRNAs. They contain pieces of the code of other genes. Such a microRNA molecule can form a double-stranded structure and activate the RNA interference machinery to block protein synthesis. The expression of that particular gene is silenced. We now understand that genetic regulation by microRNAs plays an important role in the development of the organism and the control of cellular functions. 3. New opportunities in biomedical research, gene technology and health care. siRNA. How does RNAi work Genetic and biochemical data indicate a possible two-step mechanism for RNA interference (RNAi): an initiation step and an effector step. a | In the first step, input double-stranded (ds) RNA is processed into 21–23-nucleotide guide sequences . Whether they are single- or double-stranded remains an open question. An RNA amplification step (shaded box) has been suggested on the basis of the unusual properties of the interference phenomenon in whole animals, but this has not been reproduced definitively in vitro. b | The guide RNAs are incorporated into a nuclease complex, called the RNA-induced silencing complex (RISC), which acts in the second effector step to destroy mRNAs that are recognized by the guide RNAs through base-pairing interactions. We also suggest the incorporation of an active mechanism to search for homologous mRNAs. (Endo, endonucleolytic nuclease; exo, exonucleolytic nuclease; recA, homology-searching activity related to E. coli recA.)")
34
regulación traduccional
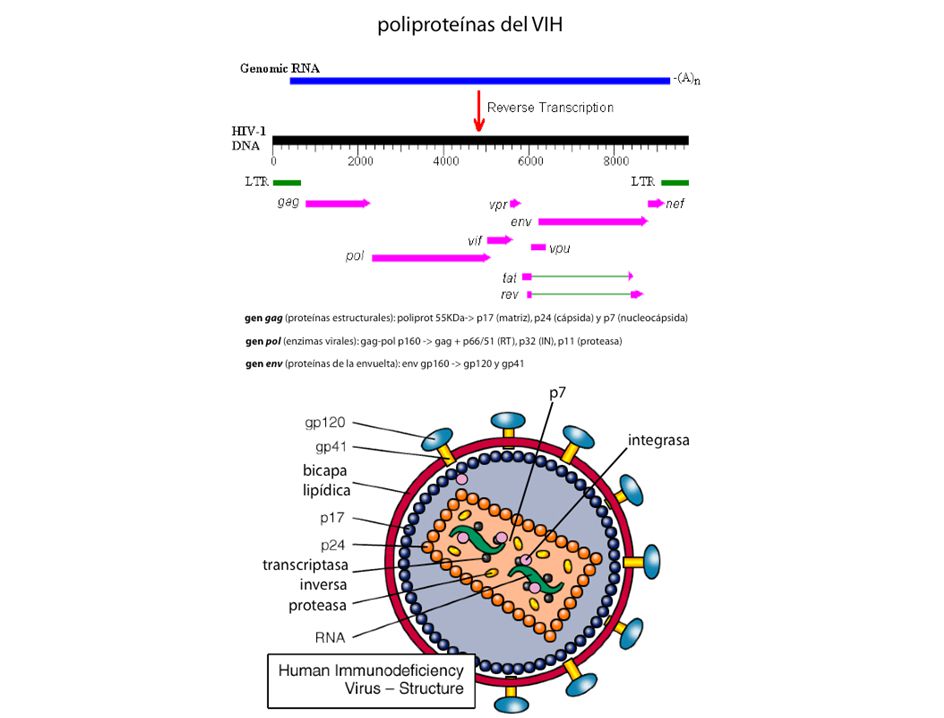
localización de ribosomas (preproteínas, péptidos líder) miRNA (microRNA) regulación postraduccional tutores moleculares (chaperonas) modificación: fosforilación, metilación metales pesados: dedos Zn, puños Cu preproteínas. Ej: insulina poliproteínas. Ej: VIH secuencias en cis localización de ribosomas (preproteínas, péptidos líder) microRNA tutores moleculares (carabinas o chaperonas) modificación: fosforilación, metilación metales pesados: Zn, Cu, Fe proproteínas: insulina poliproteínas: VIH
 miRNA (microRNA) regulación postraduccional. tutores moleculares (chaperonas) modificación: fosforilación, metilación. metales pesados: dedos Zn, puños Cu. preproteínas. Ej: insulina. poliproteínas. Ej: VIH. secuencias en cis. localización de ribosomas (preproteínas, péptidos líder) microRNA. tutores moleculares (carabinas o chaperonas) modificación: fosforilación, metilación. metales pesados: Zn, Cu, Fe. proproteínas: insulina. poliproteínas: VIH.")
35
regulación traduccional y post.
Figure: 16-12b Overview of Gene Regulation in Eukaryotes Various levels of regulation that are possible during the expression of the genetic material in eukaryotes.

