Descargar la presentación
La descarga está en progreso. Por favor, espere

1
Introducción a la Microbiología
La microbiología es hoy una ciencia dinámica que tiene ramificaciones en prácticamente todos los aspectos de la vida humana, tales como la medicina, la agricultura y el medio ambiente. La microbiología es el estudio de los microorganismos, un grupo amplio y diverso de organismos microscópicos que existen como células aisladas o asociadas; también incluye el estudio de los virus, que son microscópicos pero no celulares. Las células microbianas se distinguen pues de las células de animales y plantas, en que son incapaces de vivir aisladas en la naturaleza y sólo existen formando parte de organismos multicelulares . En general, a diferencia de los macroorganismos, los microorganismos son capaces de realizar sus procesos vitales de crecimiento, generación de energía y reproducción, independientemente de otras células, sean de la misma clase o de otra diferente. La ciencia de la microbiología La microbiología trata de las células vivas y su funcionamiento. Estudia los microorganismos especialmente las bacterias, un amplio grupo de células con una enorme importancia básica y aplicada. Trata de la diversidad microbiana y de la evolución, de cómo surgieron las diferentes clases de microorganismos y por qué. Analiza también lo que los microorganismos hacen en el mundo en general, en la sociedad humana, en el cuerpo humano, y en los cuerpos de animales y plantas.

2
De un modo u otro, los microorganismos influyen en todas las formas vivas de la Tierra y, por tanto, la ciencia de la microbiología tiene una gran trascendencia. La microbiología gira en torno a dos temas fundamentales, uno básico y otro aplicado: Como ciencia biológica básica La microbiología proporciona algunas de las herramientas de investigación más adecuadas para estudiar la naturaleza de los procesos vitales. El avanzado conocimiento que ahora tenemos de las bases físicas y químicas de la vida procede del estudio de los microorganismos. Se debe en parte a que las células microbianas comparten muchas propiedades bioquímicas con las células de organismos pluricelulares; de hecho, todas las células tienen mucho en común; unido al hecho de que las células microbianas pueden crecer hasta alcanzar una elevada densidad en cultivo en el laboratorio y son fácilmente manipulables en estudios bioquímicos y genéticos, hace de ellas excelentes modelos para el conocimiento de las funciones celulares en organismos superiores. 2. Como ciencia biológica aplicada, la microbiología trata de muchos problemas prácticos importantes en la medicina, la agricultura y la industria. Muchas de las enfermedades más importantes del hombre, de otros animales y de las

3
plantas, son producidas por microorganismos.
Los microorganismos también desempeñan una función destacada en la fertilidad del suelo y en la producción de animales domésticos. Además, muchos procesos industriales y biotecnológicos a gran escala, como la producción de antibióticos o de proteínas humanas, se basan en la utilización de microorganismos. Importancia de los microorganismos Los hombres, las plantas y los animales están íntimamente ligados a las actividades microbianas en cuanto al reciclado de los nutrientes esenciales o a la degradación de materia orgánica. En efecto, ninguna otra forma de vida tiene una importancia similar a la de los microorganismos en el mantenimiento de la vida sobre la Tierra. Los microorganismos existieron en la Tierra durante miles de millones de años antes de que aparecieran las plantas y los animales, cómo sus características fisiológicas los convierten en los mayores químicos de la Tierra, y cómo los microorganismos han establecido relaciones con los organismos superiores que pueden ser muy beneficiosas o extremadamente dañinas. Los microorganismos como células La célula es la unidad de vida fundamental. Una célula es una entidad aislada de otras células por una membrana celular (y tal vez por una pared celular) que contiene en su interior diversos compuestos y estructuras subcelulares (Fig. 1).
 que contiene en su interior diversos compuestos y estructuras subcelulares (Fig. 1).")
4
Fig.1 La membrana celular es la barrera que separa el interior de la célula del exterior. Dentro de la membrana celular se encuentran las diversas estructuras y componentes que hacen posible que la célula funcione. Son estructuras clave el núcleo o nucleoide, donde se guarda la información genética, el ácido desoxirribonucleico (DNA) necesario para hacer nuevas células, y el citoplasma, donde se encuentra la maquinaria para el crecimiento y otras funciones celulares. Todas las células están formadas por cuatro tipos de componentes químicos: proteínas, ácidos nucleicos, lípidos y polisacáridos. En conjunto, se denominan macromoléculas. La naturaleza química y la disposición de las macromoléculas de una célula de un organismo es lo que diferencia una célula de otro organismo.
 necesario para hacer nuevas células, y el citoplasma, donde se encuentra la maquinaria para el crecimiento y otras funciones celulares. Todas las células están formadas por cuatro tipos de componentes químicos: proteínas, ácidos nucleicos, lípidos y polisacáridos. En conjunto, se denominan macromoléculas. La naturaleza química y la disposición de las macromoléculas de una célula de un organismo es lo que diferencia una célula de otro organismo.")
5
Aunque cada tipo de célula tiene una estructura y un tamaño definido, una célula es una unidad dinámica, que realiza constantes cambios reemplazando sus componentes. Incluso cuando no está creciendo, una célula puede estar tomando materiales del medio para incorporarlos a su propia estructura. Al mismo tiempo, libera productos de desecho en el medio. Por tanto, una célula es un sistema abierto en constante cambio y, sin embargo, permanece igual. ¿De dónde proceden las primeras células? De alguna forma, la primera célula debió de originarse a partir de algo no celular, algo anterior a la célula, una estructura precelular. Una vez que la primera célula apareció se produjeron una serie de sucesos altamente probables, tales como el crecimiento y la división, originándose poblaciones de células a partir de las cuales tuvo lugar la evolución mediante la selección de nuevas características y mayor diversidad. De ese modo, a través de miles de millones de años de cambio evolutivo, surgió la impresionante diversidad de tipos celulares que existen en la actualidad. Debido a que todas las células están construidas con las cuatro clases de macromoléculas básicas mencionadas anteriormente y comparten muchos rasgos en común, se piensa que todas las células descienden de un antecesor común, el antecesor universal de la vida.

6
Fig. 2 Características de los sistemas vivos De lo que sabemos hoy en biología, podemos identificar varias características que son compartidas por la mayoría de los sistemas vivos, y que se resumen en la (Fig. 2). Todos los organismos celulares son estructuras altamente organizadas que muestran alguna forma de metabolismo. Esto es, las células toman sustancias químicas del medio y las transforman, conservan parte de la energía de dichas sustancias de modo que las células puedan usarla, y luego eliminan los productos de desecho. Todas las células poseen reproducción, es decir, son capaces de dirigir una serie de reacciones bioquímicas que conduce a su propia síntesis.
. Todos los organismos celulares son estructuras altamente organizadas que muestran alguna forma de metabolismo. Esto es, las células toman sustancias químicas del medio y las transforman, conservan parte de la energía de dichas sustancias de modo que las células puedan usarla, y luego eliminan los productos de desecho. Todas las células poseen reproducción, es decir, son capaces de dirigir una serie de reacciones bioquímicas que conduce a su propia síntesis.")
7
Como resultado de los procesos metabólicos, una célula crece y se divide para formar dos células.
Muchas células sufren diferenciación, un proceso por el que se forman nuevas sustancias o estructuras. A menudo, la diferenciación celular es parte de un ciclo vital en el que las células forman estructuras especiales relacionadas con la reproducción, la dispersión o la supervivencia. Las células responden a señales químicas en su medio ambiente, tales como las producidas por otras células. Por tanto pueden comunicarse e incluso estimar su propio número en el ambiente circundante por medio de pequeñas moléculas que se difunden y pasan entre células vecinas. Con frecuencia los organismos vivos tienen movimiento por autopropulsión, y en el mundo microbiano veremos diferentes mecanismos responsables de la movilidad.

8
Finalmente, a diferencia de las estructuras inertes, las células pueden evolucionar. A través del proceso de evolución las células pueden cambiar permanentemente sus características y transmitir las nuevas propiedades a su descendencia. Las células como máquinas y como sistemas codificados Las células se pueden considerar bajo dos aspectos. Por un lado, las células pueden ser consideradas como máquinas químicas que llevan a cabo transformaciones químicas dentro de los límites de la estructura celular. Los catalizadores de esta máquina química son las enzimas, proteínas capaces de acelerar notablemente la velocidad de reacciones químicas específicas. Por otro lado, las células también pueden ser consideradas como sistemas codificados, análogos a las computadoras, que guardan y procesan la información genética (DNA) que pasa finalmente a la descendencia durante la reproducción (Fig. 3).
 que pasa finalmente a la descendencia durante la reproducción (Fig. 3).")
9
Fig. 3 En realidad, las células son máquinas químicas y sistemas codificados, y el enlace entre estos dos atributos es el crecimiento. En condiciones adecuadas, una célula viable aumenta de tamaño y luego se divide para formar dos células (Fig. 3). En el proceso ordenado que supone la división celular, la cantidad de todos los constituyentes de la célula se duplica.
. En el proceso ordenado que supone la división celular, la cantidad de todos los constituyentes de la célula se duplica.")
10
Esto implica el funcionamiento de la maquinaria química de la célula que suministra la energía y los precursores necesarios para la biosíntesis de las macromoléculas. Pero cuando una célula se divide, cada una de las dos células resultantes debe contener toda la información genética necesaria para la formación de más células y, por tanto, durante el proceso de crecimiento debe haber también una duplicación del DNA (Fig. 3). En consecuencia, tanto la máquina como el código deben estar funcionalmente coordinados para lograr que una célula se reproduzca con fidelidad. Además de la coordinación, las diversas funciones de la máquina y del código están sujetas a una regulación, de modo que la célula se adapta a su medio ambiente. Los microorganismos y sus ambientes naturales En la naturaleza, las células viven asociadas a otras en conjuntos llamados poblaciones. Tales poblaciones se componen de grupos de células relacionadas, que generalmente derivan de una única célula parental por divisiones celulares sucesivas. El lugar donde vive una población microbiana en un determinado ambiente se denomina hábitat. Los microorganismos pueden encontrarse tanto en ambientes familiares como en lugares poco comunes, como aquellos tan extremos que se consideran inadecuados para formas de vida superiores. Las poblaciones celulares raramente viven solas en la naturaleza, antes bien se relacionan con otras formando las llamadas comunidades microbianas (Fig. 4).
. En consecuencia, tanto la máquina como el código deben estar funcionalmente coordinados para lograr que una célula se reproduzca con fidelidad. Además de la coordinación, las diversas funciones de la máquina y del código están sujetas a una regulación, de modo que la célula se adapta a su medio ambiente. Los microorganismos y sus ambientes naturales. En la naturaleza, las células viven asociadas a otras en conjuntos llamados poblaciones. Tales poblaciones se componen de grupos de células relacionadas, que generalmente derivan de una única célula parental por divisiones celulares sucesivas. El lugar donde vive una población microbiana en un determinado ambiente se denomina hábitat. Los microorganismos pueden encontrarse tanto en ambientes familiares como en lugares poco comunes, como aquellos tan extremos que se consideran inadecuados para formas de vida superiores. Las poblaciones celulares raramente viven solas en la naturaleza, antes bien se relacionan con otras formando las llamadas comunidades microbianas (Fig. 4).")
11
Fig. 4 Estas comunidades pueden estar integradas por células libres en medios acuáticos, pero a menudo forman los llamados biofilms sobre superficies vivas o inertes. Ej: en lagos ó en aguas residuales. Efecto de los organismos entre sí y en sus hábitat Las poblaciones de las comunidades microbianas se relacionan de varios modos y tales interacciones pueden ser perjudiciales o beneficiosas. En muchos casos, las poblaciones interaccionan y cooperan en sus funciones nutricionales

12
con los productos de desecho derivados de las actividades metabólicas de algunas células sirviendo como nutrientes para otras. Los organismos de un hábitat también se relacionan con su ambiente físico y químico. Los hábitat tienen características diferentes, y un hábitat que favorece el crecimiento de un organismo puede ser dañino para otro. Por tanto, la composición de una comunidad microbiana en un hábitat concreto está determinada en gran parte por las características físicas y químicas de ese medio. En conjunto, denominamos ecosistema a los organismos y a los componentes físicos y químicos de su medio. Hay importantes ecosistemas microbianos acuáticos (océanos, estanques, lagos, corrientes, fuentes termales...), terrestres (suelos, rocas...) e incluso asociados a organismos superiores, plantas o animales. Las propiedades de un ecosistema están controladas en gran parte por las actividades microbianas. Los organismos obtienen los nutrientes del medio para sus procesos metabólicos y los usan para formar nuevas células. Al mismo tiempo, los organismos eliminan al medio los productos de desecho de su metabolismo. Así, con el tiempo, un ecosistema microbiano puede cambiar gradualmente, tanto desde el punto de vista físico como químico. El oxígeno gaseoso constituye un buen ejemplo. El oxígeno molecular, O2, es un nutriente vital para algunos microorganismos mientras que es venenoso para otros.
, terrestres (suelos, rocas...) e incluso asociados a organismos superiores, plantas o animales. Las propiedades de un ecosistema están controladas en gran parte por las actividades microbianas. Los organismos obtienen los nutrientes del medio para sus procesos metabólicos y los usan para formar nuevas células. Al mismo tiempo, los organismos eliminan al medio los productos de desecho de su metabolismo. Así, con el tiempo, un ecosistema microbiano puede cambiar gradualmente, tanto desde el punto de vista físico como químico. El oxígeno gaseoso constituye un buen ejemplo. El oxígeno molecular, O2, es un nutriente vital para algunos microorganismos mientras que es venenoso para otros.")
13
Como las células microbianas aisladas son demasiado pequeñas para observar a simple vista, el conocimiento de los microorganismos en la naturaleza comienza con estudios que utilizan el microscopio. El examen de materiales naturales, como el suelo o el agua, siempre pone de manifiesto la presencia de células microbianas. Aunque esas células tan diminutas pueden parecer triviales, son capaces de multiplicarse rápidamente y de producir grandes poblaciones que pueden tener un gran impacto en el hábitat. Así, pese a que los microorganismos pueden pasar por ser en la naturaleza componentes minoritarios, son parte muy importante de cada ecosistema. La importancia de la vida microbiana Resulta común suponer que como los microorganismos son tan pequeños, su biomasa en la Tierra también debe ser pequeña en comparación con la biomasa de organismos superiores. Sin embargo, esto no es probablemente cierto. Cálculos detallados sobre el número total de células microbianas en la Tierra, concretamente, sobre el número total de procariotas (también llamadas bacterias, células pequeñas que carecen de núcleo indican que este número es del orden de 5 X 1030 células. La cantidad total de carbono que está presente en este número tan grande de células tan pequeñas equivale al de todas las plantas de la Tierra, mientras que el contenido total de

14
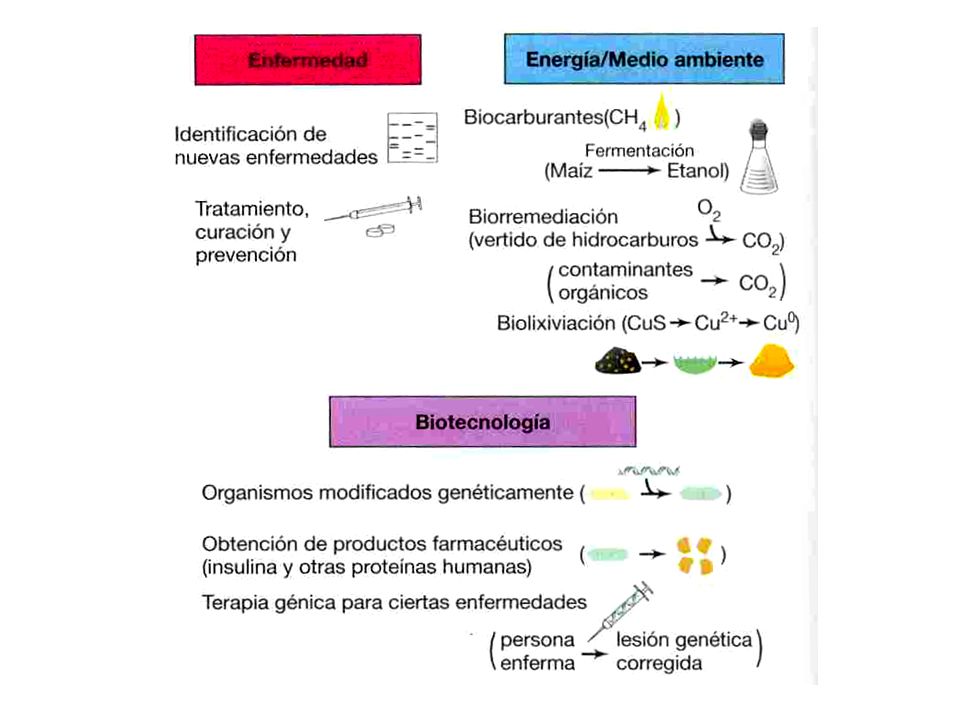
nitrógeno y fósforo en estas células procarióticas es 10 veces mayor que la de toda la biomasa vegetal. Por tanto, las células procarióticas, por pequeñas que sean, constituyen la mayor porción de biomasa sobre la Tierra y son reservas de nutrientes esenciales para la vida. Es interesante señalar que la mayor parte de las células procarióticas no se encuentra en la superficie de la Tierra, sino en zonas bajo la superficie en los ambientes oceánicos y terrestres. El impacto de los microorganismos sobre el hombre Uno de los objetivos de los microbiólogos es comprender cómo trabajan los microorganismos y, a través de ese conocimiento, diseñar modos mediante los cuales su efecto beneficioso pueda ser aumentado y el perjudicial reducido. Los microbiólogos han tenido mucho éxito en conseguir estos fines y la microbiología ha sido muy importante en los avances de la salud humana y el bienestar. Una visión global del impacto de los microorganismos en las actividades humanas se muestra en la (Fig. 5).
.")
15
Los microorganismos como agentes etiológicos de enfermedades
Al comienzo del siglo xx las causas de muerte más frecuentes eran las enfermedades infecciosas; en la actualidad tales enfermedades son mucho menos importantes. El control de las enfermedades infecciosas se ha logrado por un conocimiento integrado de los procesos infecciosos, por la mejora de las prácticas sanitarias y por el descubrimiento y uso de los agentes antimicrobianos. Enfermedades microbianas tan difundidas como la malaria, la tuberculosis, el cólera, la enfermedad del sueño africana y síndromes diarreicos severos, todavía producen muertes de seres humanos. Por tanto, los microorganismos todavía constituyen una seria amenaza. No obstante, hay que destacar que la mayor parte de los microorganismos no son perjudiciales para el hombre. De hecho, la gran mayoría son en realidad beneficiosos y llevan a cabo procesos de enorme valor para la sociedad. A continuación, consideramos algunos de estos efectos favorables.