36
secreción de proteínas en eucariotas
Secretory pathway in eukaryotic cells. Proteins whose synthesis begins in the cytosol are transported into the lumen of the endoplasmic reticulum. After further modification in the Golgi apparatus, the proteins are secreted. The lumen of the endoplasmic reticulum is topologically equivalent to the cell exterior. Proteins to be secreted are translated directly into the ER. These proteins are detected by signal recognition particles that halt translation and direct the ribosome to the membrane of the ER before translation can resume (See Figure 22.32).
.")
37
secreción de proteínas (bacterias)
")
38
péptidos señal de proteínas que se secretan (eucariotas)
Signal peptides from secreted proteins. Hydrophobic residues are shown in blue, and arrows mark the sites where the signal peptide is cleaved from the precursor. These sequences are the intial portion of the protein translated from the mRNA. The signal peptide is bound by a signal recognition protein that directs the continued translation only into the ER lumen (see Figure 22.32). This signal peptide is not found in the final, mature protein but is hydrolytically cleaved. en azul aa hidrofóbicos, flecha punto de corte
. This signal peptide is not found in the final, mature protein but is hydrolytically cleaved. en azul aa hidrofóbicos, flecha punto de corte.")
39
microRNA

40
metales pesados: dedo de Zinc
Figure: 16-16 Zinc Finger (a) A zinc finger where cysteine and histidine residues bind to a Zn++ atom. (b) The amino acid chain loops into a fingerlike configuration.
 A zinc finger where cysteine and histidine residues bind to a Zn++ atom. (b) The amino acid chain loops into a fingerlike configuration.")
41
preproteína: modificación de la proinsulina a insulina

43
virus hepatitis HCV

44
regulación a nivel de DNA
amplificación génica: rDNA genes en piezas o fragmentados reorganización cromosómica: Ig eu/heterocromatina: cromosoma X DNA-Z epigenética: impronta genética, paramutación rDNA: genes de rRNA en tandem (copias contiguas) genes en piezas o fragmentados = exones-intrones inmunoglobulinas: tenemos de 10E6-10E8 y sólo unos genes! eu/heterocromatina: expresión reglada por condensación/compactación del DNA DNA-Z: interacción diferente de DNA-B con proteínas EPIGENETICA: información que lleva el DNA pero no a nivel de secuencia (son alteraciones heredables). Impronta genómica + paramutación. Los factores epigenéticos incluyen tanto patrones espaciales, como la organización espacial del ADN alrededor de las proteínas histónicas (cromatina), como la marcación bioquímica (metilación de DNA, acetilación/metilación de histonas).
 genes en piezas o fragmentados = exones-intrones. inmunoglobulinas: tenemos de 10E6-10E8 y sólo unos genes! eu/heterocromatina: expresión reglada por condensación/compactación del DNA. DNA-Z: interacción diferente de DNA-B con proteínas. EPIGENETICA: información que lleva el DNA pero no a nivel de secuencia (son alteraciones heredables). Impronta genómica + paramutación. Los factores epigenéticos incluyen tanto patrones espaciales, como la organización espacial del ADN alrededor de las proteínas histónicas (cromatina), como la marcación bioquímica (metilación de DNA, acetilación/metilación de histonas).")
45
amplificación génica: rDNA y nucleolo

46
reorganización de genes: inmunoglobulinas

47
reorganización de genes: inmunoglobulinas
cadenas ligeras: genes V: 150 genes J: 5 uniones V-J: 100 cadenas pesadas: genes V: 80 genes D: 50 genes J: 6 unionesV-D-J: 100 Inmunoglobulinas: 291 genes = 18x109