16
Fig. 5

18
Microorganismos y agricultura
En general, nuestros sistemas de agricultura dependen en muchos aspectos de las actividades microbianas. Un gran número de cosechas se debe al cultivo de miembros de un grupo de plantas llamadas leguminosas, que viven en asociación muy estrecha con bacterias específicas que forman estructuras en sus raíces llamadas nódulos. En estos nódulos radiculares, el nitrógeno atmosférico (N2) se convierte por fijación en compuestos nitrogenados que las plantas utilizan para crecer. De este modo, las actividades de las bacterias contenidas en los nódulos de las raíces reducen la necesidad de fertilizantes costosos para plantas. También tienen gran importancia agrícola los microorganismos que son esenciales en el proceso digestivo de los rumiantes, como las vacas y las ovejas. Estos animales poseen un órgano digestivo especial llamado rumen, donde los microorganismos realizan el proceso digestivo. Sin estos microorganismos las vacas y las ovejas no podrían digerir su alimento y, por tanto, no podrían desarrollarse sobre sustancias tan pobres en nutrientes como la hierba o el heno. Los microorganismos también desempeñan funciones críticas en el reciclado de elementos importantes para la nutrición vegetal, en particular del carbono, nitrógeno y azufre. En el suelo y en el agua, los microorganismos convierten estos elementos en formas asimilables por las plantas. Además de beneficios, los microorganismos también acarrean perjuicios a la agricultura. Las enfermedades microbianas de animales y plantas tienen un importante impacto económico.
 se convierte por fijación en compuestos nitrogenados que las plantas utilizan para crecer. De este modo, las actividades de las bacterias contenidas en los nódulos de las raíces reducen la necesidad de fertilizantes costosos para plantas. También tienen gran importancia agrícola los microorganismos que son esenciales en el proceso digestivo de los rumiantes, como las vacas y las ovejas. Estos animales poseen un órgano digestivo especial llamado rumen, donde los microorganismos realizan el proceso digestivo. Sin estos microorganismos las vacas y las ovejas no podrían digerir su alimento y, por tanto, no podrían desarrollarse sobre sustancias tan pobres en nutrientes como la hierba o el heno. Los microorganismos también desempeñan funciones críticas en el reciclado de elementos importantes para la nutrición vegetal, en particular del carbono, nitrógeno y azufre. En el suelo y en el agua, los microorganismos convierten estos elementos en formas asimilables por las plantas. Además de beneficios, los microorganismos también acarrean perjuicios a la agricultura. Las enfermedades microbianas de animales y plantas tienen un importante impacto económico.")
19
Microorganismos y alimentación
Una vez producidas las cosechas, productos agrícolas, o los animales en explotaciones ganaderas, éstos deben llegar a los consumidores con calidad sanitaria. De ahí que los microorganismos tengan una gran importancia en la industria alimentaria. El deterioro de los alimentos ocasiona anualmente pérdidas económicas inmensas. Las industrias de enlatado, congelado y desecado de alimentos tienen como finalidad preparar alimentos de tal modo que no sufran deterioro por microorganismos. Las enfermedades transmitidas por alimentos también son dignas de consideración. Los alimentos deben estar adecuadamente preparados y controlados para evitar la transmisión de enfermedades, ya que el alimento apto para el consumo humano puede servir también para sustentar el crecimiento de muchos microorganismos. Sin embargo, no todos los microorganismos tiene efectos indeseables sobre los alimentos o sobre los consumidores. Por ejemplo, los productos lácteos que se manufacturan en gran medida gracias a actividades microbianas, tales como el queso, el yogurt o la mantequilla, son productos de gran valor económico. De modo similar, la col ácida, los pepinillos y algunas formas de salchichas deben también sus propiedades a los microorganismos. Los alimentos de panadería se elaboran usando levaduras. Las bebidas alcohólicas, ampliamente difundidas en nuestra sociedad, también son producidas por las levaduras.

20
Microorganismos, energía y medio ambiente
En lo que respecta a la energía, los microorganismos desempeñan funciones clave. La mayor parte del gas natural (metano) es un producto bacteriano, derivado de las actividades de las bacterias metanogénicas. Los microorganismos fotótrofos pueden utilizar la luz como fuente de energía para la producción de biomasa, es decir, energía acumulada en forma de organismos vivos. La biomasa microbiana y los materiales de desecho, como la basura doméstica, los excedentes de cosechas y los residuos animales, se pueden convertir en «biocombustibles», como el metano y el etanol, por las actividades degradativas de los microorganismos. Los microorganismos también se pueden usar para ayudar a eliminar la polución originada por las actividades humanas, un proceso que se denomina biorremediación. Se han aislado varios microorganismos de la naturaleza que consumen vertidos de petróleo, disolventes, pesticidas y otros productos tóxicos que contaminan el ambiente, bien sea directamente en el sitio del vertido o bien posteriormente, cuando el material tóxico ha penetrado en el suelo o alcanzado el agua subterránea. La enorme diversidad de los microorganismos en la Tierra permite disponer de grandes recursos genéticos que solucionen la limpieza del medio ambiente; éste es un área de intensa investigación en la actualidad. Los microorganismos y el futuro La biotecnología implica el uso de microorganismos en procesos industriales a gran escala, utilizando por lo general microorganismos modificados genéticamente y capaces de sintetizar productos específicos de elevado valor comercial.
 es un producto bacteriano, derivado de las actividades de las bacterias metanogénicas. Los microorganismos fotótrofos pueden utilizar la luz como fuente de energía para la producción de biomasa, es decir, energía acumulada en forma de organismos vivos. La biomasa microbiana y los materiales de desecho, como la basura doméstica, los excedentes de cosechas y los residuos animales, se pueden convertir en «biocombustibles», como el metano y el etanol, por las actividades degradativas de los microorganismos. Los microorganismos también se pueden usar para ayudar a eliminar la polución originada por las actividades humanas, un proceso que se denomina biorremediación. Se han aislado varios microorganismos de la naturaleza que consumen vertidos de petróleo, disolventes, pesticidas y otros productos tóxicos que contaminan el ambiente, bien sea directamente en el sitio del vertido o bien posteriormente, cuando el material tóxico ha penetrado en el suelo o alcanzado el agua subterránea. La enorme diversidad de los microorganismos en la Tierra permite disponer de grandes recursos genéticos que solucionen la limpieza del medio ambiente; éste es un área de intensa investigación en la actualidad. Los microorganismos y el futuro. La biotecnología implica el uso de microorganismos en procesos industriales a gran escala, utilizando por lo general microorganismos modificados genéticamente y capaces de sintetizar productos específicos de elevado valor comercial.")
21
La biotecnología depende en gran medida de la ingeniería genética, una disciplina que trata de la manipulación artificial de genes y de sus productos. Los genes procedentes de diversos organismos se pueden escindir en fragmentos y modificados utilizando enzimas microbianas como instrumentos moleculares. Mediante técnicas de ingeniería genética, resulta también posible hacer genes completamente artificiales. Una vez que un gen deseado se ha creado o seleccionado puede insertarse en un microorganismo y se puede expresar allí originando el producto génico deseado. Por ejemplo, la insulina humana, una hormona que se encuentra en cantidades anormalmente bajas en sujetos con diabetes, puede ser producida microbiológicamente a partir del gen de la insulina humana expresado en un microorganismo. Tenemos muchas razones para considerar a los microorganismos y sus actividades (Fig. 5). Como dijo uno de los fundadores de la. microbiología, el eminente científico francés Louis Pasteur: «En la naturaleza, el papel de lo infinitamente pequeño es infinitamente grande».
. Como dijo uno de los fundadores de la. microbiología, el eminente científico francés Louis Pasteur: «En la naturaleza, el papel de lo infinitamente pequeño es infinitamente grande».")
22
Los Caminos del Descubrimiento en Microbiología
Como cualquier ciencia, la microbiología moderna debe mucho a su pasado. Aunque se pueden encontrar raíces más profundas, la ciencia de la microbiología no se desarrolló hasta el siglo XIX. Desde entonces se ha extendido hasta originar nuevos campos relacionados entre sí. Ahora reconsideraremos algunos de los caminos que llevaron a importantes descubrimientos. Raíces históricas de la microbiología Aunque durante mucho tiempo se sospechó la existencia de criaturas demasiado pequeñas para ser percibidas a simple vista, su descubrimiento estuvo relacionado con la invención del microscopio. En 1664 Robert Hooke describió los cuerpos fructíferos de mohos (Fig. 6), pero la primera persona que vió microorganismos con detalle fue el holandés Antonie van Leeuwenhoek, aficÍonado a construir microscopios, quien en 1684 utilizó microscopios simples fabricados por él mismo (Fig. 7). Comparados con los actuales, los microscopios de Leeuwenhoek eran bastante primitivos pero mediante una cuidadosa manipulación y un buen enfoque, fue capaz de ver microorganismos tan pequeños como los procariotas. Describió sus observaciones en una serie de cartas dirigidas a la Royal Society de Londres, publicadas en 1684 en inglés. En la (Fig. 7) se muestran dibujos de algunos de sus observaciones que fueron confirmadas por otros investigadores, pero los avances en la comprensión de la naturaleza e importancia de estos minúsculos seres fueron muy lentos.
, pero la primera persona que vió microorganismos con detalle fue el holandés Antonie van Leeuwenhoek, aficÍonado a construir microscopios, quien en 1684 utilizó microscopios simples fabricados por él mismo (Fig. 7). Comparados con los actuales, los microscopios de Leeuwenhoek eran bastante primitivos pero mediante una cuidadosa manipulación y un buen enfoque, fue capaz de ver microorganismos tan pequeños como los procariotas. Describió sus observaciones en una serie de cartas dirigidas a la Royal Society de Londres, publicadas en 1684 en inglés. En la (Fig. 7) se muestran dibujos de algunos de sus observaciones que fueron confirmadas por otros investigadores, pero los avances en la comprensión de la naturaleza e importancia de estos minúsculos seres fueron muy lentos.")
23
Fig 6 La lente objetivo se encajaba al final de un fuelle ajustable, y la iluminación se concentraba en la muestra por una lente sencilla. Moho azul creciendo en la superficie de un trozo de cuero.

24
Fig 7 Dibujos de bacterias y un extendido de sangre humana No fue hasta el siglo XIX cuando los microscopios fueron mejorados generalizándose su uso, lo que permitió poner de manifiesto el alcance y la naturaleza de las formas microbianas de vida.

25
La lente se montaba sobre la placa de latón cerca del extremo del tornillo ajustable de enfoque.
Ferdinand Cohn y la ciencia de la bacteriología La microbiología no se desarrolló como ciencia hasta que las mejoras en microscopía permitieron una mejor observación de las bacterias y se idearon técnicas básicas de laboratorio para el estudio de los microorganismos. El desarrollo de esas técnicas esenciales de laboratorio se vio favorecido por la investigación llevada a cabo durante el siglo XIX en torno a dos temas inquietantes. Uno de estos temas era la cuestión de la generación espontánea. Durante siglos, la idea de que la materia inerte podía originar seres vivos tuvo serios defensores. La segunda incógnita se centraba en la naturaleza de las enfermedades infecciosas. Se sabía que las enfermedades infecciosas se transmitían de un individuo a otro pero los mecanismos de la transmisión eran desconocidos. Aunque las respuestas a estas preguntas se suelen asociar con las figuras de Louis Pasteur y Robert Koch, respectivamente, fue el botánico alemán Ferdinand Cohn (18281898), un contemporáneo de aquéllos, quien fundó la bacteriología (el estudio de las bacterias) y colocó a la incipiente microbiología en un buen punto de partida.
, un contemporáneo de aquéllos, quien fundó la bacteriología (el estudio de las bacterias) y colocó a la incipiente microbiología en un buen punto de partida.")
26
Su interés por la microscopía le llevó primero al estudio de plantas unicelulares, las algas, y más tarde a bacterias fotosintéticas, las cianobacterias. Cohn pensaba que todas las bacterias, incluso aquellas que carecen de pigmentos fotosintéticos, eran miembros del reino vegetal y sus estudios de microscopía de plantas y algas derivaron gradualmente al estudio de diversas bacterias, como la bacteria oxidante del azufre Beggiatoa (Fig 8). Fig 8 Cohn se interesó especialmente por las bacterias resistentes al calor, lo que le llevó a descubrir el género Bacillus y el proceso de formación de esporas. Ahora sabemos que las endosporas bacterianas son estructuras muy resistentes al calor, de hecho las más resistentes de todas las formas microbianas si exceptuamos unas pocas bacterias que crecen mejor a temperaturas notablemente elevadas. Cohn describió el ciclo de vida completo de Bacillus (célula vegetativa ~ endospora ~ célula vegetativa y descubrió que las células vegetati vas, pero no las endosporas, podían morir mediante ebullición.
. Fig 8. Cohn se interesó especialmente por las bacterias resistentes al calor, lo que le llevó a descubrir el género Bacillus y el proceso de formación de esporas. Ahora sabemos que las endosporas bacterianas son estructuras muy resistentes al calor, de hecho las más resistentes de todas las formas microbianas si exceptuamos unas pocas bacterias que crecen mejor a temperaturas notablemente elevadas. Cohn describió el ciclo de vida completo de Bacillus (célula vegetativa ~ endospora ~ célula vegetativa y descubrió que las células vegetati vas, pero no las endosporas, podían morir mediante ebullición.")
27
Estos hallazgos de Cohn ayudaron a explicar por qué algunos científicos anteriores, como John Tyndall, observaron que la ebullición era a menudo una técnica efectiva de esterilización, pero no siempre era así. Cohn continuó trabajando con bacterias hasta su retiro, contribuyendo de muchas maneras al desarrollo de la bacteriología, aportando las bases experimentales para un esquema de clasificación de las bacterias y fundando una importante revista científica. En su época, Cohn fue un entusiasta defensor de las técnicas y las investigaciones desarrolladas por el fundador de la microbiología médica Robert Koch. También se debe a Cohn haber ideado métodos simples y efectivos para evitar la contaminación de medios de cultivo estériles, como el uso del algodón para cerrar los tubos y los matraces. Estos métodos fueron usados posteriormente por Koch y le permitieron grandes avances como el aislamiento y la caracterización de varias bacterias causantes de enfermedades. Pasteur y el fin de la generación espontánea En el siglo XIX tuvo lugar una gran polémica sobre la teoría de la generación espontánea. La idea básica de la generación espontánea puede comprenderse fácilmente. El alimento se pudre si permanece durante cierto tiempo a la intemperie. Cuando este material putrefacto se examina al microscopio se encuentra que está repleto de bacterias. ¿De dónde provienen estas bacterias que no se ven en el alimento fresco? Algunos pensaban que provenían de semillas o gérmenes que llegaban al alimento a través del aire, mientras otros opinaban que se originaban espontáneamente a partir del material inerte. El adversario más ferviente de la generación espontánea fue el químico francés Louis Pasteur ( ), cuyo trabajo sobre este problema fue el más riguroso y convincente.
, cuyo trabajo sobre este problema fue el más riguroso y convincente.")
28
En primer lugar, Pasteur demostró que en el aire había estruturas que se parecían mucho a los microorganismos encontrados en el material putrefacto. Descubrió que el aire normal contiene continuamente una diversidad de células microbianas que son indistinguibles de las que se encuentran en mucha mayor cantidad en los materiales en putrefacción. Por tanto, concluyó que los organismos encontrados en tales materiales se originaban a partir de microorganismos presentes en el aire. Además postuló que dichas células en suspensión se depositan constantemente sobre todos los objetos. Pasteur pensó que si sus conclusiones eran correctas, entonces no debería estropearse un alimento tratado, de tal modo que todos los organismos que lo contaminaran fueran destruidos. Pasteur empleó el calor para eliminar los contaminantes, pues ya se sabía que el calor destruye con efectividad los organismos vivos. De hecho, otros investigadores ya habían mostrado que si una solución de nutrientes se introducía en un matraz de vidrio, se sellaba y se calentaba luego hasta ebullición, nunca se descomponía. Los defensores de la generación espontánea criticaban tales experimentos argumentando que se necesitaba aire fresco para la generación espontánea y que el aire dentro del matraz cerrado se modificaba por el calentamiento, de modo que no era capaz de permitir la generación espontánea.