48
eu/heterocromatina: cromosoma X

49
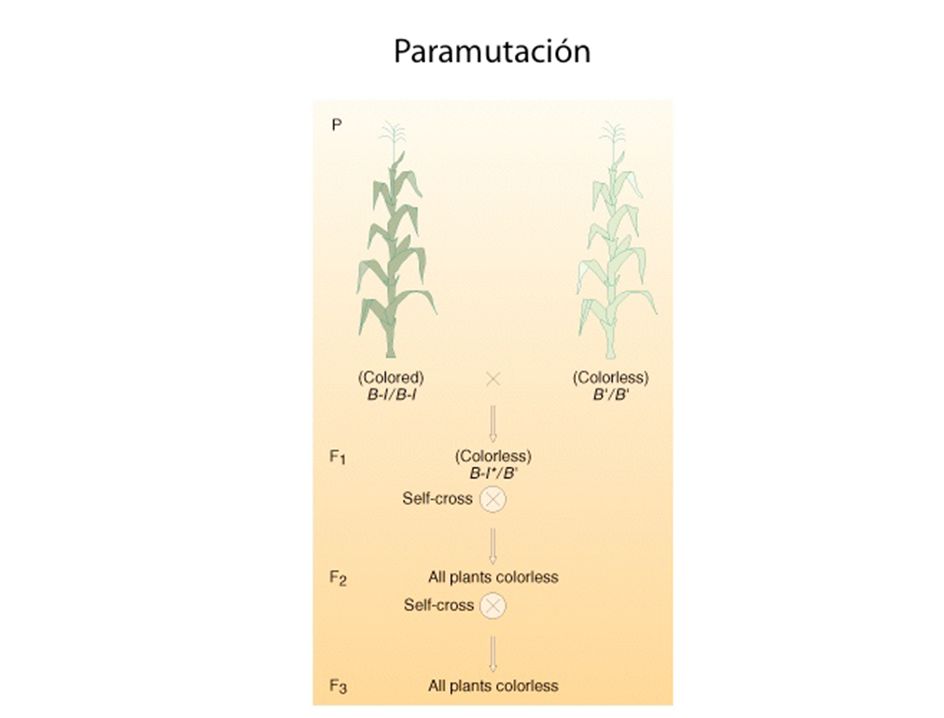
impronta genética La impronta genética es un fenómeno genético por el que ciertos genes son expresados de un modo específico que depende del sexo del progenitor. En los organismos diploides las células somáticas tienen dos copias del genoma. Por lo tanto, cada gen autosómico está representado por dos copias o alelos, cada una de ellas heredada de un progenitor en la fertilización. En la gran mayoría de los genes de los autosomas, la expresión de ambos alelos sucede simultáneamente. Sin embargo, una pequeña proporción de los genes (<1%) está "impresa", es decir, que su expresión depende de sólo uno de los alelos. La expresión del alelo depende, por tanto, de su origen parental. Un ejemplo de enfermedades genéticas humanas relacionadas con la impronta son el síndrome de Angelman y el síndrome de Prader-Willi, ambos ligados a la misma región del cromosoma 15. Un gen impreso, es decir, cuando tiene uno de los dos alelos silenciado,es funcionalmente haploide, lo que elimina la protección que confiere ser diploide contra mutaciones recesivas, además, su expresión puede ser desregulada epigenéticamente. Por tanto, estos genes representan locus suceptibles de ser alterados funcionalmente tanto genética como epigenéticamente. Actualmente, existen métodos para predecir el estado de impresión del genoma, diferenciando entre genes expresados monoalélicamente y bialélicamente y estudiando ciertas características típicas de estos genes desveladas gracias el estudio de determinadas regiones donde la impronta genética es bien conocida, como la banda 11p15.5 que presenta genes impresos tales como H19 e IGF2. Así por ejemplo, se conoce que los genes impresos son deficientes en secuencias repetitivas (short interspersed transposable elements: SINEs) como Alu. El síndrome de Angelman es una enfermedad genética rara que ocasiona un desorden neurológico en el cual se detectan dificultades severas de aprendizaje que están asociadas con características de apariencia facial y de comportamiento determinadas. El síndrome de Prader-Willi (SPW) es una alteración genética que ocasiona un cuadro clínico de obesidad, talla baja, hipogonadismo, criptorquidia y alteraciones en el aprendizaje tras una etapa de hipotonía muscular pre- y postnatal, dando la impresión de una lesión cerebral severa. El síndrome de Angelman es causado por la pérdida de una región del cromosoma 15, que principalmente es aportada de forma materna. Otra de las causas incluye la disomía uniparental, la translocación o la mutación puntual de un gen de esa región. Una persona sana recibe dos copias del cromosoma 15, uno de aportación materna y el otro por vía paterna. Sin embargo, la contribución materno-paterna es diferente ya que existen ciertos genes que se expresan de forma muy diferente dependiendo del sexo. Esto está muy relacionado con el fenómeno de impronta genética; este mecanismo consiste en la metilación del ADN. Si la contribución materna se pierde o muta, el resultado es el síndrome Angelman. Si por el contrario la pérdida o mutación ocurren en la contribución paterna el resultado se conoce como síndrome de Prader-Willi. El síndrome de Angelman también puede ser el resultado de una mutación puntual de un único gen. Este gen se conoce por el nombre de Ube3a, que es parte de la ruta de la ubiquitina. El gen está presente tanto en los cromosomas 15 de la madre como el del padre, pero difieren en el patrón de metilación (impronta). El patrón de silenciamiento génico del gen Ube3a ocurre de manera específica de las distintas regiones cerebrales; el alelo materno está siempre activo exclusivamente en el hipocampo y en el cerebelo. La gran mayoría de los casos se producen por una deleción de aproximadamente 4 Mb de la región materna 15q11-13, causando la ausencia de expresión de Ube3a en las regiones cerebrales especificadas anteriormente. Ube3a codifica para la proteína E6-AP ligasa de ubiquitina.La ausencia de esta proteína va a propiciar que se acumulen proteínas no degradadas. Esta enzima es muy selectiva y se han determinado cuatro sustratos dando un mecanismo molecular por el cual se llega al estado asociado al síndrome Angelman.