29
Pasteur superó esta objeción de modo simple y brillante construyendo un matraz con forma
de cuello de cisne, que ahora se designa como un matraz Pasteur (Fig. 9) En tales recipientes, las soluciones nutritivas se podían calentar hasta ebullición; luego, cuando el matraz se enfriaba, el aire podía entrar de nuevo, pero la curvatura del cuello del matraz evitaba que el material particulado, las bacterias y otros microorganismos, alcanzasen el interior del matraz. El material esterilizado en tal recipiente no se descomponía y no aparecían microorganismos mientras el cuello del matraz no contactara con el líquido estéril.
 En tales recipientes, las soluciones nutritivas se podían calentar hasta ebullición; luego, cuando el matraz se enfriaba, el aire podía entrar de nuevo, pero la curvatura del cuello del matraz evitaba que el material particulado, las bacterias y otros microorganismos, alcanzasen el interior del matraz. El material esterilizado en tal recipiente no se descomponía y no aparecían microorganismos mientras el cuello del matraz no contactara con el líquido estéril.")
30
Sin embargo, bastaba con que el matraz se inclinara lo suficiente como para permitir que el líquido estéril contactara con el cuello, para que ocurriera la putrefacción y el líquido se llenara de microorganismos. Este sencillo experimento bastó para aclarar definitivamente la controversia sobre la generación espontánea. Eliminar todas las bacterias o microorganismos de un objeto es un proceso que ahora denominamos esterilización. Los procedimientos que usaron Pasteur, Cohn y otros investigadores fueron finalmente mejorados y aplicados a la investigación microbiológica.

31
El fin de la teoría de la generación espontánea condujo, por tanto, al desarrollo de procedimientos eficaces de esterilización, sin los cuales la microbiología no podría haberse desarrollado como ciencia. Pasteur consiguió muchos otros éxitos en microbiología y medicina. Entre los principales destaca el desarrollo de vacunas para enfermedades como el carbunco, el cólera aviar y la rabia, durante el periodo de Estos avances médicos y veterinarios no sólo tuvieron importancia por sí mismos, sino que permitieron que arraigara el concepto de la teoría microbiana de las enfermedades infecciosas, cuyos principios estaban siendo desarrollados por entonces por otro científico contemporáneo de Pasteur, Robert Koch. Koch y la teoría microbianade las enfermedades infecciosas La demostración de que los microorganismos podían causar enfermedades impulsó el desarrollo inicial de la ciencia de la microbiología. En realidad, ya en el siglo XIX se pensaba que se podía transmitir «algo» de una persona enferma a una sana, y producir en esta última la enfermedad de la primera. Muchas enfermedades parecían diseminarse por la población y se llamaban contagiosas mientras que el agente desconocido que causaba la diseminación fue llamado contagio. Tras el descubrimiento de los microorganismos, se sospechaba que éstos pudieran ser responsables de enfermedades, pero faltaban las pruebas definitivas.

32
Los descubrimientos de Ignaz Semmelweis y Joseph Lister suministraron algunas pruebas indirectas sobre la importancia de los microorganismos como causa de enfermedades en el hombre, pero la teoría microbiana de las enfermedades infecciosas no fue claramente concebida y experimentalmente probada hasta los trabajos del médico Robert Koch ( ). En su trabajo inicial Koch estudió el carbunco, una enfermedad del ganado que en ocasiones también afecta al hombre. Esta enfermedad está causada por una bacteria formadora de endosporas, Bacillus anthracis, y la sangre de un animal con carbunco está llena de células de esta bacteria. Mediante cuidadosos estudios de microscopía, Koch puso de manifiesto que la bacteria estaba siempre presente en la sangre de los animales enfermos. Sin embargo, la mera asociación de la bacteria con la enfermedad no demostraba que la bacteria fuera la causa de la enfermedad; por el contrario, podría ser un efecto de la enfermedad. Por eso, Koch demostró que era posible tomar una pequeña cantidad de sangre de un ratón enfermo, inyectada en un segundo ratón y provocar en éste la enfermedad y la muerte. Tomando sangre de este segundo animal e inyectándola en otro, obtenía de nuevo los síntomas característicos de la enfermedad. Repitiendo experimentos de este tipo, Koch demostró por microscopía que la sangre del animal enfermo contenía gran cantidad de la bacteria formadora de endosporas. Koch llevó este experimento aun más lejos.

33
También demostró que la bacteria podía ser cultivada en caldos nutritivos fuera del animal y que, incluso después de muchas resiembras o transferencias de cultivo, la bacteria podía causar la enfermedad aún cuando se reinoculaba a un animal. Es decir, la bacteria procedente de un animal enfermo y la mantenida en cultivo inducían los mismos síntomas de enfermedad tras su inoculación. Basándose en éste y en otros experimentos, Koch formuló los siguientes criterios, conocidos en la actualidad como postulados de Koch, para demostrar que un tipo concreto de microorganismo es el agente etiológico de una enfermedad específica. Postulados de Koch 1. El organismo debe estar siempre presente en los animales que sufran la enfermedad y no en individuos sanos. 2. El organismo debe cultivarse en cultivo axénico o puro fuera del cuerpo del animal. 3. Cuando dicho cultivo se inocula a un animal susceptible, debe iniciar en él los síntomas característicos de la enfermedad. 4. El organismo debe aislarse nuevamente de estos animales experimentales y cultivarse nuevamente en el laboratorio, tras lo cual debe mostrar las mismas propiedades que el microorganismo original. Los postulados de Koch se resumen en la (Fig. 10).
.")
34
Fig 10

35
Koch y los cultivos puros
Para relacionar un microorganismo determinado con un proceso específico, como el caso de una enfermedad, el microorganismo debe ser primero aislado en un cultivo; es decir, el cultivo debe de ser axénico o puro. Este concepto fue recogido por Koch en la formulación de sus famosos postulados y desarrolló varios métodos ingeniosos para obtener bacterias en cultivo puro. Koch empezó estos estudios de forma rudimentaria, usando nutrientes sólidos como la superficie de una rebanada de patata para cultivar bacterias, pero pronto diseñó métodos más fiables, muchos de los cuales aún se usan en la actualidad. Koch observó que cuando se exponía al aire la superficie de un nutriente sólido se desarrollaban colonias bacterianas que tenían formas y colores característicos. Dedujo que cada colonia se originaba a partir de una sola célula bacteriana que había caído sobre la superficie, había encontrado los nutrientes apropiados y se había multiplicado; es decir, cada colonia representaba un cultivo axénico o puro. Koch se dio cuenta de que este descubrimiento suponía un sencillo procedimiento para obtener cultivos puros. Como muchos organismos no crecen en rebanadas de patata, Koch ideó caldos nutritivos más uniformes y reproducibles solidificados con gelatina y más tarde con agar. Actualmente el agar es el agente solidificante más usado en los laboratorios de microbiología para obtener y mantener cultivos puros de muchos microorganismos, especialmente de bacterias.

36
Cultivo puro de aire con colonias de hongos y bacterias.
Koch y la Tuberculosis Aunque en aquel tiempo se sospechaba que la tuberculosis era una enfermedad contagiosa, el organismo responsable de la enfermedad no se había visto, ni en los tejidos de enfermo ni en cultivo. Desde el principio de su estudio sobre la tuberculosis, el objetivo de Koch fue detectar el agente causante de la enfermedad y para ello empleó todos los métodos que había desarrollado previamente: microscopía, tinción de tejidos, aislamiento en cultivo puro e inoculación en animales. Como sabemos ahora, el «bacilo de la tuberculosis», Mycobacterium tuberculosis, es muy difícil de teñir debido a que posee grandes cantidades de lípidos en su superficie. Pero Koch diseñó un procedimiento para teñir M. tuberculosis en muestras de tejidos usando azul de metileno alcalino y un segundo colorante (marrón Bismark) que teñía sólo el tejido.
 que teñía sólo el tejido.")
37
El método de Koch fue el precursor de la tinción de Ziehl-Nielsen usada hoy para teñir bacterias ácido-alcohol resistentes como M. tuberculosis. Usando su nuevo método de tinción, Koch observó las células bacilares de M. tuberculosis teñidas de azul en tejidos tuberculosos, quedando estos últimos teñidos de marrón claro. Fig 11 Sin embargo, por su trabajo anterior sobre el carbunco, Koch era consciente de que identificar un microorganismo asociado a la tuberculosis no bastaba; debía cultivar el microorganismo para demostrar que era la causa específica de la tuberculosis. La obtención de cultivos de M. tuberculosis no fue tarea fácil, pero finalmente Koch tuvo éxito al obtener colonias de este organismo sobre suero de sangre coagulada. Posteriormente usó agar, que acababa de ser introducido como agente solidificante. Los cobayas pueden ser infectados fácilmente con M. tuberculosis y posteriormente mueren de tuberculosis sistémica. Koch demostró que los cobayas enfermos contenían masas celulares de M. tuberculosis en sus tejidos y que los cultivos puros obtenidos de dichos animales transmitían la enfermedad a animales sanos.