 está impresa , es decir, que su expresión depende de sólo uno de los alelos. La expresión del alelo depende, por tanto, de su origen parental. Un ejemplo de enfermedades genéticas humanas relacionadas con la impronta son el síndrome de Angelman y el síndrome de Prader-Willi, ambos ligados a la misma región del cromosoma 15. Un gen impreso, es decir, cuando tiene uno de los dos alelos silenciado,es funcionalmente haploide, lo que elimina la protección que confiere ser diploide contra mutaciones recesivas, además, su expresión puede ser desregulada epigenéticamente. Por tanto, estos genes representan locus suceptibles de ser alterados funcionalmente tanto genética como epigenéticamente. Actualmente, existen métodos para predecir el estado de impresión del genoma, diferenciando entre genes expresados monoalélicamente y bialélicamente y estudiando ciertas características típicas de estos genes desveladas gracias el estudio de determinadas regiones donde la impronta genética es bien conocida, como la banda 11p15.5 que presenta genes impresos tales como H19 e IGF2. Así por ejemplo, se conoce que los genes impresos son deficientes en secuencias repetitivas (short interspersed transposable elements: SINEs) como Alu. El síndrome de Angelman es una enfermedad genética rara que ocasiona un desorden neurológico en el cual se detectan dificultades severas de aprendizaje que están asociadas con características de apariencia facial y de comportamiento determinadas. El síndrome de Prader-Willi (SPW) es una alteración genética que ocasiona un cuadro clínico de obesidad, talla baja, hipogonadismo, criptorquidia y alteraciones en el aprendizaje tras una etapa de hipotonía muscular pre- y postnatal, dando la impresión de una lesión cerebral severa. El síndrome de Angelman es causado por la pérdida de una región del cromosoma 15, que principalmente es aportada de forma materna. Otra de las causas incluye la disomía uniparental, la translocación o la mutación puntual de un gen de esa región. Una persona sana recibe dos copias del cromosoma 15, uno de aportación materna y el otro por vía paterna. Sin embargo, la contribución materno-paterna es diferente ya que existen ciertos genes que se expresan de forma muy diferente dependiendo del sexo. Esto está muy relacionado con el fenómeno de impronta genética; este mecanismo consiste en la metilación del ADN. Si la contribución materna se pierde o muta, el resultado es el síndrome Angelman. Si por el contrario la pérdida o mutación ocurren en la contribución paterna el resultado se conoce como síndrome de Prader-Willi. El síndrome de Angelman también puede ser el resultado de una mutación puntual de un único gen. Este gen se conoce por el nombre de Ube3a, que es parte de la ruta de la ubiquitina. El gen está presente tanto en los cromosomas 15 de la madre como el del padre, pero difieren en el patrón de metilación (impronta). El patrón de silenciamiento génico del gen Ube3a ocurre de manera específica de las distintas regiones cerebrales; el alelo materno está siempre activo exclusivamente en el hipocampo y en el cerebelo. La gran mayoría de los casos se producen por una deleción de aproximadamente 4 Mb de la región materna 15q11-13, causando la ausencia de expresión de Ube3a en las regiones cerebrales especificadas anteriormente. Ube3a codifica para la proteína E6-AP ligasa de ubiquitina.La ausencia de esta proteína va a propiciar que se acumulen proteínas no degradadas. Esta enzima es muy selectiva y se han determinado cuatro sustratos dando un mecanismo molecular por el cual se llega al estado asociado al síndrome Angelman.")
51
flujo de la información genética
DNA RNA proteína DNA RNA proteína 1. transcripción inversa DNA RNA proteína 2. autorreplicación del RNA (RNA replicasa) "The central dogma, enunciated by Crick in 1958 and the keystone of molecular biology ever since, is likely to prove a considerable over-simplification". a añadir: flecha directa DNA -> proteínas (el DNA cómo mensajero). Esto sólo se da en condiciones de laboratorio DNA RNA proteína 3. priones
 The central dogma, enunciated by Crick in 1958 and the keystone of molecular biology ever since, is likely to prove a considerable over-simplification . a añadir: flecha directa DNA -> proteínas (el DNA cómo mensajero). Esto sólo se da en condiciones de laboratorio. DNA RNA proteína. 3. priones.")
Presentaciones similares