38
El nitrógeno se halla tan sólo por detrás del carbono, el hidrógeno y el oxígeno en su contribución a la masa de los seres vivos. La mayor parte de este nitrógeno forma parte de aminoácidos y nucleótidos. Aspectos generales del metabolismo del nitrógeno Las rutas biosintéticas de los aminoácidos y de los nucleótidos comparten la necesidad de nitrógeno. Por lo general, los compuestos de nitrógeno solubles y utilizables biológicamente escasean en el entorno natural. Por este motivo, la mayor parte de organismos mantienen una estricta economía en el uso del amoniaco, aminoácidos y nucleótidos. En efecto, los aminoácidos libres, purinas y pirimidinas, formados durante el recambio metabólico, normalmente son recuperados y reutilizados. En primer lugar examinaremos las vías por las cuales el nitrógeno del medio se introduce en los sistemas biológicos. El ciclo del nitrógeno mantiene una reserva de nitrógeno disponible biológicamente La fuente más abundante de nitrógeno es el aire, que en sus cuatro quintas partes es nitrógeno molecular (N2). A pesar de ello, tan sólo unas pocas especies pueden convertir el nitrógeno atmosférico en formas utilizables por los organismos vivos. Los procesos metabólicos de las diferentes especies actúan de forma interdependiente para recuperar y reutilizar el nitrógeno disponible biológicamente en un amplio ciclo del nitrógeno .
. A pesar de ello, tan sólo unas pocas especies pueden convertir el nitrógeno atmosférico en formas utilizables por los organismos vivos. Los procesos metabólicos de las diferentes especies actúan de forma interdependiente para recuperar y reutilizar el nitrógeno disponible biológicamente en un amplio ciclo del nitrógeno .")
39
Ciclo del Nitrógeno Fig 12

40
El primer paso en el ciclo del nitrógeno consiste en la fijación (reducción) del nitrógeno atmosférico por bacterias fijadoras de nitrógeno para proporcionar amoníaco (NH3 o NH4+). Aunque el amoníaco puede ser utilizado por la mayor parte de organismos vivos, las bacterias del suelo, que obtienen su energía de la oxidación del amoníaco a nitrito (N02-) y finalmente a nitrato (N03 -), son tan abundantes y activas que prácticamente todo el amoníaco que alcanza el suelo es oxidado a nitrato. Este proceso se conoce como nitrificación. Las plantas y muchas bacterias pueden incorporar y reducir fácilmente nitrato y nitrito por acción de las nitrato y nitrito reductasas. El amoníaco así formado es incorporado a los aminoácidos por las plantas. Los animales a su vez usan las plantas como fuente de aminoácidos, tanto esenciales como no esenciales, para fabricar sus proteínas. Cuando los organismos mueren, la degradación microbiana de sus proteínas devuelve el amoníaco al suelo, en donde bacterias nitrificantes lo convierten de nuevo en nitrito y nitrato. Las bacterias que convierten el nitrato en N2 bajo condiciones anaeróbicas mantienen un equilibrio entre el nitrógeno fijado y el nitrógeno atmosférico. En este proceso, denominado desnitrificación (Fig. 12), estas bacterias del suelo utilizan N03- en lugar de O2 como aceptar final de electrones en una serie de reacciones que (al igual que la fosforilación oxidativa) generan un gradiente transmembrana de protones, que se aprovecha para sintetizar ATP. A continuación examinaremos el proceso de fijación del nitrógeno, que es el primer paso del ciclo del nitrógeno.
 y finalmente a nitrato (N03 -), son tan abundantes y activas que prácticamente todo el amoníaco que alcanza el suelo es oxidado a nitrato. Este proceso se conoce como nitrificación. Las plantas y muchas bacterias pueden incorporar y reducir fácilmente nitrato y nitrito por acción de las nitrato y nitrito reductasas. El amoníaco así formado es incorporado a los aminoácidos por las plantas. Los animales a su vez usan las plantas como fuente de aminoácidos, tanto esenciales como no esenciales, para fabricar sus proteínas. Cuando los organismos mueren, la degradación microbiana de sus proteínas devuelve el amoníaco al suelo, en donde bacterias nitrificantes lo convierten de nuevo en nitrito y nitrato. Las bacterias que convierten el nitrato en N2 bajo condiciones anaeróbicas mantienen un equilibrio entre el nitrógeno fijado y el nitrógeno atmosférico. En este proceso, denominado desnitrificación (Fig. 12), estas bacterias del suelo utilizan N03- en lugar de O2 como aceptar final de electrones en una serie de reacciones que (al igual que la fosforilación oxidativa) generan un gradiente transmembrana de protones, que se aprovecha para sintetizar ATP. A continuación examinaremos el proceso de fijación del nitrógeno, que es el primer paso del ciclo del nitrógeno.")
41
El nitrógeno es fijado por enzimas del complejo de la nitrogenasa (Fig 13)
Tan sólo algunos procariotas pueden fijar el nitrógeno atmosférico. Entre ellos se encuentran las cianobacterias del suelo y de las aguas dulces y saladas, otros tipos de bacterias del suelo, tales como algunas especies de Azotobacter, y las bacterias fijadoras de nitrógeno que viven como simbiontes en los nódulos de las raíces de las plantas leguminosas. El primer producto importante de la fijación del nitrógeno es el amoníaco, que puede ser utilizado por otros organismos, ya sea directamente o tras su conversión en otros compuestos solubles, tales como nitritos, nitratos o aminoácidos. La reducción del nitrógeno a amoníaco es una reacción exergónica: N2 + 3 H NH3 ∆G = kJ / mol Sin embargo, el triple enlace N - N es muy estable, con una energía de enlace de 942 kJ/mol. La fijación del nitrógeno tiene, por tanto, una energía de activación extremadamente alta,

42
con lo que el nitrógeno atmosférico es prácticamente inerte en condiciones normales.
El amoníaco se produce industrialmente por el procedimiento de Haber (llamado así en honor de Fritz Haber, su descubridor), que requiere temperaturas entre 400 y 500°C y presiones de N2 y H2 de decenas de miles de kilopascales (varios centenares de atmósferas) para conseguir la energía de activación necesaria. La fijación biológica del nitrógeno ha de tener lugar a 0,8 atm de nitrógeno y a temperaturas biológicas. La elevada barrera de activación se supera, al menos en parte, por la unión y la hidrólisis del ATP. La reacción completa puede escribirse así: N H e ATP 2 NH ADP Pi + H2 La fijación biológica de nitrógeno se realiza por un complejo de proteínas muy conservado, denominado complejo de la nitrogenasa (Fig. 13). Los componentes clave de este complejo son la dinitrogenasa reductasa y la dinitrogenasa (Fig. 13). La dinitrogenasa reductasa (PM ) es un dímero formado por dos subunidades idénticas. Contiene un único centro redox del tipo 4Fe-4S (Fig. 14) y puede ser oxidado y reducido por un electrón. Tiene también dos centros de unión de ATP / ADP. La dinitrogensa (PM ) es un tetrámero formado por dos copias de dos subunidades distintas. La dinitrogenasa contiene hierro y molibdeno; sus centros redox tienen un total de 2 Mo, 32 Fe y 30 S por tetrámero.
, que requiere temperaturas entre 400 y 500°C y presiones de N2 y H2 de decenas de miles de kilopascales (varios centenares de atmósferas) para conseguir la energía de activación necesaria. La fijación biológica del nitrógeno ha de tener lugar a 0,8 atm de nitrógeno y a temperaturas biológicas. La elevada barrera de activación se supera, al menos en parte, por la unión y la hidrólisis del ATP. La reacción completa puede escribirse así: N H+ + 8 e ATP 2 NH ADP + 16 Pi + H2. La fijación biológica de nitrógeno se realiza por un complejo de proteínas muy conservado, denominado complejo de la nitrogenasa (Fig. 13). Los componentes clave de este complejo son la dinitrogenasa reductasa y la dinitrogenasa (Fig. 13). La dinitrogenasa reductasa (PM ) es un dímero formado por dos subunidades idénticas. Contiene un único centro redox del tipo 4Fe-4S (Fig. 14) y puede ser oxidado y reducido por un electrón. Tiene también dos centros de unión de ATP / ADP. La dinitrogensa (PM ) es un tetrámero formado por dos copias de dos subunidades distintas. La dinitrogenasa contiene hierro y molibdeno; sus centros redox tienen un total de 2 Mo, 32 Fe y 30 S por tetrámero.")
43
Fig 14 Aproximadamente la mitad del Fe y del S se encuentra en dos centros 4Fe-4S conectados, denominados agregados P. El resto forma parte de un cofactor de hierro-molibdeno de nuevo tipo. Se ha descubierto una forma de nitrogenasa que contiene vanadio en lugar de molibdeno; algunas especies de bacterias pueden producir ambos tipos de nitrogenasa. La enzima de vanadio puede ser el sistema primario de fijación de nitrógeno en algunas condiciones ambientales, pero todavía no está bien caracterizado. La fijación del nitrógeno se lleva a cabo por una forma altamente reducida de la dinitrogenasa y precisa ocho electrones: seis para la reducción del N2 y dos para formar una molécula de H2 como parte obligada del mecanismo de reacción. La dinitrogenasa se reduce por transferencia de electrones procedentes de la dinitrogenasa reductasa (Fig. 13 ). La dinitrogenasa tiene dos centros de unión para la reductasa y los ocho electrones necesarios se transfieren a la dinitrogenasa de uno en uno.
. La dinitrogenasa tiene dos centros de unión para la reductasa y los ocho electrones necesarios se transfieren a la dinitrogenasa de uno en uno.")
44
La forma reducida de la reductasa se une a la dinitrogenasa y la forma oxidada de la reductasa se disocia en un ciclo repetitivo. Este ciclo precisa de la hidrólisis de ATP por la reductasa. La fuente inmediata de electrones para reducir la dinitrogenasa reductasa es variable, pudiéndose utilizar ferredoxina reducida , flavodoxina reducida y quizás otras fuentes. En al menos un caso, la fuente principal de electrones para reducir la ferredoxina es el piruvato (Fig.15). El papel del ATP en este proceso es interesante, pues resulta ser catalítico en lugar de termodinámico. Recuérdese que el ATP puede proporcionar no sólo energía química, a través de la hidrólisis de uno o más de sus enlaces fosfoanhídrido, sino también energía de enlace mediante interacciones no covalentes que reducen la energía de activación. En la reacción llevada a cabo por la dinitrogenasa reductasa tanto la unión del ATP como la
. El papel del ATP en este proceso es interesante, pues resulta ser catalítico en lugar de termodinámico. Recuérdese que el ATP puede proporcionar no sólo energía química, a través de la hidrólisis de uno o más de sus enlaces fosfoanhídrido, sino también energía de enlace mediante interacciones no covalentes que reducen la energía de activación. En la reacción llevada a cabo por la dinitrogenasa reductasa tanto la unión del ATP como la.")
45
hidrólisis del ATP provocan cambios conformacionales que sin duda ayudan a superar la elevada energía de activación de la fijación del nitrógeno. La unión del ATP a la reductasa altera el potencial de reducción (E'O) de esta proteína de -250 a -400 mV, un incremento de su potencial reductor necesario para transferir electrones a la dinitrogenasa. Dos moléculas de ATP son hidrolizadas a continuación, justo antes de la transferencia efectiva de cada electrón de la dinitrogenasa reductasa a la dinitrogenasa. Otra característica importante del complejo de la nitrogenasa es su extrema labilidad en presencia de oxígeno. La reductasa se inactiva en contacto con el aire, con una vida media de 30 seg; la dinitrogenasa tiene una vida media de 10 min en contacto con el aire. Las bacterias libres que fijan nitrógeno solucionan este problema de diferentes maneras. Algunas viven solamente en condiciones anaeróbicas o reprimen la síntesis de la nitrogenasa en presencia de oxígeno. Algunas bacterias aeróbicas, tales como Azotobacter vinelandii, desacoplan parcialmente el transporte electrónico de la síntesis de ATP, de manera que el oxígeno es quemado tan pronto como entra en la célula. De hecho, cuando fijan nitrógeno los cultivos de estas bacterias se calientan como resultado de sus esfuerzos para eliminar el oxígeno. Algunas cianobacterias filamentosas fijadoras de nitrógeno utilizan otro procedimiento: una de cada nueve células se diferencia formando un heterocisto, especializado en la fijación del nitrógeno, con paredes suficientemente gruesas para impedir la entrada de oxígeno.

46
La relación simbiótica entre las plantas leguminosas y las bacterias fijadoras de nitrógeno en los nódulos de sus raíces (Fig. 16) (nódulos fijadores de N 2 en raíces de trébol), resuelve a la vez las necesidades energéticas de la reacción y la labilidad de los enzimas frente al oxígeno. La energía necesaria para la fijación del nitrógeno fue probablemente la fuerza impulsora que a lo largo de la evolución condujo a esta asociación de plantas y bacterias. Las bacterias en los nódulos de las raíces tienen acceso a una gran reserva de energía en forma de glúcidos y de intermedios del ciclo del ácido cítrico suministrados por la planta. Ello permite a las bacterias fijar centenares de veces más nitrógeno que sus parientes próximos que viven en los suelos. Para solventar el problema de la toxicidad del oxígeno, las bacterias de los nódulos de las raíces están impregnadas de una disolución de una hemoproteína fijadora de oxígeno, denominada leghemoglobina. Esta proteína es producida por la planta (aunque el grupo hemo puede que sea proporcionado por la bacteria).
.")
47
La leghemoglobina fija todo el oxígeno, de manera que éste no puede interferir con la fijación del nitrógeno y, a la vez, suministra eficazmente oxígeno al sistema de transporte de electrones de la bacteria. La eficacia de la simbiosis entre los vegetales y las bacterias es evidente por el enriquecimiento de nitrógeno en el suelo propiciado por las plantas leguminosas. Este enriquecimiento es la base del procedimiento rotatorio de cosechas utilizado por muchos agricultores, en el que el cultivo de plantas no leguminosas (como el maíz), que aprovechan el nitrógeno fijado presente en el suelo, se alterna cada pocos años con cultivos de leguminosas tales como la alfalfa, los guisantes o el trébol. La fijación del nitrógeno es objeto de intensos estudios debido a su enorme importancia práctica. La producción industrial de amoniaco para su utilización en fertilizantes necesita mucha energía; ello ha estimulado los esfuerzos para desarrollar organismos recombinantes o transgénicos capaces de fijar el nitrógeno. Las técnicas del DNA recombinante están siendo utilizadas para transferir el DNA que codifica los enzimas de la fijación del nitrógeno a bacterias no fijadoras de nitrógeno y a plantas. El éxito de estos esfuerzos depende de la solución del problema de la toxicidad del oxígeno en cualquier célula que produzca nitrogenasa.
, que aprovechan el nitrógeno fijado presente en el suelo, se alterna cada pocos años con cultivos de leguminosas tales como la alfalfa, los guisantes o el trébol. La fijación del nitrógeno es objeto de intensos estudios debido a su enorme importancia práctica. La producción industrial de amoniaco para su utilización en fertilizantes necesita mucha energía; ello ha estimulado los esfuerzos para desarrollar organismos recombinantes o transgénicos capaces de fijar el nitrógeno. Las técnicas del DNA recombinante están siendo utilizadas para transferir el DNA que codifica los enzimas de la fijación del nitrógeno a bacterias no fijadoras de nitrógeno y a plantas. El éxito de estos esfuerzos depende de la solución del problema de la toxicidad del oxígeno en cualquier célula que produzca nitrogenasa.")
48
CICLO DEL CARBONO Y DEL OXíGENO
El ciclo global del carbono depende de la actividad tanto de los microorganismos como de los macroorganismos y está estrechamente ligado al ciclo del oxígeno. Debido al aumento de las emisiones de CO2 a causa de la actividad humana, el ciclo del carbono es objeto de un renovado interés. Los científicos tratan de entender mejor la magnitud de los reservorios de carbono, de los principales sumideros de CO2 y las velocidades de reciclado dentro y entre compartimentos, para impedir catástrofes planetarias, como el calentamiento global. Ciclo del carbono Considerado globalmente, el carbono circula por todos los reservorios de la Tierra que lo contienen: la atmósfera, el medio terrestre, los mares y otros ambientes acuáticos, sedimentos y rocas, y en la biomasa (Fig 17 ). Tal como veremos, el ciclo del carbono y el del oxígeno están estrechamente relacionados, ya que la fijación de C02 por fotótrofos oxigénicos libera O2 y mucha de la materia orgánica es oxidada a CO2 a través de la respiración aeróbica (Fig 17).
. Tal como veremos, el ciclo del carbono y el del oxígeno están estrechamente relacionados, ya que la fijación de C02 por fotótrofos oxigénicos libera O2 y mucha de la materia orgánica es oxidada a CO2 a través de la respiración aeróbica (Fig 17).")
49
Reservorios de carbono
El mayor reservorio de carbono lo constituyen las rocas y sedimentos de la corteza terrestre, pero el tiempo de reciclado es tan grande que las salidas de dicho compartimento son relativamente insignificantes consideradas a escala humana. Tomando como referencia los organismos vivos, una gran cantidad de carbono orgánico se encuentra en las plantas terrestres.

50
Esto representa el carbono de los bosques y praderas y constituye el principal lugar de fijación fotosintética de CO2. Sin embargo, la materia orgánica muerta, llamada humus, contiene mayor cantidad de carbono que los seres vivos. El humus es una mezcla compleja de materia orgánica. Procede parcialmente de los constituyentes de los microorganismos del suelo que han resistido la descomposición y, en parte, también del material resistente de las plantas. Algunas sustancias húmicas son muy estables, con un tiempo de reciclado de unos cuarenta años, aunque otras se descomponen mucho más rápidamente. Por ejemplo, algunas sustancias húmicas pueden ser aceptores de electrones en la respiración anaeróbica. El medio más rápido de transferencia global del carbono es mediante el CO2 de la atmósfera. El CO2 es retirado de la atmósfera principalmente por la fotosíntesis de las plantas terrestres y vuelve a ella a través de la respiración de los animales y de los microorganismos quimioorganotróficos. El análisis de los diversos procesos indica que el aporte de CO2 más importante que llega a la atmósfera proviene de la descomposición de la materia orgánica muerta, incluyendo el humus, llevada a cabo por microorganismos. Recientemente, sin embargo, la actividad humana ha aumentado alarmantemente la reserva de CO2 atmosférico. Por ejemplo, en los últimos 40 años, los niveles de CO2 han aumentado un 12%.

51
Importancia de la fotosíntesis en el ciclo del carbono
La única vía importante de producción de carbono orgánico nuevo en nuestro planeta procede de la fotosíntesis y de la quimiosíntesis (fijación de CO2 por quimiolitótrofos); probablemente, la mayor parte del carbono procede de la fotosíntesis. Por tanto, los organismos fototróficos se encuentran en la base del ciclo del carbono (Fig 17). Los organismos fototróficos se encuentran en la naturaleza casi exclusivamente en hábitat donde hay luz disponible. De ahí que las grandes profundidades marinas y otros hábitat que están permanentemente en la oscuridad carezcan de fotótrofos. Los organismos fototróficos oxigénicos pueden dividirse en dos grandes grupos: plantas superiores y microorganismos. Las plantas superiores son los organismos fototróficos que predominan en los ambientes terrestres, mientras que los microorganismos fototróficos son los fotosintetizadores más abundantes en los ambientes acuáticos. La (Fig 18) muestra el ciclo redox del carbono. Empezamos por la fotosíntesis. La ecuación global de la fotosíntesis oxigénica es: CO2 + H (CH2O) + O2 (luz) en la cual (CH2O) representa materia orgánica en el estado de oxidación de material celular, como los polisacáridos (la principal forma en que la materia orgánica fotosintetizada se almacena en la célula). Los organismos fototróficos también llevan a cabo la respiración, tanto en presencia de la luz
; probablemente, la mayor parte del carbono procede de la fotosíntesis. Por tanto, los organismos fototróficos se encuentran en la base del ciclo del carbono (Fig 17). Los organismos fototróficos se encuentran en la naturaleza casi exclusivamente en hábitat donde hay luz disponible. De ahí que las grandes profundidades marinas y otros hábitat que están permanentemente en la oscuridad carezcan de fotótrofos. Los organismos fototróficos oxigénicos pueden dividirse en dos grandes grupos: plantas superiores y microorganismos. Las plantas superiores son los organismos fototróficos que predominan en los ambientes terrestres, mientras que los microorganismos fototróficos son los fotosintetizadores más abundantes en los ambientes acuáticos. La (Fig 18) muestra el ciclo redox del carbono. Empezamos por la fotosíntesis. La ecuación global de la fotosíntesis oxigénica es: CO2 + H20 (CH2O) + O2 (luz) en la cual (CH2O) representa materia orgánica en el estado de oxidación de material celular, como los polisacáridos (la principal forma en que la materia orgánica fotosintetizada se almacena en la célula). Los organismos fototróficos también llevan a cabo la respiración, tanto en presencia de la luz.")
52
como en la oscuridad. La ecuación general de la respiración es la inversa de la anterior:
(CH2O) + O CO H2O en la que (CH2O) representa también polisacáridos de reserva. Para que un organismo crezca (es decir, aumente en número de células o de masa) fototróficamente, la tasa de fotosíntesis debe ser superior a la tasa de respiración. Si eso ocurre, parte del carbono fijado a partir del CO2 en los polisacáridos puede dedicarse a la biosíntesis. El ciclo completo del carbono se construye a partir de un equilibrio neto positivo de la tasa de fotosíntesis sobre la tasa de respiración. Descomposición El carbono fijado fotosintéticamente es degradado finalmente por varios organismos. En la degradación, se observan dos estados principales de oxidación del carbono: metano (CH4) y dióxido de carbono (C02) (Fig 18).
 + O2 CO2 + H2O. en la que (CH2O) representa también polisacáridos de reserva. Para que un organismo crezca (es decir, aumente en número de células o de masa) fototróficamente, la tasa de fotosíntesis debe ser superior a la tasa de respiración. Si eso ocurre, parte del carbono fijado a partir del CO2 en los polisacáridos puede dedicarse a la biosíntesis. El ciclo completo del carbono se construye a partir de un equilibrio neto positivo de la tasa de fotosíntesis sobre la tasa de respiración. Descomposición. El carbono fijado fotosintéticamente es degradado finalmente por varios organismos. En la degradación, se observan dos estados principales de oxidación del carbono: metano (CH4) y dióxido de carbono (C02) (Fig 18).")
53
Fig 18

54
El CH4 se produce por la actividad de los metanógenos y el CO2 por diversos quimioorganótrofos mediante fermentación, por respiración anaeróbica o por respiración aeróbica. En hábitat anóxicos, el CH4 resulta de la reducción de CO2 con H2 y de algunos compuestos como el acetato. Sin embargo, prácticamente, cualquier compuesto orgánico puede ser convertido a CH4 a partir de la actividad combinada de bacterias fotosintéticas y metanógenos. El H2 generado por la degradación fermentativa de compuestos orgánicos lo consumen los metanógenos. El metano producido en hábitat anóxicos es muy insoluble, por lo que es fácilmente transportado a ambientes óxicos, donde es oxidado a CO2 por los metanotrofos (Fig 18). En consecuencia, todo el carbono orgánico revierte posteriormente a CO2, a partir del cual el metabolismo autotrófico empieza una vez más el ciclo del carbono. El balance entre la parte oxidativa y la reductora del ciclo del carbono es crítica; los productos del metabolismo de algunos organismos son sustratos para otros. El ciclo necesita mantenerse en equilibrio si es que va a continuar en la forma en que lo ha hecho durante miles de millones de años. Cualquier cambio significativo en las concentraciones de las formas gaseosas del carbono puede tener consecuencias globales (como hemos experimentado debido al aumento de CO2 atmosférico por la deforestación y la quema de combustibles fósiles). En términos de descomposición, el CO2 que libera la actividad microbiana excede en mucho al liberado por los eucariotas.
. En consecuencia, todo el carbono orgánico revierte posteriormente a CO2, a partir del cual el metabolismo autotrófico empieza una vez más el ciclo del carbono. El balance entre la parte oxidativa y la reductora del ciclo del carbono es crítica; los productos del metabolismo de algunos organismos son sustratos para otros. El ciclo necesita mantenerse en equilibrio si es que va a continuar en la forma en que lo ha hecho durante miles de millones de años. Cualquier cambio significativo en las concentraciones de las formas gaseosas del carbono puede tener consecuencias globales (como hemos experimentado debido al aumento de CO2 atmosférico por la deforestación y la quema de combustibles fósiles). En términos de descomposición, el CO2 que libera la actividad microbiana excede en mucho al liberado por los eucariotas.")
55
Bibliografía Madigan Michael T, Martinko John M, Parker Jack. “ Brock Biología de los Microorganismos”. Pearson – Prentice Hall. 2004 Nelson David L, Cox Michael M. “ Lehninger Principios de Bioquímica”. Ediciones Omega. 2001

Presentaciones similares